Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Protein moonlighting wikipedia , lookup
Western blot wikipedia , lookup
Molecular evolution wikipedia , lookup
Deoxyribozyme wikipedia , lookup
Protein adsorption wikipedia , lookup
Two-hybrid screening wikipedia , lookup
Protein–protein interaction wikipedia , lookup
Nucleic acid analogue wikipedia , lookup
Photosynthetic reaction centre wikipedia , lookup
List of types of proteins wikipedia , lookup
Biochemistry wikipedia , lookup
Nuclear magnetic resonance spectroscopy of proteins wikipedia , lookup
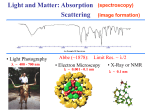
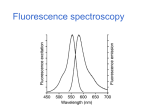
Lecture 32: Spectroscopy (continued) Review o Two basic rules of spectroscopy o Transition probability o Lifetime broadening Today UV visible spectroscopy o Fundamentals o Beer-Lambert’s law o Examples o Proteins Secondary structure o DNA Fluorescence o Fundamentals o Examples o Quantitative aspects o Fluorescence lifetime and quantum yield o Excitation transfer o Quenching Rules of spectroscopy In spectroscopy we deal with interaction of radiation with molecules. Given that quantum mechanics gives us the energy levels and the associated wavefunctions, we can summarize two fundamental aspects of the spectroscopy. Whether a given transition of the molecular systems between the energy levels 1 to 2 will take place or not depends on whether the following integral is nonzero or not. 2* i 1d In this formula, is the dipole moment, i=x,y,z. If this integral is nonzero we call the transition from energy level 1 to 2 is “dipole” allowed. In UV-visible and IR spectroscopies, the selection rules are governed by the dipole interaction. However, the selection rule is of general validity. For example, a given NMR transition is allowed or not depends on the angular momentum operator Ix. Similarly there are quadrupole allowed transitions. Second important rule concerns the linewidth of a spectral line. The uncertainty principle can be restated as follows: E.t h / 2 or t 1 / 4 Longer the lifetime of the excited state, sharper/narrower is the linewidth. UV-visible spectroscopy Here we are concerned mainly with the electronic transitions, as shown below. Within each electronic level, there are the vibrational energy levels approximated as harmonic oscillator. During the excitation of molecule, it may end up in one of the vibrationally excited state. This is not very pronounced for the spectra taken from solutions due to broadening caused by collisions with the solvent molecule. Note in the diminished linewidth broadening in vapor phase UV-visible spectroscopy of mixtures: Beer-Lambert law Example of the isobestic points Since isobestic points occur if and only if there are only two absorbing species, it can be a very useful diagnostic technique, as shown below. The reason for this unique property of the isobestic point is that the chances of having the same extinction coefficient for three compounds are extremely low. Typical Absorption spectra of proteins and DNA In proteins, two maxima (280 and 200nms) have been ascribed to pi-pi* transitions in aromatic rings and the amide bonds respectively. Unlike proteins, nucleic acids generally show one maximum at 260nm, whose intensity decreases in organized helical structure. This is the so-called hypo-chromic effect. The ratio of A260/A280 has been used to determine nucleic acid to protein concentration ratio. Additionally, the UV-visible spectroscopy is sensitive to the secondary structure of protein: Fluorescence We will use the kinetics of the first order process to describe terminology of fluorescence. Kinetics of Fluorescence However, the kd contains many contributions. Contributions to Kd and the quantum yield Fluorescence quenching Many molecules that having energy level separation comparable to the excited state energy of the photo-excited molecules. They can quench the fluorescence. Molecular oxygen is one prime example of fluorescence quencher. In this case the kinetic scheme is described as: Excitation transfer Using We can easily show that Thus one tries to detect the fluorescence of the acceptor. The key to excitation transfer process is the distance between the donor and the acceptor. Forrester pointed out that it is due to dipolar interactions and has r6 dependence. Molecular Ruler The basic idea that the excitation transfer depends on the distance of separation between the donor and the acceptor species can be used to design molecular ruler. Beautiful studies of Stryer et al demonstrated the potential of this approach in the studies of nucleic acids. These researchers labeled two termini of the oligo-proline strands and observed the excitation transfer efficiency as the fluorescent acceptor to donor distance was systematically varied. Shown below are their results. Note the excellent fit to R6 model. The key criteria for this method to work is that absorption spectrum of the acceptor must overlap the emission spectrum of the donor. It is only then the resonance energy transfer can take place. Single molecule fluorescence One of the difficulties in performing the fluorescent energy transfers, and hence the studies of inter-molecular interaction, is that one needs fluorescent labeled materials. Covalent attachment of these marker molecules is time consuming and requires special synthetic techniques. However the beauty of the fluorescent-labeled studies is that concentration of the material needed is small. As matter of fact, single molecule fluorescence can be observed owing to the extraordinary sensitivity of the photon counting instruments. An example of application of this technique to study ATP synthesase is shown below. Single molecule fluorescence In their experiment Noji et al attached the “motor “ portion of the protein to the glass slide by chemical bonding. And a fluorescently labeled actin molecule replaced the transmembrane part of the protein. When the slide was placed in solution containing ATP they observed rotation of actin filament by the monitoring emission of light of the probe under a fluorescent microscope. It is hindered rotation occurring in jumps of 120 degrees. Modern DNA and Protein chips also make use of this extraordinary sensitivity. A complementary strands on DNA are synthesized on the chip surface as shown below Fluorescently labeled complementary strand is then introduced. Binding event only occurs when the sequence introduced is complementary to the structure present on the chip. In this way, today genes are detected.