Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
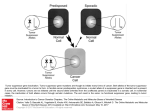
t(11;14)(q13;q32) in multiple myeloma Atlas of Genetics and Cytogenetics in Oncology and Haematology Huret JL, Laï JL OPEN ACCESS JOURNAL AT INIST-CNRS Gene Section Review CHD5 (chromodomain helicase DNA binding protein 5) Venkatadri Kolla, Mayumi Higashi, Tiangang Zhuang, Garrett M Brodeur Children's Hospital of Philadelphia, Oncology Research, CTRB Rm 3018, 3501 Civic Center Blvd, Philadelphia, PA 19104, USA (VK, MH, TZ, GMB) Published in Atlas Database: January 2012 Online updated version : http://AtlasGeneticsOncology.org/Genes/CHD5ID44521ch1p36.html DOI: 10.4267/2042/47336 This article is an update of : Brodeur GM, White PS. CHD5 (Chromodomain-helicase-DNA binding protein 5). Atlas Genet Cytogenet Oncol Haematol 2010;14(10):934-936. This work is licensed under a Creative Commons Attribution-Noncommercial-No Derivative Works 2.0 France Licence. © 2012 Atlas of Genetics and Cytogenetics in Oncology and Haematology finger motifs are thought to play roles in chromatinassociated transcriptional regulation, whereas chromodomains are frequently observed in proteins involved in recruitment of transcriptional regulatory complexes. The central portion of the protein includes a predicted DEAH-box type helicase domain (703-999) and a putative SNF2 domain (1054-1138). DEAH ATP helicases are ATP-dependent and are usually associated with nucleic acid unwinding. SNF2 domains are commonly observed in, although not restricted to, proteins involved in chromatin unwinding, DNA repair and recombination, and transcriptional regulation. A number of potential nuclear localization signals are also present (50-67, 54-71, 97-114, 100-117, 240-257, 253-270, 918-935) (Thompson et al., 2003). Identity Other names: DKFZp434N231, KIAA0444 HGNC (Hugo): CHD5 Location: 1p36.31 DNA/RNA Description Gene is encoded by 42 exons, spanning 78331 bp (NCBI). mRNA: NM_015557.2. Transcription The CHD5 cDNA sequence spans 9646 bp, with a 5' UTR of 100 bp, a single open reading frame of 5865 bp, and a 3' UTR of 3681 bp (Thompson et al., 2003). Expression Detectable expression of CHD5 was limited to all neural-derived tissues (fetal brain, total brain, cerebellum) and adrenal gland, with no expression detected in placenta, liver, fetal liver, spleen, bone marrow, thyroid, thymus, salivary gland, stomach, pancreas, small intestine, colon, or prostate (Thompson et al., 2003; Okawa et al., 2008; Wikimedia Commons). Expression is also found in testis (Potts et al., 2011; BioGPS; Brodeur, unpublished observations). Pseudogene None known. Protein Description The 5' portion of CHD5 is predicted to contain two zinc-fingers of the PHD class (amino-acid positions 345-390 and 418-463), followed closely by two chromodomains (510-525 and 596-625). PHD zinc- Atlas Genet Cytogenet Oncol Haematol. 2012; 16(6) 392 CHD5 (chromodomain helicase DNA binding protein 5) Kolla V, et al. The functional domains of the chromodomain-helicase-DNA binding protein 5 (CHD5) are shown above. There are two PHD type zinc fingers, two chromodomains, a split DEAH-box ATP-helicase domain, and a poorly defined DNA binding domain (NP_056372.1). in a panel of 101 primary tumors (Fujita et al., 2008). These data suggest that CHD5 is a bona fide tumor suppressor gene in neuroblastomas. Deletion of 1p36 is associated with loss of CHD5 expression, and the CHD5 promoter is preferentially methylated in lines with 1p36 deletion. Low CHD5 expression is also associated with a worse outcome in neuroblastoma patients. Furthermore, low CHD5 expression is associated with a worse outcome in neuroblastoma patients (Koyama et al., 2012). Most recent protein expression studies showed that CHD5 acted as immunohistochemical marker in NB in 90 primary NTs suggesting a strong association of CHD5 expression with favorable prognostic variables that were tested in a blind set of 32 NB tumors (Garcia et al., 2010). In addition to these observations this brain specific protein has been shown to regulate neural gene expression (Potts et al., 2011). Moreover, CHD5 forms a NuRD complex similar to Mi2β/CHD4, and it presumably acts as chromatin remodeling enzyme in regulating gene expression (Kolla et al., unpublished observations). Localisation Intracellular expression is preferentially (Brodeur, unpublished observations). nuclear Function CHD family proteins are thought to play a role in chromatin remodeling. CHD5 is most homologous to CHD3 and CHD4, which participate in the formation of a nucleosome remodeling and deacetylation (NuRD) complex, in association with other proteins. This complex is generally thought to repress transcription. Homology Strongest homology is to CHD3 and CHD4, with weaker homology to CHD1, CHD2, CHD6, CHD7, CHD8 and CHD9. Mutations Note No examples of homozygous inactivation have been identified, and no nonsense or frameshift mutations have been identified in any cancers to date. Several missense mutations have been detected, but it is unclear if any of these have an effect on the function of the protein (Okawa et al., 2008; Fujita et al., 2008; Ng et al., 2008; Koyama et al., 2012). Gliomas Note CHD5 is one of several genes that maps to the 700 kb region of consistent deletion in 17 glioma cell lines (Law et al., 2005). Loss of CHD5 expression is associated with deletion of 1p36 in a panel of 54 glial tumors (Bagchi et al., 2007). These data are consistent with a role for CHD5 as a tumor suppressor gene in these tumors. Implicated in Neuroblastoma Note CHD5, a new member of the chromodomain gene family, is preferentially expressed in the nervous system (Thompson et al., 2003). CHD5 maps to 1p36.3, a region of consistent deletion in neuroblastoma and other tumors (White et al., 2005). CHD5 is one of 23 genes that map to the 800 kb region of consistent deletion on 1p36.3 in an analysis of over 1200 primary neuroblastomas (Okawa et al., 2008). Functional analysis of CHD5 after transfection into neuroblastoma cell lines demonstrates that CHD5 functions as a tumor suppressor gene, suppressing both clonogenicity and tumorigenicity. The promoter was methylated especially between -780 and -450 in neuroblastoma cell lines with 1p36 deletions, but not those with two copies of CHD5. Furthermore, high CHD5 expression is highly correlated with favorable clinical and biological risk features as well as outcome Atlas Genet Cytogenet Oncol Haematol. 2012; 16(6) Melanoma Note A subset of malignant melanoma families show linkage to 1p36, and CHD5 is a tumor suppressor gene that maps to this region. However, screening of CHD5 for mutations in eight melanoma-prone families linked to 1p36 revealed no deleterious coding or splice site mutations (Ng et al., 2008). Moreover, to understand the role of CHD5 in familial melanoma, studies have shown CHD5 variants in familial cutaneous melanoma that could block or alter the ability of CHD5 to regulate the cell cycle pathway and to effect cellular control. But only one out of 47 families studied has this variant. Thus, it appears to be a rare event and further elaborative studies are required to confirm the role of CHD5 in melanoma pathogenesis (Lang et al., 2011). This suggests that CHD5 is not a major melanoma or 393 CHD5 (chromodomain helicase DNA binding protein 5) Kolla V, et al. subcutaneous melanoma susceptibility gene, at least in the families screened. Lung cancer Note Role of CHD5 in a lung cancer and epigenetic modification as well as its tumor-suppressive capability has been studied by measuring CHD5 mRNA and protein expression in lung cancer cells and tissues. CHD5 expression ranged from low to absent in the lung cancer cell lines and tissues examined correlated the results with CHD5 promoter hypermethylation. Clonogenicity and tumor growth were abrogated in lung cancer cell lines A549 and H1299 upon restoration of CHD5 expression (Zhao et al., 2011). These results suggest that CHD5 serves as a potential tumor suppressor gene in lung cancer that is inactivated via an epigenetic mechanism. Ovarian cancer Note Mutation and methylation analysis of CHD5 gene was undertaken in 123 ovarian cancers, whereas no such mutations were identified in 60 primary breast cancers. Somatic heterozygous missense mutations were identified in 3 samples, and promoter methylation was identified in another 3 of 45 samples tested (Gorringe et al., 2008). Recently, correlation of CHD5 with clinicopathological features of the tumor is also shown by quantitative RT-PCR methods. CHD5 is downregulated in a certain number of ovarian cancers and appears to be an adverse predictor candidate of ovarian cancer disease-free and total survival (Wong et al., 2011). These data suggest that CHD5 may play a role as a tumor suppressor gene in a subset of ovarian cancers, but there may be other suppressors on 1p36 as well. Prostate cancer Note Comprehensive genomic survey has been conducted for somatic events in systemic metastatic prostate tumors using both high-resolution copy number analysis and targeted mutational survey of 3508 exons from 577 cancer-related genes using next generation sequencing. Novel somatic point mutations have been shown in CHD5 along with MTOR, BRCA2, and ARHGEF12 (Robbins et al., 2011). This study indicates a deep genomic analysis of advanced metastatic prostate tumors that lead to somatic alterations, possibly contributing to lethal prostate cancer. Colorectal cancer Note The methylation status of a set of cancer-related genes was studied in 102 colon cancers from Iranian and African-American populations (51 each). The methylation status of the promoters of three genes (CHD5, ICAM5 and GPNMB) was significantly higher in cancers from the African-American population compared to the Iranian patients (Mokarram et al., 2009). This suggests that these genes may play a role in the incidence or aggressiveness of colorectal cancer in this population. Laryngeal squamous cell carcinoma Note Tumor suppressive role of CHD5 has been observed in 65 patients with laryngeal squamous cell carcinomas (LSCC) where both CHD5 RNA and protein expressions were significantly lower. These observations were also in correlation with promoter hypermethylation. Interestingly, ectopic expression of CHD5 in laryngeal cancer cells led to significant inhibition of growth and invasiveness (Wang et al., 2011). These data suggest that CHD5 acts as a tumor suppressor gene that is epigenetically downregulated in LSCC. Gastric cancer Note The methylation status of the CHD5 promoter was examined in 15 primary gastric cancers and 7 gastric cancer cell lines. CHD5 expression was downregulated in 7/7 cell lines, and methylation of the promoter was found in all 7 lines, and a similar correlation was found in 11/15 primary tumors. Ectopic expression of CHD5 led to significant growth inhibition. These results suggest that CHD5 plays a role as a tumor suppressor gene in gastric cancer, and its expression may be downregulated epigenetically. Breakpoints Note None known. Various cancers (colon cancer, breast cancer, gliomas) References Note The promoter of all nine current members of the CHD family were analyzed for methylation, and only the CHD5 promoter showed CpG hypermethylation in a subset of primary cancers, especially colon cancer, breast cancer and gliomas (Mulero-Navarro and Esteller, 2008). Thus, epigenetic inactivation of CHD5 expression may contribute to the pathogenesis of various cancers. Atlas Genet Cytogenet Oncol Haematol. 2012; 16(6) Thompson PM, Gotoh T, Kok M, White PS, Brodeur GM. CHD5, a new member of the chromodomain gene family, is preferentially expressed in the nervous system. Oncogene. 2003 Feb 20;22(7):1002-11 Law ME, Templeton KL, Kitange G, Smith J, Misra A, Feuerstein BG, Jenkins RB. Molecular cytogenetic analysis of chromosomes 1 and 19 in glioma cell lines. Cancer Genet Cytogenet. 2005 Jul 1;160(1):1-14 394 CHD5 (chromodomain helicase DNA binding protein 5) Kolla V, et al. White PS, Thompson PM, Gotoh T, Okawa ER, Igarashi J, Kok M, Winter C, Gregory SG, Hogarty MD, Maris JM, Brodeur GM. Definition and characterization of a region of 1p36.3 consistently deleted in neuroblastoma. Oncogene. 2005 Apr 14;24(16):2684-94 Casala C, Galván P, de Torres C, Mora J, Lavarino C. Expression of the neuron-specific protein CHD5 is an independent marker of outcome in neuroblastoma. Mol Cancer. 2010 Oct 15;9:277 Bagchi A, Papazoglu C, Wu Y, Capurso D, Brodt M, Francis D, Bredel M, Vogel H, Mills AA. CHD5 is a tumor suppressor at human 1p36. Cell. 2007 Feb 9;128(3):459-75 Lang J, Tobias ES, Mackie R. Preliminary evidence for involvement of the tumour suppressor gene CHD5 in a family with cutaneous melanoma. Br J Dermatol. 2011 May;164(5):1010-6 Fujita T, Igarashi J, Okawa ER, Gotoh T, Manne J, Kolla V, Kim J, Zhao H, Pawel BR, London WB, Maris JM, White PS, Brodeur GM. CHD5, a tumor suppressor gene deleted from 1p36.31 in neuroblastomas. J Natl Cancer Inst. 2008 Jul 2;100(13):940-9 Potts RC, Zhang P, Wurster AL, Precht P, Mughal MR, Wood WH 3rd, Zhang Y, Becker KG, Mattson MP, Pazin MJ. CHD5, a brain-specific paralog of Mi2 chromatin remodeling enzymes, regulates expression of neuronal genes. PLoS One. 2011;6(9):e24515 Gorringe KL, Choong DY, Williams LH, Ramakrishna M, Sridhar A, Qiu W, Bearfoot JL, Campbell IG. Mutation and methylation analysis of the chromodomain-helicase-DNA binding 5 gene in ovarian cancer. Neoplasia. 2008 Nov;10(11):1253-8 Robbins CM, Tembe WA, Baker A, Sinari S, Moses TY, Beckstrom-Sternberg S, Beckstrom-Sternberg J, Barrett M, Long J, Chinnaiyan A, Lowey J, Suh E, Pearson JV, Craig DW, Agus DB, Pienta KJ, Carpten JD. Copy number and targeted mutational analysis reveals novel somatic events in metastatic prostate tumors. Genome Res. 2011 Jan;21(1):47-55 Isidor B, Le Cunff M, Boceno M, Boisseau P, Thomas C, Rival JM, David A, Le Caignec C. Complex constitutional subtelomeric 1p36.3 deletion/duplication in a mentally retarded child with neonatal neuroblastoma. Eur J Med Genet. 2008 Nov-Dec;51(6):679-84 Wang J, Chen H, Fu S, Xu ZM, Sun KL, Fu WN. The involvement of CHD5 hypermethylation in laryngeal squamous cell carcinoma. Oral Oncol. 2011 Jul;47(7):601-8 Wong RR, Chan LK, Tsang TP, Lee CW, Cheung TH, Yim SF, Siu NS, Lee SN, Yu MY, Chim SS, Wong YF, Chung TK. CHD5 Downregulation Associated with Poor Prognosis in Epithelial Ovarian Cancer. Gynecol Obstet Invest. 2011;72(3):203-7 Mulero-Navarro S, Esteller M. Chromatin remodeling factor CHD5 is silenced by promoter CpG island hypermethylation in human cancer. Epigenetics. 2008 Jul-Aug;3(4):210-5 Ng D, Yang XR, Tucker MA, Goldstein AM. Mutation screening of CHD5 in melanoma-prone families linked to 1p36 revealed no deleterious coding or splice site changes. BMC Res Notes. 2008 Sep 19;1:86 Zhao R, Yan Q, Lv J, Huang H, Zheng W, Zhang B, Ma W. CHD5, a tumor suppressor that is epigenetically silenced in lung cancer. Lung Cancer. 2012 Jun;76(3):324-31 Okawa ER, Gotoh T, Manne J, Igarashi J, Fujita T, Silverman KA, Xhao H, Mosse YP, White PS, Brodeur GM. Expression and sequence analysis of candidates for the 1p36.31 tumor suppressor gene deleted in neuroblastomas. Oncogene. 2008 Jan 31;27(6):803-10 Cai C, Ashktorab H, Pang X, Zhao Y, Sha W, Liu Y, Gu X. MicroRNA-211 expression promotes colorectal cancer cell growth in vitro and in vivo by targeting tumor suppressor CHD5. PLoS One. 2012;7(1):e29750 Koyama H, Zhuang T, Light JE, Kolla V, Higashi M, McGrady PW, London WB, Brodeur GM. Mechanisms of CHD5 Inactivation in neuroblastomas. Clin Cancer Res. 2012 Mar 15;18(6):1588-97 Mokarram P, Kumar K, Brim H, Naghibalhossaini F, Saberifiroozi M, Nouraie M, Green R, Lee E, Smoot DT, Ashktorab H. Distinct high-profile methylated genes in colorectal cancer. PLoS One. 2009 Sep 11;4(9):e7012 This article should be referenced as such: Wang X, Lau KK, So LK, Lam YW. CHD5 is down-regulated through promoter hypermethylation in gastric cancer. J Biomed Sci. 2009 Oct 19;16:95 Kolla V, Higashi M, Zhuang T, Brodeur GM. CHD5 (chromodomain helicase DNA binding protein 5). Atlas Genet Cytogenet Oncol Haematol. 2012; 16(6):392-395. Garcia I, Mayol G, Rodríguez E, Suñol M, Gershon TR, Ríos J, Cheung NK, Kieran MW, George RE, Perez-Atayde AR, Atlas Genet Cytogenet Oncol Haematol. 2012; 16(6) 395