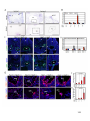
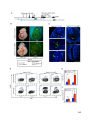
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Notch activation and downstream targets in embryonic hematopoiesis Jordi Guiu Sagarra TESI DOCTORAL UPF 2012 THESIS SUPERVISORS/DIRECTORS: Dr. Anna Bigas Dr. Lluís Espinosa Programa de Càncer. Institut Hospital del Mar d’Investigacions Mèdiques. i ii iii iv AGRAÏMENTS (ACKNOLEDGEMENTS) Gràcies a tothom amb qui he compartit aquest camí, als que anomenaré i també als que no anomenaré ja que podria omplir una tesi només de noms. A la gent del laboratori, als caps: el Lluís Espinosa, l’Anna Bigas i als companys: la Teresa D’Altri, la Mari Mulero, el Pol Margalef, la Verónica Rodilla, la Vanessa Fernandez, la Cristina Ruiz, la Júlia Inglès, l’Eva Ferrando, l’Érika López, la Leonor Norton, el Jorik, l’Albert Batet, el Dylan, l’Alba, el Ricard, l’Àlex Robert, la MELS i en general a tota la gent del Programa de Càncer. També als meus companys de laboratori Japonesos, vaig passar amb ells 4 mesos, al Hiroshi Kaneko, a l’Eri Kobayashi a la Yukie Kawatani, al Itsushi Hasegawa i al Morita, també als caps: Ritsuko Shimizu i Masayuki Yamamoto. Als serveis cientificotècnics del PRBB, especialment a la Unitat de Citometria, a l’Òscar, l’Erica i la Sabrina. Evidentment, vull donar també les gràcies a la gent de l’entorn no científic, altre cop a la Teresa als amics i a la família. També gràcies a L’Institut Hospital del Mar d’Investigacions Mèdiques per fer-se càrrec de les despeses de impressió. v vi ABSTRACT The aorta-gonad-mesonephros (AGM) is the first Hematopoietic stem cell (HSC) niche. It was previously shown that Notch pathway is required to induce the arterial fate as well as to generate Hematopoietic Stem Cells. HSC emerge at the site of arterial vessels during embryonic development. Since arterial fate precedes HSC generation, it has long been controversial whether Notch exclusively induces the arterial program and the lack of HSC is a secondary defect; or Notch is directly involved in activating both genetic programs, arterial and hematopoietic. The best-characterized Notch targets are Hes and Hes-related genes (Hrt) since they are involved in most of described Notch functions, however there is a growing number of tissuespecific transcriptional Notch-targets. In this thesis, we found that Jagged-mediated activation of Notch is required for the correct execution of the definitive hematopoietic program but not for the establishment of the arterial fate, thus demonstrating that Notch exerts a specific hematopoietic function in the embryo. Downstream of Notch pathway, we also show that embryos deficient for Hes1 and Hes5 alleles contain an intact arterial program but produce increased numbers of non-functional hematopoietic stem cells associated to higher levels of the hematopoietic regulators Runx1, Myb and Gata2. Moreover, Gata2 transcription is positively regulated by Notch and negatively controlled by vii HES-1. This creates an incoherent feed-forward loop that tightly controls Gata2 levels to generate HSC. viii RESUM La regió de l’aorta-gonada-mesonefros (AGM) és el primer nínxol de les cèl·lules mare hematopoètiques. Està descrit que la via de Notch és necessària per generar artèries i cèl·lules mare hematopoètiques. Les cèl·lules mare hematopoètiques es generen a partir de les artèries durant el desenvolupament embrionari. Tenint en compte que els vasos arterials es formen abans que les cèl·lules mare hematopoètiques, durant molt temps ha estat controvertit si la via de Notch només indueix la formació d’artèries i en canvi l’abscència de cèl·lules mare hematopoètiques és un efecte secundari o si per contra, la via de Notch participa activament en la inducció d’ambdós programes genètics, l’arterial i l’hematopoètic. Moltes de les funcions descrites de la via de Notch són conseqüència de l’expressió de les seves dianes transcripcionals: Hes i Hes-related (Hrt). No obstant cada cop s’estan identificant noves dianes transcripcionals de la via de Notch que són específiques de teixit. En aquesta tesis doctoral, demostrem que l’activació de la via de Notch per part del seu lligand Jagged1 és necessària per activar el programa hematopoètic però no cal per establir el programa arterial. Aquest fet demostra que la via de Notch juga un paper clau i directe en l’hematopoèsi embrionaria. Per sota de la via de Notch mostrem que els embrions deficients en Hes1 i Hes5 mantenen intacte el programa arterial. Cal remarcar que també generen més quantitat de cèl·lules mare ix hematopoètiques però que no són funcionals. A més tal mutants, tenen nivells més alts d’expressió dels gens Runx1, Myb i Gata2; els quals són importants reguladors de l’hematopoèsi. Per acabar demostrem que Notch activa la transcripció de Gata2 i que HES-1 l’inhibeix. Això genera un bucle de retroalimentació incoherent (Incoherent Feed-Fordward loop) , el qual regula estretament els nivells de Gata2 necessàris per generar cèl·lules mare hematopoètiques. x xi xii PREFACE This thesis is submitted to obtain the Doctor degree at Universitat Pompeu Fabra. The work shown has been supervised by Dr. Anna Bigas and Dr. Lluís Espinosa in Institut Hospital del Mar d’Investigacions Mèdiques (IMIM), Barcelona. This work was performed between September 2007 and June 2012. A relevant issue in the field of regenerative medicine is how to produce functional stem cells for future transplantation applications, which is still unknown. We addressed this issue by studying the molecular events and pathways involved in the generation of the first HSCs in the embryo since evidence shows that this is the only time that “de novo” HSC formation occurs. Dr. Spyros Artavanis-Tsakonas and Dr. John Yochem cloned the Notch gene in Drosophila melanogaster and Caenorhabditis elegans in 1983 and 1988 respectively. Then it was characterized in different animal models including mammals. Notch is a key regulator of development as well an inducer of specific cell fates in the adult. Previously it was shown that Notch is dispensable to maintain or expand HSCs in the adult but required during embryonic hematopoietic development. Intriguingly HSCs emerge from the embryonic arteries, a tissue that requires Notch activity to be generated. Consequently, it is important to understand the contribution of Notch xiii function to each of these events: arterial formation and HSC generation. To elucidate this, it is instrumental to identify Notch ligands and downstream target genes, which help to distinguish both functions. In this thesis we identified Jagged1 as specific Notch ligand as well as Notch transcriptional targets (Gata2 and Hes1) that only affect the hematopoietic program but not arterial one, which demonstrated the existence of two different functions. This is a paper-based thesis, and includes a general introduction of embryonic hematopoietic development and Notch pathway, which is the aim of our work, two scientific manuscripts (one published in EMBO JOURNAL and the other under second revisions in Journal Experimental Medicine), the conclusions and the discussion of the work. Collaborations have been important in the accomplishment of this work such as Dr M. Yamamoto, Dr. R. Shimizu, Dr. E. Bresnick, Dr. E. Dzierzak and Dr. R. Kageyama l, Dr. T. Enver and Dr. T. Gridley. This work was supported by Ministerio de Economía y Competitividad (SAF2007-60080, PLE2009-0111, SAF2010-15450), Red Temática de Investigación Cooperativa en Cáncer (RTICC) (RD06/0020/0098), Agència de Gestió d’Ajuds Universitaris i de Recerca (AGAUR) (2009SGR-23, CONES20100006 and a personal fellowship FPI BES-2008-005708. xiv xv xvi SUMMARY Page Agraïments (ACKNOLEDGEMENTS) ........................... v Abstract (ENGLISH) .................................................................... vii Resum (CATALÀ) ........................................................................ ix Preface ........................................................................................ xiii Summary ..................................................................................... xvii List of figures and tables .............................................................. xxiii CHAPTER 1. GENERAL INTRODUCTION ................................. 1 1.1. HEMATOPOIETIC SYSTEM ................................................ 3 1.1.1. Introduction to the hematopoietic system .......................... 3 1.2. ONTOGENY OF HEMATOPOIETIC SYSTEM IN THE MOUSE EMBRYO ..................................................................................... 5 1.2.1. Introduction to embryo development ................................. 5 1.2.2. Hematopoietic origin in the mouse embryo ....................... 7 1.2.3. Characterization of the hematopoietic cell types ............... 9 1.1.3.1. Phenotypic characterization ........................................... 10 1.1.3.1a) Cell surface marker expression analysis ...................... 13 1.1.3.2. Functional characterization of progenitors and HSC ...... 13 xvii 1.1.3.2.a) CFU-C ......................................................................... 13 1.1.3.2.b) BFU-E .......................................................................... 14 1.1.3.2.c) CFU-GM ...................................................................... 15 1.1.3.2.d) CFU-Mix ...................................................................... 15 1.1.3.2.e) CFU-S ......................................................................... 15 1.1.3.2.f) Short-term and Long-Term Reconstitution assay......... 16 1.1.3.2.g) Limiting dilution transplantation assay......................... 17 1.1.3.2.h) Newborn reconstitution ................................................ 18 1.1.3.2.i) Explant culture .............................................................. 19 1.1.3.2.j) Aggregation assay ........................................................ 19 1.1.3.2.k) Discrimination of cells from donor vs. recipients ......... 20 1.2.4. Primitive hematopoietic wave ............................................ 20 1.2.4.1. Yolk Sac hematopoiesis ................................................. 22 1.2.5. Definitive hematopoietic wave ........................................... 25 1.2.5.1. P-Sp/AGM and major arteries ........................................ 25 1.2.5.2. Arterial fate precedes HSC generation ........................... 28 1.2.5.3. Placenta and fetal liver hematopoiesis ........................... 29 1.3. REQUIREMENTS IN THE GENERATION OF HSC ............ 32 1.3.1. Introduction ........................................................................ 32 1.3.2. Mechanical inputs/outputs ................................................. 32 1.3.2.1. Cell budding and Rac GTPases ..................................... 33 1.3.2.2. Nos and shear stress ...................................................... 33 1.3.3. Chemical inputs/outputs .................................................... 34 xviii 1.3.3.1. Interleukine-3 .................................................................. 35 1.3.3.2. Stem Cell Factor ............................................................ 36 1.3.3.3. Sonic Hedgehog ............................................................. 36 1.3.3.4. Bone morphogenetic proteins......................................... 37 1.3.3.5. E-Twenty six transcription factors ................................... 37 1.3.3.6. Run-related transcription factor 1 ................................... 38 1.3.3.7. GATA transcription factors ............................................. 39 1.3.3.8. Wnt pathway ................................................................... 41 1.3.3.9. c-Myb .............................................................................. 42 1.3.3.10. Stem Cell Leukemia/T-cell acute leukemia-1 ............... 43 1.3.3.11. HoxB4 ........................................................................... 43 1.3.3.12. Notch pathway .............................................................. 44 1.4. NOTCH PATHWAY .............................................................. 45 1.4.1. Introduction to the Notch pathway ..................................... 45 1.4.2. Notch receptors ................................................................. 51 1.4.3. Notch receptor modifiers: Rumi, Pofut and fringe.............. 53 1.4.4. Notch Ligands .................................................................... 55 1.4.5. Receptor-ligand binding. Notch Activation......................... 56 1.4.6. Endocytosi in Notch activation........................................... 57 1.4.7. Transcription of Notch target genes .................................. 60 1.4.8. Role of Notch ..................................................................... 62 1.4.8.1. Cell fate determination .................................................... 62 1.4.8.2. Lateral inhibition ............................................................. 62 xix 1.4.8.3. Lateral induction ............................................................. 63 1.4.8.4. Notch and stem cells ...................................................... 65 1.4.8.4.a) Neurogenesis .............................................................. 65 1.4.8.4.b) Intestine ....................................................................... 65 1.4.8.4.c) Hair follicle ................................................................... 66 1.4.8.4.d) Adult hematopoiesis .................................................... 67 1.4.8.4.e) Embryonic hematopoiesis ........................................... 68 1.4.8.5. Notch and differentiated specialized cells ................... 69 1.4.8.5.a) T lymphocytes generation ........................................... 69 1.4.8.5.b) Arteries vs. veins ......................................................... 70 CHAPTER 2. OBJECTIVES ........................................................ 75 CHAPTER 3. RESULTS (PUBLICATIONS) ............................... 79 3.1. Paper 1: Impaired embryonic hematopoiesis yet normal arterial development in the absence of the Notch ligand Jagged1. ........ 81 3.1.1. Abstract ............................................................................ 83 3.1.2. Introduction ........................................................................ 84 3.2.3. Results ............................................................................... 87 3.1.3.1. Heterogeneous expression of Notch family members within the aortic haematopoietic clusters ..................................................... 87 3.1.3.2. Altered haematopoiesis in the AGM of Jagged1 but not Jagged2 mutant embryos ........................................................................... 98 xx 3.1.3.3. Arterial fate is not affected in the Jagged1 mutant embryos 91 3.1.3.4. GATA2 expression is compromised in Jagged1 mutants 95 3.1.3.5. Ectopic GATA2 expression rescues Jagged1 mutant haematopoiesis ........................................................................... 98 3.1.4. Discussion ......................................................................... 100 3.1.5. Materials and Methods ...................................................... 104 3.1.5.1. Animals ........................................................................... 104 3.1.5.2. Haematopoietic colony assay ......................................... 104 3.1.5.3. Haematopoietic liquid cultures........................................ 104 3.1.5.4. RT–PCR ......................................................................... 105 3.1.5.5. Lentiviral constructs and infection .................................. 106 3.1.5.6. Flow cytometry analysis ................................................. 106 3.1.5.7. Chromatin immunoprecipitation assay ........................... 107 3.1.5.8. Immunostaining .............................................................. 107 3.1.5.9. WISH .............................................................................. 108 3.1.5.10. Image acquisition .......................................................... 108 3.1.6. Aknowledgements ............................................................. 109 3.1.7. References ........................................................................ 109 3.1.8. Supplementary figures....................................................... 119 3.2. Paper2: Hes repressors are essential regulators of Hematopoietic Stem Cell Development downstream of Notch signaling. ........... 121 3.2.1. Abstract ............................................................................. 124 3.2.2. Introduction ........................................................................ 126 xxi 3.2.3. Results ............................................................................... 128 3.2.3.1. Increased hematopoietic cluster emergence in Hes-deficient embryonic aortas. ....................................................................... 128 3.2.3.2. Occupancy of the Gata2 promoter by HES-1 and Notch/RBP-Jκ ..................................................................................................... 130 3.2.3.3. HES-1 is a negative regulator of the Gata2 promoter in hematopoietic cells. .................................................................... 136 3.2.3.4. Notch/RBP-Jκ and HES control Gata2 expression during embryonic development ............................................................. 138 3.2.3.5. AGM hematopoietic clusters in Hes-deficient embryos lack functional progenitors and HSCs. ............................................... 140 3.2.4. Discussion ......................................................................... 146 3.2.5. Materials and Methods ...................................................... 150 3.2.5.1. Animals ........................................................................... 150 3.2.5.2. Quantitative RT–PCR ..................................................... 150 3.2.5.3. Promoter analysis, site-directed mutagenesis and luciferase assays. ....................................................................................... 151 3.2.5.4. Chromatin immunoprecipitation assay ........................... 152 3.2.5.5. Cell culture, viral particle production and viral infection.. 152 3.2.5.6. Flow cytometry analysis and sorting............................... 153 3.2.5.7. Immunostaining .............................................................. 154 3.2.5.8. In Situ Hybridization ........................................................ 155 3.2.5.9. Transgenic mice. F0 assays ........................................... 155 3.2.5.10. AGM explant culture ..................................................... 156 xxii 3.2.5.11. Hematopoietic colony assay (CFU-C) and CFU-S11 and long-term reconstitution assay ..................................................................... 156 3.2.5.12. Statistics ....................................................................... 157 3.2.6. Acknowledgements ........................................................... 157 3.2.7. References ........................................................................ 158 3.2.8. Supplementary figures....................................................... 165 CHAPTER 4: CONCLUSIONS .................................................... 171 4.1. CONCLUSIONS ................................................................... 173 4.1.2.Notch ligands and receptors ............................................... 173 4.1.3.Notch target genes ............................................................. 173 CHAPTER 5: DISCUSSION ........................................................ 177 5.1. DISCUSSION ....................................................................... 179 5.1.1. Notch regulates hematopoietic program through Jagged1 179 5.1.2. Gata2 is a direct Notch target in the AGM ......................... 180 5.1.3. Gata2 is a direct Hes-1 target in the AGM ......................... 181 5.1.4. Notch, Hes-1 and Gata2 generate an incoherent Feed-Forward loop ..................................................................................................... 182 5.1.5. Notch ligands and Notch target specificity. Arteries, veins and hematopoiesis ............................................................................. 183 xxiii CHAPTER 6: BIBLIOGRPHY ...................................................... 185 7.ABBREVIATIONS ..................................................................... 217 xxiv LIST OF FIGURES AND TABLES Introduction: Figures: Figure1: Adult hematopoiesis and lymphoid-myeloid branching points Figure2: First staged of mouse development Figure3: Picture of the main hematopoietic organs during development. Figure4: HSCs emerge from aortic endothelium. Figure5: Immunofluorescence of hematopoietic clusters emerging from the aorta. Figure6: Functional hematopoietic assays. Figure7: Hipothetical competitive transplantation assay. Figure8: Hipothetical limiting dilution transplantation assay. Figure9: Chronology of the first hematopoietic stem/progenitor cells during mouse embryo development from E6 to E10,5. Figure10: Hematopoietic organs and main hematopoietic events during embryonic development. Figure11: Human/Mouse Gata2 locus. Figure12: Notch in Drosophila wings. Figure14: Notch receptor modifications. Figure15: Notch pathway. xxiii Figure16: Notch activates target genes in the nucleus. Figure17: Lateral inhibition and lateral induction. Figure18: Notch promotes arterial fate at expenses of veins. Figure19: Lateral inhibition and induction models in the AGM. Tables: Table1: Notch signaling regulates numerous developmental processes. Results: Paper1: Impaired embryonic hematopoiesis yet normal arterial development in the absence of the Notch ligand Jagged1. Figures paper1: Figure1: Heterogeneous expression of Notch family members within the aortic haematopoietic clusters. Figure 2. Altered haematopoiesis in the AGM of Jagged1 but not Jagged2 mutant embryos. Figure 3. Arterial fate is not affected in the Jagged1 mutant embryos. Figure 4. GATA2 expression is compromised in Jagged1 mutants. Figure 5.Ectopic GATA2 expression rescues Jagged1 mutant haematopoiesis. xxiv Figure S1: Gata2 expression is increased in Jag1 Δ/Δ cells after culture on OP9 stromal cells. Figure S2: Ectopic expression od gata2 in Jag1 Δ/Δ AGM cells helps runx1 expression. Tables paper 1: Table1: Number of Gata2- and Runx1-positive cells per AGM as detected by WISH in the endothelium of the dorsal aorta in different Jagged 1 and Jagged 2 mutants. Paper2: Hes repressors are essential regulators of Hematopoietic Stem Cell Development downstream of Notch signaling. Figures paper2: Figure 1: Increased hematopoietic cluster emergence in Hes-deficient embryonic aortas. Figure 2:. Occupancy of the Gata2 promoter by HES-1 and Notch/RBP-Jκ. Figure3: HES-1 is a negative regulator of the Gata2 promoter in hematopoietic cells. Figure 4: Notch/RBP-Jκ and HES-1 regulates Gata2 promoter in vivo during embryonic development. Figure 5: AGM hematopoietic clusters in Hes-deficient embryos lack functional progenitors and HSCs. xxv Figure 6: Model for Notch control of Gata2 doses through Hes in the nascent HSC. Tables paper2: Table S1: Mouse RT-PCR primers. Table S2: Mutagenesis primers. Table S3: Mouse ChIP primers. Table S4: Hes lox-p sites primers to detect deleted bands. xxvi xxvii 1. GENERAL INTRODUCTION 1 2 1.1. HEMATOPOIETIC SYSTEM 1.1.1. Introduction to the hematopoietic system. Hematopoiesis is the process defined by the differentiation of hematopoietic precursors and production of all mature blood cells. The blood is formed by multiple cell types with a lifespan of several days (i.g. two days for neutrophils and thirty days for erythrocytes). In 1970, Metcalf and Moore and Till & McCullough demonstrated that HSC and multipotent progenitors reside in the adult Bone Marrow (BM), which is the niche for HSC maintenance and self-renewal. Two main models have been proposed to explain the adult hematopoietic hierarchy (Figure 1A and 1B). The differences in these two models are the branching points between myeloid and lymphoid lineages. Two essential characteristics of stem cells are multipotency and selfrenewal (Figure 1). Multipotency refers to their capacity to give rise to different hematopoietic cell types. Self-renewal refers to the capacity that allows the maintenance of the stem cell pool. Asymmetric division is the most probable mechanistic explanation for the balance between selfrenewal and differentiation. HSCs can alternate between symmetric and asymmetric divisions to generate differentiated blood cells and maintain the homeostasis of the system (Brummendorf, Dragowska et al. 1998; Suda, 3 Suda et al. 1984; Leary, Strauss et al. 1985; Wu, Kwon et al. 2007; Beckmann, Scheitza et al. 2007). a) b) Figure1. Adult hematopoiesis and lymphoid-myeloid branching points (labeled in gray). (a) The Akashi-Kondo-Weisman model of hematopoiesis. (b)Revised branching point between lymphoid and myeloid lineages. (LT- and ST-HSC, long and short-term HSC; MPP, multipotent progenitor; ETP, early thymic progenitor; CLP, common myeloid progenitor; CMP, common myeloid progenitor; MEP, myeloertythroid progenitor; GMP, granulocyte monocyte progenitor) (Figure adapted from Laiosa, Stadtfeld et al. 2006; Yokomizo and Dzierzak 2010) 4 1.2. ONTOGENY OF HEMATOPOIETIC SYSTEM IN THE MOUSE EMBRYO 1.2.1. Introduction to murine embryo development From a one-cell zygote to a mouse embryo many events occur (Figure 2). After three cell divisions the embryo reaches the 8-cell stage at E2. At this point, the embryo initiates a compaction event that increases cell-cell contact and cell adhesion thus generating an apical-basal polarity in each cell. The embryo continues to grow, from E2.5 to E3 (8 to 26 cells). Once it reaches the spherical shape due to the high compaction it is called morula. After the morula stage and two rounds of asymmetric divisions, there is formation of a cavity inside, the blastocoel, and the whole structure is then called blastocyst. This takes place at E3.5 (32 to 64 cells). Due to the asymmetric divisions, the blastocyst has inner and outer cells, which are called inner mass cells (IMC) and the outer epithelial layer, trophoectoderm (TE). At E4.5 the blastocyst reaches the uterus and implants into the wall of the oviduct. IMC is then segregated into primitive endoderm (PrE) and epiblast. PrE is the epithelial layer that will contribute to the endoderm. Around E6.0 the epiblast begins the differentiation program and the three germ layers (ectoderm, mesoderm and endoderm) are originated, in a process called gastrulation. 5 !"#$%&' 6?' 6>' D349%$,"9%' 1$)E34' 6?@A' 6B' 6B@A' 6C@A' 6*78349%' !"#$%&' 6,%$-&).' 1&9$-&).' 65-$-&).' ''''''''''''''''''''''''''''/01' 2)7.7:;&'65-$-&).'' '''''''''''''''''''''''()$*+&,%$-&).' <$3='94,' 234,&5%4' Figure2. First stages of mouse embryonic development. Schematic representation from the zygot to E6 when different germ layers (ectoderm, mesoderm and endoderm) are formed. Around day E7.5 the first hematopoietic cells, mainly nucleated erythrocytes are found in the yolk sac. At E8,5 the beating of the heart starts and blood circulation is established between the extraembryonic tissues (yolk sac and placenta) and the embryo proper. This event is extremely relevant when studying the embryonic hematopoiesis and the generation of HSC since circulation between the embryo and the extraembryonic tissues have extremely difficulted to establish the origin of these cells. The origin and embryonic history of the different tissues is crucial to understand the behavior of the adult cell types and tissues: 6 -Ectoderm: it gives rise to central nervous system (brain and spinal cord) and peripheral nervous system, outer surface or skin of the organism, cornea and lens of the eye, epithelium that lines the mouth and nasal cavities and the anal canal, epithelium of the pineal gland, pituitary gland, and adrenal medulla, and cells of the neural crest (which gives rise to various facial structures, pigmented skin cells called melanocytes, and dorsal root ganglia, clusters of nerve cells along the spinal cord). -Mesoderm: it gives rise to skeletal, smooth, and cardiac muscle, structures of the urogenital system (kidneys, ureters, gonads, and reproductive ducts), endothelium, hematopoietic cells, mesenchymal tissues, bone marrow, fat bone, cartilage and the lining of the body cavity. -Endoderm: it gives rise to the epithelium of the entire digestive tract (excluding the mouth and anal canal), epithelium of the respiratory tract, structures associated with the digestive tract (liver and pancreas), thyroid, parathyroid, and thymus glands, epithelium of the reproductive ducts and glands; epithelium of the urethra and bladder. 1.2.2. Hematopoietic origin in the mouse embryo Embryonic hematopoiesis starts after gastrulation, when a subset of specialized primitive streak mesodermal cells expressing Flk-1 (Shalaby, Rossant et al. 1995; Shalaby, Ho et al. 1997) migrate to the YS and around 7 E7.5 will begin to produce primitive hematopoietic cells. HSC will emerge later in another locations (see chapter 1.2.5.1). In the embryo several tissues have hematopoietic activity such as yolk sac (YS), Para-aortic splachnopleura (PSp), aorta-gonad-mesonephros (AGM), placenta and fetal liver (FL) (Figure3-4). We can divide the embryonic hematopoiesis in two main waves (Reviewed in Dzierzak and Speck 2008), which are called primitive and definitive. The different waves occur in different embryonic tissues and generate different blood cell types. Figure3. Picture of the main hematopoietic organs during embryonic development: AGM, Placenta, YS and FL. (Photo from Gekas, Dieterlen-Lievre et al. 2005) During the last decades multiple functional assays have been developed to characterize the hematopoietic cells and progenitors in the adult bone marrow. However, these assays need to be adapted and validated based on the developing nature of embryonic hematopoietic cells. 8 1.2.3. Characterization of the Hematopoietic Cell types Characterization and stratification of Hematopoietic cell types is the result of combining different assays and technologies. In general, this includes the analysis of cell morphology (Wright-Giemsa staining, Figure 4), expression of surface proteins, progenitor colony cultures and transplantation assays. In the adult, the HSC niche is the bone marrow, however in the embryo hematopoiesis takes place at different sites. Embryonic HSCs emerge from hematopoietic cluster structures present in aortic vessels (Figure 4) (de Bruijn, Speck et al. 2000). These cells express specific markers and their characterization is detailed in the next chapter. The presence of HSC or progenitors in the developing mouse tissues is studied by phenotypic characterization and laborious functional assays, which are next described in order to understand HSC biology. 1.2.3.1. Phenotypic characterization. HSCs express specific surface molecules, which are used for their identification. Surface markers expressed by adult HSC are well characterized. In mouse bone marrow HSC have the following markers: SCA-1-positive, c-Kit-positive, CD150-positive, lineage-negative and CD48negative (Kiel, Yilmaz et al. 2005). However embryonic HSC markers are still under investigation. 9 1.2.3.1a) Cell surface marker analysis: During HSC development, as previously mentioned, HSCs emerge from the aortic endothelium by a process that involves endothelium to hematopoietic transition. For this reason, it is important to identify markers that allow to distinguish the endothelial cells from endothelial-like cells with hematopoietic potential (also known as hemogenic endothelium) and already committed embryonic HSC and progenitors. Some markers have been shown to be expressed in the HSCs during mouse embryonic development (Figure5), and different combinations of markers have been used to characterize embryonic progenitors and HSC. Ly-6A.2/Ly-6E.1/SCA-1 This molecule is a well-characterized adult HSC marker, however it is weakly expressed in the AGM. Using a transgenic mouse line that express GFP under the control of the SCA-1 promoter, it was demonstrated that embryonic cells expressing this transgen were able to reconstitute the hematopoiesis of irradiated animals (de Bruijn, Ma et al. 2002). CD41/integrin alpha-IIb Studies from Robin et al (Robin, Ottersbach et al. 2011) demonstrated that HSC activity was contained only in the CD41-intermediate intensity cells, as determined by their capacity to reconstitute the hematopoisesis when transplanted into lethally irradiated mice. In these experiments neither CD41-negative nor CD41-High were able to engraft. 10 CD117/c-Kit/Tyrosine-protein kinase KIT Most of the cells from the Aortic endothelium and hematopoietic clusters of the AGM region express the CD31 (PECAM-1) marker, however the emerging clusters are clearly distinguished by the expression of c-Kit. Thus cells in the AGM aorta and clusters can be classified as endothelial cells (CD31-positive/c-Kit-negative); negative/c-Kit-positive) and hematopoietic HSCs progenitors (CD31- (CD31-positive/c-Kit-positive). Cell sorting of dissected AGMs was used to demonstrate that HSC activity with transplantation capacity resides within the CD31-positive/c-Kit-positive population. However, both CD31-positive/c-KIT-positive and CD31- negative/c-KIT-positive were able to generate hematopoietic colonies in CFU-C assay (Yokomizo and Dzierzak 2010). CD144/VE-cadherin Previous work from Medvinsky’s lab identified three different subpopulations in the AGM based on the expression of VE-cadherin, CD45 and CD41. First, they showed that the HSC activity was constrained in the VE-cadherin-positive/CD45-positive population while no activity was found in the VE-cadherin-positive/CD45-negative, indicating that HSCs are CD45positive. However, a fraction of VE-cadherin-positive/CD45-negative cells can develop HSC activity when cultured 4 days in coagregates (see chapter 1.2.3.2j) with OP9 stroma. They further demonstrate that these cells were included in the population that expresses intermediate levels of CD41 11 (Rybtsov, Sobiesiak et al. 2011). Thus, by using these three markers they distinguish between: endothelial negative/CD45-negative), intermediate/CD45-negative) cells PRE-HSCs and HSCs: (VE-cadherin-positive/CD41(VE-cadherin-positive/CD41VE-cadherin-positive/CD41- intermediate/CD45-positive. sca / CD41 C-KIT / CD31 CD45 / CD31 CD45 / CD41 / CD31 Figure5. Immunofluorescence of Hematopoietic clusters emerging from the aorta. (a) CD41 overlaps with sca positive cells. (b) Endothelial cells and hematopoietic clusters are CD31-positive/C-KIT-negative and hematopoietic clusters are CD31-positive/CKITpositive; few cells of the cluster are CD45 positive (c). (d) Most of the cluster cells are CD41-positive. (Photos extracted from Robin, Ottersbach et al. 2011; Yokomizo and Dzierzak 2010) Altogether, the current paradigm is that VE-cadherin and CD31 labels the endothelial layer and the hematopoietic clusters (see chapter 1.2.5 and 1.2.5.1), c-Kit (Yokomizo and Dzierzak 2010) and CD41 (Robin, Ottersbach et al. 2011); (Rybtsov, Sobiesiak et al. 2011) cells are mainly located in the 12 hematopoietic clusters (Figure 5) and CD45 labels some cells in the outer part of the clusters (Yokomizo and Dzierzak 2010) , most likely containing the first definitive HSC of the embryo (Rybtsov, Sobiesiak et al. 2011). The detection of surface markers although it is informative, it is not a demonstration to determine the presence of HSC activity. To assess the presence of HSC/P functional assays need to be performed. 1.2.3.2. Functional characterization of progenitors and HSC Several functional assays are used to test the presence of hematopoietic progenitors: colony-forming units in methylcellulose culture (CFU-C) and colony-forming units in the mouse spleen at day 11 post injection (CFUS11). Importantly, Long-Term-Reconstitution assays (LTR-assay) are the only measure for the presence of functional HSC. 1.2.3.2a) CFU-C: CFU-C assay is an in vitro assay commonly used to quantify the number of functional hematopoietic progenitor cells. The assay is based on the ability of hematopoietic progenitors to proliferate and differentiate into colonies in semi-solid media containing the appropriate cytokines. These colonies can be characterized according to their morphology and counted. The different types of colonies that can be identified in these assays are: Burst forming unit Erythroid (BFU-E); Colony forming unit Granulocyte and Macrophage (CFC-GM), Colony forming unit Granulocyte, Erythrocyte, Macrophage and Megakaryocyte (CFC-MIX or 13 CFU-GEMM). Each colony is originated from a single progenitor cell, thus the number of colonies is a measure of the number of progenitors for each cell lineage in the original sample. 1.2.3.2b) BFU-E (Burst forming unit-erythroid): BFU-E colonies are usually red and they are easily visualized under the microscope (Figure 6a). Smaller BFU-E may not appear red in color but are distinguishable by their morphology. 1.2.3.2c) CFU-GM (Colony forming unit-granulocyte, macrophage): The Granulocyte/Macrophage clonogenic progenitors are common progenitors for both cell types and give rise to colonies formed by a heterogeneous population of macrophages and granulocytes. These colonies are colorless and consist of round cells (granulocytes) and oval cells (macrophages). (Figure 6b). Other myeloid colonies are CFU-G (colony forming unitgranulocyte) and CFU-M (colony forming unit-macrophage), which give rise to a homogenous population of granulocytes or macrophages respectively. 1.2.3.2d) CFC-MIX or CFC-GEMM: Colony forming unit-granulocyte, erythrocyte, macrophage and megakaryocyte are multi-lineage progenitors that give rise to erythroid cells, granulocytes, macrophages, and megakaryocytes. They are identified by the reddish color that is provided by cells containing hemoglobin (erythroid), mixed with colorless/bright cells 14 (granulocytes, macrophages and megakaryocytes) in the same colony (Figure 6c). 1.2.3.2e) CFU-Spleen: Spleen colony-forming units are a measure of undifferentiated progenitors of myeloid origin that get amplified forming spleen foci after transplantation into a lethally irradiated mouse. These hematopoietic colonies are usually counted after 8-12 days from the transplantation (Figure 6d). The number of colonies obtained provides a relative quantification for the presence of undifferentiated hematopoietic progenitors (Magli, Iscove et al. 1982). 1.2.3.2f) Short-term and Long-term Reconstitution assay (STR and LTR): The only test that demonstrates the presence of HSCs in a sample involves the hematopoietic reconstitution of a lethally irradiated mouse. The irradiation leads to the depletion of the endogenous hematopoiesis in the recipient animal and permits the hematopoietic reconstitution by the injected donor cells. After 1 month, undifferentiated progenitors give rise to all lineages, which is known as STR-HSC activity. These progenitors have a limited or inexistent self-renewal capacity and get eliminated after this one-month period. In contrast, LTR-HSCs that include pluripotent hematopoietic cells with unlimited self-renewal capacity (the actual HSCs) are also able to reconstitute all lineages and this capacity is maintained 4 months after transplantation (and even later). However, to definitively test 15 the self-renewal capacity of the HSC populations, serial transplantation assays need to be performed into secondary and tertiary recipients. a) b) c) d) Figure6. Functional hematopoietic assays. (a-c) Colonies in CFU-C assay. (a)BFU-E; (b)CFU-GM; (c)CFU-Mix. (d) Colonies in Spleen at day 11 (CFU-S11). (a-c) 40x magnification. 1.2.3.2g) Competitive transplantation assay: This assay is a modification of the conventional LTR-assay. The aim of this technique is to compare the ability and the efficiency of two different HSCs pools to reconstitute irradiated animals. So, different amounts of cells from the experimental sample are injected to an adult irradiated mouse together with a constant number of competitor cells. The read-out of the experiment is the amount and percentage of reconstitution (from the cells of interest) at 1 and 4 months after transplantation (Figure 7). 16 % of donor cells HSCs Donor/competitor mix in the recipient Donor 1: HSCs: 2 33% Donor 2: HSCs: 4 50% Donor 3: HSCs: 2 66% Donor HSC Competitor HSC Figure7: Hypothetical competitive transplantation assay. Recipients are reconstituted with different amounts of HSCs from the donor and the same amount of competitor. In the figure the donor cells have normal HSC reconstitution capacity because they reach the expected percentage. 1.2.3.2h) Limiting dilution transplantation assay: This assay is a modification of the conventional LTR-assay in which decreasing numbers of donor cells are injected into lethally irradiated mice and tested for their hematopoietic reconstitution capacity. The aim of these experiments is to check the minimum amount of a specific sample that is able to reconstitute a recipient mouse. From this information it is then possible to estimate the number of HSC in the original sample. It is also used to check the changes in HSC capacity after drug treatments. In this assay, reconstitution is determined at 1 and 4 months after transplantation (Figure 8). 17 The following strategies have been also developed to allow the engraftment of the most immature or “naïve” embryonic HSCs, which are unable to reconstitute the murine hematopoiesis in conventional HSC assays. Figure8: Hipothetical limiting dilution transplantation assay. Different amounts of HSC are treated with drugs A and B. After treatment HSCs are injected into lethally irradiated mice. In this example treatment B increase the amount of HSC. 1.2.3.2i) Newborn reconstitution: This assay takes advantage of the fetal liver microenvironment still present in newborn mice. Pregnant females are injected with busulfan before delivery to immunodeplete the pups that will be the recipients for HSC testing. At day 1-2 after birth the samples of interest are injected into the facial vein of the recipients and the percent of engraftment is measured at 1 or 4 months after injection. 18 1.2.3.2j) Explant culture: Ex-vivo culture of dissected AGM is generally used to amplify the HSC population and facilitate their detection. Specifically, the embryonic tissue is dissected and cultured in an air-liquid interphase for 1-3 days (Reviewed in Medvinsky and Dzierzak 1999) and then used for transplantation assays. 1.2.3.2k) Aggregation assay: This technique has been developed as an alternative method to amplify or generate HSCs from pre-HSCs. In this case, dissected AGMs are disrupted and specific cell populations (or the whole AGM content) are then reaggregated with other AGM cells or with stromal cells such as the newborn bone marrow-derived cell line OP9 (Taoudi, Gonneau et al. 2008; Rybtsov, Sobiesiak et al. 2011). The reaggregates are cultured as explants and after 3-4 days they can be used for transplantation experiments. 1.2.3.2l) Discrimination of cells from donors vs. recipients: All hematopoietic transplantation strategies should consider the necessity to distinguish the donor cells from the remaining hematopoietic cells of the recipient. This can be achieved using different strategies: -Male/Female: The donor and recipients have different gender. A southern blot, in situ hybridization or quantitative PCR using specific probes/primers for the Y and X chromosomes is used to measure the amount of donor and recipient blood cells. 19 -Transgen Reporters: The donor cells contain an exogenous DNA that can be detected by PCR. More recently, reporter fluorescent proteins are being used, which can be directly detected by flow cytometry. -CD45 polymorphisms: CD45 is a membrane tyrosine phosphatase that is expressed in all-hematopoietic lineages except the erythrocytes. There are two different CD45 isoforms CD45.1 and CD45.2 that are nowadays widely used for detection of engrafted cells. There are specific antibodies that detect each isoform and the percentage of engraftment can be easily determined by flow cytometry. 1.2.4. Primitive hematopoietic wave The first hematopoietic cells in the embryo are found in the YS, which is a membranous sac that in reptiles, sharks and bony fishes, but not in mammals, is responsible of the embryo nourishment. In mice but not in humans, this extra-embryonic tissue is around the embryo. An endodermal and a mesodermal cell layer form the yolk sac. Being the latter the ones that give raise to the hematopoietic cells (Figure 9). 20 Figure9. Chronology of the first hematopoietic stems/progenitors cells during mouse embryo development from E6 to E10,5. It is important to note that the time point of the appearance of hematopoietic progenitor/stem cells and the site where they are detected differ according to the experimental approach used to reveal the hematopoietic potential. The hematopoietic potential of the embryonic tissues has been tested directly on freshly isolated tissues (A) or after an in vitro culture step (B) to reveal a hematopoietic potential undetectable on fresh tissues. HSC: hematopoietic stem cells, YS: yolk sac, UA: umbilical artery, VA: vitelline artery, P-Sp: para-aortic splanchnopleura, E: embryonic day postcoitus, WT: wild-type. (Figure extracted from Boisset and Robin 2012) 21 1.2.4.1. Yolk Sac hematopoiesis The first sign of mesodermal differentiation in the mouse embryo appears at E7.25 as homogeneous aggregates of cells, called hemangioblastic cords. The hemangioblastic cord differentiates to endothelial cells, and closely associated to this endothelial cells are generated primitive nucleated erythroid cells (Barker 1968), macrophages and megakaryocytes (Palis, Robertson et al. 1999; Tober, Koniski et al. 2007). The final structures that include the endothelial and hematopoietic populations are called blood islands (Figure 10). Functional HSCs, considering those cells that are able to reconstitute irradiated mice in conventional transplantation experiments, have not been detected in the YS previous to the establishment of the blood circulation. However it is still controversial whether precursors of the definitive HSC are found in the YS, and cell-tracing experiments using an inducible cre recombinase under the control of the Runx1 promoter (Runx1tm1(cre/Esr1*)Ims; see more details of Runx1 in the next chapter) crossed with a cre-recombinase reporter (ROSA26-FLOX-STOP-FLOX-LACZ) suggested this possibility (Samokhvalov, Samokhvalova et al. 2007). In this work, when induction of cre recombinase by tamoxifen was performed at E 7,5 few peripheral blood cells were found to be labeled in the adult. Most importantly, when the induction was performed after E9.5 all hematopoetic cells were labeled, indicating that HSC generation was restricted to the embryonic period. Similarly, newborn mice have been successfully reconstituted using YS cells from E9 embryos (Yoder and Hiatt 1997; Yoder, Hiatt et al. 1997), however these transplantation experiments were 22 performed after blood circulation was established and the origin of the engrafted cells cannot be determined. On the other hand, elegant studies from Dierterlen-Lièvre using quail-chick embryo chimeras demonstrated that adult blood linages were all generated from the embryonic tissues but not from the extraembryonic YS tissue (Pardanaud, Luton et al. 1996). A few years later, Patient’s lab demonstrated that primitive and definitive hematopoietic waves arose from different blastomers in Xenopus (Ciau-Uitz, Walmsley et al. 2000). In the mouse embryo, culturing YS and Paraortic-Splacnopleura (P-Sp which is the AGM precursor) before the establishment of circulation provided additional evidence that repopulating progenitors were generated only from embryonic Para-Splactnopleura tissue (Cumano, Ferraz et al. 2001). However the identification of specific factors that are required to generate adult HSCs can provide the basis to transform YS hematopoetic cells into actual HSCs. In this sense, ectopic expression of Hox-B4, a homeotic selector gene implicated in self-renewal of definitive HSCs, can confer the long-term reconstituting potential to YS-derived cells (Kyba, Perlingeiro et al. 2002). 23 Figure10. Hematopoietic organs and main hematopoietic events during embryonic development. Hematopoiesis occurs first in the yolk sac (YS blood islands and later at the aorta-gonad-mesonephros (AGM) region, placenta and fetal liver (FL). (Figure adapted from Orkin and Zon 2008) 24 1.2.5. Definitive hematopoietic wave The name definitive refers to the generation of hematopoietic cells (HSC, HPC), which are phenotypically and functionally identical to the adult hematopoietic cells. Different embryonic and extraembryonic tissues contain cells with these characteristics after day E8.5: P-Sp/AGM, umbilical and vitelin arteries, placenta and fetal liver (Figure 9-10). 1.2.5.1. P-Sp/AGM and major arteries Para-aortic splanchnopleural (P-Sp) is a mesodermal structure that develops into the aorta, gonad and mesonephros region (AGM). The AGM is comprised between the notochord and the somatic mesoderm and extends from the umbilicus to the anterior limb bud of the embryo. The AGM region contains the dorsal aorta, genital ridges and mesonephros. Around day E10,5, clusters of cells emerge in the ventral wall of the dorsal aorta of the mouse embryo that express hematopoietic markers (Taoudi and Medvinsky 2007) (de Bruijn, Ma et al. 2002; Yokomizo and Dzierzak 2010; Robin, Ottersbach et al. 2011; Rybtsov, Sobiesiak et al. 2011). These markers include CD45, SCA-1, c-Kit and CD41. AGM intra aortic hematopoietic clusters (IAHC) cells contain the first HSC that can reconstitute adult irradiated mice from E10.5 to 12 (Muller, Medvinsky et al. 1994; Medvinsky and Dzierzak 1996; Taoudi and Medvinsky 2007). 25 Medvinsky and colleagues have recently shown that, HSCs are also found in the AGM of the human embryo (Ivanovs, Rybtsov et al. 2011). Since human embryonic development, including the accomplishment of the different hematopoietic waves is much more spaced compared with the murine development, this work definitively demonstrate that the first tissue that generates HSCs in mammals is the AGM. Zebrafish has been widely used to study the origin of HSCs, which is greatly advantageous due to the transparency of the embryos that facilitates real time imaging, but also to the conservation of the process compared with mammals (Bertrand, Chi et al. 2010). These imaging techniques using zebrafish embryos have permitted to visualize the emergence of HSCs from the aortic endothelium in vivo (Boisset, van Cappellen et al. 2010; Bertrand, Chi et al. 2010). Similar results have been obtained using live imaging from mouse AGM explants. In addition to the AGM aorta, other arterial vessels such as the vitelline and umbilical arteries also contain HSC activity (de Bruijn, Peeters et al. 2000; de Bruijn, Speck et al. 2000). In conclusion, although most evidences support that definitive HSCs emerge from aortic endothelial cells, a definitive model that explain how these cells are generated is still lacking. In this sense, Dr. Godin proposed three of the four different possibilities (Reviewed in Godin and Cumano 2002): 26 1.-HSCs are produced in the aortic floor from differentiated endothelial cells, called hemogenic endothelium by a transdifferentiation process. 2.-Multipotent precursor cells generates HSC and endothelium in the aortic floor. This multipotent progenitor could be the hemangioblast or can refer to a more undifferentiated mesodermal cell. 3.-Mesodermal multipotent precursor cell generates HSC below the aortic floor in sub-aortic patches. 4.-HSC precursors are generated in the YS and after circulation is established migrate to their niche in the aortic endothelium to develop into definitive HSC with self renewal capacity. In 2009, work from Iruela-Arispe’s lab showed that HSCs are originated from cells expressing the endothelial marker VE-cadherin (Zovein, Hofmann et al. 2008), supporting either the first or second model. In this work they use the mesenchymal genes Smooth muscle 22 alpha (SM22alpha) (early mesenchymal cells) and Myocardin (Myo) (somatic mesoderm and smooth muscle cells) to drive cre recombinase expression that will then induce the ROSA26-FLOX-STOP-FLOX-LACZ reporter. In these mouse models SM22-labeling is detected in mesenchymal cells under the aorta and also in the endothelial cells while Myo-labeling is only present in the mesenchymal cells underneath the aorta but not in the endothelial layer. Contribution to adult hematopoiesis was achieved from cells labeled by SM22-alpha-cre Lac-Z but not with Myo-cre Lac-Z, thus indicating that only early mesodermal cells that contribute to endothelium generate 27 hematopoietic progeny whereas no HSC were derived from mesenchymal cells expressing Myo. However, by staining the first layer of the dorsal aorta with Oregon green injected in the lumen of the aorta, it has been recently proposed that cells underneath the endothelial layer, which express both hematopoietic and endothelial markers (VE-cadherin and CD41), can engraft and reconstitute adult irradiated animals after specific culture conditions (reaggregation assay) (Rybtsov, Sobiesiak et al. 2011). These results, that suggest that HSCs originate from cells residing in the mesenchyme, are very provocative. However, it cannot be excluded that some cells from the endothelium can migrate to the mesenchymal layers during their maturation process, reminiscent of the migration process that occurs in zebrafish (Bertrand, Chi et al. 2010). In any case, more genetic experiments need to be performed to better understand where HSCs come from. 1.2.5.2. Arterial fate precedes HSC generation. Arterial fate is determined previous to heart beating (E8,5) and HSC generation (E10,5). Arteries are the first niche for HSC (de Bruijn, Speck et al. 2000), however it is still controversial whether arteries are required to generate these cells. Evidences that support this hypothesis include the fact that most of the mutant mice that do not generate arteries show an impaired hematopoietic program. This is the case for several Notch pathway mutants such as Notch1, RBP-Jκ and Mindbomb1 (see chapter 1.4.8.5b) (Krebs, Xue et al. 2000; Shutter, Scully et al. 2000; Duarte, 28 Hirashima et al. 2004; Krebs, Shutter et al. 2004; Burns, Traver et al. 2005) and also the silentheart zebrafish mutants (see chapter 1.3.2.2) (North, Goessling et al. 2009). In contrast, COUP-TFII deficient embryos, in which veins express arterial markers, are able to generate hematopoietic clusterlike structures (see chapter 1.4.8.5b) (You, Lin et al. 2005). In mutant embryos with affected artery-vein boundaries, such as Activin Areceptor type II-like1 (acvrl1, alk1) or endoglin (CD150), contain the clusterlike structures budding from vein-like vessels that do not express the arterial marker Ephrin-B2 (Urness, Sorensen et al. 2000; Sorensen, Brooke et al. 2003). Furthermore, using the zebrafish model, Zon’s group demonstrated that overexpressing active Notch1 in Notch-deficient embryos (mindbomb-Knockout, see chapter 1.4.6), cells expressing Runx1 or Myb (putative HSC) are found in the veins in the absence of the arterial marker Ephrin-B2 (Burns, Traver et al. 2005). However the formal proof that HSC can be generated in the absence of arteries is still missing. 1.2.5.3. Placenta and fetal liver hematopoiesis In mice, HSCs are detected almost simultaneously in the placenta and the AGM (Gekas, Dieterlen-Lievre et al. 2005; Ottersbach and Dzierzak 2005) around E11 (Figure 10) and it was hypothesized that HSCs were independently originated in the placenta and AGM. However in human embryos, it has been recently shown that the first tissue that contains HSCs is the AGM (Ivanovs, Rybtsov et al. 2011). HSCs are detected at Carnegie 29 stage 15 (day 33) in the AGM, and only after week six of gestation (day 42) HSCs are detected in the human placenta (Robin, Bollerot et al. 2009). However whether the murine placenta can autonomously generate HSC remains undefined as well as their contribution to the adult hematopoiesis. In Ncx (sodium-calcium pump1) knockout embryos, which lacks heartbeat and circulation, hematopoietic progenitors are detected in the placenta (Rhodes, Gekas et al. 2008), however the presence of functional HSCs was not investigated in these conditions. In fact, the requirement for blood circulation that activates the nitric oxide cascade was later demonstrated for HSC formation (see in chapter 1.3.2.2), invalidating the heartbeat mutants as a model to study the site of HSC generation (Adamo, Naveiras et al. 2009; North, Goessling et al. 2009). Once HSCs have been generated, they colonize the fetal liver from late E9 (Johnson and Moore 1975; Houssaint 1981; Zovein, Hofmann et al. 2008) where they are amplified (Johnson and Moore 1975; Ema and Nakauchi 2000; Kumaravelu, Hook et al. 2002; Takeuchi, Sekiguchi et al. 2002). Fetal liver represents the main reservoir of HSC from E12,5 and E18,5 (Gekas, Dieterlen-Lievre et al. 2005). After E18, bone marrow is colonized by HSC in a β1-Integrin-dependent process (Potocnik, Brakebusch et al. 2000). The HSCs that colonize the bone marrow are from embryonic origin, since cells labeled around E10,5 using cell tracing experiments are the ones that give rise to all hematopoietic progeny detected in the adult bone marrow 30 (Gothert, Gustin et al. 2005; Samokhvalov, Samokhvalova et al. 2007; Zovein, Hofmann et al. 2008) 31 1.3.REQUIREMENTS IN THE GENERATION OF HSC 1.3.1. Introduction Many efforts have been made the last years to identify key genes involved in the HSC generation process, which are summarized in this chapter. Still many questions remain unknown about the molecular mechanisms that govern this process. The aim of this thesis is to decipher some of these mechanisms. Elements involved in HSC generation have been divided into chemical and mechanical inputs and outputs, Chemical inputs refer to ligand proteins, cytokines or transcription factors while mechanical inputs refer to cell movements, tensions or cell shapes (see summary in Figure 9). 1.3.2. Mechanical inputs/outputs We can distinguish mechanical inputs that induce chemical outputs and chemical inputs that induce mechanical outputs. 32 Figure9. Mechanical and chemical inputs/outputs. Gene networks regulate all biological processes. Gene networks are activated in the cells by mechanical or chemical inputs and result in mechanical or chemical outputs, which activates the same or another gene network in the same or in another cell. 1.3.2.1. Cell budding and Rac GTPases The HSC bud from the AGM to migrate to other tissues such as FL where they are amplified. Rac is a GTPase from the Rho family involved in migration, invasion, tissue remodeling (Reviewed in Bosco, Mulloy et al. 2009). It was shown that p21-Rac1 is essential for the budding of hematopoietic clusters in the AGM. In p21-Rac1-deficient mutants, the YS hematopoiesis was not affected, while the progenitor cells in the blood circulation and in the FL were reduced (Ghiaur, Ferkowicz et al. 2008). Unfortunately, the effect on HSC in the AGM was not assayed in these mutants. 1.3.2.2. Nos and shear stress It was shown in 2009 that blood flow produce a shear stress, which is required to generate functional HSC (Adamo, Naveiras et al. 2009; North, 33 Goessling et al. 2009). These mechanisms are directly related to the effects of blood circulation in the activation of the Nitric Oxide (NO) pathway. In zebrafish, it was shown that compounds that increase blood flow enhanced the production of Runx1+/Myb+ (HSC markers) cells, whereas chemicals that decreased blood flow, reduced the number of these cells. Moreover, silent heart zebrafish embryos, that lack heartbeating and fail to establish blood circulation, had impaired HSCs formation (North, Goessling et al. 2009). In the mouse, NO synthase 3 (NOS-3; eNos) is expressed in the AGM endothelium and the hematopoietic clusters, and marks LTR-HSCs. Intrauterine NOS inhibition reduced the HSC activity and similar results were shown by using the NOS-3 knockout embryos. However, one alternative explanation for all these hematopoietic phenotypes is the absence of arterial development. In fact, the arterial marker efn-b2a is reduced in silent heart zebrafish (North, Goessling et al. 2009) and the presence of intact arteries in NOS-3 knockout embryos has not been tested. Thus, the direct relationship between circulation and HSC generation is still unknown. 1.3.3. Chemical inputs/outputs Several signaling pathways are involved in the generation of HSC: Cytokine signaling induced by Interleukine-3 (IL-3) or Stem Cell Factor (SCF), Sonic hedgehog (SHH), Bone Morphogenetic protein4 (BMP4), Wnt and the Notch Pathway. These pathways regulate the transcription of specific 34 factors such as Gata factors, Runx1, ERG, SCl/Tal, Hox-B4, Myb, which are important for the hematopoietic differentiation/commitment. 1.3.3.1. Interleukin 3 IL-3 it is a cytokine involved in adult and embryonic HSC regulation. In the adult, it has been shown that IL-3 increase HSC expansion and selfrenewal, assessed by LTR-assay (Bryder and Jacobsen 2000), however it can also decrease HSC activity (van der Loo, Slieker et al. 1995; Peters, Kittler et al. 1996; Yonemura, Ku et al. 1996). The discrepancies could be due to IL-3 dose effects and/or culture conditions. However disruption of the IL-3 gene (Lantz, Boesiger et al. 1998) or the IL-3 receptor (Nishinakamura, Nakayama et al. 1995; Robb, Drinkwater et al. 1995) in mice revealed only minor alterations in adult hematopoiesis. In 2006 E. Dzierzak and colleagues demonstrated that IL-3 is expressed in the IAHC-like structures. Using Runx1 heterozygous (which contain reduced HSCs numbers) they demonstrated that IL-3 increases HSCs but does not affect de novo HSC generation since it cannot rescue Runx1 knockout phenotype (see chapter 1.3.3.6 for Runx1 role). Also was shown that IL-3 knockout, drastically reduce HSC reconstituting potential from AGM (Robin, Ottersbach et al. 2006). Most likely the few HSCs present in IL-3 knockouts are enough to colonize BM and produce adult hematopoiesis. This possibility should be tested in limiting dilution transplantation assays. 35 1.2.3.2. Stem Cell Factor The Stem Cell Factor (SCF) is the c-Kit (tyrosine kinase receptor) ligand (Huang, Nocka et al. 1990; Williams, Eisenman et al. 1990). c-Kit is one of the first HSC markers in the embryo as described in Chapter 1.2.3 (Sanchez, Holmes et al. 1996; Yokomizo and Dzierzak 2010). Importantly, it has been shown that SCF is overexpressed in placental endothelial cells, which are a niche for HSC during embryonic development (Gekas, Dieterlen-Lievre et al. 2005). Blocking of c-Kit cascade reduces the expression of key HSC genes such as Runx1, Myb and Gata2 (Sasaki, Mizuochi et al. 2010). In contrast, c-Kit-deficient animals do not show obvious HSC defects, demonstrating that the SCF/c-KIT pathway is not required to generate or expand HSCs (Takeda, Shimizu et al. 1997). 1.3.3.3. Sonic Hedgehog Positional localization of a cell determines its exposure to a chemical signal. The preferentially ventral position of the hematopoietic clusters in the lumen of the aorta suggests that embryonic polarity affects the HSC specification. Dissecting the AGM into dorsal (including neural tube) or ventral tissues (including gut) and culturing them in explants, it was shown that HSC activity resides in ventral tissues and it decreased in the dorsal AGM regions (Peeters, Ottersbach et al. 2009). On the other hand, hedgehog (SHH) protein increased the HSC activity in AGM cultures, whereas inhibiting SHH with a blocking antibody decreased this activity. SHH and its 36 transcription factor Gli1 are expressed around the AGM region and in fact, BMP4, a downstream target, is also expressed ventrally (Dyer, Farrington et al. 2001; Durand, Robin et al. 2007). Hedgehog is a family of secreted proteins composed by: Indian, Desert and Sonic. Hedgehog proteins bind the receptor Patched and this results on the activation of Smoothened (Smo), which transduces the signal through Gli (Reviewed in Baron 2001). Indian and Smo null mice induce hemato-vascular defects in early yolk sac development, but primitive erythrocytes can still develop (Byrd et al 2002; Dyer et al 2001). Altogether indicates that Hedgehog is required for definitive but not for primitive hematopoiesis (Gering and Patient 2005). 1.3.3.4. Bone morphogenetic proteins Bone morphogenetic proteins (BMPs) belong to the TGF-Beta family of growth factors, originally discovered by their ability to induce the formation of bone and cartilage (Nakagawa, Lee et al. 2009). BMPs transduce their signals through transmembrane serin/threonine kinase receptors, which in turn, phosphorylate the intracellular effectors, Smad (Reviewed in Larsson and Karlsson 2005). BMP4 is mainly expressed in the ventral mesenchyme of the aorta and inhibition of BMP4 by gremlin reduces the number of HSCs and CFU-S11 progenitors in the AGM (Durand, Robin et al. 2007). 1.3.3.5. E-Twenty six transcription factors ERG is a protein from the family of ETS transcription factors. Erg is expressed in the hemogenic endothelium (VE-cadherin-positive/CD45- 37 negative), including hematopoietic clusters (VE-cadherin-positive/CD45positive) but not in the differentiated hematopoietic cells (VE-cadherinnegative/CD45-positive). ERG is upstream of the hematopoietic regulators Gata2 and Runx1 as demonstrated by Chromatin Immunoprecipitation. (Taoudi, Bee et al. 2011). Importantly, in embryo kimeras ERG wild type and ERG knockout it has been shown that cells from both genotypes can generate HSCs and hematopoietic cells in the embryo (E14), but after birth only ERG wild type cells are maintained in BM and Peripheral blood. 1.3.3.6. Run-related transcription factor 1 Runx1 was characterized as the key transcription factor for the establishment of definitive hematopoiesis, in the AGM (North, Gu et al. 1999; Yokomizo, Ogawa et al. 2001), in the fetal liver (Okuda, van Deursen et al. 1996; Okada, Watanabe et al. 1998) and also for the primitive hematopoiesis of the YS (Yokomizo, Ogawa et al. 2001). Using conditional Runx1 deletion in endothelial cells using VE-cadherin-cre or in hematopoietic cells using Vav1-cre, it has been shown that Runx1 is required for the endothelial to hematopoietic cell transition but not thereafter. So, once HSCs are formed, Runx1 is not required (Chen, Yokomizo et al. 2009). Downstream of Runx1 several genes have been identified as hematopoietic inducers. In Runx1 knockout AGMs at E11.5, genes such as PU.1, Flt-3, Vav1 (Okada, Watanabe et al. 1998) and IL-3 (Uchida, Zhang et al. 1997; Mao, Frank et al. 1999) are not expressed. 38 1.3.3.7. GATA transcription factors Gata2 is a member of the GATA family of transcription factors. In 1994, Orkin and colleagues demonstrated that GATA-2 is required to generate definitive HSC (Tsai, Keller et al. 1994) by generation of ES-cell chimeras (Gata2 +/+ and Gata2-/-) and analysis of the contribution to different hematopoietic tissues. In these conditions, no defects were detected in primitive erythroid cells. In a second study using Gata2 haploid mice, a gene-dosage effect was observed in the HSCs activity of the AGM but not in the YS, FL or adult bone marrow HSCs (Ling, Ottersbach et al. 2004). This data suggests that the hematopoietic defects associated with Gata2 deficiency (observed also in chimeras (Tsai, Keller et al. 1994) could be due to the HSC failure in the AGM. GATA-2 is upstream of Runx1 (Burns, Traver et al. 2005), and in the AGM, it is also a component of the gene-regulatory network composed by the transcription factors SCL/TAL-1 and Fli-1. In this triad each factor has selfactivation, and activates the expression of the other members. This process allows the maintenance of expression of all three members of the triad, a prerequisite for HSC specification (Pimanda, Ottersbach et al. 2007). In the adult, Gata2 is expressed in HSC (Suzuki, Ohneda et al. 2006) and oscillates during the cell cycle (GATA-2 is high in S phase and low in G1/S and M). Immunoprecipitation demonstrated that GATA-2 followed by immunoblot analysis was phosphorylated at a cyclin-dependent 39 kinase (Cdk)–consensus motif. Moreover, GATA-2 interacts with Cdk2/cyclin A2, and Cdk4/cyclin D1, which can phosphorylate GATA-2 in vitro, suggesting that GATA-2 phosphorylation by Cdk/cyclins could mediate cell-cycle specific “on-off” response of GATA2 that may control hematopoietic-cell proliferation (Koga, Yamaguchi et al. 2007). Previous work from our lab demonstrated that Notch/RBP-Jκ was upstream of Gata2 transcription factor in the AGM and we proposed Gata2 as a transcriptional Notch target (Robert-Moreno, Espinosa et al. 2005). Our results, however, did not formally demonstrate this possibility, mainly due to the complexity of Gata2 gene transcription regulation, which has been a matter of investigation by several groups. Gata2 have two different promoters that regulate two different transcripts in mouse (Minegishi, Ohta et al. 1998) and humans (Pan, Minegishi et al. 2000)(Figure 11). The formal demonstration that Gata2 is a downstream target of Notch is the aim of Chapter 3.2 (Results) of this thesis. Figure11. Human/Mouse Gata2 locus. Gata2 is a six-exon gene. There are two alternative first exons, IS and IG, and consequently two transcripts are generated. Both codify the same Gata2 protein. (Gray: non-codifying regions; black: codifying regions) 40 1.3.3.8. Wnt pathway Wnt proteins are secreted glycoproteins, which are involved in many developmental processes (Reviewed in Cadigan and Nusse 1997). In the absence of Wnt, β-catenin is localized in a destruction complex with APC, Axin and glycogen synthetase 3β (GSK3β) that mediates the phosphorylation of β-catenin to induce its degradation. The binding of Wnt ligands to the Frizzled receptors inhibits the β-catenin destruction complex. This process results in β-catenin stabilization, accumulation and translocation to the nucleus where it binds to LEF/TCF family of transcription factors to activate target genes. In zebrafish embryonic hematopoiesis, Wnt16 is required to generate HSCs (Clements, Kim et al. 2011) assessed with the absence of CD41, Myb and Runx1 expressing cells in the AGM region. The possibility that Wnt plays a role in mice or human embryonic hematopoiesis has been studied by our laboratory. We have shown that β-catenin is required during the generation of HSC, but not thereafter (Ruiz-Herguido, Guiu et al. 2012). In the adult, the role of Wnt in HSCs is controversial. Loss of function experiments by knocking-out Wnt3a (Luis, Weerkamp et al. 2009) or conditionally deleting β-catenin in hematopoietic cells (with VAV1-cre) (Zhao, Blum et al. 2007) showed a reduction of HSC activity. However following deletion of β-catenin (Cobas, Wilson et al. 2004) or double 41 deletion of β-catenin and γ-catenin (Jeannet, Scheller et al. 2008; Koch, Wilson et al. 2008) with Mx-cre (induced by interferon), hematopoiesis was not affected. Similarly, controversial results have been reported using enhanced Wnt activity models. It was found that retroviral transduction or Wnt3a treatments show an increase of HSC activity (Reya, Duncan et al. 2003; Willert, Brown et al. 2003; Baba, Garrett et al. 2005; Malhotra and Kincade 2009). On the other hand, Wnt activation by Knock-in active βcatenin (β-catS33Y) in Rosa-26 locus or inducing β-catenin-exon3 knockout with Mx-cre, which can not be degraded, results in the exhaustion of HSC pool followed by a failure in their capacity to repopulate when transplanted in secondary recipients (Kirstetter, Anderson et al. 2006; Scheller, Huelsken et al. 2006). Recently, Staal’s group demonstrated that β-catenin is required for HSC maintenance in the adult bone marrow but the effects are dosedependent (Luis, Naber et al. 2011). 1.3.3.9. Myb Myb is a transcription factor, which is required to generate definitive but not primitive hematopoiesis (Mucenski, McLain et al. 1991). Myb-deficient embryos die at E15 with severe hematopoietic defects in fetal liver (Mucenski, McLain et al. 1991). It has been shown that Myb is expressed in cluster-like structures in the AGM suggesting a role in HSC origin. In agreement with this, cell cultures from Myb-deficient AGMs fail to produce hematopoietic cells (Mukouyama, Chiba et al. 1999). Recently it has been shown that Myb is required to generate HSC but it is not required to 42 generate specific populations of macrophages that persist in the adult tissues such as skin (Schulz, Gomez Perdiguero et al. 2012). 1.3.3.10. Stem Cell Leukemia /T-cell acute leukemia-1 (SCL/Tal-1) SCL/Tal-1 gene encodes a basic helix-loop-helix (bH-L-H) transcription factor SCL-deficient embryos die at E9,5 due to the complete absence of hematopoiesis (Robb, Lyons et al. 1995; Shivdasani, Mayer et al. 1995), but SCL is dispensable for adult HSC engraftment and self-renewal. (Mikkola, Klintman et al. 2003; Curtis, Hall et al. 2004). On the other hand, SCL is required to generate megakaryocytes and erythrocytes linages (Hall, Curtis et al. 2003; Mikkola, Klintman et al. 2003). 1.3.3.11. HoxB4 Hox-B4 is a homeobox gene. Hox genes have a pivotal role for embryonic development in determining of segment structures and body plan. Using a YFP Knocked-in the Hox-B4 gene, it was demonstrated that Hox-B4 is expressed in emerging HSC of the AGM, but also in FL (at lower levels) and BM HSCs (Hills, Gribi et al. 2011). Importantly, ectopic expression of Hox-B4 in YS progenitors induces a switch of these cells to definitive HSC since they become able to reconstitute adult irradiated mice (Kyba, Perlingeiro et al. 2002). Overexpression of Hox-B4 expands the HSC pool in adult bone marrow (Antonchuk, Sauvageau et al. 2002) as well as in human cord blood without affecting their repopulating capacity (Buske, Feuring-Buske et al. 2002). 43 Despite all these data, loss of function of Hox-B4 in the embryo results in minor hematopoietic defects. Hox-B4 knockout mice have reduced cellularity in BM and spleen, but HSCs defects are not detected (Brun, Bjornsson et al. 2004). In double Hox-B4/Hox-B3-deficient mice, a lower repopulating capability has been reported (Bjornsson, Larsson et al. 2003), suggestive of functional compensation between different Hox homologues. 1.3.3.12. Notch pathway Notch pathway is involved in cell fate specification and tissue homeostasis in multiple systems. The aim of this work is to understand the role of Notch in the generation of HSCs. This pathway is the subject of the next chapter. 44 1.4.NOTCH PATHWAY 1.4.1. Introduction to the Notch pathway T. H. Morgans’s described in 1926 a mutant of Drosophila named Notch due to serrations on the wing margin (Figure12). 57 years later, the Drosophila Notch locus was sequenced (Artavanis-Tsakonas, Muskavitch et al. 1983; Kidd, Lockett Figure 12. Notch Drosophila wings. Left to right: average, extreme condition, nearly normal. (Extracted from Morgan 1919) et al. 1983; Artavanis-Tsakonas 1988). Later, it was also sequenced in Caenorhabditis elegans (Yochem, Weston et al. 1988; Yochem and Greenwald 1989) From these early cloning studies, a plethora of functions regulated by the Notch have been identified. A summary is provided in Table 1 (Table1) (Reviewed in Andersson, Sandberg et al. 2011). 45 Table1. Notch signaling regulates numerous developmental processes. (Adapted from Andersson, Sandberg et al. 2011). Organ Processes regulated References Tissue Brain Controls the balance between gliogenesis (Ohata, Aoki et al. 2011) and neurogenesis; stem cell maintenance; (Tanigaki and Honjo 2010) apicobasal polarity of neuroepithelial cells. Breast During pregnancy: alveolar development, (Buono, Robinson et al. maintenance of luminal cell fate, and 2006) prevention of uncontrolled basal cell proliferation. Craniofacial Palate morphogenesis: loss of Notch Jag2 (Jiang, Lan et al. 1998), structures signaling results in cleft palate, fusion of Jag2/Notch1 (Casey, Lan et the tongue with the palatal shelve and al. 2006), Dll3/Notch1 other craniofacial defects (Loomes, Stevens et al. 2007), Jag1 (Li, Krantz et al. 1997), tooth development (Mitsiadis, Regaudiat et al. 2005) Ear Defines epithelium, the presumptive determines hair sensory cell and supporting cell fates. CSL (Yamamoto, Chang et al. 2011), Jag1 (Kiernan, Xu et al. 2006) (Reviewed by Cotanche and Kaiser 2010) Esophagus Regulates esophageal epithelial (Ohashi et al., 2010) homeostasis. 46 Eye Fiber cell differentiation in the lens development. CSL/Notch1 (Jia, Lin et al. 2007; Rowan, Conley et al. 2008), Jag1 (Le, Conley et al. 2009) Heart Cardiac patterning, differentiation, ventricular valve trabeculation, cardiomyocyte development, outflow (Reviewed by MacGrogan, Nus et al. 2010) tract development. Hematopoiet ic system Required for the second wave of (Reviewed by Bigas, Robert- hematopoiesis in development; controls Moreno et al. 2010; Bigas the balance of B cell versus T-cell and Espinosa 2012) development. Intestine Controls proliferation and differentiation (Reviewed by (Heath 2010) (including absorptive fate versus secretory choices) Kidney Limbs Notch2 defines cell fate podocytes and (Cheng, Nefedova et al. proximal tubules 2007) Apical ectodermal ridge (AER) formation Notch1/Notch2(Pan, Liu et al. and 2005), Jag1 (McGlinn, van digit morphogenesis, especially regulation of apoptosis. Bueren et al. 2005), Jag2(Jiang, Lan et al. 1998) , Hairy (Notch target gene) (Vasiliauskas, Laufer et al. 2003) Liver Regulates ductal plate formation and Notch2 (Geisler, Nagl et al. intrahepatic bile duct morphogenesis in 2008; Zong, Panikkar et al. mice. 2009), Notch2/Jag1(Lozier, McCright et al. 2008), Jag1 47 (Li, Krantz et al. 1997) Muscle Promotes transition of activated satellite (Ghabrial and Krasnow 2006) cells to highly proliferative myogenic precursor cells and myoblasts; prevents myoblast differentiation into myotubes after injury Lungs Lateral inhibition between tracheal cells (Reviewed in Tsivitse 2010) prevents extra cells from assuming the leas position during tracheal branching morphogenesis. Neural crest Controls patterning of neural crest (Reviewed by Jain, precursors for the outflow tract region of Rentschler et al. 2010); the heart; regulates the transition from (Woodhoo, Alonso et al. Schwann cell precursor to Schwann cell, 2009; Schouwey and controls Schwann cell proliferation and Beermann 2008) inhibits myelination; controls melanocyte stem cell maintenance. Pancreas Specifies endocrine cell differentiation (Reviewed by Kim, Shin et al. through lateral 2010) lineage cells inhibition: endocrine inhibit endocrine differentiation of their neighboring cells; maintains pancreatic endocrine precursor cells, inhibits differentiation; terminal acinar controls cell pancreatic epithelium branching and bud size. Pituitary Regulates pituitary melanotrope growth/proliferation, specification gonadotrope differentiation. and Hes1 (Monahan, Rybak et al. 2009) (Raetzman, Ross et al. 2004), Notch2 (Raetzman et al., 48 2006) (Reviewed by Davis 2010) Placenta Controls fetal circulatory angiogenesis, system maternal development, (Reviewed by Gasperowicz and Otto 2008) spongiotrophoblast development. Prostate Required for epithelial differentiation and (Wang, Leow et al. 2006; growth; expressed by progenitors that are Wang, Shou et al. 2004; Orr required for branching morphogenesis et al., 2009) (Notch1); stromal survival [Notch2 and Delta-like 1 homolog (Dlk1)]. Sex organs and germ cells Maintenance of Leydig progenitor cells in (Tang, Brennan et al. 2008; testis; Hayashi et al., 2001; regulation controls of oocyte spermatogenesis; growth actomyosindependent via cytoplasmic streaming and oocyte cellularization. Nadarajan, Govindan et al. 2009) (Reviewed by Barsoum and Yao 2010) Skin Regulates cell proliferation, adhesion, hair follicle control or of (Reviewed by Hayashi, feather Kageyama et al. 2001) papillae differentiation and homeostasis. Spine/spinal Somite segmentation through oscillation (Reviewed byDunwoodie cord/somites of genes. 2009); (Kageyama, Niwa et al. 2010) Spleen Regulates generation of T lineage- restricted progenitors and marginal zone (MZ) B-cell development; (Reviewed by Yuan, Kousis et al. 2010) controls homeostasis of CD8– dendritic cells in the spleen 49 Stomach Thymus Acts as a switch in choice between (Matsuda, Wakamatsu et al. luminal and glandular cell fates. 2005) Thymic morphogenesis, differentiation of (Jiang, Lan et al. 1998) gamma delta lineage T-cells. Thyroid Regulates the numbers of thyrocyte and Hes1 (Carre, Rachdi et al. C-cell 2011) progenitors and regulates differentiation and endocrine function of thyrocytesand C-cells. Vasculature Regulates arteriovenous specification and (Reviewed by Gridley 2010) differentiation of endothelial cells and vascular smooth muscle cells; regulates blood vessel sprouting and branching. Notch pathway involves different proteins including the Notch receptors, the Delta and Jagged ligands, the nuclear transcription factor RBP-Jκ and other general and specific cofactors. Notch family members are highly conserved during evolution, from nematodes to mammals. Activation of the pathway is initiated when the Notch receptor physically interacts with one of its ligands present on a neighboring cell. This interaction induces several proteolytic events on the receptor that releases the active Notch fragment corresponding to the intracellular domain (NICD). After cleavage, NICD translocates to the nucleus, associates with RBP-Jκ, a DNA binding protein, and assembles a transcription complex containing the coactivator Mastermind-like protein 1 (Mam-1) to activate the transcription of downstream targets. 50 1.4.2. Notch Receptors Notch receptors are cell surface transmembrane proteins. In mammals, there are four different receptors (Notch1-4) (Figure 13). The extracellular part of the Notch receptor is formed by several epidermal growth factors (EGF)-like repeats (29-36). Repeats 11-12 are required to interact with the ligands presented by neighboring cells (Rebay, Fleming et al. 1991). Cterminal to the EGF-like repeats, there is an extracellular negative regulatory region (NRR). NRR contains the Lin12/Notch repeats (LNR) and is involved in maintaining the heterodimeric structure by disulphide bridges, thus preventing receptor activation in the absence of the ligand (Weng, Ferrando et al. 2004). Different proteolytic events regulate Notch signaling. Notch receptors are first cleaved by furin-like convertases in the Golgi at site 1 (S1) located in the heterodimerization domain (Figure13-14). This proteolytic event converts Notch into a heterodimeric receptor composed by Notch extracellular domain (NECD) and Notch transmembrane and intracellular domain (NTMIC). Notch heterodimeric receptors remain associated by noncovalent interactions (disulphide bridges). RAM domain is at C-terminal of the transmembrane domain and it is required to interact with the Notch nuclear partner RBP-Jκ. The Nuclear localization signals (NLS) and the ankyrin repeats (ANK domain) are the functional core of the Notch receptor. 51 At the C-terminal end, the OPA/PEST domain contains degradation signals that regulate protein stability (Fryer, Lamar et al. 2002; Thompson, Buonamici et al. 2007). 1.4.3. Notch receptor modifiers: Rumi, Pofut and fringe Figure13.Notch receptors and ligands. (A) Notch receptors contain multiple extracellular EGF-like repeats. Drosophila Notch (dNotch) and the four mammalian Notch paralogs (mNotch1–4) differ in the number of repeats (29–36). EGF repeats are followed by the negative regulatory region (NRR), which is composed of three cysteine-rich Lin12-Notch repeats (LNR-A, -B, and -C) and a heterodimerization domain (HD). Notch also contains a transmembrane domain (TMD), a RAM (RBPjκ association module) domain, nuclear localization sequences (NLSs), a seven ankyrin repeats (ANK) domain, and a transactivation domain (TAD) that harbors conserved proline/glutamic acid/serine/threonine-rich motifs (PEST). Furin-like convertases clave notch at site 1 (S1), which converts the Notch polypeptide into an NECD-NTMIC (Notch extracellular domain-Notch transmembrane and intracellular domain) heterodimer that is held together by noncovalent interactions between the N- and Cterminal halves of the heterodimerization domain. B) Known ligands and putative ligands of Notch receptors can be divided into several groups on the basis of their domain composition. Classical DSL ligands contain the DSL (Delta/Serrate/LAG-2), DOS (Delta and OSM-11-like proteins), and EGF (epidermal growth factor) motifs (Adapted from Kopan and Ilagan 2009). 52 Notch receptors are modified post-translationally by different enzymes in the Golgi apparatus and endoplasmatic reticulum (ER) before it reaches the cytoplasmic membrane. Pofut1 (O-Fucosil-transferase), Fringe proteins (Beta1,3-GlcNAc-transferase) and Rumi (O-glucosyl-transferase) are known Notch modifiers. O-Fucose-, O-Glucose- and Beta1,3-GlcNAcmodifications occur at specific specific sites of the extracellular domain (see figure 14). Pofut1 and Rumi modifications are produced in the ER while Fringe works in the Golgi. Rumi adds glucose moieties to EGF repeats. Notch receptor without O-Glucose modifications, can bind the ligand but cannot be activated (Tan, Xu et al. 2009). O-Glucose and O-fucose can be elongated by the sequential action of other glycosyltransferases to yield, respectively, a trisaccharide or tetrasaccharide structure. First step is OFucose addition to EGF repeats, produced by POFUT1. Rumi adds OGlucose in another EGF repeats. After this, Fringe adds to the O-fucose GlcNac moieties. In the mouse embryo, it has been shown that Rumi deficiency is lethal at E9.5 and the mutants display a stronger phenotype compared with other Notch mutants, which suggests that Rumi plays a role in the glycosilation of other proteins (Fernandez-Valdivia, Takeuchi et al. 2011). 53 Figure14. Notch receptor modifications (a) The extracellular domain (ECD) of mouse Notch1, EGF repeats containing the consensus sequence for O-glucosylation, Ofucosylation, or both, are filled with blue, red, or blue and red hatched lines, respectively. Mapped O-fucose and O-glucose glycans are indicated on each EGF repeat. (lightly shaded red triangles indicate O-fucose sites not yet confirmed). Symbols are based on Consortium for Functional Glycomics guidelines: glucose, blue circle; xylose, orange star; fucose, red triangle; GlcNAc, blue square; Galactose, yellow circle; sialic acid, purple diamond (Adapted from Rana and Haltiwanger 2011). Fringe Glycosiltransferases (Radical, Lunatic, manic) are responsible for glycosylation of the receptor and modulating ligand specificity. Fringe modifications favors Notch interactions with Delta-type ligands and interfere with Jagged-type ligand binding (Yang, Nichols et al. 2005; Benedito, Roca 54 et al. 2009; Ilagan, Lim et al. 2011). In the embryo it has been shown that Lunatic Fringe is required to generate somite boundaries (Serth, SchusterGossler et al. 2003). In adult hematopoiesis, Fringe is required to generate T cells (Reviewed in Zuniga-Pflucker 2004; Yuan, Kousis et al. 2010). Pofut1 and the O-fucosylation of Notch is essential for Notch signaling and Notch-ligand interactions (Okajima and Irvine 2002; Okamura and Saga 2008). Pofut1 Knockouts die at E9.5 and at this stage they exhibit severe defects in somitogenesis, vascuologenesis, cardiogenesis and neurogenesis at similar extend than RBP-Jκ knockout (Shi and Stanley 2003). To elucidate the role of Pofut1 in adult hematopoiesis conditional knockout were generated (Mx-cre, interferon inducible cre-ERT was used). In these mutants, a reduction in T lymphopoiesis and in the marginal-zone B cell production was observed, in addition to myeloid hyperplasia (Yao, Huang et al. 2011). 1.4.4. Notch Ligands Binding of Notch to one of the ligands triggers the signaling. In the mouse there are five known ligands, Jagged1-2 (JAG-1 or -2) and Delta-like 1-3-4 (Delta1,2 or 3) (Figure 13). Notch ligands are transmembrane proteins that contain: DSL (Delta, Serrate, Lag2) domain, and EGF-like repeats and DOS EGF-like repeats (Delta and OSM-11-like proteins; (Komatsu, Chao et al. 2008) are also present in some ligands (Figure 13). DSL and DOS domains are involved in the receptor binding, however it has been shown 55 that peptides containing DSL domain are sufficient to activate Notch in some systems (Shimizu, Chiba et al. 2000; Shimizu, Chiba et al. 2000). The cysteine-rich domain, which is at C-terminal of the DSL domain in the extracellular region, is exclusive of Jagged-type ligands. Many functions have been already associated to specific Notch ligands, which are included in Table 1, and are explained, with more detail in Chapters 1.8.8.4 and 1.8.8.5. 1.4.5. Receptor-ligand binding and Notch activation After ligand binding, Notch receptors suffer two proteolytic events, which results in the release of NICD (De Strooper, Saftig et al. 1998) that translocates to the nucleus where activates target genes (Figure 15). The cleaveage in site 2 (S2) takes place 13 amino acids N-terminal of the transmembrane domain. S2 cleavage is mediated by ADAM metalloproteases called TACE (Tumor necrosis factor alpha converting enzyme) in vertebrates. The cleavage generates an intermediate product called NEXT (from Notch Extracellular Truncation). Then a multiprotein complex with γ-secretase activity (Haass and Steiner 2002; Edbauer, Winkler et al. 2003) and composed by presenilin (Haass and Steiner 2002), nicastrin (Yu, Nishimura et al. 2000), Aph1 (Goutte, Tsunozaki et al. 2002; Gu, Chen et al. 2003) and Pen-2 (Francis, McGrath et al. 2002; Steiner, Winkler et al. 2002) in a 1:1:1:1 stoichiometry (Sato, Diehl et al. 2007) produces a dual cleavage of the Notch molecule (Okochi, Steiner et al. 56 2002). One proteolytic event occurs in site 3(S3) of NEXT, close to the inner plasma membrane (Okochi, Steiner et al. 2002). The second cleavage occurs near the transmembrane domain at site 4 (S4) (Okochi, Steiner et al. 2002). It is important to know that only after γ-secretase cleavage, NICD is released and translocates to the nucleus. Thus, pharmacological inhibition of γ-secretase activity results in notch signaling blockage (Zhang, Nadeau et al. 2000). Currently γ-secretase inhibitors such as DAPT, L685,458 and Compound E are widely used in the laboratory for experimental research (Ilagan, Lim et al. 2011), but also similar compounds are used in clinical trials for leukemia or cancer treatment (Reviewed in Garber 2007). 1.4.6. Endocytosis in Notch activation Endocytosis is an important process during Notch activation. It was shown that endocytosis of the DSL ligands bound to extracellular domain of notch is required to produce the S2 cleavage (Nichols, Miyamoto et al. 2007). Endocytosis is triggered by monoubiquitination, which is mediated by the E3 ubiquitin ligases Neuralized (Deblandre, Lai et al. 2001; Lai, Deblandre et al. 2001; Pavlopoulos, Pitsouli et al. 2001) and Mindbomb1 (Itoh, Kim et al. 2003). Epsins are conserved multidomain proteins that interact with core components of the endocytic machinery and recognize poly-ubiquitinated proteins (Clathrin, AP2, PIP2) as well as with mono-ubiquitinated ones (Chen, Fre et al. 1998; De Camilli, Chen et al. 2002; Polo, Sigismund et al. 57 2002). Ligand endocytosis is required for proteolytic Notch activation. Selective internalization of the NECD by ligand expressing cells is called transendocytosis (Parks, Klueg et al. 2000; Nichols, Miyamoto et al. 2007). Recently it has been shown that ligand endocytosis exerts a pulling-force to the associated Notch receptor, required to activate the signaling. The molecular mechanism is based in clathrin-mediated endocytosis (CME) requiring epsin adaptors and actin to pull on Notch (Meloty-Kapella, Shergill et al. 2012). Notch endocytosis in the receptor-expressing cell is also important for Notch signaling. The receptors that do not bind the ligand are constitutively endocytosed and recycled to the cell surface (McGill, Dho et al. 2009) or degraded in the lysosome (Jehn, Dittert et al. 2002) in an ubiquitination dependent process. Numb is a Notch negative regulator involved in its endocytosis (McGill, Dho et al. 2009). In fact Numb behaves as scaffold that links α-adaptin, component of AP-2 complex required to generate clathrin vesicles, with Notch (Berdnik, Torok et al. 2002). Entry into endosomal vesicles requires ubiquitination of transmembrane proteins. It has been shown that Numb promotes Notch ubiquitination (McGill, Dho et al. 2009). Another Notch regulator is Musashi, which inhibits Numb translation through binding to its 3’UTR (Imai, Tokunaga et al. 2001; Kanemura, Mori et al. 2001), resulting in an increase of Notch activation. Other ubiquitin ligases involved in Notch signaling are: Itch, an E3 ubiquitin ligase that targets Notch to degradation (Cornell, Evans et al. 1999) and 58 (S3)(S4) Figure15. Notch pathway. Notch is O-fucosylated and O-glucosylated in the ER (i), and elongated by additional glycosyltransferases in the Golgi (ii). Notch is also cleaved by a furin-like convertase in the Golgi (S1 cleavage, iii), generating the mature heterodimeric receptor that is subsequently expressed at the cell surface. Following interaction with a DSL (Delta, Serrate, Lag2) ligand presented on an opposing cell, Notch ECD undergoes a conformational change exposing the protease site for an ADAM protease (Kuzbanian in Drosophila) at the membrane interface (S2 cleavage, iv), and is subsequently cleaved by gamma-secretase just inside the cell membrane (S3 cleavage and S4, v), causing Notch Intracellular Domain (NICD) release. Upon S3-S4 cleavage, the NICD is free to translocate to the nucleus (vi), bind to a member of the CSL (CBF/SuH/LAG-1) family, and Mastermind-like (MAML)family cofactors to actívate Notch target genes. (Adapted from Rana and Haltiwanger 2011) Deltex, a protein containing a RING-H2 finger motif often found in E3ubiquitin ligase proteins (Takeyama, Aguiar et al. 2003), which antagonizes the negative effects of Itch, inducing a shift from degradation to recycling 59 (Matsuno, Diederich et al. 1995; Hori, Fostier et al. 2004). Thus, Deltex is a positive regulator of Notch signaling (Xu and Artavanis-Tsakonas 1990). Surprisingly, Deltex1-null mice are viable and developmental defects are not detected (Storck, Delbos et al. 2005). 1.4.7. Transcription of Notch target genes In the absence of Notch activation, it has been shown that in Drosophila Su(H) (RBP-J in mouse) actively represses its target promoters by interacting with the correpressors Groucho and CtBP (Nagel, Krejci et al. 2005). Similarly, mammalian RBP-Jκ interacts with a corepressor complex containing SMRT (silencing mediator of retinoid and thyroid hormone receptors) and the histone deacetylase HDAC-1 in the absence of Notch signaling (Kao, Ordentlich et al. 1998). After γ-secretase cleavage, NICD translocates to the nucleus, where it forms a complex with RBP-Jκ and displaces the co-repressor proteins bound to RBP-Jκ such as SMRT, CtBP and Groucho (Figure16) (Morel, Lecourtois et al. 2001; Nagel, Krejci et al. 2005). Conversely, the nuclear NICD and RBP-Jκ complex interacts with the coactivator Mastermind that enhances target gene expression by recruiting Histone acetiyl transferases co-activators, such as p300, and remodeling enzymes. This correlates with the presence of active chromatin marks such as H3K9ac or H3K4me. On the other hand, NICD recruitment antagonizes with the polycom repressive complex2 (PCR2) and correlates with the removal of the H3K27 repressive marks (Ntziachristos, Tsirigos et 60 al. 2012). Other proteins are recruited to this complex such as SKIP in a process still not completely understood (Zhou, Fujimuro et al. 2000; Zhou and Hayward 2001). The best-characterized and conserved Notch target genes are the Hairy Enhancer of Split (Hes) and Hes Related genes (Hrt). Figure16. Notch activates target genes in the nucleous. (Adapted from Lubman, Korolev et al. 2004)) In the abcence of NICD, RBP-J is bound to the DNA with a corepressor complex. (RBP-J/SMRT/CtBP/Groucho). In the nucleus NICD displaces the corepressor complex in a manner that is only partially understood and interacts with RBP-J. First the Notch/RBP-J interface recruits Mastermind, then the triprotein complex recruits histone acetylases (p300, others). The H3K27m3 repressive mark is removed and activation marks (H3K4m and H3K9ac) are added to histon H3. Then RNA polymerase II is recruited, resulting in activation of of Hes/E(spl) and other downstream targets of Notch. NICD ubiquitination and subsequent degradation by the proteasome is thought to terminate the activation phase, permitting reassembly of the repressor complex and the addition of the repressive mark H3K27m3 to the histon H3 tail by PCR2 . Abbreviations: SMRT, silencing mediator of retinoid and thyroid hormone receptors;; HDAC, histone deacetlyase; HAT, histone acetylase; Gro:Groucho. 61 There are seven Hes genes (hes1-7) in mammals, based on homology, however only Hes1(Jarriault, Brou et al. 1995), Hes5 (Ohtsuka, Ishibashi et al. 1999) and Hes7 (Hirata, Ohtsuka et al. 2000) are canonical Notch targets. Other Notch target genes are also Hrt (1-2) (Iso, Sartorelli et al. 2001; Iso, Sartorelli et al. 2001; Iso, Chung et al. 2002). 1.4.8. Role of Notch 1.4.8.1. Cell fate determination. Notch governs many biological processes such as neurogenesis, inner ear development, limb-bud formation, intestinal homeostasis, artery-vein formation, hair follicle and T cell differentiation, among others (see Table 1). In general, Notch promotes cell diversity by either maintaining the undifferentiated state or inducing differentiation through one lineage. Two different mechanisms have been proposed to explain how Notch regulates cell diversity: lateral inhibition and lateral induction. Both models are based in cell-cell interactions. 1.4.8.2. Lateral inhibition The lateral inhibition model refers to a cell-cell interaction in which one cell inhibits the differentiation of its neighboring cell into a specific lineage. The initial situation involves a population of equivalent cells that express similar levels of Notch receptor and ligand, however stochastic/unknown events result in the upregulation of the ligand in one of these cells (sending cell) 62 creating a directional response on the neighboring cells and activating the receptor (receiving cell). As a result of a negative feedback loop, the cell that activates Notch downregulates the ligand, further amplifying the process. This results in the acquisition of distinct fates from a homogenous population, thus generating a salt and pepper mosaic (Figure 17a). One classical example of lateral inhibition occurs during the neuro-epidermal differentiation process in Drosophila. Notch activation induces the expression of the Enhancer of split Complex, which represses expression of proneural genes leading to the maintenance of the epidermal fate in the majority of the cell populations, whereas the sending cell (that does not activate Notch) differenciates into the neural program (Parks, Huppert et al. 1997). In mammals, Notch signaling also represses neurogenesis, intestinal mucosecretory differentiation and myogenesis. One general mechanism by which Notch inhibits specific transcription factors is through the activation of Hes repressors (Figure17) (Jarriault, Brou et al. 1995; de la Pompa, Wakeham et al. 1997; Kim and Shivdasani 2011). 1.4.8.3. Lateral induction In the lateral induction model, Notch mediates a unidirectional signaling between the sending cells and the adjacent cell compartment. In Drosophila, lateral induction occurs during the formation of the R7 photoreceptor from the R8 cell during eye development and also in the formation of the vulval precursor cells from the anchor cell in the C. elegans gonad (figure 17b) (Reviewed in Artavanis-Tsakonas, Matsuno et al. 1995). 63 In mammals, somite formation is also an example of lateral induction. (Conlon, Reaume et al. 1995; Ferjentsik, Hayashi et al. 2009). Figure 17. Lateral inhibition and lateral induction (A) Lateral inhibition. Among a field of initially equivalent wild-type (Notch+) cells, random fluctuations in a putative signaling component are amplified cell-autonomously such that differences arise between adjacent cells. These differences are enhanced by the production of a locally acting inhibitory signal, and result in a spaced pattern composed of two distinct cell types. In Notch- cells that lack the proposed receptor for the inhibitory signal (right panel), all cells adopt the same fate. (B) Lateral induction. Cell fates are determined by signals transmitted between nonequivalent cells, with the final pattern dependent on the timing, spatial arrangement, and specificity of the inductive interactions. In Notch- cells (right panel), the absence of Notch function might prevent inductive interactions from occurring (columns at left), or permit aberrant inductive events to occur (columns at right). (Figure adapted from Artavanis-Tsakonas, Matsuno et al. 1995) 64 1.4.8.4. Notch and stem cells As mentioned before Notch is involved in the maintenance of undifferentiated cellular compartment in different systems such as neurogenesis, intestine, hair follicle and hematopoietic system. 1.4.8.4.a) Neurogenesis: In neurogenesis, Notch signaling induces the activation of the transcriptional repressors Hes1 and Hes5, which mediate most of the Notch-dependent neurogenic functions by repressing proneural genes (Ohtsuka, Ishibashi et al. 1999). Activation of Notch signaling leads to maintenance of the neural stem cell population, whereas its inactivation induces neuronal differentiation and exhaustion of the neural stem cell population (Reviewed in Bertrand, Castro et al. 2002; Ross, Greenberg et al. 2003; Kageyama, Ohtsuka et al. 2008) 1.4.8.4.b) Intestine: Notch is involved in the regulation of intestinal homeostasis. In the RBP-Jκ knockout mice, cells from the small intestinal crypts stop proliferation and differentiate to mucosecretory cells (Jarriault, Brou et al. 1995; van Es, van Gijn et al. 2005; Kim, Kim et al. 2011). On the other hand, expression of the constitutively active form of Notch (NICD) in the intestinal epithelium dramatically impairs cell differentiation and increases the number of cycling cells (Fre, Huyghe et al. 2005). The key Notch ligands in the maintenance of intestinal homeostasis are Delta1 and Delta4 (Pellegrinet, Rodilla et al. 2011). Double Delta1 and Delta4 mutants display a clear intestinal phenotype that is characterized by the loss of the 65 intestinal stem cells (ISC), as shown by the absence of the ISC markers Lgr5, Olfm4, Ascl2. Recently, it was shown that Hes mediates most of the differentiation effects to the mucosecretory lineage downstream of Notch. In this sense, Hes1, Hes3, Hes5 triple mutant mice show similar defects than RBP-Jκ knockouts, including reduced cell proliferation, increased secretory cell differentiation and altered intestinal structures (Ueo, Imayoshi et al. 2012). However the Lgr5+ cells, which are proliferating intestinal stem cells (Barker, van Es et al. 2007; Sato, Vries et al. 2009; Ueo, Imayoshi et al. 2012)are maintained in these mutant animals. Whether other types of intestinal stem cells such as BMI+ cells, likely quiescent intestinal stem cells in +4 position of the crypt (Sangiorgi and Capecchi 2008; Tian, Biehs et al. 2011), are affected remains unknown. Thus, Hes genes, downstream of Notch, can explain the intestinal differentiation phenotype (Fre, Hannezo et al. 2011), however other Notch target genes should be responsible for the effect of Notch intervention on ISC, which is currently under investigation. 1.4.8.4.c) Hair follicle: Epidermal stem cells reside in the upper third of the follicle, in a structure called bulge (Reviewed Cotsarelis, Kaur et al. 1999)). The bulge stem cells are able to generate the epidermal and hair follicle cells (Cotsarelis, Kaur et al. 1999). Disruption of RBP-Jκ induces differentiation of hair follicular stem cells into epidermal cells and 66 suppresses hair formation. Consequently, Notch signaling negatively regulates stem cell differentiation in the bulge (Yamamoto, Tanigaki et al. 2003). Jagged1 is the key ligand that activates Notch pathway in the hair follicle. In this system Jagged1 is a downstream target of β-Catenin in this system (Estrach, Ambler et al. 2006; Rodilla, Villanueva et al. 2009) 1.4.8.4.d) Adult hematopoiesis: The role of Notch in the adult bone marrow HSC is controversial. The effects of activating Notch in vitro using different Notch ligands have been tested using hematopoietic progenitors from different sources, such as umbilical cord, linneage-negative/CD34positive/CD38-positive or bone marrow Lineage-negative/SCA-1-positive/cKit-positive (LSK) or CD133-positive (Karanu, Murdoch et al. 2001; Suzuki, Yokoyama et al. 2006); (Varnum-Finney, Purton et al. 1998). In all this experiments, Notch activation resulted in an increase in the pool of undifferentiated progenitors (assessed as CD34-positive/CD38-negative and LTR-assay in Delta1 experiments, CD34-positive/CD38-negative in Delta4 experiments, and CFUs in Jagged experiments). Similar effects were observed by overexpressing Hes1 in LSK derived cells from bone marrow, as determined by LTR-assays (Kunisato, Chiba et al. 2003). These results suggest that Notch has a role in maintaining the hematopoietic stem or progenitor pool of cells. However this possibility has not been confirmed in the in vivo system using loss of function experiments. In this sense, conditional deletion of Notch1, Jagged1 or RBP-Jκ by the interpheroninducible Mx-Cre has no detectable effect on the hematopoietic stem cell 67 pool (Mancini, Mantei et al. 2005). A similar negative result was obtained by expressing the dominant negative of the Notch pathway coactivator Mam-1 (Maillard, Koch et al. 2008). Together, these data unequivocally demonstrates that Notch is dispensable for HSC maintenance in the adult bone marrow under physiological conditions, but suggest that, its overactivation in stress/ex-vivo conditions could increase HSC self-renewal. In fact, in 2011, I.D.Bernstein’s lab and colleagues, showed that Notch1 and Notch2 are dispensable in HSC homeostasis, but Notch2 is able to inhibit myeloid differentiation and enhance the generation of both short(Multipotent progenitors) and long-term (HSC) repopulating cells during stressed hematopoiesis (induced by 5-fluorouracil) (Varnum-Finney, Halasz et al. 2011). 1.4.8.4.e) Embryonic hematopoiesis: HSCs are originated during embryonic life (see chapter 2.4). By analyzing Notch1- and RBP-Jκdeficient mouse embryos or Mindbomb-1-deficient zebrafish embryos, it was shown that this process depends on Notch (Kumano, Chiba et al. 2003; Burns, Traver et al. 2005; Robert-Moreno, Espinosa et al. 2005). In general, Notch pathway mutants lack definitive hematopoiesis and have impaired expression of key transcription factors such as SCL, Runx1 and Gata2, however they contain normal yolk sac hematopoietic cells and progenitors (Kumano 2003 , Burns 2005, Robert-Moreno 2005). However, and as previously discussed, Notch activity is essential for the regulation of arterial formation (see in chapter 1.2.5.2 and 1.4.8.5), which could be 68 affecting the process of definitive hematopoiesis. Thus, the question on whether Notch had a direct or indirect role in the generation of the HSCs had not resolved at the beginning of this work. The analysis of the Jagged1 mutants included in this thesis was crucial to solve this question (RobertMoreno, Guiu et al. 2008). Different ligands and receptors are expressed in the AGM region such as Delta4, Jagged2 and Jagged1 ligands, Notch1 and Notch4 receptors. The Notch target genes Hrt1, Hrt2 and Hes1 are also expressed (RobertMoreno, Espinosa et al. 2005). Before the initiation of this thesis, Runx1 and Gata2 had been proposed to be downstream of Notch in zebrafish and in the mouse embryos (Kumano, Chiba et al. 2003; Burns, Traver et al. 2005; Robert-Moreno, Espinosa et al. 2005). In fact, Runx1 expression rescues the hematopoietic potential of Notch1-deficient AGM cells (Burns, Traver et al. 2005). Also our group had shown that Notch1 was recruited in the Gata2 promoter (Robert-Moreno, Espinosa et al. 2005). Our current work has extended towards the identification and demonstration of specific Notch-target genes in the AGM, which are specifically involved in hematopoietic program. 1.4.8.5. Notch and differentiated specialized cells 1.4.8.5.a) T lymphocytes generation: During T cell development a pluripotent T-cell commited migrates from the bone marrow to the thymus, 69 where mature T cells are generated. Notch1 and Delta4 are required for this process. The first definitive evidence came from mice with an interferon-inducible deletion of Notch1 that contained large numbers of immature B cells in the thymus but lacked T cell precursors (Wilson, MacDonald et al. 2001; Visan, Tan et al. 2006). Similarly, conditional deletion of Hes1 using Mx-cre identified this repressor as one of the important Notch downstream effectors in T-cells (Shibata, Yamada et al. 2011). Another breakthrough in the field was the generation of mature Tcells in vitro by exposing bone marrow progenitors to Delta expressing cells. Although all initial studies were performed with Delta1, it was later demonstrated by genetic deletion of Delta4 in the thymic stoma, that this is, in fact, the physiological ligand that regulates Notch activity in this system (Mohtashami, Shah et al. 2010). Moreover, constitutive activation of Notch1 in bone marrow HSC/P results in the ectopic development of immature T cells. When transplanted into irradiated mice, these N1IC-expressing cells rapidly generate leukemic clones that are present in peripheral blood in less than one month. In fact, this is a widely used animal model for studying Tacute lymphoblastic leukemia. Moreover, Notch mutations are present in most T-ALL patients (Lee, Kumano et al. 2005; Mansour, Linch et al. 2006). 1.4.8.5.b) Arteries vs. veins: Arteries and veins are vessels with different characteristics that are formed during the early stages of development (before heart beating at 8.5 and before HSC emergence). In vertebrates 70 has been widely characterized the role of Notch in the arterial fate determination. Notch pathway members (Delta4, Notch1, RBP-Jκ, Mindbomb1) are required to generate arteries (Krebs, Xue et al. 2000; Shutter, Scully et al. 2000; Duarte, Hirashima et al. 2004; Krebs, Shutter et al. 2004; Burns, Traver et al. 2005). Delta1 mutants have downregulation of arterial markers SMA, EphrinB2 (Efn-B2) and Neuropilin-1 (Sorensen, Adams et al. 2009). Gridley’s lab shown that Jagged1 mutants have vascular malformations (Xue, Gao et al. 1999), but unfortunately arterial markers where not analyzed in this paper, and it is characterized in this thesis work. Arteries are the HSC niche during mouse development. In fact HSCs emerge from them (see 1.2.5.1 and 1.2.5.2). The link between arterial specification and HSCs generation (explained in chapter 1.2.5.2.) has complicated the study of the Notch functions in hematopoietis independently of the effects of Notch deficiency on arterial development (Reviewed in Bigas and Espinosa 2012). The fact that several Notch-family mutants are included in the group of artery deficient embryos clearly demonstrate the role for Notch in the induction of the arterial program but precludes the use of these mutants to provide definitive evidence for a hematopoietic function of Notch. One of the aims of this thesis is to identify Notch pathway members, which helps to distinguish specific Notch-arterial and Notch-hematopoietic programs; this key issue is described in the results and discussion chapters. Notch acts downstream of VEGF (Figure18) that activates the arterial 71 marker neuropilin-1 (Soker, Takashima et al. 1998; Gluzman-Poltorak, Cohen et al. 2000; Gluzman-Poltorak, Cohen et al. 2001; Lawson, Vogel et al. 2002). Notch pathway induces the expression of Efn-B2 and the Hes related genes Hrt1 and Hrt2. Hrt1 and Hrt2 are repressors of EphB4, which is a vein marker (Efn/Eph signaling mediates repulsion) (You, Lin et al. 2005). In fact Hrt1 and Hrt2 double Knockouts do not express arterial markers such as Efn-B2, CD44, and Neuropilin-1(Fischer, Schumacher et al. 2004). In veins the transcriptional repressor COUP-TFII inhibits Neuropilin-1(You, Lin et al. 2005). In this context Notch pathway is not active and EphB4 is overexpressed. 72 73 74 2.OBJECTIVES: 75 76 2.OBJECTIVES: In the embryonic aorta, loss-of-function mutations of the Notch pathway affect the development of the arterial program and the hematopoietic program (as detailed in chapter 1.4.8.4.d and 1.4.8.5.b). The aim of this thesis is to determine whether Notch has a direct role in hematopoietic program during the generation of the first HSCs as well as identify key downstream targets. The following specific objectives were proposed for this thesis: -Identify specific Notch ligands that parcicipate in the regulation of the hematopoietic program in the AGM. -Characterize Notch target genes involved in the generation of HSCs. 77 78 CHAPTER 3. RESULTS (PUBLICATIONS) 79 80 3.1. Impaired embryonic hematopoiesis yet normal arterial development in the absence of the Notch ligand Jagged1. This article has been published in “The EMBO Journal”, June 2008 Robert-‐Moreno, A., J. Guiu, et al. (2008). "Impaired embryonic haematopoiesis yet normal arterial development in the absence of the Notch ligand Jagged1." The EMBO journal 27(13): 1886-‐1895. 81 Robert-Moreno A, Guiu J, Ruiz-Herguido C, Lopez ME, Ingles-Esteve J, Riera L, et al. Impaired embryonic haematopoiesis yet normal arterial development in the absence of the Notch ligand Jagged1. Supplementary data. EMBO J. 2008 Jul 9;27(13):1886-1895. 120 3.2. Hes repressors are essential regulators of Hematopoietic Stem Cell Development downstream of Notch signaling. This article is under second revision in Journal Experimental Medicine. 121 Hes repressors are essential regulators of Hematopoietic Stem Cell Development downstream of Notch signaling. Jordi Guiu1, Ritsuko Shimizu2, Teresa D’Altri1, Stuart T. Fraser3&, Jun Hatakeyama5, Emery H. Bresnick4, Ryoichiro Kageyama5, Elaine Dzierzak6, Masayuki Yamamoto7, Lluis Espinosa1 and Anna Bigas1. 1 Program in Cancer Research. IMIM-Hospital del Mar, Parc de Recerca Biomèdica de Barcelona, Barcelona, Spain. 2 Molecular Hematology, Tohoku University School of Medicine, Sendai, Japan. 3 Department of Molecular Genetics, Kyoto University, Kyoto, Japan 4 Department of Cell and Regenerative Biology. University of Wisconsin School of Medicine and Public Health, Madison, WI 53705 5 Institute for Virus Research, Kyoto University, Kyoto, Japan 6 Department of Cell Biology and Genetics. Erasmus University, Rotterdam, The Netherlands. 7 Medical Biochemistry, Tohoku University School of Medicine, Sendai, Japan. & Disciplines of Physiology, Anatomy and Histology, School of Medical Sciences, University of Sydney, Camperdown, Australia. Running title: HES regulates HSC development. 122 Corresponding author: Anna Bigas IMIM-Hospital del Mar Dr. Aiguader 88 08003 Barcelona Phone: 34 933160440 Fax: 34 9321600410 e-mail:[email protected] & Present address: University of Sydney, Sydney, Australia. 123 3.2.1. Abstract Although previous in vivo studies have identified Notch as a key regulator of Hematopoietic Stem Cell (HSC) development in the vertebrate embryo, the underlying downstream mechanisms remain unknown. The Notch target Hes1 is widely expressed in the Aortic endothelium and hematopoietic clusters, though Hes1-deficient mice show no overt hematopoietic abnormalities. We now demonstrate that Hes is required for the development of HSC in the mouse embryo, a function previously undetected due to de novo expression of Hes5 in the Aorta/Gonad/Mesonephros region of Hes1 mutants that results in functional compensation. By analysis of double genetic mutants, we found that embryos deficient for Hes1 and Hes5 alleles contain an intact arterial program with overproduction of non-functional hematopoietic progenitors and total absence of HSC activity. These alterations were associated with increased expression of the hematopoietic regulators Runx1, c-myb and the previously identified Notch-target Gata2. By analyzing the Gata2 locus, we have discovered several functional RBP-Jκ and HES-binding sites that specifically recruit Notch/RBP-Jκ and HES-1 proteins in hematopoietic cells. Mutation of RBP-Jκ-binding sites results in loss of Gata2 reporter expression in transgenic embryos whereas mutation of HES-binding sites leads to specific Gata2 upregulation in the aortic hematopoietic cluster cells. Together our studies reveal that Notch activation in the embryonic aorta triggers both Gata2 and Hes1 transcription, next HES-1 protein 124 represses Gata2, thus creating an Incoherent Feed-Forward Loop required to restrict Gata2 expression in the emerging HSCs. 125 3.2.2. Introduction Hematopoietic Stem Cells (HSCs) originate during embryonic life in association with arterial vessels including the aorta in the Aorta/Gonad/Mesonephros (AGM) region and the umbilical and vitelline arteries (Dzierzak and Speck 2008). The first HSCs are detected around embryonic day (E)10 of murine development in hematopoietic clusters emerging from the ventral wall of the aorta in the AGM and are defined by their unique capacity to reconstitute hematopoiesis of immunodepleted adult recipients in transplantation assays. The Notch pathway is required for HSC development in the vertebrate embryo, upstream of Runx1 (Burns, Traver et al. 2005) and GATA-2 (Robert-Moreno, Espinosa et al. 2005), but is dispensable for yolk sac primitive hematopoiesis (Robert-Moreno, Espinosa et al. 2007; Bertrand, Cisson et al. 2010; Bigas, Robert-Moreno et al. 2010). Importantly, Gata2 but not Runx1 expression was totally impaired in the aorta of Jag1-/- mutant mouse embryos, which fail to generate hematopoiesis (Robert-Moreno, Espinosa et al. 2005; Robert-Moreno, Guiu et al. 2008). Re-expression of GATA-2 partially rescued generation of hematopoietic cells from Jag1-/- AGM cells (Robert-Moreno, Guiu et al. 2008), consistent with its pivotal role in definitive embryonic and adult hematopoiesis (Tsai, Keller et al. 1994). The Notch signalling pathway and its function in controlling cell fate diversification is conserved through evolution. Notch receptors interact with their ligands that are expressed on neighboring cells, and this interaction 126 induces two sequential proteolytic events, which releases the intracellular Notch (ICN) fragment. Once activated, Notch associates with its specific coactivator Mastermind (Mam) and the nuclear factor RBP-Jκ to induce a transcriptional response, generally associated with the inhibition of a particular cell fate destiny from a population of equivalent cells (Reviewed in (Kopan and Ilagan 2009). Classical examples of Notch-dependent fate inhibition are: B-cell differentiation in the hematopoietic system; secretory lineage in the adult intestine and the neural fate from neural/glial precursors. Most of these Notch responses are context-dependent and associated with regulation of tissue-specific targets, however a group of Notch target genes belonging to the Hairy/Enhancer of split (HES) family are known to mediate Notch actions in a wide range of cell types and organisms (Kageyama, Ohtsuka et al. 2007). We previously found that activation of Notch in the AGM was concomitant with the transcriptional expression of several HES family genes (Robert-Moreno, Espinosa et al. 2005). HES proteins are bHLH transcriptional repressors including the Notchtargets HES-1, HES-3, HES-5, HES-7 and the HES Related proteins HRT-1 and HRT-2 (Reviewed in Kageyama, Ohtsuka et al. 2007). In the hematopoietic system, HES-1 has a major function in normal T-cell development but it is also directly involved in the maintenance of Notchinduced T-cell leukemias (Tomita, Hattori et al. 1999; Wendorff, Koch et al. 2010) (Espinosa, Cathelin et al. 2010). In addition, ectopic HES-1 inhibits 127 differentiation of bone-marrow HSC when cultured in vitro (Kunisato, Chiba et al. 2003), however nothing is known about a putative role of HES in regulating embryonic hematopoiesis or HSC generation. Here, we investigate the mechanisms that control Hematopoietic Stem Cell development downstream of Notch and demonstrate that Hes repressors are essential for regulating the levels of Gata2 in the AGM precursors and generate functional hematopoietic cells. 3.2.3. Results 3.2.3.1. Increased hematopoietic cluster emergence in Hes-deficient embryonic aortas. Hes1 is expressed in the aortic endothelium and the presumptive hematopoietic clusters of the AGM region (Figure 1A) (Robert-Moreno, Guiu et al. 2008), suggestive of a hematopoietic function. However, Hes1null embryos contain functional HSCs and progenitors in the fetal liver that are able to reconstitute all hematopoietic lineages, except T-cells, when transplanted into RAG2-null mice. We explored the possibility that other Hes members compensate for Hes1 deficiency during hematopoietic development, similarly to what occurs in the nervous system (Ohtsuka, Ishibashi et al. 1999). To this purpose, we used two genetic models, the first one carrying a constitutive deletion of Hes1 and Hes5 alleles in different combinations, and the second with constitutive deletion of one or 128 two copies of Hes5 combined with Hes1-loxP allele/s, that can be deleted after tamoxifen treatment when crossed with mice carrying the β-ActinCRE-ERT transgen. We first determined whether deletion of several Hes1 and Hes5 alleles in the AGM affects the expression of other members of this family of repressors. Importantly we found that Hes5, which is not expressed in the wildtype AGM (Figure 1A), was highly induced after genetic deletion of Hes1 (Figure 1B). In contrast, other Hes-family genes such as Hrt1/Hey1 and Hrt2/Hey2 were not affected (Figure 1B). Notch activity but also HRT-1 and HRT-2 functions are essential to induce the arterial program (Fischer, Schumacher et al. 2004), that constitutes the niche for HSC generation. Thus, we tested whether composed Hes1/5 deficiency precludes arterial development in the AGM. Analysis of E10.5 embryo sections stained with the pan-endothelial/hematopoietic CD31 marker did not reveal any grouse vascular abnormality in the AGM aorta at this stage (not shown and Figure 1E). Moreover, by immunostaining and qRT-PCR we found that different arterial-associated genes including EphrinB2 and SMA remained unaffected in these mutant embryos (Figure 1C and 1D), These results indicate that HES repressors are dispensable for maintaining the arterial program in the embryo. A more accurate analysis of the AGM aorta stained for CD31 (labeling endothelial and hematopoietic cells) demonstrated that Hes1-/-Hes5+/- and Hes1-/-Hes5-/- mutant embryos have much larger hematopoietic cluster structures when compared to wildtype embryos (Figure 1E). We confirmed the hematopoietic nature of these clusters by staining with anti-C-Kit (Figure 129 S1) and CD41 (Figure 1E). Quantification of CD41+ cells in the AGM from different wildtype and mutant embryos demonstrated that the number of Hes1 and Hes5 deleted alleles inversely correlated with the total number of hematopoietic progenitors per AGM present in the aortic endothelium (up to 10-fold) (p<0.001) (Figure 1F) and the number of enlarge clusters with more than 4 cells (p=0.004) (Figure 1G), indicating functional compensation between Hes1 ad Hes5 in the AGM. 3.2.3.2. Occupancy of the Gata2 promoter by HES-1 and Notch/RBP-Jκ Next, we determined the expression levels of different hematopoietic transcription factors in the AGM clusters from wildtype and mutant embryos. We found that Runx1, c-myb and Gata2 were over-expressed in Hes-deficient embryos compared with the wildtype (Figure 2A). Importantly, expression levels of Gata2, but not of Runx1 and c-myb, in the embryonic AGM directly and significantly correlates with the number of deleted Hes1/5 alleles in the mutants (p<0.001) (Figure 2A). Increase in Gata2 levels was also detected by in situ hybridization of the whole AGM region of Hes1-/Hes5+/- embryos. Specifically, Gata2 mRNA was upregulated in the hematopoietic clusters, mesenchyme, mesonephros, urogenital ridge and mesentery lining of these mutant embryos when compared with the wildtype (Figure 2B). 130 131 Figure 1: Increased hematopoietic cluster emergence in Hes-deficient embryonic aortas. (A) Transversal sections of embryos at 10.5 days showing expression of Hes5 and Hes1 by in situ hybridization. Scale bars = 200µm (SG: sympathetic ganglia; Ao: aorta; IAHC: intra aortic hematopoietic cluster; m: mesonephros; p2: p2 progenitors; head arrows indicate the structures). (B) Relative expression of the indicated genes from E10.5 AGM after 3 days of 4-OH-Tamoxifen treatments in explant cultures by quantitative RT-PCR. Fold increase to the wildtype control after β2-Microglobulin and GAPDH normalization. Bars represent the average ± sd determination from one representative of two experiments showing the fold increase versus the wildtype AGM. (C) Representative immunostaining of CD44 and SMA (Arterial markers) in the AGM region in wildtype or Hes mutant embryos (Ao: aorta in AGM region; A: dorsal aorta; m: mesonephros; nt: neural tube; cv: cardinal vein). (D) Relative expression of endothelial (PECAM1), arterial markers (EphrinB2, Smooth muscle actin (SMA) and CD44) and vein marker (EphB4) in E10.5 AGM after 3 days of 4-OH-Tamoxifen treatment in explant culture from embryos with the indicated genotypes by quantitative qRT-PCR. Bars represent the average ± sd determination from one representative of two experiments showing the fold increase versus the wildtype AGM. (E) Representative immunostaining of CD31 (endothelial and hematopoietic) and CD41 (hematopoietic precursors) in the AGM region in wildtype or Hes mutant embryos (Ao: aorta; IAHC: intra aortic hematopoietic cluster; m: mesonephros; nt: neural tube; head arrows indicate the structures). (F) Quantification of CD41+ cells per µm of AGM. Speraman test was used to assess the significance between wildtype allele number and CD41+ cells per µm. Correralion wildtype allele number and CD41+ cells per µm was significant p < 0,001 and ρ = 0,878. Bars represent the average of counted cells in two different embryos ± sem. (G) CD41+ clusters containing more than 4 cells per µm of AGM. Quantification of CD41+ cells per µm of AGM. Spearman test was used to assess the significance between wildtype alleles number and CD41+ clusters containing more than 4 cells per µm. Correralion between wildtype alleles number and CD41+ clusters containing more than 4 cells per µm was significant p = 0,004 and ρ = 0,976. Bars represent the average of counted clusters in two different embryos ± sem. Previous data from our group indicated that Gata2 is a downstream target of Notch in the AGM (Robert-Moreno, Espinosa et al. 2005). By sequence analysis of the Gata2 locus using Genomatix software, we have now 132 identified the presence of two RBP-Jκ and three HES motifs that are conserved in both the human and murine Gata2 gene (Figure 2C and 2D). Since HES-1 is the main Notch target and a well-characterized transcriptional repressor, we hypothesized that Notch might activate Gata2 transcription that is next antagonized by Notch-mediated induction of HES1. To address this possibility we first conducted Chromatin Immunoprecipitation (ChIP) analysis using 32D myeloid progenitor cells to determine the recruitment of RBP-Jκ, ICN1 and HES-1 proteins to the Gata2 locus. We tested Gata2 occupancy using primers that amplify 15 regions spanning from -3033 to +5827 relative to the TSS at the IS promoter (see Figure 2E). Of note that Gata2 can be transcribed from two different promoters (IS and IG), both yielding the same protein, but using a different first untranslated exon (Minegishi, Ohta et al. 1999), being IS hematopoietic/neural specific (Nagai, Harigae et al. 1994) and IG the general Gata2 promoter. Our analysis revealed RBP-Jκ and ICN1 occupancy in the -800 to +14 region that contains two of the three predicted RBP-Jκ binding sites, whereas HES-1 protein was found associated at the +3059 to +5203 region, which contains the three predicted HES-1 sites (Figure 2E). A similar pattern for Notch1, RBP-Jκ and HES-1 association was detected using dissected AGM tissue (Figure 2F) and human K562 myeloerythroid cells (not shown). Interestingly, we failed to detect RBP-Jκ or HES binding to the Gata2 promoter in murine NIH-3T3 fibroblasts (Figure S2). These results indicated that RBP-Jκ/Notch proteins occupy the IS 133 promoter while HES-1 binds the IG promoter of the Gata2 gene specifically in hematopoietic cells. 134 135 Figure 2:. Occupancy of the Gata2 promoter by HES-1 and Notch/RBPj (A) Relative expression of the indicated genes from E10.5 AGM after 3 days of 4-OH-Tamoxifen treatments in explant cultures by quantitative RT-PCR. Fold increase to the wildtype control after β2-Microglobulin and GAPDH normalization. Spearman test was used to assess the significance between wildtype alleles number and gene expression. Correralion between number of wildtype Hes alleles and Gata2 expression was significant p ≤ 0.001 and ρ = 0,985. Bars represent the average ± sd determination from one representative of two experiments showing the fold increase versus the wildtype AGM. (B) Representative in situ hybridization of Gata2 in the wildtype (n=2) or Hes mutant embryos (n=2, each genotype). Images show transversal section of E10.5 embryos in the AGM region. Scale bars in left panels = 200µm. Scale bars in right panels = 20µm. (C) Graphical representation of the mouse Gata2 promoter showing the putative RBPJ or HES binding consensus found by bioinformatic analysis. Asterisks indicate the conserved sites in the human Gata2 locus (IS: Specific promoter; IG: general promoter). (D) Sequence and location of the human and mouse conserved RBPJ and HES binding sites relative to IS and IG promoters. (E) Chromatin immunoprecipitation with the anti-HES-1, anti-ICN, anti-RBPJ and anti-IgG antibodies from myeloid 32D cells and analysis of the Gata2 locus by qPCR. One representative of two experiments is shown. (F) Chromatin immunoprecipitation with the anti-HES-1, anti-ICN1, antiRBPJ and anti-IgG antibodies from E10.5 AGM cells and analysis of the Gata2 locus by qPCR. One representative from 3 experiments is shown. 3.2.3.3.HES-1 is a negative regulator of the Gata2 promoter in hematopoietic cells. Previously, we demonstrated that embryos with deficient Notch activity failed to express Gata2 and we presented evidence that Gata2 is a transcriptional Notch-target (Robert-Moreno, Espinosa et al. 2005). However, ectopic expression of ICN1 in AGM cells does not increase Gata2 transcription (Figure S3), which could be explained by a negative regulation of the Gata2 promoter by Hes1. To test this possibility, we analyzed different Gata2 regulatory regions (see Figure 3A) for their capacity to respond to ectopically expressed ICN1 or HES-1. We found that Gata2p-IS, containing RBP-Jκ sites, but not of Gata2p-IG, was transcriptionally induced by the Notch co-activator Mam1, in a dose-dependent manner and 136 repressed by a dominant negative-RBP-Jκ (Figure S3B). Furthermore, Gata2p-IS promoter was activated 2 to 4 fold by ICN1 after inhibition of endogenous Notch activity by DAPT (Figure 3B and S2C). By mutational analysis we found that both predicted RBP-Jκ binding sites in the Gata2pIS promoter were functional, since only elimination of the two sites impaired the capacity of this reporter to be activated by Notch (Figure 3B). In similar experiments, Gata2p-IG reporter was repressed by ectopic HES-1 expression and activated in cells transduced with Hes1-specific shRNA (Figure 3C and 3D). Mutational analysis demonstrated that all three HESbinding sites were functional and sufficient to repress Gata2 reporter expression, since only the combined mutation of all three sites relieved Gata2 repression by HES-1 (Figure 3D). Together, these results indicate that Notch and HES co-regulate expression of the Gata2 promoter in vitro in an opposite manner. 137 Figure3: HES-1 is a negative regulator of the Gata2 promoter in hematopoietic cells. (A) Scheme of the Gata2 locus indicating the conserved RBPJ and HES binding sites (vertical arrows). Promoter regions used in the luciferase assay. (B) Luciferase activity from the Gata2 IS promoter with indicated point mutations in the RBPJ-binding consensus (bold/underlined) measured in HEK-293T cells. DAPT incubation was used to reduce endogenous Notch activity. (C) Luciferase activity of the Gata2 IG promoter in the presence or absence of HES-1. (D) Luciferase activity from the Gata2 IG promoter with the indicated point mutations (bold/underlined) in the HES-binding consensus. Average of three independent experiments is shown in all the plots (B-D) t test was used to assess the significance (*p<0,05). 3.2.3.4.Notch/RBP-Jκ and HES control Gata2 expression during embryonic development To test whether Notch and HES control Gata2 expression in the embryo, we generated point mutations for both RBP-Jκ sites (p7.0IS RBP12) on the 138 previously characterized Gata2-IS reporter construct (p7.0IS WT) (Figure 4A) (Minegishi, Ohta et al. 1999; Minegishi, Suzuki et al. 2003) and microinjected mutant or wildtype DNA into fertilized oocytes. The expression patterns of GFP driven by these promoters in the transgenic F0 (founders) mice were examined at E10.5 (Figure 4B). In embryos carrying the wildtype reporter, we found specific expression of GFP in the dorsal aorta and neural tissue (3/4 transgene-positive embryos as determined by PCR) as previously reported (Minegishi, Suzuki et al. 2003). Importantly, mutations in the core of the RBP-Jκ binding sites abolished the expression of the reporter gene in both the aorta and the neural tissue (0/9 transgenepositive embryos by PCR). Immunostaining with anti-GFP in embryo sections confirmed the expression pattern for the Gata2 reporter in wildtype embryos and the absence of GFP in the aorta or neural tube of embryos carrying the RBP-Jκ mutant reporter (Figure 4C). These results indicate that Notch/RBP-Jκ activity is required for driving Gata2 transgene expression from the IS promoter in both hematopoietic and neural tissues. We performed comparable experiments using the p4.0 reporter with mutations of all three HES-sites. This reporter contains the -4 to +5kb sequence of the murine Gata2 locus (including IS and IG promoter regions) and recapitulates the expression pattern of endogenous Gata2 gene in transgenic embryos (including expression in the AGM and neural tube) (Minegishi, Ohta et al. 1999; Minegishi, Suzuki et al. 2003). Based on our in vitro data, we predicted that HES-binding site mutations might either increase the reporter levels within a particular cell population or change the 139 cell-type specificity of the reporter. Therefore, as readout for these experiments, we determined GFP levels in specific hematopoietic (CD31+/C-Kit+) and endothelial (CD31+C-Kit-CD45-) cell populations from E9.5 to 10.5 transgenic embryos (26-35 somite pairs). Our results demonstrate that the wildtype reporter was expressed at low levels in both the endothelial (CD31+C-Kit-CD45-) and hematopoietic (CD31+C-Kit+) AGM cells, at similar percentages. Interestingly, HES-mutant reporter led to increased percentage of GFP-positive cells that was not significant in the endothelial lineage (from 8 ± 2.6% to 18.1 ± 4.6%; p=0.07) but reached statistical significance in the hematopoietic population (from 6.6 ± 3.7% to 32.8 ± 9%; p = 0.02) (Figure 4D and 4E). Moreover, hematopoietic cells carrying the mutant reporter showed increased expression levels of GFP (Figure 4D and 4F). These results demonstrate that Notch is required for Gata2 expression in the AGM whereas HES restricts the levels of Gata2 specifically within the presumptive AGM hematopoietic cells. 3.2.3.5. AGM hematopoietic clusters in Hes-deficient embryos lack functional progenitors and HSCs. We have shown that Hes deficiency results in enlarged hematopoietic clusters in the developing AGM. To test the functionality of these mutant hematopoietic cells, we used the mouse line containing the tamoxifeninducible β-actin-CRE-ERT transgene in different combinations of the loxPHes1 and the Hes5 deleted alleles. E10.5 AGMs of the different genotypes were dissected and cultured as explants for 3 days in the presence of 4OH- 140 tamoxifen to induce the deletion. Dissociated AGM cells were then plated in methylcellulose to determine the number of hematopoietic progenitors by CFC assays. We found that AGM cells containing three deleted Hes alleles showed significantly different number of functional progenitors (CFC) when compared with those carrying one or two deleted alleles 141 142 Figure 4: Notch/RBPJ and HES-1 regulates Gata2 promoter in vivo during embryonic development. (A) Schematic representation of the Gata2 promoter constructs analyzed in vivo. (B) Representative bright-field (Left) and fluorescent (right) micrographs of embryos containing the wildtype or RBPJ mutant Gata2p7.0IS construct at E10.5. Summary of the data from at least 3 independent experiments is indicated in the lower panel (me: mesencephalon, FL: fetal liver, AGM: aorta-gonadmesonephros region, head arrows indicate the structures) Fisher test was used to assess the significance. (C) Transversal section of the AGM region and neural tube of E10.5 embryos containing the indicated constructs showing expression of GFP as detected by immunostaining. Nuclear staining by DAPI is represented in blue. Scale bars = 200µm (nt: neural tube; m: mesonephros, Ao: aorta, IAHC: intra aortic hematopoietic cluster, p2: neural tube p2 domain, head arrows indicate the structures). (D) Flow cytometry analysis of GFP in the CD31+ subpopulation of E10.5 AGM cells from 2 representative embryos containing the p4.0 wildtype or mutant for HES sites. (E-F) Average percentage (E) and mean intensity (F) of GFP+ cells in the CD31+ C-Kit+ and CD31+ C-Kit- CD45- populations from 3 transgene-positive embryos from at least 2 independent experiments. (Figure 5A). These results suggest that a minimum threshold of HES levels (2 wildtype alleles) is required to maintain the functionality of hematopoietic progenitors. Analysis of the colonies by PCR confirmed the deletion of the inducible alleles after tamoxifen treatment (Figure 5B). Similarly, hematopoietic progenitors (defined as CD34+C-Kit+) isolated from constitutive Hes1-/-Hes5-/- embryos were unable to produce sca1+C-Kit+ progenitors and CD45+ hematopoietic cells when cultured on OP9 stroma, compared with Hes5-/- cells (Figure 5C). To measure the number and functionality of Hes-defficient hematopoietic progenitors in an in vivo system, we determined the capacity of dissociated AGM cells from tamoxifen-treated explants of different genotypes to generate spleen colonies (CFU-S11) in lethally irradiated mice. Since Hes1f/fHes5-/- mutant animals were obtained on a mixed background, we 143 used immunodeficient SCID-Beige mice as recipient for the transplants to avoid graft rejection. In these experiments, we obtained an average number of 16.6 ± 1.4 CFU-S11 from one wildtype E10.5 AGM explant (1 embryo equivalent, 1ee). In contrast, the number of colonies obtained from AGM explants with all four Hes alleles deleted was comparable to the noninjected/irradiated controls (4 ± 0.3) (Figure 5D and 5E), and the few colonies detected did not contain Hes1 deletion (Figure 5D, lower panels). Next, we determined the presence of HSC after induction of Hes1 deletion in E10.5 AGM explants from the β-actin-CRE-ERT loxP-Hes1 R26-YFP f/+ Hes5 +/- background compared with the β-actin-CRE-ERT R26-YFP f/+. To increase the repopulating capacity of E10.5 AGM, we injected two embryo equivalents (2 ee) per each SCID-Beige recipient together with spleen cells as a support. In these experiments we did not detect any HSC activity from Hes-deficient AGM cells (3 alleles deleted) while all mice transplanted with control cells showed hematopoieitic reconstitution after 8 weeks (n=4, considering >1% of donor cells in peripheral blood). Together, these results indicate that HES proteins are essential downstream effectors of Notch signaling in the generation of HSC during embryonic development, being the control of Gata2 levels one of the mechanisms underlying this function. 144 145 Figure 5: AGM hematopoietic clusters in Hes-deficient embryos lack functional progenitors and HSCs. (A) Determination of CFU-C hematopoietic progenitors obtained from E10.5 AGM after 3 days of explant culture in the presence of 4-OHTamoxifen to induce Hes1 deletion. Kruskal-Wallis and Mann-Whitnay U test was used to assess the significance. Average and sem from 3 or more AGM explants from at least 3 independent experiments is shown. (B) PCR from isolated colonies to test Hes1 deletion. (C) Flow cytometry analysis from CD34+C-KIT+ sorted cells from E9.5 and E10.5 embryos with the indicated genotypes cultured on OP9 with factors (SCF+IL3+Epo+G-CSF) for 10 days. Dot plots are representative of three independent experiments. (D) Spleen colonies from 4-OH-Tamoxifen treated E10.5 AGM explants from the indicated genotypes injected in irradiated SCID-Beige mice. Average of (CFU-S11) from 3 or more injected animals is shown. t test was used to asses significance (E), representative spleen images (upper panel) and PCR from dissected spleen colonies to test for Hes1 deletion (lower panel) are shown. (F) Flow cytometry analysis at two months of SCID-Beige transplanted animals. Each mouse was transplanted with 2 embryo equivalent AGMs after 3 days of 4-OH-Tamoxifen treatments in explant culture. Two representative controls (n=4) and two Hes1 f/f Hes5 +/- R26-YFP f/+ β-ACTIN-CRE-ERT (n=2) are shown. Controls with more than 1% of YFP cells were considered as reconstituted. 3.2.4. Discussion Deciphering the molecular events that control hematopoietic development is a crucial issue for understanding how functional HSCs are produced and maintained. In this work, we focused on the role of HES proteins, which are the principal mediators of Notch pathway in multiple systems. Fetal liver of Hes1-deficient embryos had been previously analyzed and no major defects on HSC activity were found (Tomita, Hattori et al. 1999), although a direct measure of HSC activity was not performed. Importantly, here demonstrate that Hes5 (that is functionally equivalent to Hes1) is expressed the novo in the aorta of Hes1 mutants, which suggest the possibility of functional compensation between 146 both proteins. In agreement with this, we found that cumulative deletion of Hes1 and Hes5 alleles leads to the generation of non-functional hematopoietic progenitors in the developing AGM and the total absence of HSC activity. In addition, we demonstrate that the Gata2 gene is directly activated by Notch/RBPJκ but repressed by HES-1 specifically in the C-Kit+ cell population of the emerging AGM clusters. Thus, in Hes deficient mutants, Gata2 is upregulated, the number of hematopoietic cluster cells increased but they only contain nonfunctional hematopoietic cells, in agreement with previous work demonstrating that Gata2 levels need to be tightly regulated to generate and maintain functional HSC (Ling, Ottersbach et al. 2004; Rodrigues, Janzen et al. 2005). Current experiments are focused on exploring the possibility to revert hematopoietic function in the Hes mutants by limiting Gata2 levels. Mechanistically, one possibility is that overexpression of Gata2 in AGM cells imposes an impairment to cell cycle entry in the hematopoietic progenitor populations, as it was demonstrated in the cord blood HSC (Tipping, Pina et al. 2009). Analysis of the Gata2 regulatory regions led to the identification of two functional DNA-binding sites for Notch/RBP-Jκ and three for HES and we demonstrated their relevance in the activation and repression of this gene in the embryonic aorta, the niche of nascent HSCs. Since Hes is robustly induced by Notch, activation of this pathway in the hematopoietic progenitors might induce both positive and negative signals in the Gata2 gene resulting in the so-called Type I incoherent FeedForward Loop (I1-FFL) (Mangan and Alon 2003), that from our predictions would limit Gata2 activation in this tissue (Figure 6A). These type of circuits have previously been found to regulate Notch-dependent cell fate decisions in Drosophila (Krejci, Bernard et al. 2009). Here, we propose that following Notch 147 activation, likely through Jagged1, Gata2 and Hes genes start to be transcribed until HES protein reaches the threshold for repressing the Gata2 promoter. Then Gata2 transcription decreases and its mRNA and protein concentration drops, resulting in a pulse-like dynamics that it is required to achieve limited/transient (producing functional progenitors) rather than high/sustained activation (producing non-functional progenitors) (see model in Figure 6B). I1-FFL are also known to regulate biphasic responses where the output levels (in this case Gata2) depends on the input dose (Notch intensity) (Kim, Kwon et al. 2008). By computer modeling, it has been demonstrated that I1-FFL regulate time-dependent or dosedependent biphasic behaviors in a mutually exclusive manner (Kim, Kwon et al. 2008), and further work is needed to address whether Gata2 regulation by Notch and Hes follows one of these two models. In this work, we have dissected the effects of a single Notch pathway mediator (HES) during the generation of HSC. However, this is an extremely simplified picture of the Notch-dependent regulatory networks occurring during this process and many other elements and pathways will be sequentially incorporated to the final model. One example is the restriction imposed by the availability of Notch ligands to the Notch-responding cell that will depend on other upstream signals such as WNT (Estrach, Ambler et al. 2006; Rodilla, Villanueva et al. 2009), the specificity of Notch for its ligands that is modulated by Fringe-mediated glycosylation (Koch, Lacombe et al. 2001) or other interactions with the niche, which will be crucial to achieve the proper intensity of Notch signal in individual AGM cells. Another variable is HES-1 protein stability that is regulated by the JAK/STAT pathway and directly impacts on HES oscillation (Yoshiura, Ohtsuka et al. 2007). In addition, Hes1 transcription itself is controlled by a negative auto-regulatory feedback loop (Hirata, Yoshiura et al. 2002). Finally, a complex network of interactions that involves FLI1 and SCL and 148 operates downstream of Notch and GATA-2 in the HSC and hematopoietic precursors, has already been modeled (Narula, Smith et al. 2010). In summary, we have here deciphered the first mechanism downstream of Notch signaling that regulates HSC development. Figure 6: Model for Notch control of Gata2 doses through Hes in the nascent HSC. (A) Hematopoietic clusters and endothelium express different levels of Notch and ligands that signal to activate the Notch receptor. Once activated Notch induces the expression of Gata2 and also Hes1. Subsequently, HES-1 protein is produced that directly stops the production of Gata2 mRNA. (B) In the absence of Hes1, Gata2 levels and domains are expanded and result in the generation of a great number of non-functional progenitors. (C) When HES-1 is produced Gata2 levels are limited to the just-right dose to generate functional HSC/Progenitors. 149 3.2.5. Matherials and methods 3.2.5.1. Animals CD1, C57B6/J wildtype and Jagged1-/- (Xue, Gao et al. 1999), Hes1-/;Hes5-/- (Ohtsuka, Ishibashi et al. 1999), Hes1 f/f (Imayoshi, Shimogori et al. 2008) Rbpj -/- (Oka, Nakano et al. 1995) and β-actin-CRE-ERT (Hayashi and McMahon 2002) strains were used. R26-RYFP f/f (Strain Name: B6.129X1-GT(ROSA)26Sortm1(EYFP)Cos/J) was obtained from Jackson Laboratories. C57BL6/J oocytes were used in F0 assays. SCID-Beige animals were used as recipients in CFU-S11 and long-term reconstitution experiments. Animals were kept under pathogen-free conditions and all procedures were approved by the Animal Care Committee of PRBB (regulation of Generalitat de Catalunya). Embryos were obtained from timed pregnant females and staged by somite counting: E10.5 (31-40 sp) and E11.5 (43-48 sp). The detection of vaginal plug was designated as day 0.5. Mice and embryos were all genotyped by PCR. 3.2.5.2. Quantitative RT–PCR Total RNA was extracted with Qiagen kit and RT-First Strand cDNA Synthesis kit (Amersham Pharmacia Biotech, GE Healthcare, Buckinghamshire, UK) was used to produce cDNA. qRT–PCR was performed in LightCycler480 system using SYBR Green I Master kit (Roche, Basel, Switzerland). Primers used are in Table S1. 150 3.2.5.3. Promoter analysis, site-directed mutagenesis and luciferase assays. Human and murine Gata2 promoters were analyzed using Genomatix software. For luciferase assays, Gata2p-IG was previously described (Minegishi, Ohta et al. 1998) which covers from -482 of IG trasnscript to exon2. Gata2p-IS was generated cloning the region from -794 to +93 of exon IS into the pGL3 vector and verified by sequencing. The primers used were: Fwd (5’AAAACTCGAGGGTGCTACAAGGATGTGGTTGC3’) and Rev (5’AAAAAAGCTTGCTATGGT TGAGGTGATCTTAGGC3’). Mutations were introduced into wildtype plasmids using the QuikChange Site-Directed Mutagenesis kit from Stratagene (La Jolla, CA, USA) according to the manufacturer’s instructions. The mutations introduced and primers used are specified in the Table S2. All mutations were verified by DNA sequencing. Luciferase reporter assays were performed in HEK-293T cells. Cells were seeded in 12-well plates at a density of 5 × 104 cells/well. Equal amounts (150ng) of the different Gata2 reporter constructs. ICN1, Mastermind, dnRbpj, shHes1 (MISSION, TRCN0000018989) or irrelevant DNA and 150 ng CMV-β-Galactosidase plasmids were transfected in triplicate wells. Cells were transfected using Polyethylenimine (PEI) (Polysciences, Inc.). To reduce endogenous ICN1 levels, HEK-293T cells were treated with γsecretase inhibitor, DAPT (Calbiochem), 50µM final concentration for 72 hours before transfection and during the assay. Luciferase was measured after 48h of transfection following manufacturer instructions (Luciferase 151 Assay System; Promega). Expression levels of transfected proteins were verified by western blot. 3.2.5.4. Chromatin immunoprecipitation assay Chromatin was obtained from a pool of 40 dissected AGMs at E10.5, 32D myeloid progenitor cells line. Chromatin immunoprecipitation (ChIP) was performed as previously described (Aguilera et al, 2004) with minor modifications. In brief, cross-linked chromatin from E10.5 dissected AGMs or 32D cell line was sonicated for 10 minutes, medium-power, 0.5-interval; with a Bioruptor (Diagenode) and precipitated with anti-RBP-Jκ (Chu and Bresnick 2004) anti-ICN1(abcam) and anti-HES1 (sc-13844 or AVIVA). After crosslinkage reversal, DNA was used as a template for PCR. RT-PCR was performed with SYBR Green I Master (Roche) in LightCycler480 system. Primers used are in Table S3. 3.2.5.5. Cell culture, viral particle production and viral infection 32D were cultured in Iscoves 10%FBS supplemented with 10% WeHi-IL3 conditioned media. HEK-293T cells were seeded at a density of 5x105 cells/well and transfected with ICN1-IRES-GFP with PEI (Polysciences, Inc.). Hematopoietic cells from disrupted AGMs were incubated in Iscove's, 10% FBS, 10% IL3- and 10% SCF-conditioned medium plus 0.2 µg/ml IL6, 0.1 µg/ml flt3. Recombinant lentivirus were produced by transient transfection of HEK-293T cells according to Tronolab protocols (http://tcf.epfl.ch/page-6764.html). Briefly, subconfluent HEK-293T were 152 cotransfected with 20 µg of transfer vector, 15 µg of packaging plasmid (psPAX2), and 6µg of envelope plasmid (pMD2.G). After 3 days, supernatant was ultracentrifuged in Beckman L-70 at 26000rpm for 2 hours at 4ºC and viral pellet resuspended in 100 ml of PBS. 20µl of fresh viral suspension was used per infection. To assess hematopoietic potential of sorted populations from Hes1-/-Hes5-/- mutant mice, 500 CD34+ C-Kit+ cells were FACS-purified and added to confluent cultures of OP9 stromal cells and cultured for up to 10 days in the presence of 100ng/mL Interleukin-3 (IL-3), Granulocyte colony 100U/mL Stem stimulating cell factor factor (G-CSF) (SCF), 100ng/mL and 200ng/mL Erythropoietin (Epo). Supplemented differentiation medium was replaced every 4 days. Cultures were maintained for up to 12 days. Cells were harvested by pipetting and examined for hematopoietic development by flow cytometry. 3.2.5.6. Flow cytometry analysis and sorting The AGM region was dissected from E10.5 embryos and dissociated in 0.12% collagenase (Sigma) in PBS supplemented with 10% FBS for 30min at 37°C. The cells were analysed by flow cytometry in a FACScalibur (Becton & Dickinson) or FACSaria (Becton & Dickinson) and FlowJo software. Dead cells were excluded by 7-aminoactinomycin-D staining (Molecular Probes) in the FACScalibur analysis and with Hoechst or DAPI in the FACSaria analysis. The yolk sacs and lower trunk regions were harvested from individually dissected embryos washed of maternal blood 153 (Fraser et al., 2002). The anterior regions were used for genotyping (Ohtsuka et al., 1999). Yolk sacs and lower trunk regions were dissociated with 0.1% w/v collagenase types I and IV (Sigma, U.S.A.) for 1 hr at 30°C. Cells were then blocked with normal mouse serum and incubated with fluorescence-conjugated antibodies. For flow cytometric analysis, cells were resuspended in Hank’s buffered Saline Solution (Gibco BRL, Grand Island, NY, U.S.A.) containing Propidium Iodide (Sigma, U.S.A.) for dead cell exclusion. Flow cytometry was performed using a FACS VantageTM (Becton Dickinson,USA) with the Cellquest (Becton Dickinson) and data analysed using the Flowjo (Treestar, CA, USA) software package. The antibodies CD31-PE, Sca1-PE, CD45-FITC, Ter119-PE, CD45- PerCPCy5.5, CD117-APC were purchased from Pharmingen. VE-cadherinAPC was generated inhouse. 3.2.5.7. Immunostaining E10.5 embryos were fixed overnight in 4% paraformaldehyde (Sigma) at 4°C and frozen in Tissue-tek (Sakura). Sections (5 µm) were fixed with 20°C methanol for 15 min and block-permeabilized in 10% FBS, 0.3% Surfact-AmpsX100 (Pierce) and 5% non-fat milk in PBS for 90 min at 4°C. Samples were stained with CD31 (PECAM1; Pharmingen at 1:50) or CD41 (Pharmingen, 1:50) or C-Kit (CD117; Pharmingen at 1:50) in 10% FBS, 5% non-fat milk in PBS overnight and HRP-conjugated secondary antibody (Dako) at 1:100 for 90 min and developed with Cy3 or coupled tyramide (PerkinElmer). Anti-GFP (MBL) was used at 1:100 and Alexa Fluor 488- 154 conjugated donkey-anti-rabbit (Invitrogen, 1:500) as a secondary antibody. Sections were mounted in Vectashield medium with 4’6-diamidino-2phenylindole (DAPI). 3.2.5.8. In Situ Hybridization For ISH, precisely timed embryos were fixed overnight at 4°C in 4% paraformaldehyde and embedded in OCT (Tissue-Tek). Embryos were sectioned in a Leica-RM2135 cryostat at 16µm. Digoxygenin (DIG)-labeled probes were generated from a Bluescript plasmid (Stratagene, La Jolla, CA) containing a partial mouse Gata2 cDNA (nt 170 to 871) and from plasmid containing Hes1 cDNA and Hes5 cDNA. Images were acquired with an Olympus BX-60. Representative images were edited on Adobe Photoshop CS4 software. 3.2.5.9. Transgenic mice. F0 assays Gata2 promoter inserts from plasmids (p4.0, p4.0muHes, p7.0 and p7.0 muRBP) were purified by NACS PREPAC (GIBCO BRL, Rockville, MD), adjusted to 5-10 ng/mL and injected into mouse oocytes using standard strategies. Transgene integration was confirmed by PCR from genomic DNA from embryonic tissues. The wildtype constructs p4.0 (KobayashiOsaki, Ohneda et al. 2005) and p7.0 (Minegishi, Ohta et al. 1999) were previously described and point mutations were introduced into these plasmids. 155 3.2.5.10. AGM explant culture The AGM region was dissected and cultured in explants for 3 days as previously described (Medvinsky and Dzierzak 1999). Briefly, AGMs were deposited on nylon filters (Millipore) placed on metallic supports and cultured in myeloid long-term culture medium (Stem Cell Technologies) supplemented with 10µM hydrocortisone (Sigma) in an air-liquid interphase. The induction of β-ACTIN-CRE-ERT transgene was performed by adding 4OH-tamoxifen (Sigma) every 36h to the explant medium at final concentration 1µM. 3.2.5.11. Hematopoietic colony assay (CFU-C) and CFU-S11 and longterm reconstitution assay The explanted AGMs were digested in 0.12% collagenase (Sigma) in PBS supplemented with 10% fetal bovine serum (FBS) for 20 minutes at 37ºC and used for hematopoietic colony assay (CFU-C), CFU-S11 and long-term reconstitution assay experiments. In CFU-C assay were plated 20.000 cells/AGM were plated in duplicates in M-3434 semisolid medium (Stem Cell Technologies). After 7 days the presence of hematopoietic colonies was scored under the microscope. In CFU-S11 experiments, whole disrupted AGMs were injected intra-venous in SCID-Beige recipients. SCID-Beige recipients were previously irradiated at 2.5 gy. After 11 days the presence of colonies in the spleen was scored under the stereoscope. Some colonies were picked and lysed in Tris 0,01M pH=7,4, NaCl 0,15M, EDTA 0,01M, SDS 0,05%. From this lysate, Hes1 deletion was detected in 156 both types of colonies, CFU-C and CFU-S11, by PCR (primers in Table S4). In long-term reconstitution assay experiments, whole disrupted AGM were injected intra-venous in SCID-Beige recipients, with 500.000 white spleen cells, from a SCID-Beige animal, as a support cells. SCID-Beige recipients were previously irradiated at 2.5 gy. 3.2.5.12. Statistics Statistical analyses were performed using Microsoft Excel 2011 and SPSS. In Figures 3B, 3C, 3D, 4E, and 5D, Statistical significance was assessed using two-tailed t tests. In Figures 1F, 1G and 2A, statistical significance was assessed using Spearman test. In Figure 4A Statistical significance was assessed using Fisher test. In Figure 5A Kruskal-Wallis and MannWhitney U test was used to assess the significance. In all tests p values < 0.05 were considered significant. In all experiments: *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001. 3.2.6. Acknowledgements We thank Jessica González, Chris Vink (Dzierzak lab), Alex Maas (Erasmus University), Y. Kawatani and H. Suda (Shimizu and Yamamoto lab) for technical assistance. We would like to thank James Sharpe (CRG) for helpful genetic circuit consultation and members of the Bigas lab for critically reading the manuscript and helpful discussions. JG is a recipient of FPI BES-2008-005708 LE is an investigator of ISCIII program (02/30279). 157 This research was funded by the Ministerio de Economía y Competitividad (SAF2007-60080, PLE2009-0111, SAF2010-15450), Red Temática de Investigación Cooperativa en Cáncer (RTICC) (RD06/0020/0098), Agència de Gestió d’Ajuds Universitaris i de Recerca (AGAUR) (2009SGR-23, CONES2010-0006 to AB and Grants-in-Aids for Scientific Research from the Japanese Ministry of Education, Science, Sports and Culture to RS and MY. 3.2.7. References Bertrand, J.Y., Cisson, J.L., Stachura, D.L., and Traver, D. (2010). Notch signaling distinguishes 2 waves of definitive hematopoiesis in the zebrafish embryo. Blood 115, 2777-2783. Bigas, A., Robert-Moreno, A., and Espinosa, L. (2010). The Notch pathway in the developing hematopoietic system. Int J Dev Biol 54, 1175-1188. Burns, C.E., Traver, D., Mayhall, E., Shepard, J.L., and Zon, L.I. (2005). Hematopoietic stem cell fate is established by the Notch-Runx pathway. Genes Dev 19, 2331-2342. Chu, J., and Bresnick, E.H. (2004). Evidence that C promoter-binding factor 1 binding is required for Notch-1-mediated repression of activator protein-1. J Biol Chem 279, 12337-12345. Dzierzak, E., and Speck, N.A. (2008). Of lineage and legacy: the development of mammalian hematopoietic stem cells. Nat Immunol 9, 129136. 158 Espinosa, L., Cathelin, S., D'Altri, T., Trimarchi, T., Statnikov, A., Guiu, J., Rodilla, V., Ingles-Esteve, J., Nomdedeu, J., Bellosillo, B., et al. (2010). The Notch/Hes1 pathway sustains NF-kappaB activation through CYLD repression in T cell leukemia. Cancer Cell 18, 268-281. Estrach, S., Ambler, C.A., Lo Celso, C., Hozumi, K., and Watt, F.M. (2006). Jagged 1 is a beta-catenin target gene required for ectopic hair follicle formation in adult epidermis. Development 133, 4427-4438. Fischer, A., Schumacher, N., Maier, M., Sendtner, M., and Gessler, M. (2004). The Notch target genes Hey1 and Hey2 are required for embryonic vascular development. Genes Dev 18, 901-911. Hayashi, S., and McMahon, A.P. (2002). Efficient recombination in diverse tissues by a tamoxifen-inducible form of Cre: a tool for temporally regulated gene activation/inactivation in the mouse. Dev Biol 244, 305-318. Hirata, H., Yoshiura, S., Ohtsuka, T., Bessho, Y., Harada, T., Yoshikawa, K., and Kageyama, R. (2002). Oscillatory expression of the bHLH factor Hes1 regulated by a negative feedback loop. Science 298, 840-843. Imayoshi, I., Shimogori, T., Ohtsuka, T., and Kageyama, R. (2008). Hes genes and neurogenin regulate non-neural versus neural fate specification in the dorsal telencephalic midline. Development 135, 2531-2541. Kageyama, R., Ohtsuka, T., and Kobayashi, T. (2007). The Hes gene family: repressors and oscillators that orchestrate embryogenesis. Development 134, 1243-1251. 159 Kim, D., Kwon, Y.K., and Cho, K.H. (2008). The biphasic behavior of incoherent feed-forward loops in biomolecular regulatory networks. Bioessays 30, 1204-1211. Kobayashi-Osaki, M., Ohneda, O., Suzuki, N., Minegishi, N., Yokomizo, T., Takahashi, S., Lim, K.C., Engel, J.D., and Yamamoto, M. (2005). GATA motifs regulate early hematopoietic lineage-specific expression of the Gata2 gene. Mol Cell Biol 25, 7005-7020. Koch, U., Lacombe, T.A., Holland, D., Bowman, J.L., Cohen, B.L., Egan, S.E., and Guidos, C.J. (2001). Subversion of the T/B lineage decision in the thymus by lunatic fringe-mediated inhibition of Notch-1. Immunity 15, 225236. Kopan, R., and Ilagan, M.X. (2009). The canonical Notch signaling pathway: unfolding the activation mechanism. Cell 137, 216-233. Krejci, A., Bernard, F., Housden, B.E., Collins, S., and Bray, S.J. (2009). Direct response to Notch activation: signaling crosstalk and incoherent logic. Sci Signal 2, ra1. Kunisato, A., Chiba, S., Nakagami-Yamaguchi, E., Kumano, K., Saito, T., Masuda, S., Yamaguchi, T., Osawa, M., Kageyama, R., Nakauchi, H., et al. (2003). HES-1 preserves purified hematopoietic stem cells ex vivo and accumulates side population cells in vivo. Blood 101, 1777-1783. Ling, K.W., Ottersbach, K., van Hamburg, J.P., Oziemlak, A., Tsai, F.Y., Orkin, S.H., Ploemacher, R., Hendriks, R.W., and Dzierzak, E. (2004). GATA-2 plays two functionally distinct roles during the ontogeny of hematopoietic stem cells. J Exp Med 200, 871-882. 160 Mangan, S., and Alon, U. (2003). Structure and function of the feed-forward loop network motif. Proc Natl Acad Sci U S A 100, 11980-11985. Medvinsky, A., and Dzierzak, E. (1999). Embryonic beginnings of definitive hematopoietic stem cells. Blood 94, 3613-3614. Minegishi, N., Ohta, J., Suwabe, N., Nakauchi, H., Ishihara, H., Hayashi, N., and Yamamoto, M. (1998). Alternative promoters regulate transcription of the mouse GATA-2 gene. J Biol Chem 273, 3625-3634. Minegishi, N., Ohta, J., Yamagiwa, H., Suzuki, N., Kawauchi, S., Zhou, Y., Takahashi, S., Hayashi, N., Engel, J.D., and Yamamoto, M. (1999). The mouse GATA-2 gene is expressed in the para-aortic splanchnopleura and aorta-gonads and mesonephros region. Blood 93, 4196-4207. Minegishi, N., Suzuki, N., Yokomizo, T., Pan, X., Fujimoto, T., Takahashi, S., Hara, T., Miyajima, A., Nishikawa, S., and Yamamoto, M. (2003). Expression and domain-specific function of GATA-2 during differentiation of the hematopoietic precursor cells in midgestation mouse embryos. Blood 102, 896-905. Nagai, T., Harigae, H., Ishihara, H., Motohashi, H., Minegishi, N., Tsuchiya, S., Hayashi, N., Gu, L., Andres, B., Engel, J.D., et al. (1994). Transcription factor GATA-2 is expressed in erythroid, early myeloid, and CD34+ human leukemia-derived cell lines. Blood 84, 1074-1084. Narula, J., Smith, A.M., Gottgens, B., and Igoshin, O.A. (2010). Modeling reveals bistability and low-pass filtering in the network module determining blood stem cell fate. PLoS Comput Biol 6, e1000771. 161 Ohtsuka, T., Ishibashi, M., Gradwohl, G., Nakanishi, S., Guillemot, F., and Kageyama, R. (1999). Hes1 and Hes5 as notch effectors in mammalian neuronal differentiation. Embo J 18, 2196-2207. Oka, C., Nakano, T., Wakeham, A., de la Pompa, J.L., Mori, C., Sakai, T., Okazaki, S., Kawaichi, M., Shiota, K., Mak, T.W., et al. (1995). Disruption of the mouse RBP-J kappa gene results in early embryonic death. Development 121, 3291-3301. Robert-Moreno, A., Espinosa, L., de la Pompa, J.L., and Bigas, A. (2005). RBPjkappa-dependent Notch function regulates Gata2 and is essential for the formation of intra-embryonic hematopoietic cells. Development 132, 1117-1126. Robert-Moreno, A., Espinosa, L., Sanchez, M.J., de la Pompa, J.L., and Bigas, A. (2007). The notch pathway positively regulates programmed cell death during erythroid differentiation. Leukemia 21, 1496-1503. Robert-Moreno, A., Guiu, J., Ruiz-Herguido, C., Lopez, M.E., Ingles-Esteve, J., Riera, L., Tipping, A., Enver, T., Dzierzak, E., Gridley, T., et al. (2008). Impaired embryonic haematopoiesis yet normal arterial development in the absence of the Notch ligand Jagged1. EMBO J 27, 1886-1895. Rodilla, V., Villanueva, A., Obrador-Hevia, A., Robert-Moreno, A., Fernandez-Majada, V., Grilli, A., Lopez-Bigas, N., Bellora, N., Alba, M.M., Torres, F., et al. (2009). Jagged1 is the pathological link between Wnt and Notch pathways in colorectal cancer. Proc Natl Acad Sci U S A 106, 63156320. 162 Rodrigues, N.P., Janzen, V., Forkert, R., Dombkowski, D.M., Boyd, A.S., Orkin, S.H., Enver, T., Vyas, P., and Scadden, D.T. (2005). Haploinsufficiency of GATA-2 perturbs adult hematopoietic stem-cell homeostasis. Blood 106, 477-484. Tipping, A.J., Pina, C., Castor, A., Hong, D., Rodrigues, N.P., Lazzari, L., May, G.E., Jacobsen, S.E., and Enver, T. (2009). High GATA-2 expression inhibits human hematopoietic stem and progenitor cell function by effects on cell cycle. Blood 113, 2661-2672. Tomita, K., Hattori, M., Nakamura, E., Nakanishi, S., Minato, N., and Kageyama, R. (1999). The bHLH gene Hes1 is essential for expansion of early T cell precursors. Genes Dev 13, 1203-1210. Tsai, F.Y., Keller, G., Kuo, F.C., Weiss, M., Chen, J., Rosenblatt, M., Alt, F.W., and Orkin, S.H. (1994). An early haematopoietic defect in mice lacking the transcription factor GATA-2. Nature 371, 221-226. Wendorff, A.A., Koch, U., Wunderlich, F.T., Wirth, S., Dubey, C., Bruning, J.C., MacDonald, H.R., and Radtke, F. (2010). Hes1 is a critical but context-dependent mediator of canonical Notch signaling in lymphocyte development and transformation. Immunity 33, 671-684. Xue, Y., Gao, X., Lindsell, C.E., Norton, C.R., Chang, B., Hicks, C., Gendron-Maguire, M., Rand, E.B., Weinmaster, G., and Gridley, T. (1999). Embryonic lethality and vascular defects in mice lacking the Notch ligand Jagged1. Hum Mol Genet 8, 723-730. Yoshiura, S., Ohtsuka, T., Takenaka, Y., Nagahara, H., Yoshikawa, K., and Kageyama, R. (2007). Ultradian oscillations of Stat, Smad, and Hes1 163 expression in response to serum. Proc Natl Acad Sci U S A 104, 1129211297. 164 3.2.8. Supplementary Figures 165 166 167 168 169 170 4.CONCLUSIONS 171 172 4.1.CONCLUSIONS 4.1.1. Notch ligands and receptors: 1. Jagged1 but not Jagged2 is the key ligand to activate hematopoietic program in the AGM. Moreover Jagged1 is dispensable to determine arterial program. 2. There is active Notch in hematopoietic cluster like structures in the AGM region. 4.1.2. Notch target genes: 3. Gata2 and Hes1 but not Hes5 are expressed in the hematopoietic clusters in the AGM region. 4. Notch/RBP-Jκ is required to activate Gata2 expression through two RBP-Jκ binding sites in the IS promoter at E10,5 in the AGM and the neural tube. 5. Hes1 and Hes5 can repress Gata2 expression through three Hes binding sites in the hematopoietic clusters (cKITposCD31pos) of the AGM at E10,5. 6. Hes5 compensates for Hes1 deficiency in the AGM of Hes1 knockouts. 173 7. Hes1 and Hes5 null mice have enlarged but non-fuctional hematopoietic progenitors and HSC that cannot reconstitute adult irradiated mice. Moreover Hes1 and Hes5 double knockouts have no defects in the arterial determination. 174 175 176 5.DISCUSSION 177 178 5.1. DISCUSSION 5.1.1. Notch regulates hematopoietic program through Jagged1 One of the best-characterized functions of Notch is the induction of the arterial program during development. Delta4, Notch1, RBP-Jκ and Mindbomb1, are required to generate arteries (Krebs, Xue et al. 2000; Shutter, Scully et al. 2000; Duarte, Hirashima et al. 2004; Krebs, Shutter et al. 2004; Burns, Traver et al. 2005) characterized by the expression of specific markers such as EphrinB2, CD44 and SMA. Our group previously showed that Delta4, JAG-1 and JAG-2 are expressed in the AGM region but not Delta1 (E9.5 and E10.5)(Robert-Moreno, Espinosa et al. 2005). Delta1 is expressed at latter (E13,5-E18,5) stages, once arteries have already been formed and the AGM is not a hematopoietic organ anymore. Delta1 has a role in maintenance of the arteries because embryos deficient for Delta1 have miss-regulation of arterial markers SMA, EphrinB2 and neuropilin-1 (Sorensen, Adams et al. 2009). In Notch1, RBP-Jκ and Mindbomb1 knockout mice and zebrafish, arterial program is affected, and there is not hematopoiesis (Kumano, Chiba et al. 2003; Burns, Traver et al. 2005; Robert-Moreno, Espinosa et al. 2005). Now we have identified Jag1 as the key ligand to activate hematopoietic program without affecting arterial formation. The fact that Jaged1 knockouts failed to generate blood 179 but contained intact arteries undoubtedly demonstrated that Notch was required for definitive hematopoiesis in vivo, and uncoupled this Notch function from its role in arterial specification. In contrast, we did not detect any hematopoietic phenotype in Jag2 knockout mice. 5.1.2. Gata2 is a direct Notch target in the AGM. Once Notch is activated in the presumptive hematopoetic precursors, different Notch targets are expressed whose function is not well understood. Candidate Notch targets involved in the hematopoietic program includes Hes genes, Gata2 and Runx1. Runx1 is a transcription factor required to generate HSC from hemogenic endothelium but not thereafter (Chen, Yokomizo et al. 2009). It has been propose that Runx1 could be responsible of some Notch1 Knockout defects(Nakagawa, Ichikawa et al. 2006). However non-direct relation between Notch and Runx1 has been shown. GATA-2 is involved in regulating Runx1 expression (Nottingham, Jarratt et al. 2007). It may explain the result we found in Jagged1 mutants, in which we demonstrate that Gata2 expression is abolished; Runx1 is downregulated in half of the embryos. As discussed in chapter 1.3.3.7. GATA-2 is an essential regulator of the hematopoietic program, and in the absence of Gata2, HSC are not 180 generated during embryonic development (Tsai, Keller et al. 1994). We have here analyzed the Gata2 promoter and identified 2 functional Notch/RBP-Jκ binding sites. We found that Notch is required to activate Gata2 through these 2 RBP-Jκ binding sites. Moreover, we have shown that the hematopoietic defects of Jagged1-deficient AGM cells can be partially rescued by overexpressing Gata2, which indicates the importance of Gata2 but suggests that other Notch targets are required in the hematopoietic program. Based on our data, one of our candidate genes is Hes1. 5.1.3. Gata2 is direct Hes-1 target in the AGM. In this thesis we have shown that Hes-1 is expressed in IAHC. Morover we have identified 3 Hes binding sites in Gata2 promoter. In addition to this we demonstrate that HES-1 protein regulates the Gata2 promoter and the Gata2 mRNA levels in the hematopoietic clusters. Fetal liver of Hes1-deficient embryos had been previously analyzed. When HSCs from fetal liver are transplanted, T-cell development is blocked in the engrafted cells (Tomita, Hattori et al. 1999). However, no defects are detected in the HSC compartment. In this thesis work, we have shown that in Hes1 knockout AGMs, Hes5 is upregulated and partially compensate the phenotype associated to Hes1 deficiency. Thus, deletion of Hes1 and Hes5 alleles led to the accumulation of non-functional hematopoietic progenitors 181 and HSC in the developing AGM associated with increase of Gata2 expression. In adult BM, it has been shown that sustained overexpression of Gata2 results in HSC quiescence (G0) (Tipping, Pina et al. 2009). Further experiments such as cell cycle analysis in embryonic HSCs should be done to check if this is the case in Hes1/Hes5 null mice. 5.1.4. Notch, Hes1, and Gata2 generate an incoherent FeedForward Loop From our results, we propose a model in which Notch activates both, Gata2 and Hes1 in the hematopoietic progenitors and hematopoietic stem cells, and then, Hes1 repress Gata2 generating a genetic circuit that is called Type I incoherent Feed-Forward Loop (I1FFL) (Mangan and Alon 2003). This type of circuit has previously been found to regulate Notch-dependent cell fate decisions in Drosophila (Krejci, Bernard et al. 2009) and is likely responsible of generating a Gata2 pulse. More experiments such as ChIP sequencing precipitating chromatine with Hes1 should be done to identify other Hes1 targets. 182 5.1.5. Notch ligands and Notch target specificity. Arteries, veins and hematopoiesis. Activation of Notch, by Delta4 is required to determine the arterial fate during embryonic development, whereas Jagged1-mediated Notch activation is mandatory for HSC specification. These results suggest that Notch is able to activate different target genes in response to specific ligands to regulate different genetic programs. Due to the Delta4 knockout phenotype, most likely Hrt1, Hrt2 and Ephrin-B2 are downstream Notch targets controlling the arterial phenotype. HRT-1 and HRT-2 specifically repress EphB4. EphB4 is required to develop veins and inhibit arterial formation (Reviewed in Adams 2003). In this thesis we identify Gata2 and most likely Hes1 as specific Jagged1/Notch targets that affects hematopoietic program but not arterial. Since arteries are the niche of HSC during hematopoietic development, it is tempting to speculate that after arterial specification, arterial cells expressing Jagged1 can activate the Notch-dependent hematopoietic program. However, the inter-dependence between the arteriaI and the hematopoietic programs is not obvious. In this sense, mutant veins that aberrantly express arterial markers generate IAHC-like structures comparable to the hematopoietic clusters of the AGM (You, Lin et al. 2005). Also in embryos in which artery-vein boundaries are affected, such as Activin A-receptor type II-like1 (acvrl1, alk1) or endoglin (CD150), there are IAHC-like structures budding from vein-like vessels that do not express the arterial marker EphrinB2, but express 183 arterial/hematopoietic CD34 (Urness, Sorensen et al. 2000; Sorensen, Brooke et al. 2003). All this data suggest that the arterial program is upstream of hematopoietic program or that veins inhibit the hematopoietic program, but the formal proof is still missing. In this thesis we have added new molecular data that have helped to understand how Notch participates in embryonic HSC generation independent of Notch functions in the aortic program. Specifically, we have demonstrated that the Jagged1/Notch axis is crucial to regulate the hematopoietic program that generates the first HSCs, in a process independent of Notch arterial fate determination. Downstream of Jagged1/Notch we identified Hes1 and Gata2 genes as key elements of this process and demonstrated that both targets are related since Hes1 represses Gata2 expression. 184 6.BIBLIOGRAPHY 185 186 6.BIBLIOGRAPHY Adamo, L., O. Naveiras, et al. (2009). "Biomechanical forces promote embryonic haematopoiesis." Nature 459(7250): 1131-‐1135. Adams, R. H. (2003). "Molecular control of arterial-‐venous blood vessel identity." Journal of anatomy 202(1): 105-‐112. Andersson, E. R., R. Sandberg, et al. (2011). "Notch signaling: simplicity in design, versatility in function." Development 138(17): 3593-‐3612. Antonchuk, J., G. Sauvageau, et al. (2002). "HOXB4-‐induced expansion of adult hematopoietic stem cells ex vivo." Cell 109(1): 39-‐45. Artavanis-‐Tsakonas, S. (1988). "The molecular biology of the Notch locus and the fine tuning of differentiation in Drosophila." Trends in genetics : TIG 4(4): 95-‐100. Artavanis-‐Tsakonas, S., K. Matsuno, et al. (1995). "Notch signaling." Science 268(5208): 225-‐232. Artavanis-‐Tsakonas, S., M. A. Muskavitch, et al. (1983). "Molecular cloning of Notch, a locus affecting neurogenesis in Drosophila melanogaster." Proceedings of the National Academy of Sciences of the United States of America 80(7): 1977-‐1981. Baba, Y., K. P. Garrett, et al. (2005). "Constitutively active beta-‐catenin confers multilineage differentiation potential on lymphoid and myeloid progenitors." Immunity 23(6): 599-‐609. Barker, J. E. (1968). "Development of the mouse hematopoietic system. I. Types of hemoglobin produced in embryonic yolk sac and liver." Developmental biology 18(1): 14-‐29. Barker, N., J. H. van Es, et al. (2007). "Identification of stem cells in small intestine and colon by marker gene Lgr5." Nature 449(7165): 1003-‐ 1007. Baron, M. (2001). "Induction of embryonic hematopoietic and endothelial stem/progenitor cells by hedgehog-‐mediated signals." Differentiation; research in biological diversity 68(4-‐5): 175-‐185. Barsoum, I. B. and H. H. Yao (2010). "Fetal Leydig cells: progenitor cell maintenance and differentiation." Journal of andrology 31(1): 11-‐15. 187 Beckmann, J., S. Scheitza, et al. (2007). "Asymmetric cell division within the human hematopoietic stem and progenitor cell compartment: identification of asymmetrically segregating proteins." Blood 109(12): 5494-‐5501. Benedito, R., C. Roca, et al. (2009). "The notch ligands Dll4 and Jagged1 have opposing effects on angiogenesis." Cell 137(6): 1124-‐1135. Berdnik, D., T. Torok, et al. (2002). "The endocytic protein alpha-‐Adaptin is required for numb-‐mediated asymmetric cell division in Drosophila." Developmental cell 3(2): 221-‐231. Bertrand, J. Y., J. L. Cisson, et al. (2010). "Notch signaling distinguishes 2 waves of definitive hematopoiesis in the zebrafish embryo." Blood 115(14): 2777-‐2783. Bertrand, J. Y., N. C. Chi, et al. (2010). "Haematopoietic stem cells derive directly from aortic endothelium during development." Nature 464(7285): 108-‐111. Bertrand, N., D. S. Castro, et al. (2002). "Proneural genes and the specification of neural cell types." Nature reviews. Neuroscience 3(7): 517-‐530. Bigas, A. and L. Espinosa (2012). "Hematopoietic stem cells: to be or Notch to be." Blood 119(14): 3226-‐3235. Bigas, A., A. Robert-‐Moreno, et al. (2010). "The Notch pathway in the developing hematopoietic system." The International journal of developmental biology 54(6-‐7): 1175-‐1188. Bigas, A., A. Robert-‐Moreno, et al. (2010). "The Notch pathway in the developing hematopoietic system." Int J Dev Biol 54(6-‐7): 1175-‐1188. Bjornsson, J. M., N. Larsson, et al. (2003). "Reduced proliferative capacity of hematopoietic stem cells deficient in Hoxb3 and Hoxb4." Molecular and cellular biology 23(11): 3872-‐3883. Boisset, J. C. and C. Robin (2012). "On the origin of hematopoietic stem cells: progress and controversy." Stem cell research 8(1): 1-‐13. Boisset, J. C., W. van Cappellen, et al. (2010). "In vivo imaging of haematopoietic cells emerging from the mouse aortic endothelium." Nature 464(7285): 116-‐120. Bosco, E. E., J. C. Mulloy, et al. (2009). "Rac1 GTPase: a "Rac" of all trades." Cellular and molecular life sciences : CMLS 66(3): 370-‐374. 188 Brummendorf, T. H., W. Dragowska, et al. (1998). "Asymmetric cell divisions sustain long-‐term hematopoiesis from single-‐sorted human fetal liver cells." The Journal of experimental medicine 188(6): 1117-‐1124. Brun, A. C., J. M. Bjornsson, et al. (2004). "Hoxb4-‐deficient mice undergo normal hematopoietic development but exhibit a mild proliferation defect in hematopoietic stem cells." Blood 103(11): 4126-‐4133. Bryder, D. and S. E. Jacobsen (2000). "Interleukin-‐3 supports expansion of long-‐term multilineage repopulating activity after multiple stem cell divisions in vitro." Blood 96(5): 1748-‐1755. Buono, K. D., G. W. Robinson, et al. (2006). "The canonical Notch/RBP-‐J signaling pathway controls the balance of cell lineages in mammary epithelium during pregnancy." Developmental biology 293(2): 565-‐ 580. Burns, C. E., D. Traver, et al. (2005). "Hematopoietic stem cell fate is established by the Notch-‐Runx pathway." Genes & development 19(19): 2331-‐2342. Burns, C. E., D. Traver, et al. (2005). "Hematopoietic stem cell fate is established by the Notch-‐Runx pathway." Genes Dev 19(19): 2331-‐ 2342. Buske, C., M. Feuring-‐Buske, et al. (2002). "Deregulated expression of HOXB4 enhances the primitive growth activity of human hematopoietic cells." Blood 100(3): 862-‐868. Cadigan, K. M. and R. Nusse (1997). "Wnt signaling: a common theme in animal development." Genes & development 11(24): 3286-‐3305. Carre, A., L. Rachdi, et al. (2011). "Hes1 is required for appropriate morphogenesis and differentiation during mouse thyroid gland development." PloS one 6(2): e16752. Casey, L. M., Y. Lan, et al. (2006). "Jag2-‐Notch1 signaling regulates oral epithelial differentiation and palate development." Developmental dynamics : an official publication of the American Association of Anatomists 235(7): 1830-‐1844. Ciau-‐Uitz, A., M. Walmsley, et al. (2000). "Distinct origins of adult and embryonic blood in Xenopus." Cell 102(6): 787-‐796. Clements, W. K., A. D. Kim, et al. (2011). "A somitic Wnt16/Notch pathway specifies haematopoietic stem cells." Nature 474(7350): 220-‐224. 189 Cobas, M., A. Wilson, et al. (2004). "Beta-‐catenin is dispensable for hematopoiesis and lymphopoiesis." The Journal of experimental medicine 199(2): 221-‐229. Conlon, R. A., A. G. Reaume, et al. (1995). "Notch1 is required for the coordinate segmentation of somites." Development 121(5): 1533-‐ 1545. Cornell, M., D. A. Evans, et al. (1999). "The Drosophila melanogaster Suppressor of deltex gene, a regulator of the Notch receptor signaling pathway, is an E3 class ubiquitin ligase." Genetics 152(2): 567-‐576. Cotanche, D. A. and C. L. Kaiser (2010). "Hair cell fate decisions in cochlear development and regeneration." Hearing research 266(1-‐2): 18-‐25. Cotsarelis, G., P. Kaur, et al. (1999). "Epithelial stem cells in the skin: definition, markers, localization and functions." Experimental dermatology 8(1): 80-‐88. Cumano, A., J. C. Ferraz, et al. (2001). "Intraembryonic, but not yolk sac hematopoietic precursors, isolated before circulation, provide long-‐ term multilineage reconstitution." Immunity 15(3): 477-‐485. Curtis, D. J., M. A. Hall, et al. (2004). "SCL is required for normal function of short-‐term repopulating hematopoietic stem cells." Blood 103(9): 3342-‐3348. Chen, H., S. Fre, et al. (1998). "Epsin is an EH-‐domain-‐binding protein implicated in clathrin-‐mediated endocytosis." Nature 394(6695): 793-‐ 797. Chen, M. J., T. Yokomizo, et al. (2009). "Runx1 is required for the endothelial to haematopoietic cell transition but not thereafter." Nature 457(7231): 887-‐891. Cheng, P., Y. Nefedova, et al. (2007). "Regulation of dendritic-‐cell differentiation by bone marrow stroma via different Notch ligands." Blood 109(2): 507-‐515. Chu, J. and E. H. Bresnick (2004). "Evidence that C promoter-‐binding factor 1 binding is required for Notch-‐1-‐mediated repression of activator protein-‐1." J Biol Chem 279(13): 12337-‐12345. Davis, G. E. (2010). "Vascular balancing act: EGFL7 and Notch." Blood 116(26): 5791-‐5793. 190 de Bruijn, M. F., X. Ma, et al. (2002). "Hematopoietic stem cells localize to the endothelial cell layer in the midgestation mouse aorta." Immunity 16(5): 673-‐683. de Bruijn, M. F., M. C. Peeters, et al. (2000). "CFU-‐S(11) activity does not localize solely with the aorta in the aorta-‐gonad-‐mesonephros region." Blood 96(8): 2902-‐2904. de Bruijn, M. F., N. A. Speck, et al. (2000). "Definitive hematopoietic stem cells first develop within the major arterial regions of the mouse embryo." The EMBO journal 19(11): 2465-‐2474. De Camilli, P., H. Chen, et al. (2002). "The ENTH domain." FEBS letters 513(1): 11-‐18. de la Pompa, J. L., A. Wakeham, et al. (1997). "Conservation of the Notch signalling pathway in mammalian neurogenesis." Development 124(6): 1139-‐1148. De Strooper, B., P. Saftig, et al. (1998). "Deficiency of presenilin-‐1 inhibits the normal cleavage of amyloid precursor protein." Nature 391(6665): 387-‐390. Deblandre, G. A., E. C. Lai, et al. (2001). "Xenopus neuralized is a ubiquitin ligase that interacts with XDelta1 and regulates Notch signaling." Developmental cell 1(6): 795-‐806. Duarte, A., M. Hirashima, et al. (2004). "Dosage-‐sensitive requirement for mouse Dll4 in artery development." Genes & development 18(20): 2474-‐2478. Dunwoodie, S. L. (2009). "The role of Notch in patterning the human vertebral column." Current opinion in genetics & development 19(4): 329-‐337. Durand, C., C. Robin, et al. (2007). "Embryonic stromal clones reveal developmental regulators of definitive hematopoietic stem cells." Proceedings of the National Academy of Sciences of the United States of America 104(52): 20838-‐20843. Dyer, M. A., S. M. Farrington, et al. (2001). "Indian hedgehog activates hematopoiesis and vasculogenesis and can respecify prospective neurectodermal cell fate in the mouse embryo." Development 128(10): 1717-‐1730. 191 Dzierzak, E. and N. A. Speck (2008). "Of lineage and legacy: the development of mammalian hematopoietic stem cells." Nature immunology 9(2): 129-‐ 136. Dzierzak, E. and N. A. Speck (2008). "Of lineage and legacy: the development of mammalian hematopoietic stem cells." Nat Immunol 9(2): 129-‐136. Edbauer, D., E. Winkler, et al. (2003). "Reconstitution of gamma-‐secretase activity." Nature cell biology 5(5): 486-‐488. Ema, H. and H. Nakauchi (2000). "Expansion of hematopoietic stem cells in the developing liver of a mouse embryo." Blood 95(7): 2284-‐2288. Espinosa, L., S. Cathelin, et al. (2010). "The Notch/Hes1 pathway sustains NF-‐ kappaB activation through CYLD repression in T cell leukemia." Cancer Cell 18(3): 268-‐281. Estrach, S., C. A. Ambler, et al. (2006). "Jagged 1 is a beta-‐catenin target gene required for ectopic hair follicle formation in adult epidermis." Development 133(22): 4427-‐4438. Ferjentsik, Z., S. Hayashi, et al. (2009). "Notch is a critical component of the mouse somitogenesis oscillator and is essential for the formation of the somites." PLoS genetics 5(9): e1000662. Fernandez-‐Valdivia, R., H. Takeuchi, et al. (2011). "Regulation of mammalian Notch signaling and embryonic development by the protein O-‐ glucosyltransferase Rumi." Development 138(10): 1925-‐1934. Fischer, A., N. Schumacher, et al. (2004). "The Notch target genes Hey1 and Hey2 are required for embryonic vascular development." Genes & development 18(8): 901-‐911. Fischer, A., N. Schumacher, et al. (2004). "The Notch target genes Hey1 and Hey2 are required for embryonic vascular development." Genes Dev 18(8): 901-‐911. Francis, R., G. McGrath, et al. (2002). "aph-‐1 and pen-‐2 are required for Notch pathway signaling, gamma-‐secretase cleavage of betaAPP, and presenilin protein accumulation." Developmental cell 3(1): 85-‐97. Fre, S., E. Hannezo, et al. (2011). "Notch lineages and activity in intestinal stem cells determined by a new set of knock-‐in mice." PloS one 6(10): e25785. Fre, S., M. Huyghe, et al. (2005). "Notch signals control the fate of immature progenitor cells in the intestine." Nature 435(7044): 964-‐968. 192 Fryer, C. J., E. Lamar, et al. (2002). "Mastermind mediates chromatin-‐specific transcription and turnover of the Notch enhancer complex." Genes & development 16(11): 1397-‐1411. Garber, K. (2007). "Notch emerges as new cancer drug target." Journal of the National Cancer Institute 99(17): 1284-‐1285. Gasperowicz, M. and F. Otto (2008). "The notch signalling pathway in the development of the mouse placenta." Placenta 29(8): 651-‐659. Geisler, F., F. Nagl, et al. (2008). "Liver-‐specific inactivation of Notch2, but not Notch1, compromises intrahepatic bile duct development in mice." Hepatology 48(2): 607-‐616. Gekas, C., F. Dieterlen-‐Lievre, et al. (2005). "The placenta is a niche for hematopoietic stem cells." Developmental cell 8(3): 365-‐375. Gering, M. and R. Patient (2005). "Hedgehog signaling is required for adult blood stem cell formation in zebrafish embryos." Developmental cell 8(3): 389-‐400. Ghabrial, A. S. and M. A. Krasnow (2006). "Social interactions among epithelial cells during tracheal branching morphogenesis." Nature 441(7094): 746-‐749. Ghiaur, G., M. J. Ferkowicz, et al. (2008). "Rac1 is essential for intraembryonic hematopoiesis and for the initial seeding of fetal liver with definitive hematopoietic progenitor cells." Blood 111(7): 3313-‐3321. Gluzman-‐Poltorak, Z., T. Cohen, et al. (2000). "Neuropilin-‐2 is a receptor for the vascular endothelial growth factor (VEGF) forms VEGF-‐145 and VEGF-‐165 [corrected]." The Journal of biological chemistry 275(24): 18040-‐18045. Gluzman-‐Poltorak, Z., T. Cohen, et al. (2001). "Vascular endothelial growth factor receptor-‐1 and neuropilin-‐2 form complexes." The Journal of biological chemistry 276(22): 18688-‐18694. Godin, I. and A. Cumano (2002). "The hare and the tortoise: an embryonic haematopoietic race." Nature reviews. Immunology 2(8): 593-‐604. Gothert, J. R., S. E. Gustin, et al. (2005). "In vivo fate-‐tracing studies using the Scl stem cell enhancer: embryonic hematopoietic stem cells significantly contribute to adult hematopoiesis." Blood 105(7): 2724-‐ 2732. 193 Goutte, C., M. Tsunozaki, et al. (2002). "APH-‐1 is a multipass membrane protein essential for the Notch signaling pathway in Caenorhabditis elegans embryos." Proceedings of the National Academy of Sciences of the United States of America 99(2): 775-‐779. Gridley, T. (2010). "Notch signaling in the vasculature." Current topics in developmental biology 92: 277-‐309. Gu, Y., F. Chen, et al. (2003). "APH-‐1 interacts with mature and immature forms of presenilins and nicastrin and may play a role in maturation of presenilin.nicastrin complexes." The Journal of biological chemistry 278(9): 7374-‐7380. Haass, C. and H. Steiner (2002). "Alzheimer disease gamma-‐secretase: a complex story of GxGD-‐type presenilin proteases." Trends in cell biology 12(12): 556-‐562. Hall, M. A., D. J. Curtis, et al. (2003). "The critical regulator of embryonic hematopoiesis, SCL, is vital in the adult for megakaryopoiesis, erythropoiesis, and lineage choice in CFU-‐S12." Proceedings of the National Academy of Sciences of the United States of America 100(3): 992-‐997. Hayashi, S. and A. P. McMahon (2002). "Efficient recombination in diverse tissues by a tamoxifen-‐inducible form of Cre: a tool for temporally regulated gene activation/inactivation in the mouse." Dev Biol 244(2): 305-‐318. Hayashi, T., Y. Kageyama, et al. (2001). "Requirement of Notch 1 and its ligand jagged 2 expressions for spermatogenesis in rat and human testes." Journal of andrology 22(6): 999-‐1011. Heath, J. K. (2010). "Transcriptional networks and signaling pathways that govern vertebrate intestinal development." Current topics in developmental biology 90: 159-‐192. Hills, D., R. Gribi, et al. (2011). "Hoxb4-‐YFP reporter mouse model: a novel tool for tracking HSC development and studying the role of Hoxb4 in hematopoiesis." Blood 117(13): 3521-‐3528. Hirata, H., T. Ohtsuka, et al. (2000). "Generation of structurally and functionally distinct factors from the basic helix-‐loop-‐helix gene Hes3 by alternative first exons." The Journal of biological chemistry 275(25): 19083-‐19089. 194 Hirata, H., S. Yoshiura, et al. (2002). "Oscillatory expression of the bHLH factor Hes1 regulated by a negative feedback loop." Science 298(5594): 840-‐ 843. Hori, K., M. Fostier, et al. (2004). "Drosophila deltex mediates suppressor of Hairless-‐independent and late-‐endosomal activation of Notch signaling." Development 131(22): 5527-‐5537. Houssaint, E. (1981). "Differentiation of the mouse hepatic primordium. II. Extrinsic origin of the haemopoietic cell line." Cell differentiation 10(5): 243-‐252. Huang, E., K. Nocka, et al. (1990). "The hematopoietic growth factor KL is encoded by the Sl locus and is the ligand of the c-‐kit receptor, the gene product of the W locus." Cell 63(1): 225-‐233. Ilagan, M. X., S. Lim, et al. (2011). "Real-‐time imaging of notch activation with a luciferase complementation-‐based reporter." Science signaling 4(181): rs7. Imai, T., A. Tokunaga, et al. (2001). "The neural RNA-‐binding protein Musashi1 translationally regulates mammalian numb gene expression by interacting with its mRNA." Molecular and cellular biology 21(12): 3888-‐3900. Imayoshi, I., T. Shimogori, et al. (2008). "Hes genes and neurogenin regulate non-‐neural versus neural fate specification in the dorsal telencephalic midline." Development 135(15): 2531-‐2541. Iso, T., G. Chung, et al. (2002). "HERP1 is a cell type-‐specific primary target of Notch." The Journal of biological chemistry 277(8): 6598-‐6607. Iso, T., V. Sartorelli, et al. (2001). "HERP, a new primary target of Notch regulated by ligand binding." Molecular and cellular biology 21(17): 6071-‐6079. Iso, T., V. Sartorelli, et al. (2001). "HERP, a novel heterodimer partner of HES/E(spl) in Notch signaling." Molecular and cellular biology 21(17): 6080-‐6089. Itoh, M., C. H. Kim, et al. (2003). "Mind bomb is a ubiquitin ligase that is essential for efficient activation of Notch signaling by Delta." Developmental cell 4(1): 67-‐82. Ivanovs, A., S. Rybtsov, et al. (2011). "Highly potent human hematopoietic stem cells first emerge in the intraembryonic aorta-‐gonad-‐ 195 mesonephros region." The Journal of experimental medicine 208(12): 2417-‐2427. Jain, R., S. Rentschler, et al. (2010). "Notch and cardiac outflow tract development." Annals of the New York Academy of Sciences 1188: 184-‐190. Jarriault, S., C. Brou, et al. (1995). "Signalling downstream of activated mammalian Notch." Nature 377(6547): 355-‐358. Jeannet, G., M. Scheller, et al. (2008). "Long-‐term, multilineage hematopoiesis occurs in the combined absence of beta-‐catenin and gamma-‐catenin." Blood 111(1): 142-‐149. Jehn, B. M., I. Dittert, et al. (2002). "c-‐Cbl binding and ubiquitin-‐dependent lysosomal degradation of membrane-‐associated Notch1." The Journal of biological chemistry 277(10): 8033-‐8040. Jia, J., M. Lin, et al. (2007). "The Notch signaling pathway controls the size of the ocular lens by directly suppressing p57Kip2 expression." Molecular and cellular biology 27(20): 7236-‐7247. Jiang, R., Y. Lan, et al. (1998). "Defects in limb, craniofacial, and thymic development in Jagged2 mutant mice." Genes & development 12(7): 1046-‐1057. Johnson, G. R. and M. A. Moore (1975). "Role of stem cell migration in initiation of mouse foetal liver haemopoiesis." Nature 258(5537): 726-‐728. Kageyama, R., Y. Niwa, et al. (2010). "Ultradian oscillations in Notch signaling regulate dynamic biological events." Current topics in developmental biology 92: 311-‐331. Kageyama, R., T. Ohtsuka, et al. (2007). "The Hes gene family: repressors and oscillators that orchestrate embryogenesis." Development 134(7): 1243-‐1251. Kageyama, R., T. Ohtsuka, et al. (2008). "Dynamic Notch signaling in neural progenitor cells and a revised view of lateral inhibition." Nature neuroscience 11(11): 1247-‐1251. Kanemura, Y., K. Mori, et al. (2001). "Musashi1, an evolutionarily conserved neural RNA-‐binding protein, is a versatile marker of human glioma cells in determining their cellular origin, malignancy, and proliferative activity." Differentiation; research in biological diversity 68(2-‐3): 141-‐ 152. 196 Kao, H. Y., P. Ordentlich, et al. (1998). "A histone deacetylase corepressor complex regulates the Notch signal transduction pathway." Genes & development 12(15): 2269-‐2277. Karanu, F. N., B. Murdoch, et al. (2001). "Human homologues of Delta-‐1 and Delta-‐4 function as mitogenic regulators of primitive human hematopoietic cells." Blood 97(7): 1960-‐1967. Kidd, S., T. J. Lockett, et al. (1983). "The Notch locus of Drosophila melanogaster." Cell 34(2): 421-‐433. Kiel, M. J., O. H. Yilmaz, et al. (2005). "SLAM family receptors distinguish hematopoietic stem and progenitor cells and reveal endothelial niches for stem cells." Cell 121(7): 1109-‐1121. Kiernan, A. E., J. Xu, et al. (2006). "The Notch ligand JAG1 is required for sensory progenitor development in the mammalian inner ear." PLoS genetics 2(1): e4. Kim, D., Y. K. Kwon, et al. (2008). "The biphasic behavior of incoherent feed-‐ forward loops in biomolecular regulatory networks." Bioessays 30(11-‐ 12): 1204-‐1211. Kim, T. H., B. M. Kim, et al. (2011). "Endodermal Hedgehog signals modulate Notch pathway activity in the developing digestive tract mesenchyme." Development 138(15): 3225-‐3233. Kim, T. H. and R. A. Shivdasani (2011). "Genetic evidence that intestinal Notch functions vary regionally and operate through a common mechanism of Math1 repression." The Journal of biological chemistry 286(13): 11427-‐11433. Kim, W., Y. K. Shin, et al. (2010). "Notch signaling in pancreatic endocrine cell and diabetes." Biochemical and biophysical research communications 392(3): 247-‐251. Kirstetter, P., K. Anderson, et al. (2006). "Activation of the canonical Wnt pathway leads to loss of hematopoietic stem cell repopulation and multilineage differentiation block." Nature immunology 7(10): 1048-‐ 1056. Kobayashi-‐Osaki, M., O. Ohneda, et al. (2005). "GATA motifs regulate early hematopoietic lineage-‐specific expression of the Gata2 gene." Mol Cell Biol 25(16): 7005-‐7020. 197 Koch, U., T. A. Lacombe, et al. (2001). "Subversion of the T/B lineage decision in the thymus by lunatic fringe-‐mediated inhibition of Notch-‐1." Immunity 15(2): 225-‐236. Koch, U., A. Wilson, et al. (2008). "Simultaneous loss of beta-‐ and gamma-‐ catenin does not perturb hematopoiesis or lymphopoiesis." Blood 111(1): 160-‐164. Koga, S., N. Yamaguchi, et al. (2007). "Cell-‐cycle-‐dependent oscillation of GATA2 expression in hematopoietic cells." Blood 109(10): 4200-‐4208. Komatsu, H., M. Y. Chao, et al. (2008). "OSM-‐11 facilitates LIN-‐12 Notch signaling during Caenorhabditis elegans vulval development." PLoS biology 6(8): e196. Kopan, R. and M. X. Ilagan (2009). "The canonical Notch signaling pathway: unfolding the activation mechanism." Cell 137(2): 216-‐233. Krebs, L. T., J. R. Shutter, et al. (2004). "Haploinsufficient lethality and formation of arteriovenous malformations in Notch pathway mutants." Genes & development 18(20): 2469-‐2473. Krebs, L. T., Y. Xue, et al. (2000). "Notch signaling is essential for vascular morphogenesis in mice." Genes & development 14(11): 1343-‐1352. Krejci, A., F. Bernard, et al. (2009). "Direct response to Notch activation: signaling crosstalk and incoherent logic." Sci Signal 2(55): ra1. Krejci, A., F. Bernard, et al. (2009). "Direct response to Notch activation: signaling crosstalk and incoherent logic." Science signaling 2(55): ra1. Kumano, K., S. Chiba, et al. (2003). "Notch1 but not Notch2 is essential for generating hematopoietic stem cells from endothelial cells." Immunity 18(5): 699-‐711. Kumaravelu, P., L. Hook, et al. (2002). "Quantitative developmental anatomy of definitive haematopoietic stem cells/long-‐term repopulating units (HSC/RUs): role of the aorta-‐gonad-‐mesonephros (AGM) region and the yolk sac in colonisation of the mouse embryonic liver." Development 129(21): 4891-‐4899. Kunisato, A., S. Chiba, et al. (2003). "HES-‐1 preserves purified hematopoietic stem cells ex vivo and accumulates side population cells in vivo." Blood 101(5): 1777-‐1783. 198 Kyba, M., R. C. Perlingeiro, et al. (2002). "HoxB4 confers definitive lymphoid-‐ myeloid engraftment potential on embryonic stem cell and yolk sac hematopoietic progenitors." Cell 109(1): 29-‐37. Lai, E. C., G. A. Deblandre, et al. (2001). "Drosophila neuralized is a ubiquitin ligase that promotes the internalization and degradation of delta." Developmental cell 1(6): 783-‐794. Laiosa, C. V., M. Stadtfeld, et al. (2006). "Determinants of lymphoid-‐myeloid lineage diversification." Annual review of immunology 24: 705-‐738. Lantz, C. S., J. Boesiger, et al. (1998). "Role for interleukin-‐3 in mast-‐cell and basophil development and in immunity to parasites." Nature 392(6671): 90-‐93. Larsson, J. and S. Karlsson (2005). "The role of Smad signaling in hematopoiesis." Oncogene 24(37): 5676-‐5692. Lawson, N. D., A. M. Vogel, et al. (2002). "sonic hedgehog and vascular endothelial growth factor act upstream of the Notch pathway during arterial endothelial differentiation." Developmental cell 3(1): 127-‐136. Le, T. T., K. W. Conley, et al. (2009). "Jagged 1 is necessary for normal mouse lens formation." Developmental biology 328(1): 118-‐126. Leary, A. G., L. C. Strauss, et al. (1985). "Disparate differentiation in hemopoietic colonies derived from human paired progenitors." Blood 66(2): 327-‐332. Lee, S. Y., K. Kumano, et al. (2005). "Mutations of the Notch1 gene in T-‐cell acute lymphoblastic leukemia: analysis in adults and children." Leukemia : official journal of the Leukemia Society of America, Leukemia Research Fund, U.K 19(10): 1841-‐1843. Li, L., I. D. Krantz, et al. (1997). "Alagille syndrome is caused by mutations in human Jagged1, which encodes a ligand for Notch1." Nature genetics 16(3): 243-‐251. Ling, K. W., K. Ottersbach, et al. (2004). "GATA-‐2 plays two functionally distinct roles during the ontogeny of hematopoietic stem cells." The Journal of experimental medicine 200(7): 871-‐882. Ling, K. W., K. Ottersbach, et al. (2004). "GATA-‐2 plays two functionally distinct roles during the ontogeny of hematopoietic stem cells." J Exp Med 200(7): 871-‐882. 199 Loomes, K. M., S. A. Stevens, et al. (2007). "Dll3 and Notch1 genetic interactions model axial segmental and craniofacial malformations of human birth defects." Developmental dynamics : an official publication of the American Association of Anatomists 236(10): 2943-‐2951. Lozier, J., B. McCright, et al. (2008). "Notch signaling regulates bile duct morphogenesis in mice." PloS one 3(3): e1851. Lubman, O. Y., S. V. Korolev, et al. (2004). "Anchoring notch genetics and biochemistry; structural analysis of the ankyrin domain sheds light on existing data." Molecular cell 13(5): 619-‐626. Luis, T. C., B. A. Naber, et al. (2011). "Canonical wnt signaling regulates hematopoiesis in a dosage-‐dependent fashion." Cell stem cell 9(4): 345-‐ 356. Luis, T. C., F. Weerkamp, et al. (2009). "Wnt3a deficiency irreversibly impairs hematopoietic stem cell self-‐renewal and leads to defects in progenitor cell differentiation." Blood 113(3): 546-‐554. MacGrogan, D., M. Nus, et al. (2010). "Notch signaling in cardiac development and disease." Current topics in developmental biology 92: 333-‐365. Magli, M. C., N. N. Iscove, et al. (1982). "Transient nature of early haematopoietic spleen colonies." Nature 295(5849): 527-‐529. Maillard, I., U. Koch, et al. (2008). "Canonical notch signaling is dispensable for the maintenance of adult hematopoietic stem cells." Cell stem cell 2(4): 356-‐366. Malhotra, S. and P. W. Kincade (2009). "Canonical Wnt pathway signaling suppresses VCAM-‐1 expression by marrow stromal and hematopoietic cells." Experimental hematology 37(1): 19-‐30. Mancini, S. J., N. Mantei, et al. (2005). "Jagged1-‐dependent Notch signaling is dispensable for hematopoietic stem cell self-‐renewal and differentiation." Blood 105(6): 2340-‐2342. Mangan, S. and U. Alon (2003). "Structure and function of the feed-‐forward loop network motif." Proceedings of the National Academy of Sciences of the United States of America 100(21): 11980-‐11985. Mangan, S. and U. Alon (2003). "Structure and function of the feed-‐forward loop network motif." Proc Natl Acad Sci U S A 100(21): 11980-‐11985. Mansour, M. R., D. C. Linch, et al. (2006). "High incidence of Notch-‐1 mutations in adult patients with T-‐cell acute lymphoblastic leukemia." Leukemia : 200 official journal of the Leukemia Society of America, Leukemia Research Fund, U.K 20(3): 537-‐539. Mao, S., R. C. Frank, et al. (1999). "Functional and physical interactions between AML1 proteins and an ETS protein, MEF: implications for the pathogenesis of t(8;21)-‐positive leukemias." Molecular and cellular biology 19(5): 3635-‐3644. Matsuda, Y., Y. Wakamatsu, et al. (2005). "Notch signaling functions as a binary switch for the determination of glandular and luminal fates of endodermal epithelium during chicken stomach development." Development 132(12): 2783-‐2793. Matsuno, K., R. J. Diederich, et al. (1995). "Deltex acts as a positive regulator of Notch signaling through interactions with the Notch ankyrin repeats." Development 121(8): 2633-‐2644. McGill, M. A., S. E. Dho, et al. (2009). "Numb regulates post-‐endocytic trafficking and degradation of Notch1." The Journal of biological chemistry 284(39): 26427-‐26438. McGlinn, E., K. L. van Bueren, et al. (2005). "Pax9 and Jagged1 act downstream of Gli3 in vertebrate limb development." Mechanisms of development 122(11): 1218-‐1233. Medvinsky, A. and E. Dzierzak (1996). "Definitive hematopoiesis is autonomously initiated by the AGM region." Cell 86(6): 897-‐906. Medvinsky, A. and E. Dzierzak (1999). "Development of the hematopoietic stem cell: can we describe it?" Blood 94(10): 3613-‐3614. Medvinsky, A. and E. Dzierzak (1999). "Embryonic beginnings of definitive hematopoietic stem cells." Blood 94(10): 3613-‐3614. Meloty-‐Kapella, L., B. Shergill, et al. (2012). "Notch Ligand Endocytosis Generates Mechanical Pulling Force Dependent on Dynamin, Epsins, and Actin." Developmental cell. Mikkola, H. K., J. Klintman, et al. (2003). "Haematopoietic stem cells retain long-‐term repopulating activity and multipotency in the absence of stem-‐cell leukaemia SCL/tal-‐1 gene." Nature 421(6922): 547-‐551. Minegishi, N., J. Ohta, et al. (1998). "Alternative promoters regulate transcription of the mouse GATA-‐2 gene." The Journal of biological chemistry 273(6): 3625-‐3634. 201 Minegishi, N., J. Ohta, et al. (1998). "Alternative promoters regulate transcription of the mouse GATA-‐2 gene." J Biol Chem 273(6): 3625-‐ 3634. Minegishi, N., J. Ohta, et al. (1999). "The mouse GATA-‐2 gene is expressed in the para-‐aortic splanchnopleura and aorta-‐gonads and mesonephros region." Blood 93(12): 4196-‐4207. Minegishi, N., N. Suzuki, et al. (2003). "Expression and domain-‐specific function of GATA-‐2 during differentiation of the hematopoietic precursor cells in midgestation mouse embryos." Blood 102(3): 896-‐ 905. Mitsiadis, T. A., L. Regaudiat, et al. (2005). "Role of the Notch signalling pathway in tooth morphogenesis." Archives of oral biology 50(2): 137-‐ 140. Mohtashami, M., D. K. Shah, et al. (2010). "Direct comparison of Dll1-‐ and Dll4-‐ mediated Notch activation levels shows differential lymphomyeloid lineage commitment outcomes." Journal of immunology 185(2): 867-‐ 876. Monahan, P., S. Rybak, et al. (2009). "The notch target gene HES1 regulates cell cycle inhibitor expression in the developing pituitary." Endocrinology 150(9): 4386-‐4394. Morel, V., M. Lecourtois, et al. (2001). "Transcriptional repression by suppressor of hairless involves the binding of a hairless-‐dCtBP complex in Drosophila." Current biology : CB 11(10): 789-‐792. Morgan, T. H. (1919). The physical basis of heredity, by Thomas Hunt Morgan ... 117 illustrations. Philadelphia, J.B. Lippincott. Mucenski, M. L., K. McLain, et al. (1991). "A functional c-‐myb gene is required for normal murine fetal hepatic hematopoiesis." Cell 65(4): 677-‐689. Mukouyama, Y., N. Chiba, et al. (1999). "Hematopoietic cells in cultures of the murine embryonic aorta-‐gonad-‐mesonephros region are induced by c-‐ Myb." Current biology : CB 9(15): 833-‐836. Muller, A. M., A. Medvinsky, et al. (1994). "Development of hematopoietic stem cell activity in the mouse embryo." Immunity 1(4): 291-‐301. Nadarajan, S., J. A. Govindan, et al. (2009). "MSP and GLP-‐1/Notch signaling coordinately regulate actomyosin-‐dependent cytoplasmic streaming and oocyte growth in C. elegans." Development 136(13): 2223-‐2234. 202 Nagai, T., H. Harigae, et al. (1994). "Transcription factor GATA-‐2 is expressed in erythroid, early myeloid, and CD34+ human leukemia-‐derived cell lines." Blood 84(4): 1074-‐1084. Nagel, A. C., A. Krejci, et al. (2005). "Hairless-‐mediated repression of notch target genes requires the combined activity of Groucho and CtBP corepressors." Molecular and cellular biology 25(23): 10433-‐10441. Nakagawa, M., M. Ichikawa, et al. (2006). "AML1/Runx1 rescues Notch1-‐null mutation-‐induced deficiency of para-‐aortic splanchnopleural hematopoiesis." Blood 108(10): 3329-‐3334. Nakagawa, T., S. Y. Lee, et al. (2009). "Induction of chondrogenesis from human embryonic stem cells without embryoid body formation by bone morphogenetic protein 7 and transforming growth factor beta1." Arthritis and rheumatism 60(12): 3686-‐3692. Narula, J., A. M. Smith, et al. (2010). "Modeling reveals bistability and low-‐pass filtering in the network module determining blood stem cell fate." PLoS Comput Biol 6(5): e1000771. Nichols, J. T., A. Miyamoto, et al. (2007). "DSL ligand endocytosis physically dissociates Notch1 heterodimers before activating proteolysis can occur." The Journal of cell biology 176(4): 445-‐458. Nishinakamura, R., N. Nakayama, et al. (1995). "Mice deficient for the IL-‐ 3/GM-‐CSF/IL-‐5 beta c receptor exhibit lung pathology and impaired immune response, while beta IL3 receptor-‐deficient mice are normal." Immunity 2(3): 211-‐222. North, T., T. L. Gu, et al. (1999). "Cbfa2 is required for the formation of intra-‐ aortic hematopoietic clusters." Development 126(11): 2563-‐2575. North, T. E., W. Goessling, et al. (2009). "Hematopoietic stem cell development is dependent on blood flow." Cell 137(4): 736-‐748. Nottingham, W. T., A. Jarratt, et al. (2007). "Runx1-‐mediated hematopoietic stem-‐cell emergence is controlled by a Gata/Ets/SCL-‐regulated enhancer." Blood 110(13): 4188-‐4197. Ntziachristos, P., A. Tsirigos, et al. (2012). "Genetic inactivation of the polycomb repressive complex 2 in T cell acute lymphoblastic leukemia." Nature medicine 18(2): 298-‐301. 203 Ohata, S., R. Aoki, et al. (2011). "Dual roles of Notch in regulation of apically restricted mitosis and apicobasal polarity of neuroepithelial cells." Neuron 69(2): 215-‐230. Ohtsuka, T., M. Ishibashi, et al. (1999). "Hes1 and Hes5 as notch effectors in mammalian neuronal differentiation." The EMBO journal 18(8): 2196-‐ 2207. Ohtsuka, T., M. Ishibashi, et al. (1999). "Hes1 and Hes5 as notch effectors in mammalian neuronal differentiation." Embo J 18(8): 2196-‐2207. Oka, C., T. Nakano, et al. (1995). "Disruption of the mouse RBP-‐J kappa gene results in early embryonic death." Development 121(10): 3291-‐3301. Okada, H., T. Watanabe, et al. (1998). "AML1(-‐/-‐) embryos do not express certain hematopoiesis-‐related gene transcripts including those of the PU.1 gene." Oncogene 17(18): 2287-‐2293. Okajima, T. and K. D. Irvine (2002). "Regulation of notch signaling by o-‐linked fucose." Cell 111(6): 893-‐904. Okamura, Y. and Y. Saga (2008). "Pofut1 is required for the proper localization of the Notch receptor during mouse development." Mechanisms of development 125(8): 663-‐673. Okochi, M., H. Steiner, et al. (2002). "Presenilins mediate a dual intramembranous gamma-‐secretase cleavage of Notch-‐1." The EMBO journal 21(20): 5408-‐5416. Okuda, T., J. van Deursen, et al. (1996). "AML1, the target of multiple chromosomal translocations in human leukemia, is essential for normal fetal liver hematopoiesis." Cell 84(2): 321-‐330. Orkin, S. H. and L. I. Zon (2008). "Hematopoiesis: an evolving paradigm for stem cell biology." Cell 132(4): 631-‐644. Ottersbach, K. and E. Dzierzak (2005). "The murine placenta contains hematopoietic stem cells within the vascular labyrinth region." Developmental cell 8(3): 377-‐387. Palis, J., S. Robertson, et al. (1999). "Development of erythroid and myeloid progenitors in the yolk sac and embryo proper of the mouse." Development 126(22): 5073-‐5084. Pan, X., N. Minegishi, et al. (2000). "Identification of human GATA-‐2 gene distal IS exon and its expression in hematopoietic stem cell fractions." Journal of biochemistry 127(1): 105-‐112. 204 Pan, Y., Z. Liu, et al. (2005). "Notch1 and 2 cooperate in limb ectoderm to receive an early Jagged2 signal regulating interdigital apoptosis." Developmental biology 286(2): 472-‐482. Pardanaud, L., D. Luton, et al. (1996). "Two distinct endothelial lineages in ontogeny, one of them related to hemopoiesis." Development 122(5): 1363-‐1371. Parks, A. L., S. S. Huppert, et al. (1997). "The dynamics of neurogenic signalling underlying bristle development in Drosophila melanogaster." Mechanisms of development 63(1): 61-‐74. Parks, A. L., K. M. Klueg, et al. (2000). "Ligand endocytosis drives receptor dissociation and activation in the Notch pathway." Development 127(7): 1373-‐1385. Pavlopoulos, E., C. Pitsouli, et al. (2001). "neuralized Encodes a peripheral membrane protein involved in delta signaling and endocytosis." Developmental cell 1(6): 807-‐816. Peeters, M., K. Ottersbach, et al. (2009). "Ventral embryonic tissues and Hedgehog proteins induce early AGM hematopoietic stem cell development." Development 136(15): 2613-‐2621. Pellegrinet, L., V. Rodilla, et al. (2011). "Dll1-‐ and dll4-‐mediated notch signaling are required for homeostasis of intestinal stem cells." Gastroenterology 140(4): 1230-‐1240 e1231-‐1237. Peters, S. O., E. L. Kittler, et al. (1996). "Ex vivo expansion of murine marrow cells with interleukin-‐3 (IL-‐3), IL-‐6, IL-‐11, and stem cell factor leads to impaired engraftment in irradiated hosts." Blood 87(1): 30-‐37. Pimanda, J. E., K. Ottersbach, et al. (2007). "Gata2, Fli1, and Scl form a recursively wired gene-‐regulatory circuit during early hematopoietic development." Proceedings of the National Academy of Sciences of the United States of America 104(45): 17692-‐17697. Polo, S., S. Sigismund, et al. (2002). "A single motif responsible for ubiquitin recognition and monoubiquitination in endocytic proteins." Nature 416(6879): 451-‐455. Potocnik, A. J., C. Brakebusch, et al. (2000). "Fetal and adult hematopoietic stem cells require beta1 integrin function for colonizing fetal liver, spleen, and bone marrow." Immunity 12(6): 653-‐663. 205 Raetzman, L. T., S. A. Ross, et al. (2004). "Developmental regulation of Notch signaling genes in the embryonic pituitary: Prop1 deficiency affects Notch2 expression." Developmental biology 265(2): 329-‐340. Rana, N. A. and R. S. Haltiwanger (2011). "Fringe benefits: functional and structural impacts of O-‐glycosylation on the extracellular domain of Notch receptors." Current opinion in structural biology 21(5): 583-‐589. Rebay, I., R. J. Fleming, et al. (1991). "Specific EGF repeats of Notch mediate interactions with Delta and Serrate: implications for Notch as a multifunctional receptor." Cell 67(4): 687-‐699. Reya, T., A. W. Duncan, et al. (2003). "A role for Wnt signalling in self-‐renewal of haematopoietic stem cells." Nature 423(6938): 409-‐414. Rhodes, K. E., C. Gekas, et al. (2008). "The emergence of hematopoietic stem cells is initiated in the placental vasculature in the absence of circulation." Cell stem cell 2(3): 252-‐263. Robb, L., C. C. Drinkwater, et al. (1995). "Hematopoietic and lung abnormalities in mice with a null mutation of the common beta subunit of the receptors for granulocyte-‐macrophage colony-‐stimulating factor and interleukins 3 and 5." Proceedings of the National Academy of Sciences of the United States of America 92(21): 9565-‐9569. Robb, L., I. Lyons, et al. (1995). "Absence of yolk sac hematopoiesis from mice with a targeted disruption of the scl gene." Proceedings of the National Academy of Sciences of the United States of America 92(15): 7075-‐ 7079. Robert-‐Moreno, A., L. Espinosa, et al. (2005). "RBPjkappa-‐dependent Notch function regulates Gata2 and is essential for the formation of intra-‐ embryonic hematopoietic cells." Development 132(5): 1117-‐1126. Robert-‐Moreno, A., L. Espinosa, et al. (2007). "The notch pathway positively regulates programmed cell death during erythroid differentiation." Leukemia 21(7): 1496-‐1503. Robert-‐Moreno, A., J. Guiu, et al. (2008). "Impaired embryonic haematopoiesis yet normal arterial development in the absence of the Notch ligand Jagged1." The EMBO journal 27(13): 1886-‐1895. Robert-‐Moreno, A., J. Guiu, et al. (2008). "Impaired embryonic haematopoiesis yet normal arterial development in the absence of the Notch ligand Jagged1." EMBO J 27(13): 1886-‐1895. 206 Robin, C., K. Bollerot, et al. (2009). "Human placenta is a potent hematopoietic niche containing hematopoietic stem and progenitor cells throughout development." Cell stem cell 5(4): 385-‐395. Robin, C., K. Ottersbach, et al. (2011). "CD41 is developmentally regulated and differentially expressed on mouse hematopoietic stem cells." Blood 117(19): 5088-‐5091. Robin, C., K. Ottersbach, et al. (2006). "An unexpected role for IL-‐3 in the embryonic development of hematopoietic stem cells." Developmental cell 11(2): 171-‐180. Rodilla, V., A. Villanueva, et al. (2009). "Jagged1 is the pathological link between Wnt and Notch pathways in colorectal cancer." Proceedings of the National Academy of Sciences of the United States of America 106(15): 6315-‐6320. Rodilla, V., A. Villanueva, et al. (2009). "Jagged1 is the pathological link between Wnt and Notch pathways in colorectal cancer." Proc Natl Acad Sci U S A 106(15): 6315-‐6320. Rodrigues, N. P., V. Janzen, et al. (2005). "Haploinsufficiency of GATA-‐2 perturbs adult hematopoietic stem-‐cell homeostasis." Blood 106(2): 477-‐484. Ross, S. E., M. E. Greenberg, et al. (2003). "Basic helix-‐loop-‐helix factors in cortical development." Neuron 39(1): 13-‐25. Rowan, S., K. W. Conley, et al. (2008). "Notch signaling regulates growth and differentiation in the mammalian lens." Developmental biology 321(1): 111-‐122. Ruiz-‐Herguido, C., J. Guiu, et al. (2012). "Hematopoietic stem cell development requires transient Wnt/beta-‐catenin activity." The Journal of experimental medicine 209(8): 1457-‐1468. Rybtsov, S., M. Sobiesiak, et al. (2011). "Hierarchical organization and early hematopoietic specification of the developing HSC lineage in the AGM region." The Journal of experimental medicine 208(6): 1305-‐1315. Samokhvalov, I. M., N. I. Samokhvalova, et al. (2007). "Cell tracing shows the contribution of the yolk sac to adult haematopoiesis." Nature 446(7139): 1056-‐1061. 207 Sanchez, M. J., A. Holmes, et al. (1996). "Characterization of the first definitive hematopoietic stem cells in the AGM and liver of the mouse embryo." Immunity 5(6): 513-‐525. Sangiorgi, E. and M. R. Capecchi (2008). "Bmi1 is expressed in vivo in intestinal stem cells." Nature genetics 40(7): 915-‐920. Sasaki, T., C. Mizuochi, et al. (2010). "Regulation of hematopoietic cell clusters in the placental niche through SCF/Kit signaling in embryonic mouse." Development 137(23): 3941-‐3952. Sato, T., T. S. Diehl, et al. (2007). "Active gamma-‐secretase complexes contain only one of each component." The Journal of biological chemistry 282(47): 33985-‐33993. Sato, T., R. G. Vries, et al. (2009). "Single Lgr5 stem cells build crypt-‐villus structures in vitro without a mesenchymal niche." Nature 459(7244): 262-‐265. Scheller, M., J. Huelsken, et al. (2006). "Hematopoietic stem cell and multilineage defects generated by constitutive beta-‐catenin activation." Nature immunology 7(10): 1037-‐1047. Schouwey, K. and F. Beermann (2008). "The Notch pathway: hair graying and pigment cell homeostasis." Histology and histopathology 23(5): 609-‐ 619. Schulz, C., E. Gomez Perdiguero, et al. (2012). "A lineage of myeloid cells independent of Myb and hematopoietic stem cells." Science 336(6077): 86-‐90. Serth, K., K. Schuster-‐Gossler, et al. (2003). "Transcriptional oscillation of lunatic fringe is essential for somitogenesis." Genes & development 17(7): 912-‐925. Shalaby, F., J. Ho, et al. (1997). "A requirement for Flk1 in primitive and definitive hematopoiesis and vasculogenesis." Cell 89(6): 981-‐990. Shalaby, F., J. Rossant, et al. (1995). "Failure of blood-‐island formation and vasculogenesis in Flk-‐1-‐deficient mice." Nature 376(6535): 62-‐66. Shi, S. and P. Stanley (2003). "Protein O-‐fucosyltransferase 1 is an essential component of Notch signaling pathways." Proceedings of the National Academy of Sciences of the United States of America 100(9): 5234-‐ 5239. 208 Shibata, K., H. Yamada, et al. (2011). "Notch-‐Hes1 pathway is required for the development of IL-‐17-‐producing gammadelta T cells." Blood 118(3): 586-‐593. Shimizu, K., S. Chiba, et al. (2000). "Binding of Delta1, Jagged1, and Jagged2 to Notch2 rapidly induces cleavage, nuclear translocation, and hyperphosphorylation of Notch2." Molecular and cellular biology 20(18): 6913-‐6922. Shimizu, K., S. Chiba, et al. (2000). "Physical interaction of Delta1, Jagged1, and Jagged2 with Notch1 and Notch3 receptors." Biochemical and biophysical research communications 276(1): 385-‐389. Shivdasani, R. A., E. L. Mayer, et al. (1995). "Absence of blood formation in mice lacking the T-‐cell leukaemia oncoprotein tal-‐1/SCL." Nature 373(6513): 432-‐434. Shutter, J. R., S. Scully, et al. (2000). "Dll4, a novel Notch ligand expressed in arterial endothelium." Genes & development 14(11): 1313-‐1318. Soker, S., S. Takashima, et al. (1998). "Neuropilin-‐1 is expressed by endothelial and tumor cells as an isoform-‐specific receptor for vascular endothelial growth factor." Cell 92(6): 735-‐745. Sorensen, I., R. H. Adams, et al. (2009). "DLL1-‐mediated Notch activation regulates endothelial identity in mouse fetal arteries." Blood 113(22): 5680-‐5688. Sorensen, L. K., B. S. Brooke, et al. (2003). "Loss of distinct arterial and venous boundaries in mice lacking endoglin, a vascular-‐specific TGFbeta coreceptor." Developmental biology 261(1): 235-‐250. Steiner, H., E. Winkler, et al. (2002). "PEN-‐2 is an integral component of the gamma-‐secretase complex required for coordinated expression of presenilin and nicastrin." The Journal of biological chemistry 277(42): 39062-‐39065. Storck, S., F. Delbos, et al. (2005). "Normal immune system development in mice lacking the Deltex-‐1 RING finger domain." Molecular and cellular biology 25(4): 1437-‐1445. Suda, J., T. Suda, et al. (1984). "Analysis of differentiation of mouse hemopoietic stem cells in culture by sequential replating of paired progenitors." Blood 64(2): 393-‐399. 209 Suzuki, N., O. Ohneda, et al. (2006). "Combinatorial Gata2 and Sca1 expression defines hematopoietic stem cells in the bone marrow niche." Proceedings of the National Academy of Sciences of the United States of America 103(7): 2202-‐2207. Suzuki, T., Y. Yokoyama, et al. (2006). "Highly efficient ex vivo expansion of human hematopoietic stem cells using Delta1-‐Fc chimeric protein." Stem cells 24(11): 2456-‐2465. Takeda, S., T. Shimizu, et al. (1997). "Interactions between c-‐kit and stem cell factor are not required for B-‐cell development in vivo." Blood 89(2): 518-‐525. Takeuchi, M., T. Sekiguchi, et al. (2002). "Cultivation of aorta-‐gonad-‐ mesonephros-‐derived hematopoietic stem cells in the fetal liver microenvironment amplifies long-‐term repopulating activity and enhances engraftment to the bone marrow." Blood 99(4): 1190-‐1196. Takeyama, K., R. C. Aguiar, et al. (2003). "The BAL-‐binding protein BBAP and related Deltex family members exhibit ubiquitin-‐protein isopeptide ligase activity." The Journal of biological chemistry 278(24): 21930-‐ 21937. Tan, J. B., K. Xu, et al. (2009). "Lunatic and manic fringe cooperatively enhance marginal zone B cell precursor competition for delta-‐like 1 in splenic endothelial niches." Immunity 30(2): 254-‐263. Tang, H., J. Brennan, et al. (2008). "Notch signaling maintains Leydig progenitor cells in the mouse testis." Development 135(22): 3745-‐ 3753. Tanigaki, K. and T. Honjo (2010). "Two opposing roles of RBP-‐J in Notch signaling." Current topics in developmental biology 92: 231-‐252. Taoudi, S., T. Bee, et al. (2011). "ERG dependence distinguishes developmental control of hematopoietic stem cell maintenance from hematopoietic specification." Genes & development 25(3): 251-‐262. Taoudi, S., C. Gonneau, et al. (2008). "Extensive hematopoietic stem cell generation in the AGM region via maturation of VE-‐cadherin+CD45+ pre-‐definitive HSCs." Cell stem cell 3(1): 99-‐108. Taoudi, S. and A. Medvinsky (2007). "Functional identification of the hematopoietic stem cell niche in the ventral domain of the embryonic 210 dorsal aorta." Proceedings of the National Academy of Sciences of the United States of America 104(22): 9399-‐9403. Thompson, B. J., S. Buonamici, et al. (2007). "The SCFFBW7 ubiquitin ligase complex as a tumor suppressor in T cell leukemia." The Journal of experimental medicine 204(8): 1825-‐1835. Tian, H., B. Biehs, et al. (2011). "A reserve stem cell population in small intestine renders Lgr5-‐positive cells dispensable." Nature 478(7368): 255-‐259. Tipping, A. J., C. Pina, et al. (2009). "High GATA-‐2 expression inhibits human hematopoietic stem and progenitor cell function by effects on cell cycle." Blood 113(12): 2661-‐2672. Tober, J., A. Koniski, et al. (2007). "The megakaryocyte lineage originates from hemangioblast precursors and is an integral component both of primitive and of definitive hematopoiesis." Blood 109(4): 1433-‐1441. Tomita, K., M. Hattori, et al. (1999). "The bHLH gene Hes1 is essential for expansion of early T cell precursors." Genes Dev 13(9): 1203-‐1210. Tomita, K., M. Hattori, et al. (1999). "The bHLH gene Hes1 is essential for expansion of early T cell precursors." Genes & development 13(9): 1203-‐1210. Tsai, F. Y., G. Keller, et al. (1994). "An early haematopoietic defect in mice lacking the transcription factor GATA-‐2." Nature 371(6494): 221-‐226. Tsivitse, S. (2010). "Notch and Wnt signaling, physiological stimuli and postnatal myogenesis." International journal of biological sciences 6(3): 268-‐281. Uchida, H., J. Zhang, et al. (1997). "AML1A and AML1B can transactivate the human IL-‐3 promoter." Journal of immunology 158(5): 2251-‐2258. Ueo, T., I. Imayoshi, et al. (2012). "The role of Hes genes in intestinal development, homeostasis and tumor formation." Development 139(6): 1071-‐1082. Urness, L. D., L. K. Sorensen, et al. (2000). "Arteriovenous malformations in mice lacking activin receptor-‐like kinase-‐1." Nature genetics 26(3): 328-‐331. van der Loo, J. C., W. A. Slieker, et al. (1995). "Use of ER-‐MP12 as a positive marker for the isolation of murine long-‐term in vitro repopulating stem cells." Experimental hematology 23(9): 1002-‐1010. 211 van Es, J. H., M. E. van Gijn, et al. (2005). "Notch/gamma-‐secretase inhibition turns proliferative cells in intestinal crypts and adenomas into goblet cells." Nature 435(7044): 959-‐963. Varnum-‐Finney, B., L. M. Halasz, et al. (2011). "Notch2 governs the rate of generation of mouse long-‐ and short-‐term repopulating stem cells." The Journal of clinical investigation 121(3): 1207-‐1216. Varnum-‐Finney, B., L. E. Purton, et al. (1998). "The Notch ligand, Jagged-‐1, influences the development of primitive hematopoietic precursor cells." Blood 91(11): 4084-‐4091. Vasiliauskas, D., E. Laufer, et al. (2003). "A role for hairy1 in regulating chick limb bud growth." Developmental biology 262(1): 94-‐106. Visan, I., J. B. Tan, et al. (2006). "Regulation of T lymphopoiesis by Notch1 and Lunatic fringe-‐mediated competition for intrathymic niches." Nature immunology 7(6): 634-‐643. Wang, X. D., C. C. Leow, et al. (2006). "Notch signaling is required for normal prostatic epithelial cell proliferation and differentiation." Developmental biology 290(1): 66-‐80. Wang, X. D., J. Shou, et al. (2004). "Notch1-‐expressing cells are indispensable for prostatic branching morphogenesis during development and re-‐ growth following castration and androgen replacement." The Journal of biological chemistry 279(23): 24733-‐24744. Wendorff, A. A., U. Koch, et al. (2010). "Hes1 is a critical but context-‐dependent mediator of canonical Notch signaling in lymphocyte development and transformation." Immunity 33(5): 671-‐684. Weng, A. P., A. A. Ferrando, et al. (2004). "Activating mutations of NOTCH1 in human T cell acute lymphoblastic leukemia." Science 306(5694): 269-‐ 271. Wilson, A., H. R. MacDonald, et al. (2001). "Notch 1-‐deficient common lymphoid precursors adopt a B cell fate in the thymus." The Journal of experimental medicine 194(7): 1003-‐1012. Willert, K., J. D. Brown, et al. (2003). "Wnt proteins are lipid-‐modified and can act as stem cell growth factors." Nature 423(6938): 448-‐452. Williams, D. E., J. Eisenman, et al. (1990). "Identification of a ligand for the c-‐kit proto-‐oncogene." Cell 63(1): 167-‐174. 212 Woodhoo, A., M. B. Alonso, et al. (2009). "Notch controls embryonic Schwann cell differentiation, postnatal myelination and adult plasticity." Nature neuroscience 12(7): 839-‐847. Wu, M., H. Y. Kwon, et al. (2007). "Imaging hematopoietic precursor division in real time." Cell stem cell 1(5): 541-‐554. Xu, T. and S. Artavanis-‐Tsakonas (1990). "deltex, a locus interacting with the neurogenic genes, Notch, Delta and mastermind in Drosophila melanogaster." Genetics 126(3): 665-‐677. Xue, Y., X. Gao, et al. (1999). "Embryonic lethality and vascular defects in mice lacking the Notch ligand Jagged1." Human molecular genetics 8(5): 723-‐730. Xue, Y., X. Gao, et al. (1999). "Embryonic lethality and vascular defects in mice lacking the Notch ligand Jagged1." Hum Mol Genet 8(5): 723-‐730. Yamamoto, N., W. Chang, et al. (2011). "Rbpj regulates development of prosensory cells in the mammalian inner ear." Developmental biology 353(2): 367-‐379. Yamamoto, N., K. Tanigaki, et al. (2003). "Notch/RBP-‐J signaling regulates epidermis/hair fate determination of hair follicular stem cells." Current biology : CB 13(4): 333-‐338. Yang, L. T., J. T. Nichols, et al. (2005). "Fringe glycosyltransferases differentially modulate Notch1 proteolysis induced by Delta1 and Jagged1." Molecular biology of the cell 16(2): 927-‐942. Yao, D., Y. Huang, et al. (2011). "Protein O-‐fucosyltransferase 1 (Pofut1) regulates lymphoid and myeloid homeostasis through modulation of Notch receptor ligand interactions." Blood 117(21): 5652-‐5662. Yochem, J. and I. Greenwald (1989). "glp-‐1 and lin-‐12, genes implicated in distinct cell-‐cell interactions in C. elegans, encode similar transmembrane proteins." Cell 58(3): 553-‐563. Yochem, J., K. Weston, et al. (1988). "The Caenorhabditis elegans lin-‐12 gene encodes a transmembrane protein with overall similarity to Drosophila Notch." Nature 335(6190): 547-‐550. Yoder, M. C. and K. Hiatt (1997). "Engraftment of embryonic hematopoietic cells in conditioned newborn recipients." Blood 89(6): 2176-‐2183. Yoder, M. C., K. Hiatt, et al. (1997). "In vivo repopulating hematopoietic stem cells are present in the murine yolk sac at day 9.0 postcoitus." 213 Proceedings of the National Academy of Sciences of the United States of America 94(13): 6776-‐6780. Yokomizo, T. and E. Dzierzak (2010). "Three-‐dimensional cartography of hematopoietic clusters in the vasculature of whole mouse embryos." Development 137(21): 3651-‐3661. Yokomizo, T., M. Ogawa, et al. (2001). "Requirement of Runx1/AML1/PEBP2alphaB for the generation of haematopoietic cells from endothelial cells." Genes to cells : devoted to molecular & cellular mechanisms 6(1): 13-‐23. Yonemura, Y., H. Ku, et al. (1996). "Interleukin 3 or interleukin 1 abrogates the reconstituting ability of hematopoietic stem cells." Proceedings of the National Academy of Sciences of the United States of America 93(9): 4040-‐4044. Yoshiura, S., T. Ohtsuka, et al. (2007). "Ultradian oscillations of Stat, Smad, and Hes1 expression in response to serum." Proc Natl Acad Sci U S A 104(27): 11292-‐11297. You, L. R., F. J. Lin, et al. (2005). "Suppression of Notch signalling by the COUP-‐ TFII transcription factor regulates vein identity." Nature 435(7038): 98-‐104. Yu, G., M. Nishimura, et al. (2000). "Nicastrin modulates presenilin-‐mediated notch/glp-‐1 signal transduction and betaAPP processing." Nature 407(6800): 48-‐54. Yuan, J. S., P. C. Kousis, et al. (2010). "Functions of notch signaling in the immune system: consensus and controversies." Annual review of immunology 28: 343-‐365. Zhang, Z., P. Nadeau, et al. (2000). "Presenilins are required for gamma-‐ secretase cleavage of beta-‐APP and transmembrane cleavage of Notch-‐ 1." Nature cell biology 2(7): 463-‐465. Zhao, C., J. Blum, et al. (2007). "Loss of beta-‐catenin impairs the renewal of normal and CML stem cells in vivo." Cancer cell 12(6): 528-‐541. Zhou, S., M. Fujimuro, et al. (2000). "SKIP, a CBF1-‐associated protein, interacts with the ankyrin repeat domain of NotchIC To facilitate NotchIC function." Molecular and cellular biology 20(7): 2400-‐2410. 214 Zhou, S. and S. D. Hayward (2001). "Nuclear localization of CBF1 is regulated by interactions with the SMRT corepressor complex." Molecular and cellular biology 21(18): 6222-‐6232. Zong, Y., A. Panikkar, et al. (2009). "Notch signaling controls liver development by regulating biliary differentiation." Development 136(10): 1727-‐ 1739. Zovein, A. C., J. J. Hofmann, et al. (2008). "Fate tracing reveals the endothelial origin of hematopoietic stem cells." Cell stem cell 3(6): 625-‐636. Zuniga-‐Pflucker, J. C. (2004). "T-‐cell development made simple." Nature reviews. Immunology 4(1): 67-‐72. 215 216 7.ABBREVIATIONS alk1: Activin A-receptor type II-like1 7-AAD: 7-actinomicin-D ADAM: a-disintegrin and metalloproteinase AER: Apical ectodermal ridge AGM: Aorta-Gonad-Mesonephros Ao: Aorta Aph1: Anterior pharynx-defective 1 Ascl2: Achaete-scute homolog 2 BFU-E: Erythroid burst forming unit BM: Bone marrow BMI: B lymphoma Mo-MLV insertion region 1. Polycomb ring finger oncogene BMP: Bone morfogenetic protein c-Kit: CD117. Stem cell growth factor receptor. v-kit Hardy-Zuckerman 4 feline sarcoma viral oncogene homolog Myb: Myeloblastosis viral oncogene homolog C. elegans: Caenorhabditis elegans CD133: Prominin-1 CD144: VE-cadherin CD150: Endoglin CD31: Pecam-1. Platelet endothelial cell adhesion molecule 217 CD34: Hematopoietic progenitor cell antigen CD38: ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase CD41: Integrin alpha-Iib CD45: Protein tyrosine phosphatase, receptor type, C CD48: BLAST-1 CFC-GM: Granulocyte macrophage colony forming cell CFC-MIX: Colony forming cell granulocyte-erythrocyte-macrophage- megacariocyte CFU-C: Colony forming unit in culture CFU-S11: Colony forming unit in spleen at day 11 after transplantation ChIP: Chromatin immunoprecipitation CLP: Common limfoid progenitor CMP: Common myeloid progenitor Compound E: S,S)- 2-[2-(3,5-Difluorophenyl)-acetylamino]-N-(1-methyl-2oxo-5-phenyl-2,3-dihydro-1H-benzo[e][1,4]diazepin-3-yl)-propionamide COUP: chicken ovalbumin upstream promoter-transcription factor cre: Causes Recombination CSL: CBF1, Supressor of hairless, Lag-1 CtBP: C-terminal binding protein DAPI: 4'6-diamidino-2-phenylondole DAPT: N-[N-(3,5-Difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester Delta: Delta-like DMSO: Dimethyl sulfoxide DNA: Deoxi-ribonucleic acid 218 DOS: Delta and OSM Dr: Doctor DSL: Delta, Serrate and Lag-2 E: Embryonic day post-coitus ECD: xtra cellular domain EfnB2: Ephrin-B2 EGF: Epidermal growth factor Eph: Ephrin ER: Endoplasmatic reticulum ERG: ETS-releted gene ETP: Early timic progenitors ETS: E-twenty six FBS: Fetal bovine serum FL: Fetal liver Fli: Friend leukemia integration transcription factor Flk: Fetal liver kinase Flt: Fms-like turosyne kinase G0: Gap zero phase or resting phase GFP: Green fluorescent protein GlcNAc: N-acetyl-glucosaminie Gli: Glioma-associated oncogene homolog GMP: Granulocyte monocyte progenitor Gro: Groucho. GSK3β: Glycogen synthetase 3β 219 GTP: Guanosine-5'-triphosphate HAT: Histone acetylase HDAC: Histone deacetlyase Hes: Hayri enhancer of split Hox: Homeobox Hrt: Hes releted gene HSC: Hematopoietic stem cells HSC/P: Hematopoietic stem cell and progenitors HXKXac: Histon X, lysin X, acteilation HXKXme: Histon X, lysin X, metilation I1FFL: Incoherent feed-forward loop type one IAHC: Intra aortic hematopoietic cluster IG: First exon general IL: Interleukine IMC: Inner mass cell IMIM: Institut municipal d'investigacions mèdiques IS: First exon specific ISH: In situ hibridaization JAG: Jagged KO: Knock out L685,458: Gamma secretase inhibitor. LacZ: β-galactosidase lb: Limb bud LEF: Lymphoid enhancer-binding factor 220 Lgr5: Leucine rich repeat containing G protein coupled receptor 5 Lineage Antibodies cocktail: Includes B220, Mac, Gr-1, Ter119, CD3 antibodies LNR: Lin12/Notch repeats LRP: Low-density lipoprotein receptor-related protein LT-HSC: Long term hematopoietic stem cells LTR: Long term reconstitution m: Mesonephros Mam: Mastermind-like Mash: Achaete-scute complex homolog1 Math: Athonal homolog MEP: Myeloerythorid progenitor MPP: Multipotent progenitors mRNA: Messenger ribonucleic acid Mx: Myxovirus resitance, interferon-inducible protein Myb: Myeloblastosis viral oncogene homolog Myo: Myocardin-like protein 1 MZ: Marginal zone Ncx: Sodium-calcium exchanger NECD: Notch extracellular domain NEXT: Notch extracellular truncation N1IC: Notch1 intra cellular domain NICD: Notch intra cellular domain NLS: nuclear localization signal 221 NO: Nitric oxide Nos: Nitric oxide synthase NRR: Negative regulatory region nt: Neural tube NTMIC: Notch transmembrane and intracellular domain Olfm4: Olfactomedin 4 OP9: Murin stromal cell line from a newborn op/op mouse (osteoperotic mutation) OPA: Glutamine-rich repeat region P-Sp: Paraortic splachnopleura PCR: Poly com repressive complex Pen: Presenilin PEST: Conserved proline/glutamic acid/serine/threonine-rich motifs Pofut: Protein O-fucosyltransferase PrE: Primitive endoderm PU: PU-box binding proteine. Spi, Spleen forming virus (SFFV) proviral integration oncogene. Rac: Ras-related C3 botulinum toxin substrate RAM: RBPjκ association module RBP-Jκ: Recombination signal binding protein for immunoglobulin kappa J Rho: Rhodopsin RNA: Ribonucleic acid Rumi: O-glucosyltransferase rumi Runx: Run-related transcription factor 222 S1: Site1 S2: Site2 S3: Site3 S4: Site4 SCA: Stem cell antigen SCF: Stem cell factor Scl/Tal: T-cell acute lymphocytic leukemia protein 1 homolog SHH: Sonic Hegehog SKIP: Ski-related protein SM: Smooth muscle SMA: Smooth muscle actin Smad: Mothers against decapentaplegic homolog Smo: Smoothened SMRT: Silencing mediator of retinoid and thyroid hormone receptors ST-HSC: Short term hematopoietic stem cells STR-assay: Short term reconstitution-assay TACE: Tumor necrosis factor alpha converting enzyme TCF: Transcription factor TE: Trophoectoblast TF: Transcription factor UA: Umbilical artery VA: Vitelline artery VEGF: Vascular endothelial growth factor WISH: Whole mount in situ hibridization 223 Wnt: Wingless-type MMTV integration site family WT: Wild-type YS: Yolk sac 224