Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
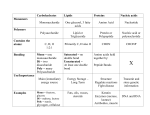
Struttura primaria: sequenza lineare degli aminoacidi Struttura secondaria: disposizione spaziale degli atomi dello scheletro polipeptidico regolare e ripetitiva in residui adiacenti Struttura terziaria: ogni ulteriore ripiegamento della catena polipeptidica; struttura tridimensionale globale del polipeptide Struttura quaternaria: disposizione spaziale delle subunità di una proteina oligomerica Protein length distribution H.sapiens Average protein length : 453 +/- 512 amino acid residues Size range: 3 - 34350 amino acid residues Protein length distribution E.coli K12 Average protein length : 316 +/- 215 amino acid residues Size range: 14 - 2358 amino acid residues phi CA—C—N—CA—C—N omega psi n: aminoacido in esame; n-1: aminoacido precedente; n+1: aminoacido seguente CA: carbonio alfa; C: carbonio carbonilico; N: azoto amidico Phi: angolo di rotazione tra carbonio alfa e azoto amidico; angolo diedro tra il piano formato dagli atomi Cn-1, Nn, CAn ed il piano Nn, CAn, Cn. Psi: angolo di rotazione tra carbonio alfa e carbonio carbonilico; angolo diedro tra il piano formato dagli atomi Nn, CAn, Cn ed il piano CAn, Cn, Nn+1. Omega: angolo di rotazione del legame peptidico = 180° Ramachandran plot psi phi -elica Space-filling model -elica In giallo: Catene laterali The structure repeats itself every 5.4 Å along the helix axis, i.e. we say that the -helix has a pitch of 5.4 Å. -helices have 3.6 amino acid residues per turn, i.e. a helix 36 amino acids long would form 10 turns. The separation of residues along the helix axis is 5.4/3.6 or 1.5 Å, i.e. the -helix has a rise per residue of 1.5 Å. 1.Every main chain C=O and N-H group is hydrogen-bonded to a peptide bond 4 residues away (i.e. On to Nn+4). This gives a very regular, stable arrangement. 2.The peptide planes are roughly parallel with the helix axis and the dipoles within the helix are aligned, i.e. all C=O groups point in the same direction and all N-H groups point the other way. Side chains point outward from helix axis and are generally oriented towards its amino-terminal end. Distortions of -helices. The majority of -helices in globular proteins are curved or distorted somewhat compared with the standard Pauling-Corey model. These distortions arise from several factors including: 1.The packing of buried helices against other secondary structure elements in the core of the protein. 2.Proline residues induce distortions of around 20 degrees in the direction of the helix axis. This is because proline cannot form a regular -helix due to steric hindrance arising from its cyclic side chain which also blocks the main chain N atom and chemically prevents it forming a hydrogen bond. Janet Thornton has shown that proline causes two H-bonds in the helix to be broken since the NH group of the following residue is also prevented from forming a good hydrogen bond. Helices containing proline are usually long perhaps because shorter helices would be destabilised by the presence of a proline residue too much. Proline occurs more commonly in extended regions of polypeptide. 3.Solvent. Exposed helices are often bent away from the solvent region. This is because the exposed C=O groups tend to point towards solvent to maximise their H-bonding capacity, i.e. tend to form H-bonds to solvent as well as N-H groups. This gives rise to a bend in the helix axis. L'enantiomero L è classificato tra i 20 amminoacidi ordinari in quanto la prolina entra nella composizione di molte catene polipeptidiche. Tra essi, è l'unica ad avere il gruppo amminico secondario, dato che il suo gruppo laterale si chiude sull'atomo di azoto formando una struttura ciclica. Per questo motivo, chimicamente, la prolina è in realtà un imminoacido, non un amminoacido. Essendo l'unico amminoacido il cui gruppo amminico è secondario, non sviluppa per reazione con la ninidrina il colore viola tipico degli altri amminoacidi, ma presenta una colorazione giallo/rossa. Most peptide bonds overwhelmingly adopt the trans isomer (typically 99.9% under unstrained conditions), chiefly because the amide hydrogen (trans isomer) offers less steric repulsion to the preceding Cα atom than does the following Cα atom (cis isomer). By contrast, the cis and trans isomers of the X-Pro peptide bond (where X represents any amino acid) both experience steric clashes with the neighboring substitution and are nearly equal energetically. Hence, the fraction of X-Pro peptide bonds in the cis isomer under unstrained conditions ranges from 10-40%; the fraction depends slightly on the preceding amino acid, with aromatic residues favoring the cis isomer slightly. From a kinetic standpoint, cis-trans proline isomerization is a very slow process that can impede the progress of protein folding by trapping one or more proline residues crucial for folding in the non-native isomer, especially when the native protein requires the cis isomer. This is because proline residues are exclusively synthesized in the ribosome as the trans isomer form. All organisms possess prolyl isomerase enzymes to catalyze this isomerization, and some bacteria have specialized prolyl isomerases associated with the ribosome. However, not all prolines are essential for folding, and protein folding may proceed at a normal rate despite having non-native conformers of many X-Pro peptide bonds. Prolil-isomerasi (rotamasi) • Isomerizzazione cis-trans Pro (10% dei residui di Pro) • Processo lento nel bilancio del folding • Note anche come immunofilline • Due famiglie: • A) ciclofilline (legano l’immunosoppressore ciclosporina A) • B) FKBP12 (legano l’immunosoppressore di origine fungina FK506) Distortions of -helices. The majority of -helices in globular proteins are curved or distorted somewhat compared with the standard Pauling-Corey model. These distortions arise from several factors including: 1.The packing of buried helices against other secondary structure elements in the core of the protein. 2.Proline residues induce distortions of around 20 degrees in the direction of the helix axis. This is because proline cannot form a regular -helix due to steric hindrance arising from its cyclic side chain which also blocks the main chain N atom and chemically prevents it forming a hydrogen bond. Janet Thornton has shown that proline causes two H-bonds in the helix to be broken since the NH group of the following residue is also prevented from forming a good hydrogen bond. Helices containing proline are usually long perhaps because shorter helices would be destabilised by the presence of a proline residue too much. Proline occurs more commonly in extended regions of polypeptide. 3.Solvent. Exposed helices are often bent away from the solvent region. This is because the exposed C=O groups tend to point towards solvent to maximise their H-bonding capacity, i.e. tend to form H-bonds to solvent as well as N-H groups. This gives rise to a bend in the helix axis. La cheratina è una proteina filamentosa ricca di zolfo, contenuto nei residui amminoacidici di cisteina; è molto stabile e resistente. È prodotta dai cheratinociti ed è il principale costituente dello strato corneo dell'epidermide, delle unghie e di appendici quali capelli, corna e piume. I residui a e d (idrofobici) si allineano sulla stesso lato dell’elica Il lato idrofobico di un’elica si associa con un’altra elica CHERATINA Passo dell’elica 5,1Å 3,5 residui per giro Due eliche si avvolgono l’una intorno all’altra in senso sinistrorso: Coiled coil (avvolgimento avvolto) Disulfide bridges In addition to intra- and intermolecular hydrogen bonds, keratins have large amounts of the sulfur-containing amino acid cysteine, required for the disulfide bridges that confer additional strength and rigidity by permanent, thermally-stable crosslinking. Human hair is approximately 14% cysteine. The pungent smells of burning hair and rubber are due to the sulfur compounds formed. Extensive disulfide bonding contributes to the insolubility of keratins, except in dissociating or reducing agents. The more flexible and elastic keratins of hair have fewer interchain disulfide bridges than the keratins in mammalian fingernails, , which are harder. Diagramma di un desmosoma Epidermolysis bullosa simplex (EBS) is a disorder resulting from mutations in the genes encoding keratin 5 or keratin 14. Blister formation of EBS is within the basal keratinocyte of the epidermis. COLLAGENO Ripetizione della tripletta Gly-X-Y, dove X è spesso Pro e Y è spesso Hyp (a volte Hyl) Ogni catena forma un’elica sinistrorsa con tre residui per giro e una distanza tra residui di 2,9 Å Tripla elica allungata Legami ad idrogeno Beta-sheet Legami ad idrogeno Catene laterali 7Å FIBROINA della seta (-Gly-Ser-Gly-Ala-Gly-Ala)n Foglietti beta antiparalleli le cui catene si estendono parallelamente all’asse della fibra. FIBROINA della seta (-Gly-Ser-Gly-Ala-Gly-Ala)n Dalla struttura del foglietto beta dipendono alcune delle proprietà meccaniche della seta. La seta , una delle fibre più resistenti, non è praticamente estensibile in quanto un suo allungamento causerebbe la rottura dei legami covalenti della molecola che si trova in una conformazione quasi completamente estesa. La seta è però flessibile perché i foglietti beta vicini sono uniti da forze deboli. Reverse turns (ripiegamenti inversi) A reverse turn is region of the polypeptide having a hydrogen bond from one main chain carbonyl oxygen to the main chain N-H group 3 residues along the chain (i.e. Oi to Ni+3). Helical regions are excluded from this definition and turns between -strands form a special class of turn known as the -hairpin. -hairpins -hairpins are one of the simplest super-secondary structures and are widespread in globular proteins. 70% of -hairpins are less than 7 residues in length with the tworesidue turns forming the most noticeable component. -a- motifs Anti-parallel -strands can be linked by short lengths of polypeptide forming -hairpin structures. In contrast, parallel -strands are connected by longer regions of chain which cross the -sheet and frequently contain -helical segments. This motif is called the a- motif and is found in most proteins that have a parallel -sheet. The loop regions linking the strands to the helical segments can vary greatly in length. The helix axis is roughly parallel with the -strands and all three elements of secondary structure interact forming a hydrophobic core. In certain proteins the loop linking the carboxy terminal end of the first -strand to the amino terminal end of the helix is involved in binding of ligands or substrates. The -a- motif almost always has a right-handed fold as demonstrated in the figure.