Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
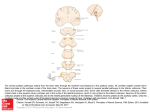
Lecture 11 1 The auditory nerve enters the brainstem at the pons-medulla junction, whereupon the first order neurons terminate on the cell bodies of the second order neurons in the cochlear nuclei. There are several nuclei within this complex but three are important in audition: 1. Anterior ventral cochlear nuclei (AVCN) 2. Posterior ventral cochlear nuclei (PVCN). 3. Dorsal cochlear nuclei (DCN). http://neuroscience.uth.tmc.edu/s2/chapte r13.html 2 Each first order neuron gives off collaterals or branches which have synapses with second order nerve cells located in different regions of the dorsal-ventral cochlear nuclei From this point on, the major ascending pathway traditionally is described as crossing the midline of the brainstem to the superior olivary complex on the opposite or contralateral side. That is, the second order neurons within the cochlear nuclei give off axons which cross over via the trapezoid body to terminate on third order neurons the cell bodies of which are located in the superior olive. 3 4 The central auditory pathways As in the case of the cochlear nuclei, the superior olivary complex is compromised of various nuclei. But only or two seem importance for hearing. The majority of fibres originating from the cochlear nuclei terminate in the medial olive in humans but in the lateral nucleus in animals such as cats or rodents. Superior olivary nuclei assume different relative sizes in different species (e.g the medial olive is largest in man) 5 The cochlear nuclei is located in the upper medulla, while the Superior olivary complex is located in the lower pons, within the hindbrain. Third order neurons in the superior olive give off axons which course centrally through the lateral lemniscus to terminate at the midbrain level in the inferior colliculus. From here, the fourth order neurons ascend to the medial geniculate body, one of several nuclei of the thalamus. 6 Axons of the fifth order neurons then terminate in the cell bodies in the auditory cortex. As noted in the previous lecture, the main area within which these cell bodies are found is the primary auditory cortex located on the superior surface of the superior temporal gyrus. This pathway is the most familiar. 7 Other ascending pathways within the brainstem are substantial: Of particular importance is the ipsilateral (same side) pathway which develops from second order neurons in the cochlear nuclei. -Crossed or uncrossed, fibers arising from the cochlear nuclei need not terminate in the superior olive but may go directly to the inferior colliculus. -These fibers ascend through the lateral lemniscus 8 Not all fibers ascending in this tract go directly to the inferior colliculus. Some fibers or collaterals may terminate in the nuclei of the lateral lemnisci. Fibers can also cross over to the opposite side at this level. Similarly, the inferior colliculus is not always a point of termination. Some ascending fibers also bypass the inferior colliculus (on either side) to terminate directly in the medial geniculate. The last stop along the ascending pathway in the brainstem before connection is made with the cortex. 9 Ascending auditory fibers also may give collaterals to other areas than nuclei which are direct constituents of the central auditory system. The auditory neurons in the brainstem may give off collaterals which go to the cerebellum. The cerebellum is involved intimately in the control of locomotion. This connection undoubtedly facilitate reflexive movements signaled by the acoustic stimuli, therefore, avoiding more time consuming pathways up to the cortex and back to this motor center. This direct input would facilitate the sudden removal of oneself from the path of oncoming car 10 Collaterals also are given off to terminate in a highly diffuse neural structure known as the reticular formation. It is through the reticular formation that an indirect route of communication is provided between various parts of the brain. The reticular formation is implicated strongly in the control of the level of consciousness or arousal and it receive information from all sensory systems. 11 There is also corticocochlear efferent system or descending pathway. It is more or less parallels the ascending pathway. However, only the pathway from the levelof the superior olive down is known well. Therefore, much attention has been focused on this more peripheral protion of the descending system which is formed by the olivocochlear bundles. 12 These efferent fibers course from the olivary region through the internal auditory meatus to innervate the inner and outer hair cells of the organ of Corti. Thus the brain not only receives information from the periphery, it also can exercise control over the peripheral system. The two temporal lobes can communicate via fibers of the commissure, the two halves of the cerebrum. 13 Thus there are points all along the central auditory pathways which allow for possible interaction between the two sides of the system. In other words, it is difficult to envision the right side of the auditory CNS doing something without the left side knowing about it. 14 Tuning curves providing best or characteristic frequencies (CFs) have been reported for all the levels of the auditory system from the cochlear nuclie through the auditory cortex. One of the most interesting aspects of the auditory pathways is the relatively systematic representation of frequency at each level. There is virtual mapping of the audible frequency range within each nuclear mass-neurons most sensitive to high frequencies are in one area, those sensitive to low frequencies are in another part, and those sensitive to intermediate frequencies are located successively between them. This orderly representation of frequency according to place is called tonotopic organisation. 15 High frequencies are represented basally in the cochlea, tapering down to low frequencies at the apex. This tonotopic arrangement is continued in the auditory nerve, where the apical fibers are found toward the core of the nerve trunk and basal fibers in the on the outside of the inferior margin. 16 Reaction of the BM to sound varies from base to apex according to the frequency of stimulation. Studies cited by Moore (1997) have shown that in normal healthy ears, each point on the BM is sharply tuned. This indicates that the response at each point is very sensitive to a limited range of frequencies, and that it requires higher intensities to produce a response if the signal frequency is outside this range. 17 The frequency that gives maximum response at a particular point along the BM is known as the characteristic frequency (CF) for that place. Pure tones produce patterns with single maxima, and the position depends on the frequency, there is a frequency to place conversion. The place frequency map which was first described by Liberman (1982) is the relationship between the tuning of the location along the cochlear partition and distance from base to apex. This is dependent on the CF. 18 Fibers with high CFs innervate the base of the cochlea while low CFs innervate the apex, tonotopic organisation is the maintenance of this neural spatial representation throughout the nuclei of the central auditory pathway. The neurons with different CF may represent the basis of a neural circuit designed to process spectral information, the ability of a single neuron to process auditory information depends on how inputs to the neurons interact., excitation or inhibition 19 Showing the tonotopic organisation of the basilar membrane when stimulated by a musical note, Image courtesy from http://www.drcameronent.com/images/hearing.jpg 20 • • The two most obvious parameters of the sound stimulus should be encoded somehow in the central auditory system: Frequency Intensity The first order neurons will be very important in this process. The intensity of the stimulus must be translated into a certain rate of discharge in neuron. 21 By virtue of the travelling wave in the cochlea, the spike rate also will be frequencydependant so it is necessary to determine the relationship between the activity of a single auditory neuron and events occurring in the cochlea. First order auditory neurons respond to a wide range of frequencies although not equally to all frequencies. The single unit response is frequency selective. As the stimulus frequency deviates from the one frequency to which the unit is more sensitive (CF) 22 Temporal (time) encoding theories as the basis of pitch perception have existed for many years. Helmholtz in the 19th century worked on proving that the tuned string like fibers along the basilar membrane constituted the place mechanism for the frequency encoding. Rutherford was working on the theory of temporal encoding. 23 Rutherford theory was that the basilar membrane acts much like the diaphragm of the microphone. The frequency of vibration of the diaphragm was presumed to be preserved in the frequency of the neural discharge, as if the neural discharges of the first order neurons followed or were perfectly in synchrony with the movements of the basilar membrane. However, by the end of the 20th century physiologists were able to demonstrate that all neurons are limited in how fast they can fire, due to the limitations imposed by refractory period. 24 This fundamental fact thus made Rutherford’s telephone or frequency theory invalid since the discharge rate commensurate (correspond) with the upper frequency limit of hearing in most animals are impossible for any neuron. For example, assuming a maximal rate of 2000 spikes/second this would mean that the upper frequency cutoff for hearing is 2000 Hz which is about one-tenth of the upper frequency limit of hearing in man 25 Only modification of the telephone theory only gained acceptance. This modification took the limitation in discharge rates of neurons into account and was called volley theory. It was postulated that whenever the frequency of the stimulus exceeds the discharge rates of neuron it is still possible to encode frequency into a time pattern of neuronal discharges by virtue of the discharge pattern of a group neurons. Even though the fiber cannot carry the information , a group can. 26 - - - A frequency can be encoded by three neurons: One fiber may discharge at one cycle of the stimulus but not recover sufficiently to fire again until several periods later. However, another fiber, which may have been refractory during the first fiber discharge might be ready to fire again. Later a third fiber may be able to discharge. 27 The encoding of frequency is likely to be the result of a combination of place and time encoding as suggested by Wever in his placevolley theory nearly three decades ago. Wever noted that the place theory is quite adequate for distinguishing between different tones for all but the low frequencies. Below about 400 Hz the selectivity of the travelling wave envelope does not appear to be sufficiently sharp to account for ability of the humans and other animals to discriminate one frequency from the other 28 Thus the periodicity of the neural discharges is properly the key component of frequency encoding and the temporal code predominates. Above 5000 Hz the place mechanism becomes totally necessary because the neurons cannot follow the frequency vibration of the basilar membrane in any fashion. Within the transition region between these two frequencies, both place and temporal encoding are involved in varying degrees in encoding of frequency and perception of pitch. 29