Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Stat Mech and Models
I. Probability
II. Ensemble Averages
III. Helix Models
Probability and the
partition function
Using the Boltzmann distribution, the probability at a given temperature of a
particular state is:
( j 1 )
Pj
nj
N
je
k BT
e
i
( i 1 )
k BT
i
The bottom is the canonical partition function.
j
Is the degeneracy
Probability rewriten
Let us take the probability expression, and rearrange
j
( j 1 )
je
Pj
i i e
k BT
( i 1 )
k BT
i
k BT
e
( i 1 ) k BT [ln(i ) ln(1 )]
k BT
i
How did I do the last step?
1
i
( j 1 ) kBT [ln( j ) ln(1 )]
e
( j 1 )
k BT
e
1
e
( i 1 )
k BT
( H j H1 ) T [ S j S1 ]
RT
e
e
( H i H1 ) T [ Si S1 ]
RT
i
What does the top look like?
Ensemble averages
To take an ensemble average:
O Ok PK
k
K
Ok k e
1
i
i
1 e
Average Energy:
E k PK
k
K
k k e
1
i
i
1 e
( k 1 )
k BT
( i 1 )
k BT
( k 1 )
k BT
( i 1 )
k BT
Ensemble averages
Average Energy:
E k PK
k
Notice:
ln Z 1 Z
T
Q T
K
k k e
1
K
i
i
1 e
( i 1 )
k BT
1 k
e
2 k
1
k BT
i
i
1 e
( k 1 )
k BT
( k 1 )
k BT
( i 1 )
k BT
What does this tell us about the utility of the partition function?
Ensemble averages
We can write macroscopic variables in terms of the parition function
Energy, pressure, entropy, helmholz free energy, and more
ln Z
S k ln Z kT
T
A kT ln Z
ln Z
p kT
V
Utility of Other
Ensembles:
Microcanonical:
S k ln
Isothermal/Isobaric:
G kT ln
Notes on Gibbs Free
Energy
Also know by some people as the free energy or by others as the
thermodynamics potential
G H T S
What might be a serious problem with simulating G?
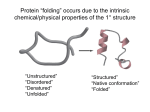
Helix Formation Models
Physical simple models for helix formation: amenable to paper and pencil
calculations {although the object of computational tests}
Each amino acid {or base pair} can exist in either unfolded or folded conformations
A unfolded amino acid is denoted by 0, and a folded amino acid is denoted by 1.
For a fixed number of amino acids {an n-mer}, we can enumerate the number of states:
N=2; 4 states
N=3; 8 states
N=m 2m states
What are the states for N=2?
What are the states for N=3?
Helix Formation Models
The most straightforward of these models are two parameter models:
These parameters are the nucleation parameter, s, and the propagation parameter, s.
Physically what do you think these parametbers mean?
To be precise:
s is the equilibrium constant for folding at the end of a sequence of folded AA
s exp(G p / RT )
What does this assume about the
energy of folding a single residue?
ss is the equilibrium constant for folding in an unfolded sequence
s s exp(Gn / RT )
Zip Model
A particular example of the simple helix formation models
The only allowed states are those with contigious stretches of 1’s:
Physically what does this mean in terms of structures? Is this realistic?
What are the states for N=2?
What are the states for N=3?
As N increases the number of allowed states compared to possible states decreases
Zip Model
Each allowed state contributes to the partition function.
A state contributes either 1 {the entirely unfolded state}, or ssm , where m is the
number of 1’s {the size of the folded stretch}. Why do we multiply?
What is the partition function for N=3?
What is the partition function for N=2?
n
Z 1 k s s k
k 1
Can you derive a expression for k?
How would this differ if we allowed all states?