Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
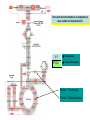
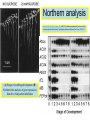
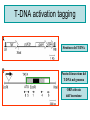
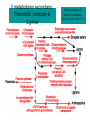
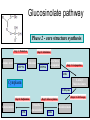
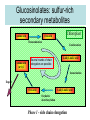
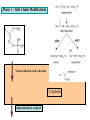
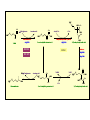
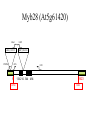
Ingegneria metabolica “smart” Strategie di attivazione parallela Come si ottiene un aumento di flusso? Aumentando S Sottraendo P Aumentando enzima Causano aumenti locali che faticano a propagarsi lungo la via (dampening) Aumentando attività Esaminiamo alcuni esempi di aumenti di flusso in vivo * In lievito nello switch tra fermentazione a respirazione (DeRisi, 1997) * Nel seme durante la mobilizzazione delle riserve lipidiche (Rylott, 2001) * Sintesi dei lipidi durante l’embriogenesi di Arabidopsis (O’Hara, 2002) * Altri esempi (vedi Fell) Diauxic shift in yeast Exploring the Metabolic and Genetic Control of Gene Expression on a Genomic Scale (DeRisi et al., 1997) Quali sono i geni che vengono attivati e quali vengono disattivati nella transizione da fermentazione a respirazione? Microarray con tutti i geni di lievito ibridato con mRNA a vari tempi di crescita Rosso = Aumento Verde = Diminuzione Seguiamo i trascritti nel tempo Passando da fermentazione a respirazione cosa cambia nel metabolismo? 4.9 PYK1 Variazione Gene interessato Rosso = Aumento Verde = Diminuzione Molti geni sono regolati in modo simile Variazione coordinata di molti geni E’ possibile classificare i geni in base alla regolazione: 6 classi Lipid mobilization in Arabidopsis germinating seeds Schematic representation of the pathways involved in storage lipid mobilization in oilseeds: 1, ACX; 2, multifuctional protein; 3, thiolase; 4, MS; 5, ICL; 6, PEPck. Northern analysis Rylott EL, Hooks MA, Graham IA. (2001) Co-ordinate regulation of genes involved in storage lipid mobilization in Arabidopsis thaliana. Biochem Soc Trans. 29:283-7. (A) Stages of seedling development (B) Northern blot analysis of gene expression from 0 to 8 days after imbibition Enzimi coinvolti ACC Malonyl-CoA transacilasi KAS III, II & I FAS - Acido grasso sintasi Lipid synthesis during embryogenesis O'Hara, P., et al. Plant Physiol. 2002;129:310-320 3-oxoacyl-ACP reductase (KR) biotin carboxylase (BC) acyl-ACP thioesterase (TE) enoyl-ACP reductase (ENR) acyl-carrier protein (ACP) FAS Components Exhibit Constant mRNA Ratios Abbondanza relativa dei trascritti It was demonstrated recently that mRNAs encoding the four subunits of heteromeric (ACCase) acetylCoA carboxylase accumulate at a constant molar ratio throughout silique development in Arabidopsis. The ratios were found to be CAC1:CAC2:CAC3:(accD-A & accD-B) = 0.14:1.0:0.17:0.06 (Ke et al., 2000) Via del triptofano in lievito Solo la simultanea espressione di molti (tutti) i geni causa un ΔJ paragonabile al ΔEi (ΔJ ≃ CJ x ΔEi ) Evidenze sperimentali Reguloni! La concentrazione dei metaboliti varia molto meno del flusso * Rate limiting step concept: more misguided than even MCA initially suggested * Agire su un solo punto è poco efficace e potrebbe essere deleterio Il metodo universale mantiene costanti le concentrazioni dei metaboliti [Si] evita effetti negativi dovuti all’aumento o alla riduzione di [Si] Referenze Referenze ai lavori sugli aumenti naturali in vivo Vedi anche Fell ultimo cap * DeRisi JL, Iyer VR, Brown PO. DeRisi JL, Iyer VR, Brown PO. (1997) Exploring the metabolic and genetic control of gene expression on a genomic scale. Science. 278:680-6. * O'Hara P, Slabas AR, Fawcett T. (2002) Fatty acid and lipid biosynthetic genes are expressed at constant molar ratios but different absolute levels during embryogenesis. Plant Physiol. 129:310-20 * Rylott EL, Hooks MA, Graham IA. (2001) Co-ordinate regulation of genes involved in storage lipid mobilization in Arabidopsis thaliana. Biochem Soc Trans. 29:283-7. * Niederberger P, Prasad R, Miozzari G, Kacser H. (1992) A strategy for increasing an in vivo flux by genetic manipulations. The tryptophan system of yeast. Biochem J. 287:473-9. * Zhao J, Last RL.(1996) Coordinate regulation of the tryptophan biosynthetic pathway and indolic phytoalexin accumulation in Arabidopsis. Plant Cell. 8:2235-44. * Eastmond PJ, Rawsthorne S. (2000) Ccoordinate changes in carbon partitioning and plastidial metabolism during the development of oilseed rape embryos. Plant Physiol. 122:767-74 •Universal method: Kacser and Acerenza (1993) A universal method for achieving increases in metabolite production Eur J. of Biochemistry 216:361-367 •Lütke-Eversloh T, Stephanopoulos G. (2008) Combinatorial pathway analysis for improved L-tyrosine production in Escherichia coli: identification of enzymatic bottlenecks by systematic gene overexpression. Metab Eng. 10:69-77. Ingegneria metabolica “in batch” (6) Espressione di fattori di trascrizione che regolano positivamente gli enzimi della via metabolica S + TF +A * Terpenoid Indole Alkaloyd (TIA) * via dei flavonoidi cere, glucinolati... + B + C Usando i fattori di trascrizione probabilmente si mantengono le “giuste proporzioni tra gli enzimi CAVEAT: ci sono limiti a questa strategia? P Certo, alcuni enzimi come già molto abbondanti (es. quelli del calvin o glicolitici) Fig. 1. Biosynthesis of TIAs in C. roseus. Solid arrows indicate single enzymatic conversions, whereas dashed arrows indicate multiple enzymatic conversions. Numerosi enzimi della via sono stati identificati e clonati. Esiste un fattore di trascrizione capace di attivarli tutti insieme? Abbreviations of enzymes: AS, anthranilate synthase; DXS, D-1-deoxyxylulose 5phosphate synthase; G10H, geraniol 10-hydroxylase; CPR, cytochrome P450-reductase; TDC, tryptophan decarboxylase; STR, strictosidine synthase; SGD,strictosidine b-D-glucosidase; D4H, esacetoxyvindoline 4-hydroxylase; and DAT, acetyl-CoA:4O-deacetylvindoline 4-O-acetyltransferase. Genes regulated by ORCA3 are underlined. T-DNA activation tagging Struttura del T-DNA Punto di inserzione del T-DNA nel genoma ORF attivata dall’inserzione Linea cellulare selezionata con inibitori delle TDC. L’inserzione del T-DNA porta ad un aumento del flusso nella via Molti altri geni della stessa via sono indotti nella linea cellulare Il metabolismo secondario: Flavonoidi, Antociani e Lignina Genes encoding all enzymes indicated in red are clock-controlled Myb transcription factor PAP1 I geni in rosso sono implicati nella biosintesi dei fenilpropanoidi e sono controllati dal ritmo circadiano Alcuni geni sembrano essere regolati in maniera molto simile dal punto di vista temporale. Può essere segno di un controllo comune mediato cioè dallo stesso fattore di trascrizione? Activation tagging Il mutante pap1-D presenta una colorazione rossa (carattere dominante) e accumula antocianine (una classe di flavonoidi) Molti geni della via dei fenilpropanoidi (e sue diramazioni: flavonoidi, antocianine) sono espressi maggiormente nel mutante. Il mutante pap1-D presenta una maggiore attività enzimatica e più lignina. La sovraespressione di Pap1 o Pap2 in Tabacco o Arabidopsis porta ad un’intensa pigmentazione Come identificare i fattori implicati nella trascrizione di vie metaboliche mutanti classici (indotti o spontanei) gene activation tagging o sovraespressione Coregolazione elementi comuni in cis elementi comuni in trans (?) identificazione del fattore tramite One-hybryd Identificazione…. Attenzione: i fattori di trascrizione sono enzimi (?) e spesso agiscono in sinergia Immagini cortesia del prof. C. Martin Lobe Tube Geni regolatori in Anthyrrinum majus Diversi geni della via sono down-regulated nel mutante delila ma solo nella zona con ridotta pigmentazione Tobacco crosses: 35S:Del x 35S:Ros1 Piante di Arabidopsis che sovraesprimono uno solo dei due fattori non mostrano accumulo. Quando sono coespressi l’aumento di flusso è notevole. Immagini cortesia del prof. C. Martin Sinergismo ! Rosea1 + Delila can give 100-fold + activation and anthocyanin levels of up to 10 mg/g fwt. They can also increase flux through pathway branches 2.5-fold. Other regulatory combinations are not so potent Immagini cortesia del prof. C. Martin Fattori di trascrizione coinvolti nella regolazione del metabolismo in pianta Broun P. (2004) Transcription factors as tools for metabolic engineering in plants. Curr Opin Plant Biol. 7:202-9. Altri esempi: - Cernac et al. (2006) The WRI1 gene encodes an AP2/EREBP transcription factor involved in the control of metabolism, particularly glycolysis, in the developing seeds. Plant Physiology 141:745757. - Xie et al. (2006) Metabolic engineering of proanthocyanidins through co-expression of anthocyanidin reductase and the PAP1 MYB transcription factor. Plant J. 45:895-907. - Metabolismo degli olii in foglia: Santos Mendoza et al., (2005) FEBS Lett. 579:4666-4670. LEAFY COTYLEDON 2 - Kannangara et al. (2007) The transcription factor WIN1/SHN1 regulates Cutin biosynthesis in Arabidopsis thaliana. Plant Cell. 2007 Apr;19(4):1278-94. - Aharoni et al. (2004) The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell. 16:2463-80. - Baud and Lepiniec (2009) Regulation of de novo fatty acid synthesis in maturing oilseeds of Arabidopsis, Plant Physiol. Biochem. 47:448–455. - Ruuska et al. (2002) Contrapuntal networks of gene expression during Arabidopsis seed filling, Plant Cell 14:1191–1206. - Shen et al. (2010) Expression of ZmLEC1 and ZmWRI1 increases seed oil production in maize, Plant Physiol. 153:980–987. - Pouvreau et al. (2011) Duplicate maize Wrinkled1 transcription factors activate target genes involved in seed oil biosynthesis, Plant Physiol. 156:674–686. - Zhang et al. (2002) Similarity of expression patterns of knotted1 and ZmLEC1 during somatic and zygotic embryogenesis in maize (Zea mays L.), Planta 215:191–194. - Maeo et al. (2009) An AP2-type transcription factor, WRINKLED1, of Arabidopsis thaliana binds to the AW-box sequence conserved among proximal upstream regions of genes involved in fatty acid synthesis, Plant J. 60:476–487. WIN1: wax inducer (biosintesi delle cere) Broun P, Poindexter P, Osborne E, Jiang C-Z, Riechmann JL: WIN1, a transcriptional activator of epidermal wax accumulation in Arabidopsis. Proc Natl Acad Sci USA 2004, 101(13):4706-11 Activation of wax production in Arabidopsis plants that overexpress WIN1, an ERF-type transcription factor, and concurrent induction of wax pathway genes. Morphological phenotype of (a) a control (wt) and (b) 35S::WIN1 plants. Note the glossy appearance of 35S::WIN1-overexpressing leaves. Scanning electron microscope (SEM) images of (c) control and (d) 35S::WIN1 leaf surfaces: WIN1 overexpressors produce wax crystals, which are absent from control leaves. (Magnification: 3000x.) Stomatal cells are shown at the centre of the images. (e) Northern analysis of the expression of wax pathway genes in 35S::WIN1 and control plants: KCS1, which encodes a putative fatty acid elongase, and CER1, encoding a putative fatty acid decarbonylase, are induced in 35S::WIN1 plants. Northern and microarray analyses of 35S::WIN1 plants indicated that several genes that are implicated in wax biosynthesis, such as ECERIFERUM1 (CER1) and 3-KETOACYL-COA SYNTHASE1 (KCS1), were upregulated in the WIN1-overexpressors wt b and c are representative of medium, and high levels of leaf glossiness 35S::WIN1 35S::WIN1 Total fatty acids per seed for the untransformed mutant (wri1) and wild type (WT) (a), and transgenic lines in the wri1 background (b) or the wild type background (c). Fatty acid composition Lipid and fatty acid compositions, after LEC2:GR induction in leaves Lipid composition. Transcriptional regulation of triacylglycerol biosynthesis in maturing seeds of Arabidopsis thaliana LEAFY COTYLEDON1 (LEC1), LEC2, ABSCISIC ACID INSENSITIVE3 (ABI3), and FUSCA3 (FUS3) arenormally expressed predominantly in seeds, can induce the deposition of seed oil in vegetative tissues when ectopically activated in seedlings. Family TF Name Summary of Role in Seed Oil Deposition B3 domain; AFL Clade ABSCISIC ACID INSENSITIVE3 (ABI3), LEAFY COTYLEDON2 (LEC2), FUSCA3 (FUS3) Master regulators of embryogenesis and seed maturation; mutation/overexpression often associated with pleiotropic effects; direct and indirect regulation of suites of genes involved in carbohydrate and lipid metabolism, including fatty acid synthesis, triacylglycerol assembly and packaging HAP3/C BP LEAFY COTYLEDON1 (LEC1), LEC1-LIKE (L1L) Subunits of CCAAT binding proteins; capable of working independently of CBP; master regulators of embryogenesis and seed maturation; direct and indirect regulation of genes involved in carbohydrate and lipid metabolism AP2 WRINKLED1 (WRI1) Direct target of master regulators having more specific role towards seed oil biosynthesis; mutants dramatically reduced in seed oil content and wrinkled appearance; direct and indirect regulation of carbohydrate and lipid metabolism genes, particularly plastidial fatty acid synthesis Dof GmDof4 GmDof11 Transgenic expression yields higher seed oil levels; direct and indirect regulation of lipid metabolism genes; possible negative regulators of seed storage proteins CHD3 PICKLE (PKL) Putative chromatin remodeling factor; represses master regulator genes at germination; associated with the repressive chromatin mark H3K27me3 PRC2 FERTILIZATION INDEPENDENT ENDOSPERM (FIE), SWINGER (SWN), EMBRYONIC FLOWER2 (EMF2) Components of Polycomb Repressive Complex 2 that catalyze deposition of H3K27me3; repressors of seed maturation genes in vegetative tissues B3 domain; HSI2 Clade HIGH-LEVEL EXPRESSION OF SUCROSE INDUCIBLE GENE2 (HSI2)/VAL1, HSI2-LIKE1 (HSIL1/VAL2), HSL2/VAL3 Act redundantly to repress AFL Clade genes and other positive regulators of seed maturation during germination and in seedlings; possible chromatin remodeling activities AP2 APETALA2 (AP2) Negative regulator of seed size, possibly via carbohydrate metabolism in the seed coat; effects on seed oil deposition likely indirect HD-ZIP GLABRA2 (GL2) Negative regulator of oil content; loss of seed mucilage proposed to make more C available for fatty acid synthesis Zhong and Ye (2009) Transcriptional regulation of lignin biosynthesis. Plant Signal Behav. 4:1028-34. According to Metabolic Control Analysis, the parallel activation (multisite modulation) of enzymes within a biochemical pathway is the optimal strategy for changing fluxes retains metabolite and control homeostasis How universal is the “universal method” in vivo? PDS (Phytoene Desaturase) If a mRNA level changes, what happens to other ones in the same metabolic pathway? mRNA is not equal to protein Two-gene flux changes over long times scatterplot Use data from many different tissues, mutants, conditions… Pearson correlation coefficient PSY (Phytoene Synthase) A square matrix At4g15560 At5g11380 At5g62790 At2g02500 At2g26930 At1g63970 At5g60600 At4g34350 At3g21500 1.00 0.12 0.01 0.03 0.04 0.20 0.25 0.19 0.19 At4g15560 0.12 1.00 0.35 0.75 0.66 0.65 0.66 0.73 0.73 At5g11380 0.01 0.35 1.00 0.35 0.31 0.40 0.21 0.18 0.15 At5g62790 0.03 0.75 0.35 1.00 0.69 0.70 0.78 0.78 0.68 At2g02500 0.04 0.66 0.31 0.69 1.00 0.77 0.72 0.67 0.56 At2g26930 0.20 0.65 0.40 0.70 0.77 1.00 0.80 0.67 0.60 At1g63970 0.25 0.66 0.21 0.78 0.72 0.80 1.00 0.88 0.74 At5g60600 0.19 0.73 0.18 0.78 0.67 0.67 0.88 1.00 0.77 PSY (Phytoene Synthase) At3g21500 PSY (Phytoene Synthase) At4g34350 0.19 0.73 0.15 0.68 0.56 0.60 0.74 0.77 1.00 From numbers to colours Gene A Gene B Gene A Essentially the same strategy published recently by Toufighi K, et al. (2005) Plant J. 43:153-63 The Botany Array Resource: e-Northerns, Expression Angling, and promoter analyses. Gene B The Red Square… Gene ABCDEFGHIJKLMNOPQRS Group 1 Group 1 & 3 Group 3 are coregulated Coregulated genes close in the list will appear as a red square Apply the correlation analysis to the entire “metabolic genome” (enzymes, transporters….) B A B Isoprenoid biosynthesis two indipendent pathways in plants: A cytosolic B plastidial Lange and Ghassemian (2003) Genome organization in Arabidopsis thaliana: a survey for genes involved in isoprenoid and chlorophyll metabolism. Plant Mol Biol. 51:925-48. Plastidial pathway: Carotenoids Phytyl Plastoquinone Phylloquinone Tocopherol Mono-terpenes Phytochrome Gibberellic acid Abscissic acid. Figure from Lange and Ghassemian (2003) 1000 5 0 0 g e n e s 1414 2000 2750 5 0 0 g e n e s Plastidial IPP Cytosolyc IPP (meval.) 1 0 0 ◄ GGPP synthases: 10 isoforms Carotenoid g e n e s Chlorophyll GA GGPP synthase GGPP Prenyl group Phytyl PP Chlorophyll (At3g20160) At3g29430 At3g32040 At4g36810 (At4g38460) At3g29430 and At3g32040 provide GGPP for… Which GGPP synthase isoform works in the carotenoid pathway? At3g29430 Migliori correlatori tra tutti i geni di Arabisopsis (R value in linear plots) At3g29430 At3g29410 At4g33720 At5g15180 At1g53940 At2g24400 At5g59680 At5g24410 At1g73780 At3g47210 At3g59370 At1g33900 At5g03570 At3g32040 At1g21210 At5g37450 At1g11540 At3g49860 At2g31085 At1g49030 At1g66020 At3g05950 At5g15725 At3g01190 At4g31875 At2g38600 At3g46400 1.0000 0.8313 0.7440 0.7352 0.7341 0.7081 0.7022 0.6942 0.6873 0.6867 0.6865 0.6857 0.6855 0.6847 0.6831 0.6827 0.6810 0.6770 0.6739 0.6734 0.6725 0.6709 0.6668 0.6644 0.6620 0.6569 0.6532 geranylgeranyl pyrophosphate synthase, putative terpene synthase/cyclase family protein pathogenesis-related protein, putative peroxidase, putative GDSL-motif lipase/hydrolase family protein auxin-responsive protein, putative / small auxin up RNA (SAUR_D) leucine-rich repeat protein kinase, putative glucosamine/galactosamine-6-phosphate isomerase-related protease inhibitor/seed storage/lipid transfer protein expressed protein expressed protein avirulence-responsive protein, putative iron-responsive transporter-related geranylgeranyl pyrophosphate synthase, putative wall-associated kinase 4 leucine-rich repeat transmembrane protein kinase, putative expressed protein ADP-ribosylation factor, putative Clavata3 / ESR-Related-6 (CLE6) expressed protein terpene synthase/cyclase family protein germin-like protein, putative expressed protein peroxidase 27 (PER27) (P27) (PRXR7) expressed protein acid phosphatase class B family protein leucine-rich repeat protein kinase, putative At3g29430 is possibly involved in terpene synthesis Calvin cycle At4g26520 At4g26530 At4g38970 At2g21330 At5g56630 At5g47810 At4g32840 At2g22480 At4g26390 At3g55440 At2g29560 At1g07110 At1g13440 At1g42970 At3g26650 At3g04120 At3g12780 At1g58150 At1g56190 At1g22170 At1g78040 At3g08590 At5g04120 At3g22960 At5g52920 At2g21170 At5g61410 At1g71100 At3g04790 At2g45290 At3g60750 At1g32060 At1g43670 At3g54050 At3g55800 At5g35790 At1g09420 At5g24420 At5g24410 At3g49360 At1g13700 At5g44520 At2g01290 At5g39320 At5g64290 At5g35630 At4g37930 At1g23310 At3g19710 At1g32450 3.5 in log scale >3000 fructose-bisphosphate aldolase, cytoplasmic fructose-bisphosphate aldolase, putative fructose-bisphosphate aldolase, putative fructose-bisphosphate aldolase, putative phosphofructokinase family protein phosphofructokinase family protein phosphofructokinase family protein phosphofructokinase family protein pyruvate kinase, putative triosephosphate isomerase, cytosolic, putative enolase, putative fructose-6-phosphate 2-kinase / fructose-2,6-bisphosphatase (F2KP) glyceraldehyde 3-phosphate dehydrogenase, cytosolic, putative glyceraldehyde-3-phosphate dehydrogenase B, chloroplast (GAPB) glyceraldehyde 3-phosphate dehydrogenase A, chloroplast (GAPA) glyceraldehyde-3-phosphate dehydrogenase, cytosolic (GAPC) phosphoglycerate kinase, putative hypothetical protein phosphoglycerate kinase, putative phosphoglycerate/bisphosphoglycerate mutase family protein pollen Ole e 1 allergen and extensin family protein 2,3-biphosphoglycerate-independent phosphoglycerate mutase phosphoglycerate/bisphosphoglycerate mutase family protein pyruvate kinase, putative pyruvate kinase, putative triosephosphate isomerase, chloroplast, putative ribulose-phosphate 3-epimerase, chloroplast, putative / ribose 5-phosphate isomerase-related ribose 5-phosphate isomerase-related transketolase, putative transketolase, putative phosphoribulokinase (PRK) / phosphopentokinase fructose-1,6-bisphosphatase, putative fructose-1,6-bisphosphatase, putative sedoheptulose-1,7-bisphosphatase, chloroplast glucose-6-phosphate 1-dehydrogenase / G6PD (APG1) glucose-6-phosphate 1-dehydrogenase, putative / G6PD, putative glucosamine/galactosamine-6-phosphate isomerase-related glucosamine/galactosamine-6-phosphate isomerase-related glucosamine/galactosamine-6-phosphate isomerase family protein glucosamine/galactosamine-6-phosphate isomerase family protein ribose 5-phosphate isomerase-related expressed protein UDP-glucose 6-dehydrogenase, putative oxoglutarate/malate translocator, putative glutamine synthetase (GS2) glycine hydroxymethyltransferase glutamate:glyoxylate aminotransferase 1 (GGT1) branched-chain amino acid aminotransferase, putative proton-dependent oligopeptide transport (POT) family protein Reducing glucosinolates in Arabidopsis Glucosinolates are sulphur rich compounds from brassicas Some beneficial, other toxic (quantity!) Upon wounding are converted into toxic products Two branches Mutants isolated Short chain Aliphatic GSL Long chain Indolic GSL Beekwilder et al., (2008) PLoS 3:e2068. Glucosinolate pathway Phase 2 - core structure synthesis Step 1: Oxidation Amino Acid Step 2: Oxidation Aldoxime CYP79s CYP83s Aci-Nitro compound Step 3: Conjugation GSTs S-Alkyl Thioidroximate Cytoplasm C-S Lyase Step 6: Sulfatation Glucosinolate ST5s Step 5: Glucosylation Desulfoglucosinolate Step 4: C-S Clevage Thioidroximate UGTs Glucosinolates: sulfur-rich secondary metabolites Amino acid Oxo-acid Chloroplast Transamination Amino acid (n+1)C Several rounds of chain elongation are possible Condensation 2-alkyl-malic acid Isomerization Export Oxo-acid 3-alkyl-malic acid Oxydative decarboxylation Phase 1 - side chain elongation Kroymann et al., Plant Physiology (2001) 127:1077–1088, Phase 3 - Side Chain Modification Various oxidations on the side chain Cytoplasm compartimentation -transport TRYPTOPHAN BIOSYNTHESIS GLUCOSINOLATE FROM TRYPTOPHAN AND PHENYLALANINE SHARED GENES (PAPS BIOSYNTHESIS,C-S LYASE AND GLUCOSYL TRANSFERASE) CYP79A2 At5g05260 CYP79B2 At4g39950 CYP79B3 At2g22330 CYP83B1 At4g31500 At1g74100 ST5a ATGSTF10 At2g30870 ATGSTU13 At1g27130 ATGSTF9 At2g30860 F17I23 At4g30530 ASA1 At5g05730 TSA1 At5g17990 TRP 1 At3g54640 IGPS At2g04400 DHS1 At4g39980 SAT52 At5g56760 OASC At3g59760 PEN2 PEN3 SUR1 UGT74B1 AKN2 AKN1 BCAT3 BCAT4 MAM1 F17J16 T9E8 MFL8 CYP83A1 ATGSTF11 ATGSTU20 ST5b ST5c B5 #1 F12P19 T3P18 F16J13 MYB28 F28J8 AOP2 AOP3 At2g04400 At1g59870 At2g20610 At1g24100 At4g39940 At2g14750 At3g49680 At3g19710 At5g23010 At3g58990 At4g13430 At2g43100 At4g13770 At3g03190 At1g78370 At1g74090 At1g18590 At2g46650 At1g65860 At1g62560 At4g12030 At5g61420 At1g21440 At4g03060 At4g03050 HOMOMETHIONINE BIOSYNTHESIS GLUCOSINOLATE FROM HOMOMETIONINE Aromatic branch CYP79A2 CYP79B2 CYP79B3 CYP83B1 Phase II – ST5a - Sulfotransferase GLS from Trp and Phe Glutathione S-Transferase Glutathione S-Transferase Glutathione S-Transferase Anthranilate synthase ASA1 -Anthranilate synthase α subunit TSA1 - Trp synthase, alpha subunit TRP Biosynthesis TRP1- P-ribosyl-anthranilate synthase IGPS Indole-3-glycerol p synthase DHS1 – DAHP synthetase 1 SAT 52 – Serine O-acetyltrasferase Cysteine Synthase Glycosil hydrolase family 1 protein ABC Transporter SUR1 - C-S Lyase UGT74B1 S-Glucosil Trasferase (PAPS–Biosynthesis, C-S AKN2 – Adenylylsulfate kinase 2 Lyase, Glucosyl Transferase) AKN1 – Adenylylsulafte kinase 1 BCAT3 Branched-chain amino BCAT4 acid aminotransferase MAMPhase 1 – 2 isopropylmalate synthase 3 I - Homomet Aconitase C-terminal domain Biosynthesis Aconitase family protein Aconitase C-terminal domain CYP83A1 Glutathione-S Transferase Phase II – GLS from Glutathione-S Transferase ST5b – Sulfotransferase Homomet ST5c – Sulfotransferase Cytochrome b5 Flavin-contaning monooxygenase Phase III, monooxygenase transport and Flavin-contaning Bile acid Sodium symporter regulation – GLS from MYB 28 HOMOMET Mutase family protein AOP2 - Dioxygenase AOP3 -Dioxygenase Phase II Shared genes Aliphatic branch GLUCOSINOLATE BIOSYNTHESIS METHIONINE SIDE-CHAIN ELONGATION At4g13770 At3g03190 At1g78370 At2g20610 At1g18590 At1g74090 At3g19710 At5g23010 At3g58990 At2g43100 At4g13430 At4g12030 At5g61420 At2g46650 At1g62560 At1g21440 CYP83A1 Monooxygenase “GLUCOSINOLATE FROM FENIL.-OMOMET.” ATGSTF11 Glutathione S-transferase SUR1 Phase II - GLS biosynthesis (Met derived) C-S Lyase “GLUCOSIN. FROM PHENILAL-TRYPT-HOMOMET.” ST5c Sulfotransferase “GLUCOSINOLATE FROM HOMOMET.” ST5b Sulfotransferase “GLUCOSINOLATE FROM HOMOMET.” ATGSTU20 Glutathione S-transferase BCAT4 Aminotransferase“HOMOMET.–LEUCINE BIOSYNTHESIS” MAM1 Phase I -BIOSYNTHESIS” GLS biosynthesis 2-isopropylmalate Synthase “HOMOMET F17J16 derived) SIDE-CHAIN Aconitase C-terminal domain(Met “LEUC.-HOMOMET.BIOSYNTHESIS” MFL8 ELONGATION Aconitase C-terminal domain“HOMOMET. BIOSYNTHESIS” T9E8 Aconitate hydratase F16J13 Sodium symporter family protein MYB28 Transcription factor T3P18 Candidate genes for transport, regulation... (MET derived GLS) Flavin conteining monooxygenase family protein F28J8 Mutase family protein B5 #1 Cytochrome b5 Phase I and II enzymes are co-regulating H C 3 O H O S O -C h e to g lu ta r a to S H C 3 O A na m in oa c id C o A A c e ty lC o A O H O S O H H C 3 N H 2 A m in o tr a n sfe r a se M e th io n in e A t3 g 1 9 7 1 0 O H O 2o x o4m e th y lth iob u ta n o ica c id M e th y lth io a lk y lm a la tesy n th a se O H A t5 g 2 3 1 0 0 2 -(2 'm e th y tio )e th y lm a lica c id BCAT4 A c o n ita se MAM1 A t3 g 5 8 8 9 0 BCAT3 A t2 g 4 3 1 0 0 O O -C h e to g lu ta r a to O H A na m in oa c id H C 3 S H C 3 H o m o m e th io n in e N H 2 N A D H O + N A D S H C 3 O H O H O S H O O 2o x o5m e th y lth iop e n ta n o ica c id C O 2 O H 3 -(2 'm e th y tio )e th y lm a lica c id Myb28 (At5g61420) LBa1 SALK_136312 LB51 BRC_H161Lb ATG PROM 1 ATG EX3 134 214 344 484 1623 TGA MAM-L 400 350 300 250 RGE Effect of knocking out Myb28? 200 150 100 50 0 RT-PCR on 2 controls and 2 KOs -50 C 2 6 9 C Leaf 2 6 9 6 9 6 9 Root MYB 28 CYP83A1 16 14 1.8 1.6 10 1.4 8 1.2 6 1.0 RGE RGE 12 4 0.8 0.6 2 0.4 0 C 2 6 9 C Leaf 2 6 0.2 9 0.0 Root -0.2 C 2 6 9 C Leaf 2 Root MYB 29 Aconitase 2.0 3.0 1.5 2.5 2.0 RGE RGE 1.0 0.5 0.0 C 2 6 9 C 2 6 9 1.5 1.0 0.5 0.0 -0.5 Leaf Root C 2 6 9 C 2 -0.5 Leaf Root 5.31 279 565.0460 -2 5.00 M09107 2.74 144 422.0223 % 98 37.26 1962 487.1212 39.66 2088 223.0983 Wt and Myb28-KO metabolome 10.00 4.42 232 436.0179 5.24 275 565.0455 27.46 1445 505.1335 10.88 572 323.1347 15.00 14.40 757 478.0881 20.00 18.96 998 739.1794 25.00 20.06 1055 492.0634 30.00 26.45 1392 339.0452 35.00 45.00 50.00 55.00 1: TOF MS ESBPI 54.25;2856;791.4733 3.43e4 32.86 1729 477.0614 27.44 1444 505.1348 10.88 572 323.1338 40.00 46.99 2474 333.1882 42.60 2242 478.0856 37.26 1962 487.1232 51.52 2712 476.1041 46.97 2473 333.1925 -2 5.00 10.00 15.00 20.00 25.00 30.00 35.00 40.00 45.00 50.00 55.00 1: TOF MS ESBPI 54.22;2854;791.4753 3.42e4 M09106 20.17;1062;447.0292 4.38 230 436.0205 % 98 5.29 278 565.0436 16.79 883 385.1132 26.41 1390 339.0430 32.80 1727 477.0533 27.44 1444 389.1235 10.91 574 323.1335 37.26 1962 487.1258 39.61 2085 223.0984 51.50 46.97 2711 2473 1046.5104 333.1924 ko -2 5.00 10.00 15.00 20.00 25.00 30.00 35.00 40.00 45.00 M09105 98 % 2.78 145 422.0237 4.40 231 436.0169 18.98 999 739.1732 14.40 GSL unknown 757 20.12 26.40 1059 1389 492.0636 339.0451 32.86 1729 477.0608 478.0864 5.27 277 565.0471 55.00 1: TOF MS ESBPI 54.34;2861;791.4747 3.43e4 Methylsulfinyloctyl 42.62 Methylsulfinylheptyl 2243 27.46 1445 389.1248 10.91 574 323.1330 37.26 1962 487.1224 478.0875 46.99 2474 333.1934 50.00 51.60 2717 492.0991 51.14 2692 552.2348 -2 5.00 10.00 15.00 20.00 25.00 30.00 35.00 40.00 45.00 50.00 wt Time 55.00 myb28, myb29 and myb28myb29 Mutating Myb28 and Myb29 Regulators Beekwilder et al., (2008) PLoS 3:e2068. Reducing glucosinolate content... ...stimulates pest growth and damage! Beekwilder et al., (2008) PLoS 3:e2068 Insect feeding Effect of the double KO Too late! 2828 genes What is the distribution of all the R values in the matrix? La spalla di valori alti e positivi di R all’interno dei geni metabolici è la testimonianza che esiste molta coregolazione Open issues Explore enzyme subsets Pathway identification Clustering of enzymes Shared cis-elements / regulators Suggest substrate for enzymes / trasporters Limitations Other levels of regulation Co-regulation does not mean necessarily… One vs. all analysis for At5g57800 CER1 protein, putative (WAX2) (Log) At5g57800 At5g20270 At2g26250 At3g43720 At1g17840 At1g68530 At4g39330 At2g26910 At5g13400 At4g25960 At5g14410 At1g02205 At1g51500 At2g04570 At5g57800 1 0.8287 0.8044 0.7894 0.7892 0.7864 0.7792 0.7755 0.7735 0.7679 0.766 0.7563 0.7379 0.7234 CER1 protein, putative (WAX2) expressed protein beta-ketoacyl-CoA synthase family (FIDDLEHEAD) (FDH) protease inhibitor/seed storage/lipid transfer protein (LTP) family protein ABC transporter family protein very-long-chain fatty acid condensing enzyme (CUT1) mannitol dehydrogenase, putative ABC transporter family protein proton-dependent oligopeptide transport (POT) family protein multidrug resistance P-glycoprotein, putative expressed protein CER1 protein (another?) ABC transporter family protein GDSL-motif lipase/hydrolase family protein CUT1 (very-long-chain fatty acid condensing enzyme, At1g68530) shows good correlation with At1g51500 (R=0.815), an ABC transporter protein Transporters WT cer5 cer5 Wax analyses of Arabidopsis stem surface (cuticle) or epidermal peel extracts (total epidermis). Cer5 (At1g51500) Pighin et al., Science (2004) 306:622-625 Programma Ripasso di cinetica enzimatica e approccio classico al controllo dei flussi [1,6]. Fondamenti di Analisi del Controllo Metabolico (MCA): proprietà locali e sistemiche, elasticità e coefficienti di controllo del flusso e delle concentrazione [1,6,7]. Trattazione dei sistemi Supply-Demand in generale [8] e dell’ATP in particolare [9]. Rate limiting steps e ingegneria metabolica [10, 11 e 12]. Tipi di ingegneria metabolica: a- Inattivazione di enzimi e allergeni (via del gossipolo [13], ODAP e glucosidi cianogenici) e review generale [14]); bCreazione di vie metaboliche ex novo o potenziamento di vie endogene già presenti (Glucosidi cianogenici [15,16], Vitamina E [17, 18], Folato [19], laurato [20, 21]); c- Aumento del demand (aumento del contenuto in aa, aumento del contenuto in zucchero) [22-24]; e- Amido in patata: strategie diverse [25]; fUtilizzo dei fattori di trascrizione (Terpenoid Indole Alkaloyd, Flavonoidi, cuticola, glucosinolati...) [10,11,26]. Bibliografia (ref 2-4 sono testi generali sul metabolismo delle piante e la sua manipolazione) Generali (MCA e metabolismo): [1] Fell, Understanding the control of Metabolism Portland Press (1997) (in Biblioteca biologica) [2] Dennis/Turpin Plant Metabolism (1998) Longman; nuova edizione. [3] Lea/Leegood Plant Biochemistry and Molecular Biology (1993) Wiley & sons. [4] Foyer e Quick (Eds) A molecular approach to primary metabolism in higher plants; Taylor and Francis (1997) Articoli originali [6] Kacser, Burns, & Fell, The control of flux (1995) Biochem. Soc. Trans. 23, 341-366 (art. del 1973). [7] Kacser e Acerenza, Eur. J. Biochem. (1993) 216:361-367 [8] Hofmeyr & Cornish-Bowden (2000) Regulating the cellular economy of supply and demand. FEBS Lett. 476:47-51. [9] Koebmann et al. (2002) The glycolytic flux in Escherichia coli is controlled by the demand for ATP. J. Bacteriol. 184:3909-16 [10] Morandini & Salamini (2003) Plant biotechnology and Breeding, allied for years to come Trends Pl. Sci. 8:70-5. [11] Morandini, Salamini & Gantet, (2005) Engineering of Plant Metabolism for Drug and Food. Curr. Med. Chem. – Immun., Endoc. & Metab. Agents 5:103-112 [12] Morandini (2009) Rethinking metabolic control. Plant Science 176:441-451 [13] Sunilkumar et al., (2005) Engineering cottonseed for use in human nutrition by tissue-specific reduction of toxic gossypol. P.N.A.S. 103:18054–18059. [14] Morandini (2010) Inactivation of allergens and toxins. N Biotechnol. 27:482-93. [15] Tattersall DB et al., (2001) Resistance to an herbivore through engineered cyanogenic glucoside synthesis. Science 293:1826-8. [16] Nielsen et al., (2008) Metabolon formation in dhurrin biosynthesis. Phytochemistry 69:88-98. [17] DellaPenna D. (2005) Progress in the dissection and manipulation of vitamin E synthesis. Trends Plant Sci 10:574-9. [18] Valentin (2006) The Arabidopsis vitamin E pathway gene5-1 mutant reveals a critical role for phytol kinase in seed tocopherol biosynthesis. Plant Cell. 18:212-24. [19] Hossain et al. (2004) Enhancement of folates in plants through metabolic engineering. Proc Natl Acad Sci USA 101:5158–5163. [20] Knutzon et al., (1999) LPAAT from coconut endosperm mediates the insertionof laurate at the sn-2 position of triacylglycerols in Lauric rapeseed oil and can increase total laurate levels. Plant Physiology 120:739746. [21] Thelen JJ, Ohlrogge JB. (2002) Metabolic engineering of fatty acid biosynthesis in plants. Metab Eng. 4:12-21. [22] Chong et al. (2007) Growth and metabolism in sugarcane are altered by the creation of a new hexosephosphate sink. Plant Biotechnol J. 5:240-53. [23] Wu (2007) Doubled sugar content in sugarcane plants modified to produce a sucrose isomer. Pl. Biotech. J. 5:109-17. [24] Basnayake S. (2012) Field performance of transgenic sugarcane expressing isomaltulose synthase. Plant Biotechnology Journal 10:217-225 [25] Geigenberger et al., (2004) Metabolic control analysis and regulation of the conversion of sucrose to starch in growing potato tubers. Plant, Cell and Environment 27:655–673. [26] Broun P. (2004) Transcription factors as tools for metabolic engineering in plants. Curr Opin Plant Biol. 7:202-9. In rosso sono evidenziati quelli da leggere con attenzione ai fini dell’esame. Ulteriori riferimenti bibliografici si trovano nei singoli file di powerpoint delle lezioni. Chiunque desiderasse gli articoli originali basta me li chieda. Cosa è naturale? L’uomo fa parte della natura? Da cosa viene la specialità dell’uomo? Su cosa si fonda? Gli esseri umani e la tecnologia sono una cosa sola? Un compito... Il docente universitario ha il compito non solo di indagare la verità e di suscitarne perenne stupore, ma anche di promuoverne la conoscenza in ogni sfaccettatura e di difenderla da interpretazioni riduttive e distorte. Porre al centro il tema della verità non è un atto meramente speculativo, ristretto a una piccola cerchia di pensatori; al contrario, è una questione vitale per dare profonda identità alla vita personale e suscitare la responsabilità nelle relazioni sociali. Di fatto, se si lascia cadere la domanda sulla verità e la concreta possibilità per ogni persona di poterla raggiungere, la vita finisce per essere ridotta ad un ventaglio di ipotesi, prive di riferimenti certi. BENEDETTO XVI Pontificia Università Lateranense, Sabato, 21 ottobre 2006 http://www.vatican.va/holy_father/benedict_xvi/speeches/2006/october/documents/hf_ben-xvi_spe_20061021_lateranense_it.html