Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
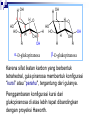
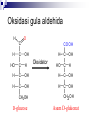
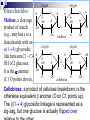
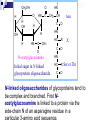
SATUAN ACARA DAN JADUAL KULIAH BIOKIMIA No. Topik Perkuliahan Tanggal Pengajar 1. Pendahuluan Konsep dasar biokimia Reaksi-reaksi biokimia 30-08-2005 Drs. Winarto Hariadi, M.Si. 2. Air dan Buffer 06-09-2005 Drs. Winarto Hariadi, M.Si. 3. Karbohidrat I Tinjauan umum Monosakarida Disakarida Polisakarida 13-09-2005 Dr. Ir. Arman Wijonarko, M.Sc. 4. Karbohidrat II Reaksi monosakarida Ikatan glikosida Fungsi karbohidrat 20-09-2005 Dr. Ir. Arman Wijonarko, M.Sc. SATUAN ACARA DAN JADUAL KULIAH BIOKIMIA No. Topik Perkuliahan Tanggal Pengajar 5. Asam Amino dan Protein I Tinjauan umum Asam amino Biosintesis asam amino 27-09-2005 Dr. Ir. Arman Wijonarko, M.Sc. 6. Asam Amino dan Protein II Peptida Struktur protein Fungsi asam amino dan protein Biosintesis protein 04-10-2005 Dr. Ir. Arman Wijonarko, M.Sc. 7. Lipida I Tinjauan umum Asam lemak jenuh dan tak jenuh Reaksi asam lemak 11-10-2005 Drs. Winarto Hariadi, M.Si. 8. Lipida II Fungsi asam lemak dan lipida Biosintesis asam lemak 18-10-2005 Drs. Winarto Hariadi, M.Si. 9. UJIAN SISIPAN 25-10-2005 Topik 1 s/d 8 SATUAN ACARA DAN JADUAL KULIAH BIOKIMIA No. Topik Perkuliahan Tanggal Pengajar 10. Asam nukleat I Tinjauan umum Nukleosida dan nukleotida 15-11-2005 Ir. Sedyo Hartono, M.P., Ph.D. 11. Asam nukleat II Struktur DNA dan RNA Nukleosida dan nukleotida 15-11-2005 Ir. Sedyo Hartono, M.P., Ph.D. 12. Enzim I Tinjauan umum Klasifikasi enzim Koenzim dan kofaktor 22-11-2005 Ir. Irfan D. Prijambada, M.Eng., Ph.D. 13. Enzim II Mekanisme dan kinetika kerja enzim Penghambatan kerja enzim 29-11-2005 Ir. Irfan D. Prijambada, M.Eng., Ph.D. SATUAN ACARA DAN JADUAL KULIAH BIOKIMIA No. Topik Perkuliahan Tanggal Pengajar 14. Metabolisme I Tinjauan umum Jalur metabolisme 06-12-2005 Ir. Irfan D. Prijambada, M.Eng., Ph.D. 15. Metabolisme II Bioenergetika Pengendalian metabolisme 06-12-2005 Ir. Irfan D. Prijambada, M.Eng., Ph.D. Mengikuti jadual Fakultas Topik 10 s/d 15 16. UJIAN AKHIR KARBOHIDRAT II * Reaksi monosakarida * Ikatan glikosida * Fungsi karbohidrat Irfan D. Prijambada, Ph.D. Lab. Mikrobiologi Tanah dan Lingkungan, Fakultas Pertanian UGM Monosakarida Memiliki atom karbon 3 sampai 7 Setiap atom karbon memiliki gugus hidroksil, keton atau aldehida. Setiap molekul monosakarida memiliki 1 gugus keton atau 1 gugus aldehida Gugus aldehida selalu berada di atom C pertama Gugus keton selalu berada di atom C kedua Monosakarida Ketosas (mis: fruktosa) biasanya Aldosa (mis: glukosa) memiliki gugus aldehida pada salah satu memiliki gugus keto pada atom C2. ujungnya. H O CH2OH C C O HO C H OH H C OH OH H C OH H C OH HO C H H C H C CH2OH CH2OH D-glucose D-fructose Notasi D vs L CH O Notasi D & L dilakukan karena adanya atom C H C OH dengan konfigurasi CH2OH asimetris seperti pada D-gliseraldehida gliseraldehida. CH O HO H C OH CH2OH D-glyceraldehyde H CH2OH L-gliseraldehida CH O Penampilan dalam bentuk gambar bagian bawah disebut Proyeksi Fischer. C CH O HO C H CH2OH L-gliseraldehida Penamaan Gula Untuk gula dengan atom C asimetrik lebih dari 1, notasi D atau L ditentukan oleh atom C asimetrik terjauh dari gugus aldehida atau keto. Gula yang ditemui di alam adalah dalam bentuk isomer D. O H C H – C – OH HO – C – H H – C – OH H – C – OH CH2OH D-glukosa O H C HO – C – H H – C – OH HO – C – H HO – C – H CH2OH L-glukosa Gula dalam bentuk D merupakan bayangan cermin dari gula dalam bentuk L. Kedua gula tersebut memiliki nama yang sama, misalnya Dglukosa & L-glukosa. O H C H – C –OH HO – C –H H – C – OH H – C – OH CH2OH D-glukosa O H C HO – C – H H – C – OH HO – C – H HO – C – H CH2OH L-glukosa Stereoisomers lainnya memiliki names yang unik, misalnya glukosa, manosa, galaktosa, dll. Jumlah stereoisomer adalah 2n, dengan n adalah jumlah pusat asimetrik. Aldosa dengan 6-C memiliki 4 pusat asimetrik, oleh karenanya memiliki 16 stereoisomer (8 gula berbentuk D dan 8 gula berbentuk L). Pembentukan hemiasetal & hemiketal Aldehida dapat bereaksi dengan alkohol membentuk hemiasetal. Keton dapat bereaksi dengan alkohol membentuk hemiketal. H C H O + R' OH R' O R OH R aldehida alkohol hemiasetal R C C R O + "R OH R' keton "R O C R' alkohol hemiketal OH Pentosa dan heksosa dapat membentuk struktur siklik melalui reaksi gugus keton atau aldehida dengan gugus OH dari atom C asimetrik terjauh. Glukosa membentuk hemiasetal intramolekular sebagai hasil reaksi aldehida dari C1 & OH dari atom C5, dinamakan cincin piranosa. 1 H HO H H 2 3 4 5 6 CHO C OH C H D-glukosa C OH (bentuk linier) C OH CH2OH 6 CH2OH 6 CH2OH 5 H 4 OH H OH 3 H O H H 1 2 OH a-D-glukosa OH 5 H 4 OH H OH 3 H O OH H 1 2 H OH b-D-glukosa Penampilan dalam bentuk gula siklik disebut proyeksi Haworth. CH2OH 1 HO H H 2C O C H C OH C OH 3 4 5 HOH2C 6 6 CH2OH D-fruktosa (linear) H 5 H 1 CH2OH O 4 OH HO 2 3 OH H a-D-fruktofuranosa Fruktosa dapat membentuk Cincin piranosa, melalui reaksi antara gugus keto atom C2 dengan OH dari C6. Cincin furanosa, melalui reaksi antara gugus keto atom C2 dengan OH dari C5. 6 C H OH 6 C H2OH 5 H 4 OH H OH 3 H 2 O H H 1 2 OH a-D-glukosa OH 5 H 4 OH H OH 3 H O OH H 1 2 H OH b -D-glukosa Pembentukan cincin siklik glukosa menghasilkan pusat asimetrik baru pada atom C1. Kedua stereoisomer disebut anomer, a & b. Proyeksi Haworth menunjukkan bentuk cincin dari gula dengan perbedaan pada posisi OH di C1 anomerik : a (OH di bawah struktur cincin) b (OH di atas struktur cincin). H OH H OH 4 6 H O HO HO H O HO H HO 5 3 H H 2 H OH 1 OH a-D-glukopiranosa H OH OH H b-D-glukopiranosa Karena sifat ikatan karbon yang berbentuk tetrahedral, gula piranosa membentuk konfigurasi “kursi" atau “perahu", tergantung dari gulanya. Penggambaran konfigurasi kursi dari glukopiranosa di atas lebih tepat dibandingkan dengan proyeksi Haworth. Turunan gula CH O COOH CH2OH H C OH H C OH HO C H H C OH HO C H H C OH H C OH H C OH H C OH H C OH H C OH CH2OH D-ribitol CH2OH Asam D-glukonat COOH Asam D-glukuronat Gula alkohol – tidak memiliki gugus aldehida atau ketone; misalnya ribitol. Gula asam –gugus aldehida pada atom C1, atau OH pada atom C6, dioksidasi membentuk asam karboksilat; misalnya asam glukonat, asam glukuronat. Oksidasi gula aldehida H O C CO O H H C OH HO C H H C H C H C OH HO C H OH H C OH OH H C OH CH2OH D-glucose Oksidator CH2 O H Asam D-glukonat Oksidasi gula aldehida Gula yang dapat dioksidasi adalah senyawa pereduksi. Gula yang demikian disebut sebagai gula pereduksi. Senyawa yang sering digunakan sebagai pengoksidasi adalah ion Cu+2, yang berwarna biru cerah, yang akan tereduksi menjadi ion Cu+, yang berwarna merah kusam. Hal ini menjadi dasar bagi pengujian Benedict yang digunakan untuk menentukan keberadaan glukosa dalam urin, suatu pengujian bagi diagnosa diabetes. Oksidasi gula aldehida Glukosa + Cu++ panas & alk . pH Gluconic acid + Cu2O (Cu2O is insol ppt) glukosa oksidase Glukosa + O2 Asam glukonat + H2O2 (H2O2 nya diukur) Glukosa + ATP heksokinase Glukosa-6-P + ADP (G-6-Pnya diukur) Turunan gula CH 2OH CH2OH O H H OH H H OH H OH OH H O H NH 2 a-D-glukosamina H H O OH OH H N C CH 3 H a-D-N-asetilglukosamina Gula amino - gugus amino menggantikan gugus hidroksil. Sebagai contoh glukosamina. Gugus amino dapat mengalami asetilasi, seperti pada N-asetilglukosamina. Ikatan Glikosida Gugus hidroksil anomerik dan gugus hidroksil gula atau senyawa yang lain dapat membentuk ikatan yang disebut ikatan glikosida dengan membebaskan air : R-OH + HO-R' R-O-R' + H2O Misalnya methanol bereaksi dengan gugus OH anomerik dari glukosa membentuk metil glukosida (metil-glukopiranosa). H OH H OH H2O H O HO HO H H H + CH3- OH H O HO HO H OH H OH a-D-glukopiranosa metanol H OH OCH3 Metil-a-D-glukopiranosa Disaccharides: Maltose, a cleavage product of starch (e.g., amylose), is a disaccharide with an a(1 4) glycosidic link between C1 - C4 OH of 2 glucoses. It is the a anomer (C1 O points down). 6 CH2OH 6 CH2OH H 5 O H OH 4 OH 3 H H H 1 H 4 4 maltose OH H H H H H 1 OH 2 OH 1 O 4 5 O H OH H H 3 H 6 CH2OH O H OH H OH 3 OH 5 O O 2 6 CH2OH H 5 2 OH 3 cellobiose H OH 1 2 OH Cellobiose, a product of cellulose breakdown, is the otherwise equivalent b anomer (O on C1 points up). The b(1 4) glycosidic linkage is represented as a zig-zag, but one glucose is actually flipped over H Other disaccharides include: Sucrose, common table sugar, has a glycosidic bond linking the anomeric hydroxyls of glucose & fructose. Because the configuration at the anomeric C of glucose is a (O points down from ring), the linkage is a(12). The full name of sucrose is a-D-glucopyranosyl(12)-b-D-fructopyranose.) Lactose, milk sugar, is composed of galactose & glucose, with b(14) linkage from the anomeric OH of galactose. Its full name is b-D- Polysaccharides CH2OH H O H OH H H H 1 O OH 6CH OH 2 5 O H 4 OH 3 H OH H H H H 1 O H OH CH2OH CH2OH CH2OH H H H O H OH H O O H H O H OH H H O OH 2 OH H OH H OH H OH amylose Plants store glucose as amylose or amylopectin, glucose polymers collectively called starch. Glucose storage in polymeric form minimizes osmotic effects. Amylose is a glucose polymer with a(14) linkages. It adopts a helical conformation. The end of the polysaccharide with an anomeric C1 not involved in a glycosidic bond is called the reducing end. CH2OH CH2OH O H H OH H H OH H O OH CH2OH H H OH H H OH H H OH CH2OH O H OH O H OH H H O O H OH H H OH H H O 4 amylopectin H 1 O 6 CH2 5 H OH 3 H CH2OH O H 2 OH H H 1 O CH2OH O H 4 OH H H H H O OH O H OH H H OH H OH Amylopectin is a glucose polymer with mainly a(14) linkages, but it also has branches formed by a(16) linkages. Branches are generally longer than shown above. The branches produce a compact structure & provide multiple chain ends at which enzymatic cleavage can CH2OH CH2OH O H H OH H H OH H O OH CH2OH H H OH H H OH H H OH CH2OH O H OH O H OH H H O O H OH H H OH H H O 4 glycogen H 1 O 6 CH2 5 H OH 3 H CH2OH O H 2 OH H H 1 O CH2OH O H 4 OH H H H H O OH O H OH H H OH H OH Glycogen, the glucose storage polymer in animals, is similar in structure to amylopectin. But glycogen has more a(16) branches. The highly branched structure permits rapid release of glucose from glycogen stores, e.g., in muscle during exercise. The ability to rapidly mobilize glucose is more essential to animals than to plants. CH2OH H O H OH H OH H 1 O H H OH 6CH OH 2 5 O H 4 OH 3 H H H 1 2 OH O O H OH CH2OH CH2OH CH2OH H H O O H OH H OH O H O H OH H OH OH H H H H H H H OH cellulose Cellulose, a major constituent of plant cell walls, consists of long linear chains of glucose with b(14) linkages. Every other glucose is flipped over, due to the b linkages. van der Waals interactions, that cause and inter-chain H-bonds This promotes intra-chain cellulose chains to be and straight & rigid, and pack with a crystalline arrangement in thick Schematic of arrangement of bundles called cellulose chains in a microfibril. CH2OH H O H OH H OH H 1 O H H OH 6CH OH 2 5 O H 4 OH 3 H H H 1 2 OH O O H OH CH2OH CH2OH CH2OH H H O O H OH H OH O H O H OH H OH OH H H H H H H H OH cellulose Multisubunit Cellulose Synthase complexes in the plasma membrane spin out from the cell surface microfibrils consisting of 36 parallel, interacting cellulose chains. These microfibrils are very strong. The role of cellulose is to impart strength and rigidity to plant cell walls, which can withstand high hydrostatic pressure gradients. Osmotic swelling is prevented. CH 2OH D-glucuronate 6 6COO H 4 5 H OH 3 H H 2 OH 1 H H OH O O H 4 O H 5 3 H 2 1 O H NHCOCH 3 N-acetyl-D-glucosamine hyaluronate Glycosaminoglycans (mucopolysaccharides) are polymers of repeating disaccharides. Within the disaccharides, the sugars tend to be modified, with acidic groups, amino groups, sulfated hydroxyl and amino groups, etc. Glycosaminoglycans tend to be negatively charged, because of the prevalence of acidic CH 2OH D-glucuronate 6 6COO H 4 5 H OH 3 H H 2 OH 1 H H OH O O H 4 O H 5 3 H 2 1 O H NHCOCH 3 N-acetyl-D-glucosamine hyaluronate Hyaluronate is a glycosaminoglycan with a repeating disaccharide consisting of 2 glucose derivatives, glucuronate (glucuronic acid) & N-acetylglucosamine. The glycosidic linkages are b(13) & b(14). CH 2OH D-glucuronate 6COO H 4 6 5 H OH 3 H H 2 OH 1 H H OH O O H 4 O H 5 3 H 2 1 O H NHCOCH 3 N-acetyl-D-glucosamine hyaluronate Proteoglycans are glycosaminoglycans that are covalently linked to specific core proteins. Some proteoglycans of the extracellular matrix in turn link non-covalently to hyaluronate via protein domains called link modules. CH 2OH D-glucuronate 6COO H 4 6 5 H OH 3 H H 2 OH 1 H H OH O O H 4 O H 5 3 H 2 1 O H NHCOCH 3 N-acetyl-D-glucosamine hyaluronate For example, in cartilage multiple copies of the aggrecan proteoglycan bind to an extended hyaluronate backbone to form a large complex. Versican, another proteoglycan that binds to hyaluronate, is in the extracellular matrix of loose connective tissues. See web sites on aggrecan and aggrecan plus N-sulfo-glucosamine-6-sulfate iduronate-2-sulfate CH2OSO3 H H COO OH O O H O H H OH H H H H OSO3 O H NHSO3 heparin or heparan sulfate - examples of residues Heparan sulfate is initially synthesized on a membrane-embedded core protein as a polymer of alternating N-acetylglucosamine and glucuronate residues. Later, in segments of the polymer, glucuronate residues may be converted to the sulfated sugar iduronic acid, while N-acetylglucosamine residues PDB 1RID Heparin, a soluble glycosaminoglycan found in granules of mast cells, has a structure similar to that of heparan sulfates, but is more highly sulfated. When released into the blood, it inhibits clot formation by interacting heparin: (IDS-SGN)5 with the protein antithrombin. C O N S Heparin has an extended helical Charge repulsion by the many negatively charged conformation. groups may contribute to this conformation. Heparin shown has 10 residues, alternating IDS (iduronate-2-sulfate) & SGN (N-sulfo-glucosamine-6- core protein heparan sulfate glycosaminoglycan transmembrane a-helix cytosol Some cell surface heparan sulfate glycosaminoglycans remain covalently linked to core proteins embedded in the plasma membrane. Proteins involved in signaling & adhesion at the cell surface recognize and bind segments of heparan sulfate chains having particular patterns of sulfation. CH2OH C O CH2 CH O O Oligosaccharides H H NH serine that are covalently H OH residue attached to OH O H proteins or to H HN C CH3 membrane lipids b-D-N-acetylglucosamine may be linear or branched chains. O-linked oligosaccharide chains of glycoproteins vary in complexity. They link to a protein via a glycosidic bond between a sugar residue & a serine or threonine OH. O-linked oligosaccharides have roles in recognition, interaction, and enzyme regulation. C CH2OH O H H OH O CH2 CH NH H O serine residue O H OH H HN C CH3 b-D-N-acetylglucosamine N-acetylglucosamine (GlcNAc) is a common Olinked glycosylation of protein serine or threonine residues. Many cellular proteins, including enzymes & transcription factors, are regulated by reversible GlcNAc attachment. Often attachment of GlcNAc to a protein OH CH2OH O O H H OH HN C HN CH2 C H H OH H HN C CH3 O N-acetylglucosamine Initial sugar in N-linked glycoprotein oligosaccharide Asn CH O HN HC R C O X HN HC R C O Ser or Thr N-linked oligosaccharides of glycoproteins tend to be complex and branched. First Nacetylglucosamine is linked to a protein via the side-chain N of an asparagine residue in a NAN NAN NAN Gal Gal Gal NAG NAG NAG Man Man Man Key: NAG NAG Asn N-linked oligosaccharide Fuc NAN = N-acetylneuraminate Gal = galactose NAG = N-acetylglucosamine Man = mannose Fuc = fucose Additional monosaccharides are added, and the Nlinked oligosaccharide chain is modified by removal and addition of residues, to yield a characteristic branched structure. Many proteins secreted by cells have attached Nlinked oligosaccharide chains. Genetic diseases have been attributed to deficiency of particular enzymes involved in synthesizing or modifying oligosaccharide chains of these glycoproteins. Such diseases, and gene knockout studies in mice, have been used to define pathways of modification of oligosaccharide chains of glycoproteins and glycolipids. Carbohydrate chains of plasma membrane glycoproteins and glycolipids usually face the outside of the cell. Lectins are glycoproteins that recognize and bind to specific oligosaccharides. A few examples: Concanavalin A and wheat germ agglutinin are plant lectins that have been useful research tools. Mannan-binding lectin (MBL) is a glycoprotein found in blood plasma. It associates with cell surface carbohydrates of disease-causing microorganisms, promoting phagocytosis of these organisms as part of the Selectins are integral proteins of mammalian cell plasma membranes with roles in cell-cell recognition & binding. A lectin-like domain is at the end of an extracellular segment that extends out from the cell surface. selectin lectin domain outside cytosol transmembrane a-helix cytoskeleton binding domain A cleavage site just outside the transmembrane ahelix provides a mechanism for regulated release of some lectins from the cell surface. A cytosolic domain participates in regulated interaction with the actin cytoskeleton.











































![Zuccheri [chimica]](http://s1.studyres.com/store/data/004545645_1-6bd8c4a2d3b0941e429f1520d29dd13d-150x150.png)