Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
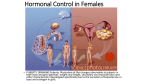
32 Animal Reproduction Concept 32.1 Reproduction Can Be Sexual or Asexual Asexual reproduction: requires no mating does not result in genetic diversity Asexually reproducing species are mostly invertebrates, sessile, and live in constant environments. Three types of asexual reproduction are budding, regeneration, and parthenogenesis. Concept 32.1 Reproduction Can Be Sexual or Asexual Budding produces new individuals that form from the bodies of older animals. A bud grows by mitotic cell division—cells differentiate before the bud breaks away. The bud is genetically identical to the parent. Figure 32.1 Three Forms of Asexual Reproduction (Part 1) Concept 32.1 Reproduction Can Be Sexual or Asexual Regeneration can replace damaged tissue or form a complete individual. Example: Echinoderms. Figure 32.1 Three Forms of Asexual Reproduction (Part 2) Concept 32.1 Reproduction Can Be Sexual or Asexual Parthenogenesis is the development of offspring from unfertilized eggs. Parthenogenesis may determine the sex of the offspring. In some species, females can act as males depending on cyclic states of estrogen and progesterone. Figure 32.1 Three Forms of Asexual Reproduction (Part 3) Concept 32.1 Reproduction Can Be Sexual or Asexual Sexual Reproduction: • two haploid cells, gametes, form a diploid individual Three fundamental steps of sexual reproduction: • Gametogenesis—making haploid gametes • Spawning or mating—getting gametes together • Fertilization—fusing gametes to form a diploid Concept 32.1 Reproduction Can Be Sexual or Asexual Sexual reproduction has a big advantage— the generation of genetic diversity. Meiosis allows genetic diversity through crossing over between homologous chromosomes and independent assortment. Concept 32.2 Gametogenesis Produces Haploid Gametes Gametogenesis occurs in the gonads—the testes in males and ovaries in females Male gametes, the sperm, move by flagella. The larger gametes of females are the ova, or eggs, and are nonmotile. Gametes are produced from germ cells— present early in development and distinct from other (somatic) cells of the body. Concept 32.2 Gametogenesis Produces Haploid Gametes Germ cells migrate to the gonads when they begin to form. Embryonic germ cells divide by mitosis to form diploid spermatogonia in males and oogonia in females. These multiply by mitosis, producing primary spermatocytes and primary oocytes—these enter meiosis and produce haploid gametes, sperm and ova. Concept 32.2 Gametogenesis Produces Haploid Gametes The production of sperm is spermatogenesis—of ova is oogenesis. The first meiotic division of a primary spermatocyte results in two secondary spermatocytes. The second division produces four haploid spermatids, which will each become a sperm. Figure 32.2 Gametogenesis (Part 1) Concept 32.2 Gametogenesis Produces Haploid Gametes A man produces over 100 million sperm per day. Concept 32.2 Gametogenesis Produces Haploid Gametes In hermaphroditic species such as earthworms, a single individual may produce sperm and ova simultaneously. An anemone fish produces sperm and ova sequentially and may function as a male or a female at different times. Concept 32.3 Fertilization Is the Union of Sperm and Ovum Fertilization is the union of a haploid sperm and a haploid egg. It creates a single diploid cell, called a zygote, which will develop into an embryo. Fertilization involves a complex series of events. Concept 32.3 Fertilization Is the Union of Sperm and Ovum Steps in fertilization: • Recognition and binding of sperm and ovum • Activation of sperm • Plasma membranes fuse • Additional sperm entry blocked • Activation of ovum • Ovum and sperm nuclei fuse Concept 32.3 Fertilization Is the Union of Sperm and Ovum Aquatic animals bring gametes together through spawning—gamete release before external fertilization occurs. Internal fertilization occurs when sperm is released directly into the female reproductive tract. Internal fertilization requires accessory sex organs, such as penis and vagina. Copulation is the joining of the male and female accessory organs. Concept 32.3 Fertilization Is the Union of Sperm and Ovum Species-specific sperm and ovum interactions are controlled by specific recognition molecules. Ova of aquatic species release chemical attractants to cause sperm to swim toward the ovum. Sperm must go through two protective layers to reach an ovum—a jelly coat and the vitelline envelope. Concept 32.3 Fertilization Is the Union of Sperm and Ovum The acrosome is a membrane-enclosed structure on the sperm head. Egg and sperm contact causes substances in the jelly coat to trigger an acrosomal reaction. Membranes in the sperm head and acrosome break down; enzymes are released and digest the jelly coat. Figure 32.3 Fertilization of Sea Urchin Egg (Part 2) Concept 32.3 Fertilization Is the Union of Sperm and Ovum An acrosomal process extends from the head of the sperm. The acrosomal process is coated with bindin—specific recognition molecules. Bindin acts on bindin receptors in the vitelline envelope. Sperm and egg plasma membranes fuse to form a fertilization cone. Concept 32.3 Fertilization Is the Union of Sperm and Ovum Internal fertilization involves species-specific mating behaviors and ovum-sperm recognition mechanisms. The ovum is surrounded by the cumulus, cells in a gelatinous matrix. The zona pellucida, or zona, is a glycoprotein envelope beneath the cumulus. Concept 32.3 Fertilization Is the Union of Sperm and Ovum When sperm make contact with the zona, species-specific glycoproteins bind to recognition molecules on the sperm. Binding triggers the acrosomal reaction, and enzymes digest the zona pellucida. When sperm reaches ovum membrane other proteins facilitate membrane fusion. Figure 32.3 Fertilization of Sea Urchin Egg (Part 1) Figure 32.4 Barriers to Mammalian Sperm Concept 32.3 Fertilization Is the Union of Sperm and Ovum Fusion and entry of a sperm into the ovum lead to: Blocks to polyspermy—mechanisms to prevent more than one sperm from entering an ovum Concept 32.3 Fertilization Is the Union of Sperm and Ovum Fast block to polyspermy: • Transient (die quickly) • Caused by change in membrane potential as sodium ions (Na+) enter plasma membrane of ovum after contact with a sperm Concept 32.4 Human Reproduction Is Hormonally Controlled Sperm are produced in the paired male gonads, or testes. The testes are located in the scrotum, outside of the body, to maintain optimal temperature for sperm production. Semen is made up of sperm and other fluids and molecules. Figure 32.5 Reproductive Organs of the Human Male (Part 1) Concept 32.4 Human Reproduction Is Hormonally Controlled Spermatogenesis occurs in the seminiferous tubules in each testis. Between the tubules are Leydig cells, which produce testosterone. Spermatogonia reside in the outermost regions of the tubules, near Sertoli cells, which provide nutrients. Figure 32.6 Spermatogenesis Takes Place in the Seminiferous Tubules (Part 1) Figure 32.6 Spermatogenesis Takes Place in the Seminiferous Tubules (Part 2) Figure 32.6 Spermatogenesis Takes Place in the Seminiferous Tubules (Part 3) Concept 32.4 Human Reproduction Is Hormonally Controlled Immature sperm cells are shed in the lumen of the seminiferous tubule. They move into the epididymis, mature, and become motile. Sperm travel in the vas deferens which joins with the semen-carrying ejaculatory duct. This joins the urethra, the common final duct, at the base of the penis and opens to the outside at the tip of the penis. Figure 32.5 Reproductive Organs of the Human Male (Part 2) Concept 32.4 Human Reproduction Is Hormonally Controlled Besides sperm, semen contains seminal fluids—the products of several accessory glands: • The paired seminal vesicles, the prostate gland, and the bulbourethral glands The prostate gland produces a fluid that is alkaline and reduces acidity in male and female reproductive tracts. The fluid also contains enzymes to thicken semen and later to dissolve it. Concept 32.4 Human Reproduction Is Hormonally Controlled The bulbourethral glands produce an alkaline secretion that: • Neutralizes acidity in the urethra • Provides lubrication and facilitates sperm movement during climax These secretions precede climax yet carry residual sperm capable of fertilization. Pregnancy can occur even when the penis is withdrawn prior to ejaculation (coitus interruptus). Concept 32.4 Human Reproduction Is Hormonally Controlled Male copulatory organ is the penis. The sensitive tip of the penis, the glans penis, is covered by the foreskin—removal of the foreskin is circumcision. Sexual stimulation triggers the nervous system to produce penile erection. Nitric oxide (NO) acts on blood vessels by stimulating production of cGMP. cGMP causes dilation of blood vessels so that spongy erectile tissue fills with blood. Concept 32.4 Human Reproduction Is Hormonally Controlled At the climax of copulation, semen is ejaculated through the vasa deferentia and urethra. Ejaculation is accompanied by feelings of intense pleasure called orgasm. After ejaculation NO release decreases and enzymes break down cGMP—blood vessels are no longer compressed, and erection declines. Concept 32.4 Human Reproduction Is Hormonally Controlled Hormones control male sexual function: GnRH (gonadotropin-releasing hormone)— released by the hypothalamus at puberty GnRH increases the release of LH (luteinizing hormone) and FSH (folliclestimulating hormone) by the anterior pituitary. Figure 32.7 Male Reproductive Hormones Concept 32.4 Human Reproduction Is Hormonally Controlled LH increases testosterone: • Increases growth rate and starts development of secondary sexual characteristics FSH and testosterone control spermatogenesis in the Sertoli cells. Sertoli cells also produce inhibin, which exerts negative feedback on cells that produce and secrete FSH. And now here’s Alex with the Reproductive system! Concept 32.2 Gametogenesis Produces Haploid Gametes Oogenesis: A primary oocyte immediately begins prophase I of meiosis. Here, development stops in many species. The primary oocyte grows larger and acquires nutrients. When meiosis resumes, the nucleus of the oocyte divides into two daughter cells of unequal sizes. The cell with more cytoplasm is the secondary oocyte—the smaller one is the first polar body. Concept 32.2 Gametogenesis Produces Haploid Gametes The second meiotic division forms daughter cells of unequal sizes. One is a large ootid, which differentiates to become a mature ovum. The other forms the second polar body. Figure 32.2 Gametogenesis (Part 2) Concept 32.2 Gametogenesis Produces Haploid Gametes Few primary oocytes complete all meiotic stages—females produce far fewer gametes than males. The average woman has about 450 menstrual cycles and releases on ovum each time, until menopause—the end of fertility. Concept 32.3 Fertilization Is the Union of Sperm and Ovum Fertilized ova may be released into the environment or retained: Oviparity—egg laying: • Oviparous animals lay eggs in the environment, and embryos develop outside the mother’s body. Viviparity—live bearing: • Viviparous animals retain the embryo in the mother’s body during early development. Concept 32.3 Fertilization Is the Union of Sperm and Ovum Viviparity differs in mammals as they have a specialized female reproductive tract: • Uterus (or womb)—holds the embryo • Placenta—develops in the uterus and enables exchange of nutrients and waste Concept 32.4 Human Reproduction Is Hormonally Controlled In females, the ovary releases an ovum into one of the oviducts, or Fallopian tubes, where it may be fertilized. The ovum is propelled towards the uterus where it will develop if fertilized. The bottom of the uterus is the narrow cervix, which opens into the vagina. Concept 32.4 Human Reproduction Is Hormonally Controlled The female reproductive cycle is about 28 days and consists of two linked cycles: • The ovarian cycle that produces mature ova and hormones • The uterine, or menstrual, cycle that prepares the uterus for the arrival of an embryo. Concept 32.4 Human Reproduction Is Hormonally Controlled Each primary oocyte and its surrounding ovarian cells constitute a follicle. At the beginning of a cycle, the anterior pituitary increases FSH and LH. 6 to 12 follicles grow in the first two weeks of the cycle—the follicular phase. Follicles increase estrogen production until the largest follicle matures completely and others die (atresia). Figure 32.9 The Ovarian Cycle (Part 1) Concept 32.4 Human Reproduction Is Hormonally Controlled Estrogen exerts negative feedback on the pituitary early in the ovarian cycle. During days 12–14 estrogen becomes a positive feedback signal and causes a surge of LH and FSH. The LH surge triggers ovulation—the follicle ruptures and the oocyte is released from the ovary. Concept 32.4 Human Reproduction Is Hormonally Controlled Follicle has two types of cells: • Granulosa cells surround the developing oocyte and are stimulated by FSH. • Thecal cells enclose the whole follicle, produce androgens when stimulated by LH. Levels of circulating estrogen increase, which feeds back negatively to the hypothalamus and anterior pituitary. FSH and LH levels then fall. Figure 32.10 The Ovarian and Uterine Cycles (Part 1) Figure 32.10 The Ovarian and Uterine Cycles (Part 2) Figure 32.10 The Ovarian and Uterine Cycles (Part 3) Figure 32.10 The Ovarian and Uterine Cycles (Part 4) Concept 32.4 Human Reproduction Is Hormonally Controlled The follicle with the most FSH receptors survives. The other follicular cells form the corpus luteum, which remains in the ovary. The corpus luteum functions as an endocrine gland and produces estrogen and progesterone for about two weeks— the luteal phase. Progesterone causes the uterine lining, the endometrium, to thicken. Concept 32.4 Human Reproduction Is Hormonally Controlled If an embryo does not arrive within 2 weeks after ovulation, then endometrium breaks down. Menstruation is the sloughing off of the endometrium through the vagina. In other mammals, estrus, or sexual receptivity, corresponds to ovulation. Most other mammals do not menstruate; the uterine lining is reabsorbed. Concept 32.4 Human Reproduction Is Hormonally Controlled Different hormones control pregnancy. After fertilization, the zygote divides and moves toward the uterus. It attaches to the endometrium as a blastocyst and burrows in—implantation. A new cover layer of cells secretes human chorionic gonadatropin (hCG). hCG causes the corpus luteum to continue to produce estrogen and progesterone, and endometrium is maintained. Concept 32.4 Human Reproduction Is Hormonally Controlled Pregnancy tests use an antibody to detect hCG in urine. The placenta forms from combined maternal and embryonic tissues and secretes progesterone and estrogen. Both hormones prevent the pituitary from releasing gonadotropins, so the ovarian cycle ceases during pregnancy. Concept 32.4 Human Reproduction Is Hormonally Controlled The onset of labor is triggered by hormonal and mechanical stimuli: • Progesterone—inhibits uterine contractions • Estrogen—stimulates contractions The ratio shifts in favor of estrogen near the end of pregnancy. The onset of labor is marked by an increase in oxytocin—a powerful stimulant of uterine contraction—by mother and fetus. Concept 32.4 Human Reproduction Is Hormonally Controlled Mechanical stimuli: • Uterine stretching by fully grown fetus • Pressure on the cervix by the head These stimuli create a positive feedback loop—cause release of oxytocin which increases contractions and puts more pressure on the cervix. Concept 32.4 Human Reproduction Is Hormonally Controlled Between contractions in early labor, hormonal and mechanical stimuli cause the cervix to dilate. It becomes large enough for the baby to pass through, about 10 cm. The head enters the vagina and passage is assisted by mother bearing down and “pushing” with abdominal muscles. Concept 32.4 Human Reproduction Is Hormonally Controlled If the baby suckles at the breast immediately after birth, additional oxytocin is secreted. This causes the uterus to continue to contract and reduce in size and helps stop bleeding. Oxytocin also promotes bonding between mother and infant. Concept 32.5 Humans Use a Variety of Methods to Control Fertility Ways to prevent fertilization or implantation (conception) are referred to as contraception. The only failure-proof methods of preventing pregnancy are complete abstinence from sexual activity or gonad removal. Other methods vary in their failure rate. Table 32.1 Methods of Contraception (Part 1) Table 32.1 Methods of Contraception (Part 2) Table 32.1 Methods of Contraception (Part 3) Concept 32.5 Humans Use a Variety of Methods to Control Fertility An abortion is a termination of the pregnancy once the fertilized egg is implanted in the uterus. A spontaneous abortion is a miscarriage, and may occur before the pregnancy is known. Abortions through medical intervention may be for therapeutic reasons or for fertility control—the embryo and some of the endometrium are removed. Concept 32.5 Humans Use a Variety of Methods to Control Fertility Infertility is the inability of a couple to conceive a child—several treatments exist. Artificial insemination—physician positions sperm in woman’s reproductive tract Assisted reproductive technologies (ARTs)—remove unfertilized eggs, combine them with sperm outside the body, and return them for implantation • In vitro fertilization (IVF)—the first ART. Concept 33.1 Fertilization Activates Development In an unfertilized frog egg: • Vegetal hemisphere—the lower half of the egg, where nutrients are concentrated • Animal hemisphere—the opposite end of the egg, contains the haploid nucleus Cytoplasmic movement after fertilization is visible because of pigments. Concept 33.2 Cleavage Repackages the Cytoplasm of the Zygote Cleavage—a rapid series of cell division, but no cell growth. Embryo becomes a ball of small cells. Blastocoel—a central fluid-filled cavity that forms in the ball. The embryo becomes a blastula and its cells are called blastomeres. Concept 33.2 Cleavage Repackages the Cytoplasm of the Zygote Complete cleavage occurs in eggs with little yolk. Cleavage furrows divide the egg completely—blastomeres are of similar size. Figure 33.3 Some Patterns of Cleavage (Part 1) Concept 33.2 Cleavage Repackages the Cytoplasm of the Zygote Incomplete cleavage occurs in eggs with a lot of yolk when cleavage furrows do not penetrate. Discoidal cleavage is common in eggs with a dense yolk—the embryo forms as a blastodisc that sits on top of the yolk. Figure 33.3 Some Patterns of Cleavage (Part 2) Concept 33.2 Cleavage Repackages the Cytoplasm of the Zygote Superficial cleavage is a type of incomplete cleavage. A syncytium, a cell with many nuclei, forms. The plasma membrane grows inward around nuclei and forms cells. Figure 33.3 Some Patterns of Cleavage (Part 3) Concept 33.2 Cleavage Repackages the Cytoplasm of the Zygote Mammalian cleavage is slow. During the fourth division, cells separate into two groups: • Inner cell mass—becomes the embryo— cells are pluripotent and in culture are embryonic stem cells (ESCs) • Trophoblast—a sac that forms from the outer cells—secretes fluid and creates the blastocoel, with inner cell mass at one end Embryo is now called a blastocyst. Figure 33.4 A Human Blastocyst at Implantation (Part 1) Concept 33.2 Cleavage Repackages the Cytoplasm of the Zygote Blastomeres become determined— committed to specific development—at different times. In mosaic development each blastomere contributes certain aspects to the adult animal. In regulative development, cells compensate for any lost cells. Figure 33.5 Fate Map of a Frog Blastula Concept 33.3 Gastrulation Creates Three Tissue Layers The blastula is transformed into an embryo during gastrulation, through movement of cells. The embryo has multiple tissue layers and distinct body axes. During gastrulation three germ layers form—also called cell layers or tissue layers Concept 33.3 Gastrulation Creates Three Tissue Layers • Endoderm—innermost layer; becomes the lining of the digestive and respiratory tracts, pancreas, and liver • Ectoderm—outer germ layer; becomes the nervous system, the eyes and ears, and the skin • Mesoderm—middle layer; contains cells that migrate between the other layers; forms organs, blood vessels, muscle, and bones Concept 33.3 Gastrulation Creates Three Tissue Layers During gastrulation: • Vegetal hemisphere flattens as cells change shape • Vegetal pole bulges inward, invaginates; cells become endoderm and form the archenteron, or gut • Some cells migrate into the central cavity and become mesenchyme—cells of the mesoderm layer Concept 33.3 Gastrulation Creates Three Tissue Layers Filopodia form and adhere to the ectoderm; pull the archenteron by contracting The mouth forms where the archenteron meets the ectoderm. The blastopore is the opening of the invagination of the vegetal pole and becomes the anus. Figure 33.6 Gastrulation in Sea Urchins (Part 1) Concept 33.3 Gastrulation Creates Three Tissue Layers . Involution occurs as bottle cells move inward and create the dorsal lip. Cells from the animal hemisphere move toward the site of involution—epiboly. At end of gastrulation, embryo has three germ layers and dorsal-ventral and anterior-posterior organization Figure 33.7 Gastrulation in the Frog Embryo (Part 1) Figure 33.7 Gastrulation in the Frog Embryo (Part 3) Concept 33.3 Gastrulation Creates Three Tissue Layers Reptiles and birds: Gastrulation occurs in a flat disk of cells called the blastodisc. Some cells enter a fluid space between the blastodisc and the yolk and form the hypoblast—a continuous layer that will contribute to extraembryonic membranes. Overlying cells form the epiblast— becomes the embryo. Concept 33.3 Gastrulation Creates Three Tissue Layers Epiblast cells move toward the midline and form a ridge called the primitive streak. The primitive groove develops along the primitive streak—cells migrate through it and become endoderm and mesoderm. Hensen’s node is the equivalent of the amphibian dorsal lip and contains many signaling molecules. Concept 33.4 Neurulation Creates the Nervous System Gastrulation produces an embryo with three germ layers that will influence each other during development. During organogenesis, organs and organ systems develop simultaneously. Neurulation is the initiation of the nervous system—occurs in early organogenesis. Concept 33.5 Extraembryonic Membranes Nourish the Growing Embryo The amniote egg, with its contained water supply, frees development from requiring an external water supply. Extraembryonic membranes surround embryos in amniote eggs. They function in nutrition, gas exchange, and waste removal. Concept 33.5 Extraembryonic Membranes Nourish the Growing Embryo In the chick, four membranes form: • Yolk sac—encloses yolk within the egg and passes nutrients to the embryo • Allantoic membrane—a sac for waste storage • Amnion—secretes fluid for protection • Chorion—reduces water loss and exchanges gases Figure 33.15 The Extraembryonic Membranes of Amniotes (Part 1) Figure 33.15 The Extraembryonic Membranes of Amniotes (Part 2) Concept 33.5 Extraembryonic Membranes Nourish the Growing Embryo Human gestation is divided into trimesters of about 12 weeks each. In the first trimester the embryo is very sensitive to damage from radiation, drugs, and chemicals. Gastrulation occurs, tissues differentiate, and the placenta forms. By the end of the first trimester, most organs have started to form and the embryo becomes a fetus. Concept 33.5 Extraembryonic Membranes Nourish the Growing Embryo During the second and third trimesters the fetus grows rapidly. Toward the end of the third trimester the organ systems mature. Birth occurs when the last of its critical organs—the lungs—matures. The end!