Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
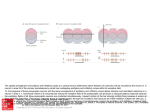
Biol. Cybern. 76, 357–363 (1997) Biological Cybernetics c Springer-Verlag 1997 Coarse coding: calculation of the resolution achieved by a population of large receptive field neurons Christian W. Eurich1 , Helmut Schwegler2 1 2 The University of Chicago, Department of Neurology, MC 2030, 5841 South Maryland Avenue, Chicago, IL 60637, USA Institut für Theoretische Physik, Universität Bremen, D-28334 Bremen, Germany Received: 7 January 1996 / Accepted in revised form: 7 January 1997 Abstract. Electrophysiological studies in various sensory systems of different species show that many neurons involved in object localization have large receptive fields. This seems to contradict the high sensory resolution and the behavioral precision observed in localization experiments. Assuming a coarse coding mechanism, the resolution obtained by an ensemble of neurons is analytically calculated as a function of receptive field size. It is shown that particularly large receptive fields yield a high resolution. 1 Introduction A substantial amount of work in the neurosciences has been dedicated to the question of how information is represented in neural systems. In this paper, we consider tasks of object localization such as visual or auditory localization of an object in the surroundings, or tactile localization of an object on the skin surface. The space in which a localization takes place will be referred to as the sensory space. Theoretical calculations of the sensory resolution for different forms of internal representation of object position lead to results which can be tested quantitatively in behavioral experiments. At the same time, electrophysiological and anatomical data about the neurons involved in the processing of sensory information are available, relating behavioral patterns to the underlying anatomy and physiology. Many species show a high accuracy in object localization. Tongue-projecting salamanders (Bolitoglossini ), for example, are able to localize mites of 0.5 mm size at distances of 15–20 cm and snap at them with high success rates (Roth 1987). Barn owls (Tyto alba) determine the direction of the sound of a prey with an accuracy of a few degrees (Konishi 1983, 1993). In human subjects, the two-point discrimination threshold for tactile stimulation is 1.4 mm, decreasing to 1.16 mm after paired peripheral tactile stimulation (PPTS; Stauffenberg et al. 1994; Dinse et al. 1995). Interestingly, in all of these cases central neurons involved in object localization have receptive fields which are Correspondence to: C. Eurich (Fax: +(312) 702-9076; e-mail: [email protected]) large compared with the sensory resolution observed in behavioral experiments. By a receptive field of a central neuron we mean the subset of the sensory space in which an appropriate stimulus elicits a reaction in the corresponding neuron. Tectal neurons in the tongue-projecting salamander Hydromantes italicus have a mean receptive field diameter of 41◦ , with a minimum of 10◦ and a maximum near 360◦ (Wiggers et al. 1995). The receptive fields of auditory neurons in the nucleus mesencephalicus lateralis dorsalis of Tyto alba range from 23◦ to “unrestricted” in elevation and have a mean diameter of 25◦ in azimuth (Knudsen and Konishi 1978; Konishi 1993). In the case of tactile stimulation, electrophysiological recordings have been obtained in the primary somatosensory cortex of the rat. The neurons react to a skin surface of 45.4(±4.6) mm2 with an increase to 79.9(±8.2) mm2 after PPTS, resulting in a higher receptive field overlap (Dinse et al. 1995). These findings suggest that large receptive field neurons may well be involved in object localization, a task which has often been ascribed exclusively to small receptive field neurons (Grüsser-Cornehls 1984; Gaillard 1985). Several mechanisms have been proposed for the neural coding of the position of a stimulus in a sensory space, X (Milner 1974; Feldman and Ballard 1982; Hinton et al. 1986; Snippe and Koenderink 1992). The main concepts are the local coding scheme, the intensity coding scheme, and the ensemble coding scheme. In the local coding scheme, X is divided into small cells the size of the space which can be resolved by the receptor cells. Each cell corresponds to one neuron which fires whenever a stimulus appears in its cell. Although a discrimination of several stimuli is fairly easy in this coding scheme, the number of neurons necessary to cover X grows exponentially with the dimension of X. This combinatorial explosion makes it unlikely that local coding is exclusively used as a localization mechanism in neural systems (Feldman and Ballard 1982). In the intensity coding scheme, each neuron encodes the position of an object along one dimension of X by firing at different rates. Thus, the number of neurons involved is relatively small, and the resolution is determined by the reliability of the neuron reaction (e.g., by the amount of noise). 358 The main drawback of the intensity coding is the time needed for defining a precise firing rate with a neural spike train; this time is much too long to explain fast animal reactions in many cases, particularly in prey capture activity. Another difficulty lies in the fact that the same firing rate may be elicited in a neuron either by a single object or by a combined stimulation originating from two or more objects; the neural system cannot discriminate between one or more stimuli in sensory space. This property of the convergence of information channels, known as metamery, takes an extreme form in the case of intensity coding and prevents an appropriate representation of the world. Finally, in the ensemble coding scheme, each neuron has a receptive field covering a region which is large compared with the sensory resolution, resulting in a receptive field overlap everywhere on X. The position of a stimulus is encoded by the ensemble of neurons contributing to the respective overlap. The abovementioned large receptive fields found empirically suggest an ensemble coding mechanism in various sensory systems. This is supported by the fact that parallel information processing plays an important role in the brain, from the simultaneous activations of many receptors to the population code for muscle control. Although metamery is also present in the ensemble coding scheme, different stimuli can still be discriminated if they are not too close to each other or if the neuron density is sufficiently high. Several attempts have been made to understand the properties of ensemble coding. Heiligenberg (1987) and Baldi and Heiligenberg (1988) considered the hyperacuity properties of an infinite, one-dimensional array of large, overlapping receptive fields. They assumed a Gaussian sensitivity profile for the neurons, i.e., the firing rate of a neuron depended on the position of the stimulus within the receptive field (channel coding). Baldi and Heiligenberg concluded that the resolution increases with receptive field size and that the system is stable with respect to noise. For the case of channel coding, Snippe and Koenderink (1992) calculated the resolution obtained by receptive fields arranged on an n-dimensional lattice in the presence of noise. Their conclusion was that receptive field size does not play a role for n = 2 but that large receptive fields and, correspondingly, substantial receptive field overlap are only advantageous for n > 2. Obviously, these findings do not account for the large receptive fields described above: Direction localization (either visual or auditory) and somatosensory localization are two-dimensional problems. Instead of channel coding, which like intensity coding suffers from the problem of the definition of an appropriate firing rate within a short amount of time, Hinton (1981) and Hinton et al. (1986) considered the coarse coding mechanism in which the neurons do not have a Gaussian sensitivity profile, but are binary in nature: They fire whenever a stimulus is within their receptive field, and are otherwise “silent”. In the coarse coding scheme, the resolution is determined by the number of different encodings (i.e., by the number of different firing patterns) in the neural population as a stimulus is moved about in the sensory space. This directly leads to the result that large receptive fields yield a high resolution, because they overlap extensively and therefore show a high number of different encodings. In a rough estima- tion, Hinton et al. (1986) showed that for X = IRk with equally distributed receptive fields all having the shape of k-dimensional spheres of radius r, the accuracy (which is the reciprocal of the resolution) is proportional to N rk−1 , i.e., for k ≥ 2, large receptive fields are advantageous compared with small receptive fields. In a biologically relevant situation, however, the space X = IRk has to be replaced by a space of finite size. In this case, r cannot be arbitrarily large. Instead, we find that the resolution is bad if r approaches the size of X, since receptive fields covering the whole sensory space (and hence, neurons firing all the time) do not convey information. On the other hand, Hinton’s formula shows that for r → 0, the resolution is also bad. Therefore, an optimal value for the receptive field size exists, the calculation of which requires a more general formalism for the evaluation of the resolution obtained by a population of neurons. Concerning the accuracy in the coarse coding scheme, Hinton et al. (1986, p. 91) wrote: “The accuracy is proportional to the number of different encodings that are generated as we move a feature point along a straight line from one side of the space to the other. Every time the line crosses the boundary of a zone, the encoding of the feature point changes because the activity of the unit corresponding to that zone changes.” The mechanism developed here suggests that it might be possible to calculate the resolution obtained by a population of neurons by mapping combinatorics of receptive fields onto the density of receptive field boundaries. Following this concept, we assume that (i) receptive field boundaries are simple, one-dimensional lines and (ii) receptive fields are “compact” in shape, e.g., circular, elliptical, or rectangular. The first assumption rules out pathological cases such as fractal receptive boundaries which – despite their infinite length – do not lead to an increase in resolution, because they do not increase the number of different encodings in the neural population. The second assumption rules out receptive fields which are widely ramified, or consist of several disconnected parts. This feature corresponds to the existence of a topological map in the sense that nearby positions in the sensory space have similar representations in the neural population, whereas stimuli in positions which are far apart activate very different ensembles of neurons. Although in the presence of such a topological map the number of different encodings in a population of N neurons is smaller than the combinatorial maximum of 2N , the evaluation of the information stored in the population – in the form of motor commands or the activation of further neural structures – is likely to be more efficient, especially in the presence of noise. Receptive fields with simple shapes are usually encountered in nature, e.g., in the visual (Gaillard 1985; Wiggers et al. 1995) or in the auditory (Knudsen and Konishi 1978) system. In the remainder of this article, the resolution obtained by a population of receptive fields is calculated analytically by considering the density of receptive field boundaries. Thereby, we restrict ourselves to circular fields in twodimensional sensory spaces. In Sect. 2, the basic concepts such as receptive field density and resolution are introduced. Section 3 gives a simple example revealing the basic mechanism responsible for a high resolution. In Sect. 4, a local formalism is developed which allows the derivation of the 359 i.e., to a different representation of the position of the object. In this way, the resolution obtained by the neuron population is related to the density of receptive field boundaries. In order to obtain an analytical expression for the resolution, we idealize the situation to a continuous field approximation with a receptive field density σ(x) with the properties g 3 f e 1 d b γ Z σ(x) ≥ 0 σ(x) dx = N c 2 a Fig. 1. A sensory space, X = IR2 , is covered with three circular receptive fields. A stimulus moving along a path, γ, elicits different reactions in the neuron population depending on the actual set of overlapping receptive fields resolution for arbitrary receptive field distributions in space and arbitrary receptive field size distributions. We close the considerations with a short discussion in Sect. 5. A forthcoming article (Eurich et al. 1997b) is concerned with the applications to special biological systems, namely the visually controlled orienting movement of salamanders towards prey positions and the tongue-projection of salamanders which requires an accurate determination of prey distance. 2 Basic notions Let X be the sensory space, i.e., the space in which the localization of an object takes place. Examples include a three-dimensional subset of Euclidean space, IR3 , for the localization of a visual or auditory object including distance, or the two-sphere, S 2 , for the determination of direction only. In the latter case, it is assumed that the two-sphere is embedded in IR3 ; the observer is positioned in the center of the sphere, so that each point x ∈ S 2 corresponds to a certain direction. Consider N receptive fields Ri ⊆ X at positions xi (i = 1, . . . , N ) where xi refers to a characteristic point of Ri (e.g., the center of a circular receptive field). The output function of a neuron is defined as 1, if x ∈ Ri (i = 1, . . . , N ) (1) fi (x) = 0, if x ∈ / Ri i.e., we consider binary neurons with the additional property of a receptive field. Note that this restricted view of neuron behavior only refers to the position of an object; a firing rate coding can be supplemented corresponding to additional features of the environment. As an illustration, Fig. 1 shows a sensory space X = IR2 with three circular receptive fields. A single stimulus moves along a path, γ, which can be divided into 7 parts, (a)– (g), according to the different firing patterns elicited in the neuron population. For example, when the stimulus is in part (b) neuron 1 fires, and when the stimulus is in part (c) neurons 1 and 2 fire. Whenever the stimulus crosses a receptive field boundary, one of the neurons is switched on or off, thus leading to a different active neuron ensemble, (2) (3) X and, for arbitrary A ⊆ X, Z σ(x) dx ≈ # (receptive fields in A) (4) A where the approximation results from the fact that the number of real receptive fields is integer while the integral on the left side takes continuous values. σ(x) can either be calculated from a probability distribution of receptive fields or from a set of experimental data; see Appendix A. In optics, resolution is defined as the minimal distance of two points which can still be separately mapped by an optical device. In our case, two points in X are separately mapped (i.e., have different representations) if there is a receptive field boundary running between them. This leads to the definition of the resolution A(x) A(x) = 1 L(x) (5) where L(x) is the density of receptive field boundaries. The main objective in the remainder of this article is the inference of a relation between the density σ(x) – which is usually determined by experimental investigations – and the density L(x). Here we confine ourselves to the geometric situation of a two-sphere with radius R, 2 = (x, y, z) ∈ IR3 : x2 + y 2 + z 2 = R2 (6) X = SR covered with circular receptive fields, i.e., with receptive fields having the shapes of spherical caps. Extensions to X = IR2 and X = S 1 (Eurich 1995) or to the cylinder (Eurich et al. 1997a) are straightforward. The consideration of arbitrary manifolds with a more general class of receptive field morphologies requires a sophisticated mathematical approach and is not required for most biological applications anyway. An immediate consequence of (5) is 1 (7) N since σ(x) ∝ N and therefore L(x) ∝ N . Equation (7) is advantageous compared with the situation of local coding with nonoverlapping receptive √ fields where the resolution is only proportional to 1/ dim X N . The result is in agreement with the abovementioned estimation in Hinton et al. (1986). A(x) ∝ 3 An introductory example As an introductory example revealing the basic mechanism responsible for the high resolution of large receptive fields, 360 γ 2ρ R 1 a b 2 . a Cross-section of S 2 showing a Fig. 2. The geometry for X = SR R receptive field with angular diameter 2%. b The sphere is divided into two sets, S1 (dark gray), a stripe of angular width 2% surrounding a great circle, γ, and the remaining spherical caps, S2 (light gray) 2 consider the case X = SR with uniformly distributed receptive fields of equal size. The normalization (3) yields N ≡ σ ∀(x∈X) (8) 4πR2 2 Figure 2a shows the cross-section of SR with a recep2 tive field which has an angular diameter 2%. In Fig. 2b, SR is depicted. The path, γ, is a great circle. The resolution is determined by the number of receptive field boundaries intersecting γ. According to the angular diameter of the re2 can be divided into two regions, S1 and ceptive fields, SR S2 , where S1 is a stripe of angular width 2% around γ, and S2 is composed of the two remaining spherical caps. Thus, receptive fields whose centers are situated within S1 intersect γ twice, whereas receptive fields in S2 do not intersect γ at all. (Receptive fields sitting exactly on the boundary between S1 and S2 are tangent to γ in one point, but they form a set of measure zero.) The number of receptive field boundaries intersecting γ, E(%), is expected to be Fig. 3. The angular resolution per neuron, N α(%), as a function of receptive field size e σ(x) = E(%) = σ (2 · F1 + 0 · F2 ) = 2N sin % (9) F1 (resp. F2 ) being the surface area of S1 (resp. S2 ). Division by the length of γ yields the density of receptive field boundaries along γ: E(%) N sin % = 2πR πR This results in a resolution 1 πR A(%) = = L(%) N sin % L(%) = (10) (11) x0 γ κ(x0,ρ) Fig. 4. The geometrical situation for the calculation of the density of re2 for the direction e which is tangential ceptive field boundaries at x0 ∈ SR to a curve γ. κ(x0 ; %) indicates the set of all positions of receptive fields which contribute with a field boundary at x0 . One of these receptive fields is shown (gray) yield the best resolution is that they have the longest possible field boundaries: Given a fixed number of neurons, they yield the highest possibility of being crossed by a stimulus moving in X. 4 Extension to more general cases In this section, a twofold extension of the previous example is developed. First, a local formalism is introduced which allows the calculation of the resolution for arbitrary receptive field densities σ(x) on the sphere. Second, instead of a fixed receptive field size for all neurons, a size distribution is considered. The distribution may vary from place to place as observed, for example, in the visual system of salamanders, where large receptive fields tend to be situated mainly in the lateral visual field. The angular resolution is α(%) = A(%) π = R N sin % (12) In Fig. 3, the function N α(%) – which can be interpreted as the angular resolution per neuron – is plotted against %. It has a minimum at % = 90◦ , which means that the best resolution is achieved by a population of neurons with a receptive field diameter of 180◦ . These are large field neurons whose receptive fields cover half of the sphere. The corresponding resolution is α(90◦ ) = 180◦ /N , i.e., with, say, N = 100 neurons a resolution of 1.8◦ is achieved. An illustrative explanation of the fact that the largest possible receptive fields 4.1 Arbitrary field densities Assume that all receptive fields possess the same angular 2 as in the diameter, 2%. Instead of a path γ across X = SR previous example, consider a local direction, e, which can be 2 . For a thought of being tangential to γ at some point x0 ∈ SR given field density σ(x), we calculate the density of receptive field boundaries at x0 , Le (x0 ; %). In general, Le (x0 ; %) varies with e, which is indicated by the subscript. The geometric situation is reproduced in Fig. 4. All receptive fields contributing with a field boundary at x0 are 361 situated on a circle, κ(x0 ; %), of angular diameter 2% around x0 . For each point x ∈ κ(x0 ; %) a weighting factor σ(x) has to be supplied. Furthermore, σ(x) has to be multiplied by a geometrical factor R sin %| cos β|, where β is an angle parametrizing κ(x0 ; %) with β = 0 corresponding to the direction e (see Fig. 5a). The geometrical factor takes into account that receptive fields lying in the directions parallel to e have a greater influence than receptive fields lying in the directions perpendicular to e. The density of receptive field boundaries finally results from an integration over β: Z2π Le (x0 ; %) = R sin % σ(ϑ(β), ϕ(β))| cos β| dβ (13) a different value of %. Ultimately, in the expression for the density of receptive field boundaries (13), σ(x) has to be replaced by σ̃(x, %), and an additional integration over the range of values of % has to be performed. Considering (18), the result is (19) Le (x0 ; w) 2π π ZZ R sin % σ(ϑ(β), ϕ(β))w(%|(ϑ(β), ϕ(β)))| cos β| dβd% = 0 0 The parameter w in Le (x0 ; w) indicates that L does not refer to a single receptive field size but to a size distribution. 0 2 where ϑ, ϕ are spherical coordinates on SR . For a more detailed derivation of (13), see Appendix B. According to (5), the resolution is the reciprocal value of the density of receptive field boundaries, Ae (x0 ; %) = 1/Le (x0 ; %), while the angular resolution results from a division of Ae (x0 ; %) by R as in (12), αe (x0 ; %) = Ae (x0 ; %)/R. As an example, let σ(x) again take the constant value 2 and e, it follows from (13) (8). Then, for arbitrary x0 ∈ SR sin % L(%) = N 4πR Z2π | cos β| dβ 0 sin % (14) =N πR which is in agreement with the result (10) obtained above. 4.2 Receptive field size distribution Receptive fields of sensory neurons usually have different sizes. For a mathematical treatment, consider the joint den2 , % ∈ R ≡ [0; π]) where w(x, %)dxd% sity w(x, %) (x ∈ SR is the probability of finding a receptive field in the range [%; % + d%] in the volume dx at x. The joint distribution is normalized as follows: Z Z w(x, %) dxd% = 1 (15) X R In practice, instead of w(x, %), a size distribution is given 2 . This is mathematically described as the for various x ∈ SR conditional probability density w(%|x) = w(x, %) w(x) (16) where w(x) is the marginal receptive field distribution (23) introduced in Appendix A. Analogous to the case (28) of constant field sizes, a receptive field density σ̃(x, %) can be introduced which comprises the dependencies of both position and size: σ̃(x, %) = N w(x, %) (17) (16), (17), and (28) yield σ̃(x, %) = w(%|x) σ(x) (18) In the local formalism, instead of a single curve κ(x0 , %) a set of such curves has to be taken into account, each for 5 Discussion A coarse coding mechanism was assumed for the encoding of a single position in a sensory space, X, by a population 2 of neurons. As a standard example for X, the two-sphere SR was chosen, describing for example the visual or auditory direction localization. For the case of “compact” receptive fields, the notion of accuracy developed in Hinton et al. (1986) was replaced by a definition of resolution based on the density of receptive field boundaries. The resolution was calculated analytically for circular receptive fields with arbitrary distributions in X and arbitrary size distributions. Even the simplest case of equally distributed receptive fields all having the same size shows that the best resolution is obtained with receptive fields which are 180◦ in diameter. In this case, receptive field overlap is maximal, leading to the highest possible number of overlap patterns, and hence to the highest number of different neural representations of positions in the sensory space. The results suggest that the large receptive field neurons found in the visual, auditory, and somatosensory system of various species contribute to the high accuracy observed in behavioral experiments. According to a previously adopted view, only small receptive field neurons are involved in object localization, whereas large receptive field neurons are responsible for large-scale movements, predator detection, etc. (Grüsser-Cornehls 1984; Gaillard 1985). However, only the detailed analysis of a given sensory system can answer the question of the relevance of neurons with different receptive field sizes. Examples for such analyses will be supplied in forthcoming articles (Eurich et al. 1997a,b). In the general case of arbitrarily distributed receptive fields, the density of receptive field boundaries (13,19) depends on the direction e in the sensory space, i.e., the resolution is angle-dependent. This anisotropy is consistent with psychophysical studies (Jastrow 1893; for a review, see Appelle 1972) showing that the performance of the visual system is poorer for diagonal stimuli than for horizontal and vertical stimuli (oblique effect). A similar effect has been demonstrated in the somatosensory system (Lechelt 1988). Further analysis comparing theoretical prediction and empirical data may show that at least part of the phenomena can be explained in terms of distributions of receptive fields. Some further topics in connection with coarse coding remain to be worked out. First, the mechanism of decoding the information carried by a population of neurons has not been 362 Appendix A Calculation of the receptive field density eϑ e W (x1 , . . . , xN ) dx1 . . . dxN = 1 (20) XN where X N is the N -fold Cartesian product of X with itself. W is symmetric with respect to permutations of its arguments since the sample of receptive fields is not ordered: x0 { R sin ρ β κ (x 0 ;ρ) ∼ β0 eϕ R cos ρ 0 κ (x 0 ;ρ) a { R sin ρ ρ x b dh κ (x 0+dl e; ρ) dF dβ e The receptive field density, σ(x), can either be obtained from a probability distribution of receptive fields on X, or from a limited set of (usually experimental) data. In the former case, consider the function W (x1 , . . . , xN ) giving the probability density of receptive field Ri being at xi (i = 1, . . . , N ) with respect to an ensemble of individuals of the same species. The normalization yields Z 0 x0+dl e x0 dl β β { tackled analytically. However, numerical calculations in the form of a neural network have been performed, indicating that the neuron population which evaluates the distributed information (e.g., in the form of muscle contractions) does not considerably outnumber the encoding neuron population (Eurich et al. 1995). Second, only a single stimulus has been considered so far. It is an open question as to what preprocessing is necessary to analyse a complex sensory scene, and eventually discriminate between several objects, with a coarse coding mechanism. R sin ρ e κ(x0; ρ) κ(x0; ρ) c d Fig. 5. a Topview and b sideview of the curve κ(x0 ; %). c The area dF (gray) traversed by the curve κ when the stimulus moves from x0 to x0 +dle. d The local geometry concerning a single stripe (gray) of dF . For further explanations, see text W (x1 , . . . , xi , . . . , xj , . . . , xN ) = W (x1 , . . . , xj , . . . , xi , . . . , xN ) ∀i,j∈{1,...,N } (21) The marginal distribution with respect to the ith receptive field is Z W (x1 , . . . , xN ) dx1 . . . dxi−1 dxi+1 . . . dxN wi (xi ) = (22) X N −1 (i = 1, . . . , N ) and (21) immediately yields w1 (x1 ) = · · · = wN (xN ) =: w(x) (23) w(x)dx is the probability of finding an arbitrary receptive field in the volume dx at x. Now define a function nA (x1 , . . . , xN ) giving the number of receptive fields in A ⊆ X: nA (x1 , . . . , xN ) = # (receptive fields in A ⊆ X) (24) Just like W (x1 , . . . , xN ), nA (x1 , . . . , xN ) is symmetric with respect to permutations of its arguments. Let IA (x) be the indicator function for the subset A ⊆ X, n IA (x) = 1, x ∈ A 0, x ∈ /A (25) Then, A comparison of (27) and (4) leads to the definition of the density of receptive fields, σ(x), σ(x) := N w(x) (28) σ(x) fulfils the requirements (2) and (3). In a slightly different interpretation, σ(x) in (28) can be regarded as the receptive field density of a “mean system”. Under the assumption that the members of the statistical ensemble do not differ significantly, σ(x) can be obtained by looking at an actual, single system which is given in an experimental situation. The question arising in this situation is: Given a finite number of receptive field positions, x0i (i = 1, . . . , N ), what does the field density σ(x) look like? Since a probability arises from a relative frequency in the limit of an infinite number of trials only, σ(x) cannot be inferred uniquely. For 2 , the following procedure yields X being the two-sphere with radius R, SR good results: For all i, replace the receptive field at x0i by a Gaussian function with variance π 2 /4 centered at x0i , w̃i (x; x0i ) = exp (− 2ε2 (x, x0i ) 2 ) (x ∈ SR ) π2 where ε(x, x0i ) is the angle between x and x0i : ε(x, x0i ) = arccos(x · x0i ) nA (x1 , . . . , xN ) = N X IA (xi ) (26) i=1 Applying (22), (23), and (26), the expected value of the number of receptive fields in A is given by Z hnAi = N Z X = i=1 Z =N N P σ(x) = N A i=1 N RP X i=1 exp (− 2ε 2 (x,x0i ) ) π2 (31) 2 0i ) exp (− 2ε (x,x ) π2 dx A change of summation and integration in the denominator of (31) yields N integrals all having the same value I which is obtained numerically as I ≈ 7.579. The resulting formula for the density σ(x) is IA (x) w(x) dx N P X w(x) dx (30) The receptive field density is obtained by summation of the Gaussian functions, normalization, and multiplication with N : nA (x1 , . . . , xN )W (x1 , . . . , xN ) dx1 . . . dxN XN (29) (27) σ(x) = exp (− 2ε i=1 I 2 (x,x0i ) ) π2 (32) 363 Appendix B Derivation of Equation (13) According to Fig. 4, all points x ∈ κ(x0 ; %) contribute with a receptive field boundary at x0 . κ(x0 ; %) is a circle which forms the basis of a spherical shell centered around x0 . A topview and a sideview of κ(x0 ; %) are shown in Fig. 5a and 5b, respectively. The circle is parametrized as follows. Each point x ∈ κ(x0 ; %) can be written as x = x0 cos % + R sin % cos(β + β0 ) eϕ0 + R sin % sin(β + β0 ) eϑ0 (33) where eϕ0 and eϑ0 are the basis vectors corresponding to spherical coordinates at x0 , Ã eϑ0 = cos ϑ0 cos ϕ0 cos ϑ0 sin ϕ0 − sin ϑ0 ! Ã , eϕ0 = − sin ϕ0 cos ϕ0 0 ! (34) and β0 is the angle between e and eϕ0 , β0 = arccos(e · eϕ0 ) (35) The circle is thus parametrized by the angle β ∈ [0; 2π] with β = 0 corresponding to the direction e (Fig. 5a). Given a fixed set (x0 , e, %), (33) yields x = (x, y, z)T as a function of β which is necessary to evaluate σ(x(β)). If the receptive field density is given as a function of spherical coordinates instead, (33) has to be combined with the well-known transformation of Cartesian and spherical coordinates to obtain σ(ϑ(β), ϕ(β)). A stimulus moving from x0 to x0 + dle leads to the curve κ traversing the area dF as indicated in Fig. 5c. For the purpose of integration, dF is divided into small stripes parallel to e (Fig. 5d). The stripes have a length dl and a width dh which depends on the angle β: dh = R sin %| cos β|dβ (36) An integration of the receptive field density, σ(x(β)) or σ(ϑ(β), ϕ(β)), over dF = dh dl yields the number of receptive field boundaries the stimulus intersects on its way, while an integration over dh alone as given by (36) results in an expression for the density of receptive field boundaries at x0 corresponding to the direction e. The latter immediately leads to (13). Acknowledgements. We thank Gerhard Roth and Wolfgang Wiggers for helpful discussions and the supply of salamander data. Manfred Fahle’s hint on the oblique effect and the comments of an anonymous referee are gratefully acknowledged. This work was supported by the Deutsche Forschungsgemeinschaft with a grant from the Schwerpunktprogramm “Physiologie und Theorie neuronaler Netzwerke”. One of us (C.W.E.) was supported by the Studienstiftung des deutschen Volkes. References 1. Appelle S (1972) Perception and discrimination as a function of stimulus orientation: the “oblique effect” in man and animals. Psychol Bull 78: 266–278 2. Baldi P, Heiligenberg W (1988) How sensory maps could enhance resolution through ordered arrangements of broadly tuned receivers. Biol Cybern 59: 313–318 3. Dinse HR, Godde B, Spengler F (1995) Short-term plasticity of topographic organizaton of somatosensory cortex and improvement of spatial discrimination performance induced by an associative pairing of tactile stimulation. (Internal Report 95-01) Institut für Neuroinformatik, Bochum 4. Eurich CW (1995) Objektlokalisation mit neuronalen Netzen. Harri Deutsch, Frankfurt 5. Eurich CW, Roth G, Schwegler H, Wiggers W (1995) Simulander: a neural network model for the orientation movement of salamanders. J Comp Physiol A 176: 379–389 6. Eurich CW, Dinse HR, Dicke U, Godde B, Schwegler H (1997a) Coarse coding accounts for improvement of spatial discrimination after plastic reorganization in rats and humans. (Submitted) 7. Eurich CW, Schwegler H, Woesler R (1997b) Coarse coding: applications to the visual system of salamanders. Biol Cybern (in press) 8. Feldman JA, Ballard DH (1982) Connectionist models and their properties. Cogn Sci 6: 205–254 9. Gaillard F (1985) Binocularly driven neurons in the rostral part of the frog optic tectum. J Comp Physiol A 157: 47–55 10. Grüsser-Cornehls U (1984) The neurophysiology of the amphibian optic tectum. In: Vanegas H (ed) Comparative neurology of the optic tectum. Plenum Press, New York, pp211–245 11. Heiligenberg W (1987) Central processing of sensory information in electric fish. J Comp Physiol A 161: 621–631 12. Hinton GE (1981) Shape representation in parallel systems. In: Proceedings of the 7th International Joint Conference on Artificial Intelligence, Vancouver, BC, Canada, pp1088–1096 13. Hinton GE, McClelland JL, Rumelhart DE (1986) Distributed representations. In: Rumelhart DE, McClelland JL (eds) Parallel distributed processing, Vol 1. MIT Press, Cambridge, Mass., pp77–109 14. Jastrow J (1893) On the judgment of angles and positions of lines. Am J Psychol 5: 214–248 15. Knudsen EI, Konishi M (1978) A neural map of auditory space in the owl. Science 200: 795–797 16. Konishi M (1983) Localization of acoustic signals in the owl. In: Ewert J-P, Capranica RR, Ingle DJ (eds) Advances in vertebrate neuroethology. Plenum Press, New York, pp227–245 17. Konishi M (1993) Listening with two ears. Sci Am 268: 34–41 18. Lechelt EC (1988) Spatial asymmetries in tactile discrimination of line orientation: a comparison of the sighted, visually impaired, and blind. Perception 17: 579–588 19. Milner PM (1974) A model for visual shape recognition. Psychol Rev 81: 521–535 20. Roth G (1987) Visual behavior in salamanders. Springer, Berlin Heidelberg New York 21. Snippe HP, Koenderink JJ (1992) Discrimination thresholds for channel-coded systems. Biol Cybern 66: 543–551 22. Stauffenberg B, Godde B, Spengler F, Dinse HR (1994) Time course and persistence of changes of human spatial discrimination performance induced by a paired tactile stimulation protocol. In: Elsner N, Breer H (eds) Sensory transduction. Thieme, Stuttgart, p262 23. Wiggers W, Roth G, Eurich CW, Straub A (1995) Binocular depth perception mechanisms in tongue-projecting salamanders. J Comp Physiol A 176: 365–377