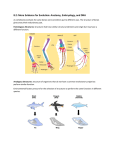
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 Phil. Trans. R. Soc. B (2009) 364, 2925–2940 doi:10.1098/rstb.2009.0099 Review The evolution of early vertebrate photoreceptors Shaun P. Collin1,*, Wayne L. Davies2, Nathan S. Hart1 and David M. Hunt3 1 School of Biomedical Sciences, The University of Queensland, Brisbane 4072, Queensland, Australia 2 Nuffield Laboratory of Ophthalmology, University of Oxford, The John Radcliffe Hospital, Headley Way, Oxford OX3 9DU, UK 3 UCL Institute of Ophthalmology, 11 – 43 Bath Street, London EC1V 9EL, UK Meeting the challenge of sampling an ancient aquatic landscape by the early vertebrates was crucial to their survival and would establish a retinal bauplan to be used by all subsequent vertebrate descendents. Image-forming eyes were under tremendous selection pressure and the ability to identify suitable prey and detect potential predators was thought to be one of the major drivers of speciation in the Early Cambrian. Based on the fossil record, we know that hagfishes, lampreys, holocephalans, elasmobranchs and lungfishes occupy critical stages in vertebrate evolution, having remained relatively unchanged over hundreds of millions of years. Now using extant representatives of these ‘living fossils’, we are able to piece together the evolution of vertebrate photoreception. While photoreception in hagfishes appears to be based on light detection and controlling circadian rhythms, rather than image formation, the photoreceptors of lampreys fall into five distinct classes and represent a critical stage in the dichotomy of rods and cones. At least four types of retinal cones sample the visual environment in lampreys mediating photopic (and potentially colour) vision, a sampling strategy retained by lungfishes, some modern teleosts, reptiles and birds. Trichromacy is retained in cartilaginous fishes (at least in batoids and holocephalans), where it is predicted that true scotopic (dim light) vision evolved in the common ancestor of all living gnathostomes. The capacity to discriminate colour and balance the tradeoff between resolution and sensitivity in the early vertebrates was an important driver of eye evolution, where many of the ocular features evolved were retained as vertebrates progressed on to land. Keywords: photoreception; cones; rods; opsin genes; visual pigments; spectral sensitivity 1. INTRODUCTION (a) The earliest vertebrates Lampreys and hagfishes are the sole survivors of the very early agnathan (jawless) stage in vertebrate evolution (Hardisty 1982). Ancient hagfishes, based on a single fossil specimen, Myxinikela siroka, found in similar deposits to fossil lampreys, are thought to date back to the Cambrian, with M. siroka surviving until the end of the Triassic era (Bardack 1991, 1998). Extant hagfishes have changed little, and are considered monophyletic and comprise 60 species within five genera: Paramyxine, Eptatretus (35 species), Myxine (19 species), Notomyxine (single species), Neomyxine (single species) and Nemamyxine (two species) (Fernholm 1998). These bottom-dwelling, cartilaginous fishes are found in deep, cold water on all continents (except Antarctica) and feed on dead or moribund fish and invertebrates with two sets of laterally * Author for correspondence ([email protected]). One contribution of 13 to a Theme Issue ‘The evolution of phototransduction and eyes’. everting and biting or scraping teeth or cusps (Fernholm 1998). Hagfishes possess a pair of lateral eyes embedded beneath an opaque patch of cranial epithelium. Although the eyes of hagfishes have traditionally been thought to be degenerate (Holmberg 1970, 1971), lacking a lens and both intra- and extraocular eye muscles, recent studies consider that they may represent a ‘missing link’ in eye evolution, lying intermediate between the non-image-forming eyes of tunicates and the image-forming eyes of lampreys (Lamb et al. 2007, 2008). Recent studies have shown that lampreys, or their very close relatives, had already evolved by the Lower Cambrian period (ca 540 Ma; Xian-Guang et al. 2002; Shu et al. 2003). While all 34 species of Northern Hemisphere (or holarctic) lampreys (Renaud 1997) are placed in a single family (the Petromyzontidae), the four species of Southern Hemisphere lampreys are separated into either the Geotriidae or Mordaciidae (Potter 1980). It has been proposed that both latter families were derived independently from stocks similar to those of contemporary representatives of the holarctic genus Ichthyomyzon (Hubbs & Potter 1971; Potter & Hilliard 1987; Gill et al. 2925 This journal is # 2009 The Royal Society Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 2926 S. P. Collin et al. Review. Vertebrate photoreceptor evolution 2003). The Geotriidae is represented solely by Geotria australis, an anadromous species, which is found in rivers in New Zealand, southern Australia, Tasmania, Chile and Argentina (Potter & Strahan 1968), while the Mordaciidae contains two anadromous species (Mordacia mordax and Mordacia lapicida), restricted to rivers in eastern Australia and Chile, respectively (Potter & Strahan 1968; Potter et al. 1978), and a non-parasitic species, Mordacia praecox. The eyes of larval lampreys or ammocoetes are similar to those of hagfishes, i.e. are buried beneath the skin, where most of the retina is relatively undifferentiated and possesses a thick neuroblastic layer (Dickson & Collard 1979; Rubinson & Cain 1989; Rubinson 1990). Recent reviews by Lamb et al. (2007) and Lamb (2009) suggest that lampreys inherited their eyes from an ancestor that they had in common with hagfish, and that this hagfish-like larval eye is present in the lamprey larva but transforms to a vertebratelike eye in the adult. The eyes of fully metamorphosed lampreys are certainly prominent and well differentiated, following closely the design of the gnathostomatous fishes, with a multi-focal lens, intra- and extraocular eye muscles and a complex retina (Collin et al. 1999; Gustafsson et al. 2008). The class Chondrichthyes is a monophyletic group with over 1100 extant species occupying a diverse range of habitats, from shallow, freshwater rivers to marine environments such as coral reefs and the open ocean including the deep sea. Extant Chondrichthyes are divisible into two sister taxa, Holocephali and Elasmobranchii, with a basal vertebrate lineage dating back to the Lower Devonian, at least 400 Ma (Grogan & Lund 2004). Close relatives of extant species today are found in the fossil record, dating back 150 Myr (Grogan & Lund 2004; Compagno et al. 2005). The eyes of both holocephalans and elasmobranchs are large, image-forming, camera-type eyes with an almost spherical lens, well-developed intra- and extraocular eyes muscles and a retina packed with photoreceptive elements (Gruber & Cohen 1978; Hodgson & Mathewson 1978; Hueter et al. 2004; Hart et al. 2006a,b). Sarcopterygian fish gave rise to the first tetrapods and are represented today by the lungfishes (the Australian, Neoceratodus forsteri; the African, Protopterus spp.; and the South American, Lepidosiren paradoxa), and the coelacanths, Latimeria chalumnae and Latimeria menadoensis. The relationship between all early Sarcopterygii remains controversial and highly debated despite the advent of phylogenetic analysis of nucleotide and amino acid sequences for a variety of diverse genes (Carroll 1997; Brinkmann et al. 2004; Takezaki et al. 2004). Some molecular analyses of sarcopterygian phylogeny reveal that the lungfishes are more related to tetrapods than to the coelacanth, L. chalumnae (Brinkmann et al. 2004), while others present an unresolved trichotomy between all three groups (Takezaki et al. 2004). Fossil forms of the Australian lungfish Ceratodus date back to the Lower Cretaceous Period 135 Ma, making this species more primitive than the lepidosireniform family of lungfishes and the only remaining member of one of the oldest extant vertebrate genera (Kemp & Molnar 1981). Therefore, Phil. Trans. R. Soc. B (2009) the well-developed visual system of N. forsteri may best reflect the visual system just prior to the emergence of land vertebrates in the Devonian period. These groups of early vertebrates represent critical periods in vertebrate evolution and studies of their visual systems in extant species provide a window into the past. Hagfishes and lampreys are the living representatives of the agnathan lineage that gave rise to gnathostomes, and therefore, studies of their eyes enable us to contemplate the type of light environment perceived by our vertebrate ancestors and how their eyes were adapted for vision, thereby laying the foundation for the emergence of jawed vertebrates. The cartilaginous holocephalans and elasmobranchs represent the basal lineage of jawed vertebrates and are apex predators, relying on vision in addition to a battery of other senses. The colonization of more dim light environments may have been one of the main reasons for the success of this large group, although more members of the Chondrichthyes need to be examined and many studies reveal a diversity of visual adaptations to their light environment, without the constraints of phylogeny (Hart et al. 2006a,b). Extant lungfish, on the other hand, represent a period later in evolution, when vertebrates were about to venture on to land and thereby be exposed to higher light intensities and a wider range of available wavelengths of light. For all of these critical groups, colour vision may have been an important factor in their evolutionary success in sampling the ancient visual landscape. (b) The ancient visual landscape Although the environmental conditions under which many of the ancestral vertebrates existed are largely hypothetical, the visual stimuli that would have driven the random selection of ocular adaptations can be surmised based on the eyes of extinct and extant vertebrates. Paleontological evidence from the Silurian and Devonian periods shows that the lateral eyes of the ancestral vertebrates were capable of image formation and were rotated within their orbits by seven extraocular eye muscles (Young 2008). These heavily armoured ostracoderms were benthic, presumably feeding by extracting food in or on the substrate. Depending on the light environment encountered by these early vertebrate ancestors, two theories have been proposed as to how multiple photoreceptor types evolved: (i) those fish that possessed two visual pigments, one matching the background and the other offset from the background illumination, could better detect targets that were spectrally different by enhancement of contrast (Lythgoe 1980; Hawryshyn 1998) and (ii) the evolution of at least two spectral classes of photoreceptors would eliminate the considerable noise associated with the significant flicker produced by light passing through the surface ripples in shallow water, thereby enabling the earlier detection of predators (Maximov 2000). Both of these theories assume that vision in the ancestral vertebrates was cone based (photopic), with colour vision evolving possibly later as a by-product, following the development of colour adaptation mechanisms via horizontal cells. Despite the absence of any Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 Review. Vertebrate photoreceptor evolution behavioural evidence of colour vision in either the ancestral or early vertebrates, it is highly likely that colour vision dates back at least 540 Myr with the ancestral vertebrates possessing four major types of vertebrate cone opsins, each presumably sensitive to different parts of the light spectrum (see below). Based on molecular genetic analysis of a range of vertebrate visual pigment genes, it was predicted that the long wavelength-sensitive/medium wavelengthsensitive (LWS/MWS) opsins evolved first and that the Rh1 opsins evolved much later, implying that photopic vision preceded scotopic vision (Yokoyama & Yokoyama 1990, 1996). Although the selection pressures underlying the origins of colour vision are still unknown, the early vertebrates lived in a shallow water environment, which was well lit, based on the finding of fossilized green algal deposits (Walker & Laporte 1970), where a broad light spectrum could be exploited by the evolution of multiple visual pigments and utilized in observing a range of complex environments and animals, some of which were inherently colourful (Parker 1998; Maximov 2000). In this review, we present a reflection on the evolution of photoreceptors, i.e. rods and cones (or their predecessors) in early extant vertebrates, based upon morphological, spectral and molecular criteria. This includes early strategies for spectral filtering and tuning and the potential for colour discrimination. The capacity for photopic (cone-based, bright light vision) and scotopic (rod-based, dim light vision) vision will also be discussed within the context of the ancestral light environment, the visual demands placed on each group to feed, avoid predation and reproduce and the need for functional studies on the photoreceptors (regardless of placing them into current visual nomenclature) in order to better understand the evolution of the visual process. 2. EVOLUTION OF EARLY PHOTORECEPTORS (a) Rods or cones? Historically, it has been established that the vertebrate retina, including that of humans, contains two independent visual systems. Schultze (1866, 1867) proposed, on the basis of finding predominantly rods in nocturnal or crepuscular animals and cones in diurnal animals, that two morphologically distinct photoreceptor types mediated scotopic and photopic vision, respectively. Schultze (1867) also recognized the role of cones in the detection of colour, and by comparison of the presence or absence of either rods or cones in a number of different species concluded that cones evolved from rods. These findings were later refuted by several authors who identified conelike receptors in the eyes of cyclostomes, i.e. lampreys (Öhman 1971; Dickson & Graves 1979; Govardovskii & Lychakov 1984; Tonosaki et al. 1989; Collin et al. 1999; Collin & Trezise 2004), but the characterization and distinction of rods and cones in the early vertebrates is not at all clear and has been the subject of great debate for over 150 years (Crescitelli 1972). The retinae of a number of Northern Hemisphere (or holarctic) lampreys have been examined morphologically and thought to contain two photoreceptor Phil. Trans. R. Soc. B (2009) S. P. Collin et al. 2927 types, a short and a long; putatively a rod and a cone photoreceptor, respectively (Crescitelli 1956, 1972). Later, Öhman (1976) classified the two receptors as ‘rods’, and despite both receptors possessing almost spherical receptor bases with only a few synaptic ribbons, the presence of membranous inclusions within the retinal pigment epithelium (RPE) and a plasma membrane surrounding the bulk of the outer segment discs, there were also cone-associated features apparent. These include tapered outer segments and the occasional infolding of the plasma membrane. On the basis of the continuity of the outer segment discs with the extracellular matrix and a pattern of protein labelling throughout the outer segment (following the incorporation of radioactively labelled amino acids to indicate the process of outer segment disc renewal), both receptor types were considered cones in the sea lamprey, Petromyzon marinus (Dickson & Graves 1979). The shape of the outer segment (cylindrical in rods and tapered in cones) has traditionally been used to characterize receptors, but this has given equivocal results in many species owing to the similarity of each receptor type in different regions of the retina, often a product of variations in receptor size and packing. This ambiguity has been especially difficult to reconcile in the Southern Hemisphere lamprey, G. australis, where five morphological types of photoreceptors have been differentiated on criteria as subtle as the ultrastructural aggregation of short wavelength-absorbing pigment within the myoid (Collin et al. 1999; Collin & Trezise 2004), a feature that could easily be overlooked (Meyer-Rochow & Stewart 1996). Based on the continuity of the outer segment discs with the extracellular matrix in all five morphological photoreceptor types in G. australis, all are considered cone-like. The short and long receptors of holarctic lampreys have also been difficult to assign rod- or cone-like status; authors have variously considered them both to be rods (Öhman 1971, 1976; Holmberg & Öhman 1976; Dickson & Graves 1982), both cones (Dickson & Graves 1979) or a rod and a cone, respectively (Crescitelli 1956, 1972; Govardovskii & Lychakov 1984; Ishikawa et al. 1987; Negishi et al. 1987; Hárosi & Kleinschmidt 1993; Yoshida & Tonosaki 1994). Other morphological features, which are less ambiguous and underlie important electrochemical signalling during the phototransduction cascade, e.g. the arrangement of the plasma membrane surrounding the outer segment (Cohen 1972), may be better features upon which to differentiate rods and cones, although these traditional criteria may not be definitive without taking into account spectral, molecular and ultimately functional features. Although spectral analyses may also be ambiguous because the spectral sensitivity of rods overlaps with a range of MWS cones, amino acid sequences of the opsin proteins may hold similar ambiguities. We consider that phototransduction genes may, in fact, hold vital clues to the identity and/or function of the earliest photoreceptors. It appears that the RhA and RhB opsins identified by Collin et al. (2003a,b) in G. australis may share features in common with the Rh1 (found in rods) and Rh2 (found in cones) Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 2928 S. P. Collin et al. Review. Vertebrate photoreceptor evolution (b) normalized absorbance (a) 439 359 1.0 492 497 560 359 0.25 (d) 517 1.0 525 555 600 0.75 0.5 0.25 (f) 477 502 504 561 514 normalized absorbance 552 560 0.5 (e) 1.0 0.75 0.5 0.25 (g) normalized absorbance 497 0.75 (c) normalized absorbance 439 366–383 479–484 (h) 540 558–584 623–656 479–484 540 558–584 623–656 1.0 0.75 0.5 0.25 300 400 500 wavelength (nm) 600 700 300 400 500 wavelength (nm) 600 700 Figure 1. Schematic representations of the spectral sensitivities of the visual pigments of the different photoreceptor types found in early vertebrates. (a) The downstream and (b) upstream migrants of the Southern Hemisphere lamprey, G. australis. The long-wavelength shift (from 492 to 552 nm) of the MWS pigment is caused by the replacement of the yellow myoidal pigment with an ellipsosome. (c) The river lamprey, L. fluviatilis. (d) The sea lamprey, P. marinus. (e) The Southern Hemisphere lamprey, M. mordax. ( f ) The giant shovelnose ray, G. typus. (g) The juvenile stage of the Australian lungfish, N. forsteri. (h) The adult stage of N. forsteri. Note the loss of the UVS pigment in the adult lungfish. Data predominantly based on microspectrophotometric (MSP) analyses but spectrophotometric analysis of recombinant pigments regenerated with 11-cis-retinal have also been included where MSP was not possible. lmax values (indicated) were obtained from Govardovskii & Lychakov (1984); Hárosi & Kleinschmidt (1993); Collin et al. (2003b, 2004) and Hart et al. (2004, 2008). The dashed lines in (g) and (h) indicate the long-wavelength shift in absorbance after taking into account the intracellular filtering produced by the oil droplets and myoidal pigment. opsins, respectively, of jawed vertebrates (see below). However, the amino acid sequence of the RhA opsin in lampreys is intermediate between rod and cone opsins with a rod-type Glu122 and a cone-type Pro189 (Imai et al. 2005). Phil. Trans. R. Soc. B (2009) The process of capturing a photon and transforming it into a biochemical signal (phototransduction) is an ancient process involving a diverse range of chemical reactions that nevertheless shows a striking similarity in its component parts in both invertebrates Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 Review. Vertebrate photoreceptor evolution human mouse platypus dunnart chicken pigeon frog green anole coelacanth lungfish 40 100 98 100 59 91 99 99 S. P. Collin et al. 87 2929 RhA/Rh1 goldfish zebrafish catshark 100 100 dogfish skate pouched lamprey 100 99 100 100 Arctic lamprey 99 100 100 100 94 chicken pigeon green anole lungfish coelacanth 100 100 77 pouched lamprey chicken 87 pigeon 95 platypus 100 green anole frog 100 63 lungfish 100 zebrafish 100 goldfish pouched lamprey human 100 mouse 100 dunnart 100 96 wallaby chicken 99 90 81 pigeon green anole 94 lungfish frog 100 pouched lamprey 100 goldfish zebrafish 100 human 97 mouse dunnart 100 100 wallaby platypus 97 97 frog 98 lungfish goldfish 100 97 84 zebrafish green anole chicken 98 100 pigeon RhB/Rh2 goldfish zebrafish 100 72 0.1 100 SWS2 SWS1 LWS1 pouched lamprey Arctic lamprey Figure 2. Phylogeny of visual pigment (opsin) gene families (LWS, SWS1, SWS2, RhB/Rh2 and RhA/Rh1) in vertebrates. The tree was generated by using a Bayesian probabilistic inference method with a Metropolis Markov chain Monte Carlo algorithm. A general time-reversal model was used with posterior probability values (represented as a percentage) indicated at the base of each node. The scale bar indicates the number of nucleotide substitutions per site. The fruitfly Rh4 was used as an outgroup. Phil. Trans. R. Soc. B (2009) Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 2930 S. P. Collin et al. Review. Vertebrate photoreceptor evolution and vertebrates. In vertebrates, many of these component processes (e.g. transducins, arrestin, phosphodiesterase 6 and the cyclic guanosine monophosphate-gated channels) are encoded by different cone- or rod-specific isoforms (Hisatomi & Tokunaga 2002). These proteins mediate the reactions at the different stages of the biochemical cascade. After photon capture and the isomerization of the chromophore from the 11-cis to all-trans form, the resulting conformational change in the opsin protein is converted via the phototransduction cascade to a biochemical signal recognizable by the other retinal neurons. Preliminary results for the lamprey G. australis reveal that, in general, only single isoforms of the phototransduction genes are present and, with the exception of transducin-a, the coding sequences are phylogenetically more cone- than rod-like (Hunt et al. 2007). Therefore, it may be that at the time in evolutionary history when the jawed and jawless vertebrates split into two lineages, the eyes of lampreys possessed a predominantly photopic visual system based on cone-like photoreceptors. However, at least one type of receptor was undergoing some sort of ‘transmutation’ to a receptor hybrid, which would subsequently become a ‘true rod’ with the age of cartilaginous fishes (Walls 1942; Collin & Trezise 2006; Pisani et al. 2006; Davies et al. 2009). (b) Spectral sensitivity The wavelength of maximum absorbance (lmax) of the visual pigment in a given photoreceptor can be a useful, but not definitive, criterion for discriminating between the spectral sensitivity of rods and cones, although in many species this value can be identical (Bowmaker 1995, 2008; figure 1). There has been no microspectrophotometric analysis of the spectral absorbance of individual photoreceptor types in hagfishes. However, behavioural experiments have revealed that swimming responses (preceded by movements of the head or body) to light stimulation were maximal between 500 and 520 nm, with little or no response above 600 nm (Steven 1955). The spectral characterization of photoreceptor types in lampreys has received more attention. Microspectrophotometric studies of the short and long photoreceptor types in two Northern Hemisphere (holarctic) species (the sea lamprey P. marinus and the river lamprey Lampetra fluviatilis) reveal different peak spectral sensitivities (lmax) values. These lie at 525 and 600 nm (in upstream P. marinus both receptor types possess visual pigments that are porphyropsins or visual pigments that utilize a vitamin-A2 chromophore; (Hárosi & Kleinschmidt 1993; figure 1d ) and at 517 and 555 nm (in upstream L. fluviatilis both receptor types possess visual pigments that utilize a vitamin A1-based chromophore; Govardovskii & Lychakov 1984; figure 1c), respectively. Therefore, it appears that the short and long photoreceptors in holarctic lampreys possess an MWS and an LWS visual pigment within their outer segment, respectively. In the Southern Hemisphere lamprey, G. australis, conventional spectrophotometric analysis of Phil. Trans. R. Soc. B (2009) recombinant opsins expressed in mammalian cells reveals five visual pigments with lmax values at 359, 439, 497, 492 and 560 nm (the LWS opsin failed to generate a functional pigment when expressed as a recombinant protein so the lmax value was based on key spectral tuning sites) (Davies et al. 2007; figure 1a,b). Earlier microspectrophotometric studies on three of the five photoreceptor types indicated that that these photoreceptors utilize an A2-based chromophore on the basis of the goodness of fit of the mean absorbance spectra to mathematical pigment templates (Govardovskii et al. 2000; Collin et al. 2003a; figure 1a). The lmax of the visual pigment within the outer segment of the single photoreceptor type found in the retina of the downstream migrant phase of M. mordax is 514 nm and is, therefore, MWS (Collin et al. 2004; figure 1e). Based on its lmax, the receptor type in M. mordax may be homologous to the short receptors of L. fluviatilis (517 nm, A1; figure 1c) and P. marinus (525 nm, A2; figure 1d ). The photoreceptors of both downstream and upstream migrants of the Pacific lamprey, Entosphenous tridentatus, possess a vitamin-A1-based visual pigment (Crescitelli 1972). When considered together with the finding of the entire complement of photoreceptors possessing visual pigments incorporating a chromophore based on vitamin-A2 in the white sturgeon, Acipenser transmontanus (an early ray-finned fish with an anadromous life cycle), it appears that migration between freshwater and saltwater may not be sufficient to induce a paired A1/A2 visual pigment system (Whitmore & Bowmaker 1989; Sillman et al. 1995). At this stage, and given the considerable interspecific variability, it is perhaps premature to predict whether vitamin A1 or A2 is the ancestral chromophore in vertebrates. However, it is interesting to note that only vitamin-A1, and not vitamin-A2, could be isolated from the liver in Myxine glutinosa (Steven 1955; Vigh-Teichmann et al. 1984). Using microspectrophotometry, three species of ray (Rhinobatos (Glaucostegus) typus, Aptychotrema rostrata and Dasyatis kuhlii) have been found to possess three cone types in the short wavelength-sensitive (SWS, lmax 459 – 477 nm), MWS (lmax 492 – 502 nm) and long- (lmax 552 – 561 nm) spectral ranges in addition to a rod (lmax 497 – 504 nm) (Hart et al. 2004; Theiss et al. 2007; figure 1f ). These findings strongly suggest the potential for colour discrimination in this class of vertebrates that was once thought to be monochromatic, i.e. Chondrichthyes, and support earlier reports of three peaks in electrophysiologically recorded spectral sensitivity at 476, 502 and 540 nm in the closely related stingray, Dasyatis pastinaca, by Govardovskii & Lychakov (1977). The retina of the Australian lungfish is duplex, containing both rod and cone photoreceptors. Microspectrophotometry reveals that juvenile lungfish possess four types of single cone photoreceptor with visual pigments maximally sensitive to ultraviolet (UVS, lmax 366 nm), SWS (lmax 479 nm), MWS (lmax 558 nm) and LWS (lmax 623 nm) and, therefore, have the potential for tetrachromatic colour vision (Bailes et al. 2006; Hart et al. 2008; Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 Review. Vertebrate photoreceptor evolution figure 1g,h). UVS cones are the only photoreceptor type not found in the retina of the adult lungfish. The retinae of both juveniles and adults also contain a single type of MWS photoreceptor (lmax 540 nm, figure 1g,h). On the basis of spectral bandwidth and the fit of the absorbance spectra to visual pigment templates (Govardovskii et al. 2000), all visual pigments in the lungfish are considered to use an A2-based chromophore (or porphyropsin), as is typical of many other freshwater fish (Hart et al. 2008). (c) Opsins and spectral tuning Molecular examination of photoreceptor visual pigments is an important and more definitive method of characterizing rods and cones (especially when combined with in situ hybridization) and assessing the potential for colour vision. As described previously, the chromophore that is bound to the visual opsins of vertebrates is derived from the aldehyde of either vitamin-A1 or vitamin-A2 to give a rhodopsin or porphyropsin pigment, respectively. Porphyropsin pigments are spectrally more long-wavelength-shifted than the corresponding rhodopsin, and this effect increases progressively at longer wavelengths (Whitmore & Bowmaker 1989; Hárosi 1994; Parry & Bowmaker 2000; Temple et al. 2008). Packed into the membranes of the discs within the outer segments, these transmembrane proteins comprise a polypeptide chain of approximately 350 amino acids, which forms seven transmembrane alpha-helices linked by extraand intracellular loops. Changes in the photoreceptor spectral sensitivity are mediated primarily by variations in the amino acid sequence of the opsin protein, each encoded by a distinct opsin gene and by different ratios of the two chromophores (Nathans et al. 1986; Partridge 1995). Vertebrate retinal opsin genes are classified into five evolutionarily distinct groups: LWS opsins found in cones (LWS/MWS), SWS 1 opsins found in cones (SWS1), SWS 2 opsins found in cones (SWS2), MWS opsins found in cones (RhB/ Rh2,), and MWS opsins primarily found in rods (RhA/Rh1) (Yokoyama 1997; Bowmaker & Hunt 2006; figure 5). Although Rh1 opsin-based pigments are usually expressed in rods and the remainder in cones, as indicated previously (and see below), the closely related RhA opsin has some cone-like characteristics. Gene duplication, followed by nucleotide substitution, provides the basis of functional differentiation of the various groups of opsins. Currently, there are no retinal visual pigments characterized in hagfishes. However, Hisatomi et al. (1991) were the first to clone and sequence a fulllength opsin gene from the river lamprey, Lampetra japonica. The deduced amino acid sequence was most similar to Rh1 rod opsin sequences of other vertebrates, showing 78 – 82% identity. The in vitro expressed lamprey pigment, reconstituted with 11-cis-retinal, showed an absorption maximum of 500 nm and reacted with an anti-bovine rod pigment (Rh1) antibody, which had previously been found to recognize the short photoreceptor type in L. japonica (Hisatomi et al. 1997). Interestingly, and unlike the rod Rh1 opsins of other vertebrates, this lamprey Phil. Trans. R. Soc. B (2009) S. P. Collin et al. 2931 pigment (initially classed as Rh1) bleached gradually in the presence of hydroxylamine, even in the dark, which is usually a characteristic of cone pigments (Okano et al. 1989). Therefore, this pigment has biochemical characteristics intermediate between rod and cone pigments. Similarly, an opsin gene in the marine lamprey, P. marinus, has a deduced amino acid sequence that shows a 92 per cent identity to the Rh1 opsin gene of the river lamprey, L. japonica (Zhang & Yokoyama 1997). The full extent of the palette of visual pigment opsin genes available to the ancestral vertebrates would appear to be realized within the Southern Hemisphere lamprey, G. australis, where all five opsin genes are expressed within the retina and differentially regulated during the lamprey life cycle (Collin et al. 2003a; Davies et al. 2007; figure 2). Based on neighbourjoining, maximum likelihood and Bayesian inference methods used to reconstruct phylogenetic trees, opsin genes were identified as the orthologues of the jawed vertebrate LWS, SWS1 and SWS2 opsin gene lineages. In addition, two Rh classes of opsin genes were identified (RhA and RhB), which appear to be most closely related to the gnathostome Rh1 and Rh2 genes, respectively. This finding suggests that (i) the most recent common ancestor of the jawed and jawless vertebrates possessed a single ancestral Rh opsin gene, the common ancestor of all the Rh opsin genes, as well as LWS, SWS1 and SWS2 opsin genes, with separate gene duplications giving rise to the RhA and RhB opsin genes in jawless (agnathan) vertebrates and the Rh1 and Rh2 opsin genes in the jawed (gnathostome) vertebrates (Collin et al. 2003a; Collin & Trezise 2004) or (ii) the RhA opsin found in the two species of lampreys examined (the pouched lamprey, G. australis, and the arctic lamprey, Lethenteron japonica) would appear to be ancestral to the Rh1 class of opsin and that the RhB opsin in G. australis is ancestral to the Rh2 opsin group, suggesting that lamprey RhA and RhB genes share ancestral orthologues with the jawed vertebrate Rh1 and Rh2 gene classes, respectively (Pisani et al. 2006; Davies et al. 2009; figure 2). Holocephalans and elasmobranchs are a large class of vertebrates that have colonized both deep sea and shallow water, especially coral reef environments. Up until recently, only the Rh1 opsin gene had been isolated and cloned in elasmobranchs, i.e. in the catshark, dogfishes and the skate (O’Brien et al. 1997; Bozzano et al. 2005). However, an analysis of the genome of the holocephalan elephant shark, Callorhinus milii, has revealed that a rod (Rh1) and three cone (Rh2, LWS1 and LWS2) opsin genes are expressed within the retina, with lmax values of 496 (Rh1), 442 (Rh2), 499 (LWS1) and 548 nm (LWS2) as determined by in vitro expression (Davies et al. 2009). The research on C. milii represents the first to reveal the molecular identity of any cone opsin gene within the Chondrichthyes. Together with the discovery of three spectrally different cone photoreceptor types in batoid elasmobranchs (Hart et al. 2004; Theiss et al. 2007), there is now impressive evidence to support the potential for trichromatic vision in cartilaginous fishes. All five opsins are also expressed in the retina of the subadult Australian lungfish (N. forsteri) and a Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 2932 S. P. Collin et al. Review. Vertebrate photoreceptor evolution (a) (b) (c) (d) Figure 3. Axial view of the retinal photoreceptor array in a range of early vertebrates viewed using Nomarski optics. (a) Upstream migrant of the Southern Hemisphere lamprey, G. australis (Agnatha). Photoreceptors are differentiated based on the presence of an ellipsosome (large circular profiles), the presence or absence of an orange/yellow, myoidal short wavelength-absorbing pigment and size. (b) Downstream migrant of the Southern Hemisphere lamprey, M. mordax (Agnatha). This species has only one photoreceptor type containing an ellipsosome arranged within a tightly packed hexagonal array and surrounded by a tapetum housed within the retinal pigment epithelial cells. (c) Epaulette shark, Hemiscyllium ocellatum (Chondrichthyes, Elasmobranchii). This rod-rich retina has a low proportion of cones differentiated on the basis of their small size and refractive properties (Litherland & Collin 2008). (d) Australian lungfish, N. forsteri (Osteichthyes, Dipnoi). Large red oil droplets and orange myoidal pigment (arrowheads) are interspersed with large rods and smaller cones containing clear oil droplets. Scale bars: (a) 20 mm; (b) 25 mm; (c) 5 mm; (d) 35 mm. phylogenetic comparison of codon-matched alignments of both nucleotide and deduced amino acid sequences reveals the presence of LWS, SWS1, SWS2, Rh2 and Rh1 genes (Bailes et al. 2007; figure 2). The characterization of four cone opsins reveals the potential for tetrachromatic vision in lungfish, although behavioural work is needed to verify if lungfish can discriminate objects based on differences in chromatic hue, in addition to whether it has four-dimensional colour space. (d) Ellipsosomes, oil droplets, myoidal pigment and tapeta A photoreceptor’s spectral sensitivity is determined primarily by the spectral absorbance of the visual pigment(s) it contains. However, many species have also evolved intra- and extracellular devices to enhance (filter or tune) the incoming light before it reaches the visual pigments located within the outer segments (reviewed by Douglas & Marshall 1999). These filters come in many forms, including corneal (Collin & Collin 2001; Siebeck et al. 2003), lenticular (Heinermann 1985; Siebeck & Marshall 2001) and retinal, i.e. ellipsosomes (large elliptical inclusions surrounded by mitochondria and lying within the inner segment) and myoidal pigment situated below the Phil. Trans. R. Soc. B (2009) mitochondria (Collin & Potter 2000; Collin et al. 2003b; figures 2a,d and 4a). However, the most notable are coloured oil droplets (which differ from ellipsosomes in both their development and biochemistry; Nag & Battacharjee 1995) located within the ellipsoid of the cone inner segment. These densely pigmented organelles usually act as long-pass cut-off spectral filters, although the location of the cut-off wavelength varies depending on the lmax of the visual pigment (Partridge 1989; Hart 2001; Hart & Vorobyev 2005; figures 2d and 3e) and even environmental factors such as ambient light intensity (Hart et al. 2006a,b). No oil droplets have been identified in the retina of any species of hagfishes or lampreys examined thus far. However, an intracellular yellow/orange photostable pigment is present in the myoid and ellipsoidal regions of at least three of the five photoreceptor types in the Southern Hemisphere lamprey G. australis (Collin et al. 1999; Collin & Trezise 2004; figures 3a and 4c). This yellow-orange pigment, which is perhaps homologous to that found within one of the cone photoreceptor types in the Australian lungfish, N. forsteri (Bailes et al. 2006; figures 3d, 4e and 5) and also the ornate lizard, Ctenophorus ornatus (Barbour et al. 2002; figure 5), acts as a spectral filter by absorbing visible wavelengths below about Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 S. P. Collin et al. Review. Vertebrate photoreceptor evolution (a) (b) 2933 (c) os os os e m m m m m n C C/R C (d) yp1 n yp2 n n C/R m yp1 n C/R n C C C (e) os os m od dm m m C C C R n R od p n p n n yp m C C od m p n C C Figure 4. Schematic representation of the complement of photoreceptor types in early vertebrates based on morphological criteria. (a) The single receptor type in the lamprey, M. mordax (Mordaciidae). (b) The long and short receptors in the Northern Hemisphere (holarctic) lampreys (Petromyzontidae). (c) The five receptor types in the lamprey, G. australis (downstream migrant) (Geotriidae). Note the inclusion of yellow myoidal pigment in three of the receptor types. (d ) The four receptor types in the giant shovelnose ray, G. typus (Rhinobatidae). (e) The five receptor types in juvenile Australian lungfish, N. forsteri (Ceratodontidae). Note the yellow myoidal pigment and both red and colourless oil droplets. Receptor identities have been based on a range of morphological, spectral and molecular studies; C/R, cone/rod hybrid or features that make it difficult to differentiate; C, cone; R, rod; dm, distended mitochondria; e, ellipsosome; m, mitochondria within the inner segment; n, nucleus; od, oil droplet; os, outer segment; p, paraboloid; yp (1 and 2), orange/yellow pigment. 550 nm (Collin et al. 2003b). This short-wavelengthabsorbing filter may have evolved originally in a vertebrate with a similar life history to G. australis, in response to the need to filter out the potentially damaging short-wavelength (high energy) light to which the eye is constantly being subjected. A short-wavelengthabsorbing filter may also prevent photostimulation of the beta-band of the visual pigments and narrow the spectral sensitivity functions of different cone types, thereby improving colour discrimination (Vorobyev 2003; Hart et al. 2008). It is also possible that absorbance of short-wavelength light, at least in some photoreceptor types, may improve contrast sensitivity (Walls 1942). Oil droplets are thought to have evolved early in vertebrate evolution, predating the emergence of Phil. Trans. R. Soc. B (2009) terrestrial vertebrates ca 400 Ma (Robinson 1994). The Australian lungfish, N. forsteri, a living fossil and the closest living relative of the first terrestrial vertebrates, possesses oil droplets within three of its four large cone types (two of which are non-pigmented and one type in the LWS cone has a dense red pigment; Robinson 1994; Bailes et al. 2006; Hart et al. 2008; Marshall et al. in press; figures 3d, 4e and 5). Both pigmented and non-pigmented oil droplets have been retained in diurnal lizards, turtles and birds (Walls 1942; figure 5). The apparent loss of oil droplets in amphibians, snakes, crocodiles and mammals (except the marsupials, which have non-pigmented oil droplets; Arrese et al. 2002), is thought to be the result of a shift from a diurnal to a nocturnal lifestyle and may function to improve photon capture and Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 2934 S. P. Collin et al. Review. Vertebrate photoreceptor evolution focus light onto the outer segment (Walls 1942; Collin & Potter 2000). Although the more widely accepted advantages of oil droplets in reducing chromatic aberration and negating the harmful effects of ultraviolet radiation may still apply (Walls & Judd 1933; Walls 1942; Douglas & Marshall 1999), the improvement in colour discrimination may have been the driving force for the evolution of coloured oil droplets in lungfish, especially those living in brightly lit environments (Vorobyev 2003; Hart et al. 2008). The pigmented oil droplets act as long-wavelength-pass cut-off filters that sharpen and shift the spectral sensitivity of the photoreceptor to longer wavelengths in order to improve spectral discrimination (Kelber et al. 2003; Hart et al. 2008). Non-pigmented oil droplets may have a function in improving photon capture and focusing light on to the outer segment (Hart et al. 2006a,b). However, we consider that the evolutionary longevity of the visual system of the Australian lungfish may also lie partly in its ability to balance the trade-off between resolution and sensitivity. While the large cone diameters (approx. 15 mm in diameter; Bailes et al. 2006; figures 3d and 4e) are most probably a by-product of the reduction in quantal flux in response to the filtering capacity of the coloured inclusions, its large rods (approx. 20 mm in diameter; figures 3d and 4e) will ensure sufficiently high sensitivity to operate in the reportedly low light levels of its crepuscular and/or nocturnal lifestyle (Kemp 1986). The presence of ellipsosomes in the photoreceptors of some lampreys indicates that these early vertebrates have the capacity to alter the light before it is absorbed by the visual pigments. These elliptically shaped intracellular organelles span the diameter of the inner segment (figures 3a,b and 4a) and may be used to focus light on to a narrow, tapered outer segment (Young & Martin 1984), in addition to reducing both photo-oxidative damage (Avery & Bowmaker 1982) and the effects of chromatic aberration (Collin & Potter 2000). Given that ellipsosomes are present in two of the extant agnathan families (Geotriidae and Mordaciidae), it appears likely that ellipsosomes may have been present in the retina in their most recent common ancestor (Collin & Potter 2000; Collin et al. 2003b; figures 3a,d, 4a and 5). Whether oil droplets evolved from such an aggregation of pigment granules is unknown and may be improbable given the packaging of the pigment: spherical membrane-bound pigment found in the ellipsoid region (oil droplets) versus stored secretory material within the endoplasmic reticulum of the myoid region (Collin et al. 1999). The presence of a tapetum is an adaptation to increase sensitivity by reflecting light back through the photoreceptors after striking a reflective layer situated either within the retinal pigment epithelial cells or behind Bruch’s membrane. Depending on the type of tapetum (specular or diffuse), the wavelengths of light re-entering the photoreceptor outer segment may also vary and be tuned to the ambient light environment. Lampreys, elasmobranchs, lungfishes, as well as teleosts, all exhibit an eyeshine produced by tapeta (Collin 1988; Collin & Potter 2000; Bailes et al. 2006; figure 5). The retina of the Southern Hemisphere lamprey, M. mordax, possesses a regular hexagonal array of a single Phil. Trans. R. Soc. B (2009) type of photoreceptor containing an ellipsoidal ellipsosome (figures 3b and 4a) surrounded by an RPE filled with two types of tapetal reflector (spheres and needles). The spherical particles characteristic of the diffuse type of reflector are most common in the vitread region surrounding the photoreceptors, whereas the needleshaped particles that are characteristic of the specular type of reflector are most abundant in the sclerad region of the RPE. It is evident that the enhanced sensitivity gained from a reflective tapetum was an early invention and one that was favoured by natural selection as particular members of each major group escaped predation by moving into perpetually dim light environments and/or needed to enhance sensitivity during crepuscular periods of changes in light intensity and thereby from cone-based to rod-based vision. This scenario is particularly applicable to elasmobranchs, many of which actively move between different light levels and possess a reflective choroidal tapetum sitting behind a retinal array of predominantly rod photoreceptors (Litherland & Collin 2008; figures 3c, 4d and 5). 3. FUNCTIONAL AND BEHAVIOURAL IMPLICATIONS FOR EARLY PHOTORECEPTION The functional divergence of the two receptor types in Northern Hemisphere lampreys has been examined by microspectrophotometry (Govardovskii & Lychakov 1984; Hárosi & Kleinschmidt 1993), visual pigment anion sensitivity (Hárosi & Kleinschmidt 1993), hydroxylamine sensitivity (Hisatomi et al. 1997) and electrophysiology (Govardovskii & Lychakov 1984). In all cases, the results predominantly indicate that both receptor types are cones but uncertainty still exists as regards their identity and function. Electroretinographic responses recorded in the river lamprey, L. fluviatilis, by Govardovskii & Lychakov (1984) show typical cone responses for both receptor types, including the short receptors (putative rods), which do not saturate at high light intensities and contribute to both photopic and scotopic vision. The short receptors are, however, approximately 100 times more sensitive than the long receptors and are the only class of receptor to be active in the dark-adapted state (Govardovskii & Lychakov 1984). The long receptors contribute to retinal function only during intense stimulation and can therefore be regarded as cones. Moreover, changes in stimulus colour always produces a large response of a complex form, irrespective of the relative strength of the stimuli, indicating that under photopic conditions the lamprey retina possesses the ability of colour discrimination. Future functional studies on the photokinetics of the complement of photoreceptor types in G. australis and M. mordax will undoubtedly help elucidate the rod/ cone hybrid nature of these photoreceptor types. No functional studies of the one or two receptor types hypothesized to exist in hagfish eyes have been undertaken. However, based on a series of testable hypotheses, Lamb et al. (2007, 2008) consider the eyes to be non-image-forming bilateral ‘pineal’ organs that have been retained for over 500 Myr as a circadian irradiance detector. Similarly, few functional studies have been undertaken in elasmobranchs. Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 Review. Vertebrate photoreceptor evolution Dowling & Ripps (1972) reveal that, in the pure-rod skate retina, the rod photoreceptors respond to incremental stimuli up to and including bright light intensities that would bleach a large fraction of the visual pigment present, much like typical cones in other jawed vertebrates. Later research using electroretinograms also confirms that the all-rod retina of the skate possesses both ON and OFF bipolar cell pathways that are functional (Chappell & Rosenstein 1996). (a) Colour vision The detection of all aspects of the aquatic visual scene including prey, predator or mate is dramatically improved by the added dimension of chromatic detail and the large variety of identified photoreceptor visual pigments in vertebrates indicates that colour vision offers a substantial selective advantage. In order to mediate colour vision, multiple sensors (each providing a means for the differential filtering of spectral information) and neuronal comparators (designed to contrast signals originating from these different sensors) must both be present if the chromatic signal is to elicit a behavioural response (Jacobs & Rowe 2004). In vertebrates, molecular phylogenetic analyses have established that vertebrate visual pigments evolved along five distinct lines and that these lineages may have been in place before the divergence of jawed and jawless fishes (Davies et al. 2009; figure 2). The five vertebrate visual pigment gene lineages presumably diverged from a single common ancestor that had evolved prior to the divergence of the protostomes and deuterostomes, which is thought to have occurred about 700 Ma (Oota & Saitou 1999). Therefore, colour vision most probably evolved in the earliest vertebrates between 400 and 700 Ma. However, our ability to pinpoint the origins of colour vision or colour discrimination within this time frame are still a matter of contention arising from the lack of data on cephalochordates, tunicates and hagfishes, which lie at the base of chordate phylogeny. However, new evidence now suggests that colour vision evolved between 700 and at least 540 Ma. This is based on the presence of five types of cone-like photoreceptors in the Southern Hemisphere lamprey, G. australis (Collin et al. 2003b; Collin & Trezise 2004; figure 4c), three cone types in chondrichthyan holocephalans and elasmobranchs (Hart et al. 2004; Theiss et al. 2007; Davies et al. 2009; figure 4d). The finding of four cone types in the Australian lungfish, N. forsteri, allows us to make inferences about the colour vision of the first terrestrial vertebrates (Bailes et al. 2006; Hart et al. 2008; figure 4e). Molecular studies in a range of teleost fishes reveal that not only are these classical cone opsin genes retained in bony fishes (including an Rh1 opsin; figure 2) but a series of consecutive gene duplication events within the Rh2 (to produce Rh2a, Rh2Ab and Rh2B in green cones) and SWS2 (to produce SWS2A and SWS2B in blue cones) gene lineages have produced up to seven cone opsin genes, which are functionally expressed in the retina (Parry et al. 2005; Trezise & Collin 2005; Phil. Trans. R. Soc. B (2009) S. P. Collin et al. 2935 Shand et al. 2008). It is proposed that the rapid and successful speciation of cichlids, which has produced about a thousand new species, in the African Great Lakes over the past 250 000 years may have been partly due to the ability of this group to adapt quickly to new ecological niches by ‘exploring new genetic space’ by a series of gene duplications (Carleton et al. 2005; Parry et al. 2005; Trezise & Collin 2005). Multiple cone opsins found in representatives of all of the early vertebrate classes, together with a range of coloured intraocular filtering mechanisms, suggest that the ancestral vertebrates relied on diurnal vision and may have utilized colour discrimination to target prey, avoid predators, communicate and attract reproductive partners. (b) The role of vision in early vertebrates Given the rather simple and poorly organized photoreceptor outer segments of hagfishes, it is of little surprise that behaviourally they are almost ‘blind’, and their weak response to light is unaffected by the removal of the eyes (Newth & Ross 1955). Lamb et al. (2007, 2008) suggest that the ‘eyes’ of hagfishes may act more like the pineal of gnathostomes (hagfishes lack a true pineal). Unfortunately, there are very few studies examining visual behaviour in lampreys. The ability to discriminate a broad range of colours and view the world tetra- (or even penta-) chromatically may be advantageous to extant lampreys living in brightly lit environments. The countershading patterns of the Southern Hemisphere lamprey G. australis with a bright iridescent, blue dorsal surface and a silver ventral surface are unique features within the Agnatha. Presumably this is an adaptation to its pelagic marine trophic phase, where it spends much of its time in the clear, blue surface waters off the coast of South Georgia feeding parasitically on a range of teleosts. Although parasitic, Nikol’skii (1956) describes L. japonica as ‘predatory’, gnawing out tissues and even internal organs with its specialized mouthparts. Possessing no paired fins and swimming by propagating a laterally directed wave along its eel-like body (Grillner et al. 1991), lampreys can steer with great precision towards a specific location in the water column to locate potential hosts, using visual cues for orientation during locomotion (Grillner et al. 1991; Ullén et al. 1995). Mordacia mordax are known to frequent deeper water, diving to depths of up to 1000 m (Bigelow & Schroeder 1953) and, like the upstream migrants of G. australis, are nocturnal. Generally, elasmobranch colours tend to be muted in comparison to the bright colours seen in some teleost fishes (Kemp 1999), although some elasmobranch species are coloured for camouflage and/or sexual selection. Until behavioural experiments are carried out, it will be difficult to understand the functional existence of up to three cone types and the potential for trichromatic colour vision in rays and elephant sharks (Hart et al. 2006a,b; Davies et al. 2009; figure 4d ). Most elasmobranchs display some degree of countershading, which tends to be most apparent in active, pelagic sharks, such as the blue shark, Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 2936 S. P. Collin et al. Review. Vertebrate photoreceptor evolution od cod mp tp Tetrapods coloured oil droplets (od) lungfishes od mp tp coelacanth cod tp colourless oil droplets (cod) cod el tp ray-finned fishes elasmobranchs tp el mp tp lampreys ellipsosomes (el) myoidal pigment (mp) tapeta (tp) hagfishes ? Figure 5. Phylogenetic schematic of the major vertebrate groups showing the evolution of tapeta (tp), myoidal short wavelength-absorbing pigment (mp), ellisosomes (el), colourless oil droplets (cod) and coloured oil droplets (od). Black bars represent the first appearance of each of the morphological traits. The schematic drawings of the photoreceptors on the right-hand side depict the complement of receptor types within at least one species within that vertebrate class/group. The lists of visual traits adjacent to these drawings indicate that at least one species possesses this feature. If a particular feature is not listed for a particular clade, it has not been identified (to our knowledge) in any species within that class/group. The phylogeny is based on that of Meyer & Zardoya (2003). At present, the relationships between hagfishes and lampreys and between lungfishes and the coelacanth are considered polytomies but have been depicted here as being monophyletic based on other visual characters. Prionace glauca, and the shortfin mako shark, Isurus oxyrhinchus, while benthic or bottom-dwelling species often have characteristic markings that contrast the background colour and appear as spots, stripes, bars, blotches or reticulations (Kemp 1999; Hart et al. 2006a,b). In many species, the role of these patterns has been interpreted as camouflage, although it is not known whether the animals themselves use differences in colour and patterning to recognize conspecifics. In the nurse shark, Ginglymostoma cirratum, females appear to select larger and darker coloured males (Pratt & Carrier 2001), suggesting that body colouration and/or contrast could be used as a cue by females to select mates in this species. The Australian lungfish N. forsteri has the potential for a complex colour vision system based on either trichromacy (adults) or tetrachromacy (juveniles). Also, Phil. Trans. R. Soc. B (2009) by reducing the overlap of the spectral sensitivities of adjacent spectral cone types by long-pass spectral filters (oil droplets), there is an improvement in colour discrimination, despite the unavoidable reduction in total photon catch (Vorobyev 1997). Previous studies have highlighted the theoretical advantages of retinal spectral filters for colour discrimination in terrestrial birds and semiaquatic freshwater turtles, but it appears that this mechanism has its origin within the aquatic realm (Hart et al. 2008). 4. CONCLUSIONS In summary, colour vision is inferred to have evolved in the earliest vertebrates more than 540 Ma, providing the basis for colour discrimination in all extant vertebrate classes found today. The evolution of multiple Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 Review. Vertebrate photoreceptor evolution cone types in the first vertebrates (the lampreys) provides important clues to the basis of visual communication, not only in fishes but in all gnathostomes. Further studies are required to establish the selective forces driving the evolution of photoreception in lampreys, hagfishes and other organisms that represent the invertebrate –vertebrate transition and to increase our understanding of the phylogenetic basis of the duplicity theory (or the evolution of the functional dichotomy of true rod- and cone-based vision) and early photoreception. It is clear that before vertebrates ventured onto land, they possessed a well-developed colour vision system able to balance the trade-off between resolution and sensitivity. Multi-disciplinary approaches will continue to identify the evolutionary constraints placed upon the shape, light responses, spectral sensitivity and molecular structure of photoreceptors in early vertebrates and their role in visual behaviour. The authors would like to thank Ian Potter, Ann Trezise, Maree Knight, Justin Marshall, Misha Vorobyev and Helena Bailes for useful discussions and/or taking part in previous research forming part of this review. This work was supported by the Australian Research Council and the UK Biotechnology and Biological Sciences Research Council. REFERENCES Arrese, C. A., Hart, N. S., Thomas, N., Beazley, L. D. & Shand, J. 2002 Trichromacy in Australian marsupials. Curr. Biol. 12, 657–660. (doi:10.1016/S09609822(02)00772-8) Avery, J. A. & Bowmaker, J. K. 1982 Visual pigments in the four-eyed fish. Anableps anableps. Nature 298, 62–64. (doi:10.1038/298062a0) Bailes, H. J., Robinson, S. R., Trezise, A. E. O. & Collin, S. P. 2006 Morphology, characterisation and distribution of retinal photoreceptors in the Australian lungfish Neoceratodus forsteri (Krefft, 1870). J. Comp. Neurol. 494, 381 –397. (doi:10.1002/cne.20809) Bailes, H. J., Davies, W. L., Trezise, A. E. O. & Collin, S. P. 2007 Visual pigments in a living fossil, the Australian lungfish Neoceratodus forsteri. BMC Evol. Biol. 7, 200. (doi:10.1186/1471 –2148–7-200) Barbour, H. R., Archer, M. A., Hart, N. S., Thomas, N., Dunlop, S. A., Beazley, L. D. & Shand, J. 2002 Retinal characteristics of the ornate dragon lizard, Ctenophorous ornatus. J. Comp. Neurol. 450, 334–344. (doi:10.1002/ cne.10308) Bardack, D. 1991 First fossil hagfish (Myxinoidea): a record from the Pennsylvanian of Illinois. Science 254, 701– 703. (doi:10.1126/science.254.5032.701) Bardack, D. 1998 Relationships of living and fossil hagfishes. In The Biology of Hagfishes (eds J. M. Jorgensen, J. P. Lomholt, R. E. Weber & H. Malte), pp. 3 –14. London, UK: Chapman and Hall. Bigelow, H. B. & Schroeder, W. C. 1953 Fishes of the Gulf of Maine. Fishery Bull. Fish. Wildl. Serv. USA 53, 1 –577. Bowmaker, J. K. 1995 The visual pigments of fish. Prog. Ret. Eye Res. 15, 1 –31. (doi:10.1016/1350-9462(95)00001-1) Bowmaker, J. K. 2008 Evolution of vertebrate visual pigments. Vis. Res. 48, 2022–2041. (doi:10.1016/j.visres. 2008.03.025) Bowmaker, J. K. & Hunt, D. M. 2006 Opsin primer. Curr. Biol. 16, R484–R489. (doi:10.1016/j.cub.2006.06.016) Phil. Trans. R. Soc. B (2009) S. P. Collin et al. 2937 Bozzano, A., Mugia, R., Vallerga, S., Hirano, J. & Archer, S. 2005 The photoreceptor system in the retinae of two dogfishes, Scyliorhinus canicula and Galeus melastomus: possible relationship with depth distribution and predatory lifestyle. J. Fish Biol. 59, 1258–1278. (doi:10.1111/ j.1095-8649.2001.tb00190.x) Brinkmann, H., Venkatesh, B., Brenner, S. & Meyer, A. 2004 Nuclear protein coding genes support lungfish and not the coelacanth as the closest living relatives of land vertebrates. Proc. Natl Acad. Sci. USA 101, 4899 – 4905. (doi:10.1073/pnas.0400609101) Carleton, K. L., Parry, J. W. L., Bowmaker, J. K., Hunt, D. M. & Seehausen, O. 2005 Colour vision and speciation in Lake Victoria cichlids of the genus Pundamilia. Mol. Ecol. 14, 4341–4353. Carroll, R. L. 1997 Patterns and processes of vertebrate evolution. Cambridge, UK: Cambridge University Press. Chappell, R. L. & Rosenstein, F. J. 1996 Pharmacology of the skate electroretinogram indicates independent ON and OFF bipolar cell pathways. J. Gen. Physiol. 107, 535–544. (doi:10.1085/jgp.107.4.535) Cohen, A. I. 1972 Rods and cones. In Handbook of sensory physiology, vol VII/2. (ed. M. G. F. Fuortes), pp. 63–110. New York, NY: Springer. Collin, S. P. 1988 The retinal structure of the shovel-nosed ray, Rhinobatos batillum (Rhinobatidae). Morphology and quantitative analysis of ganglion, amacrine and bipolar cell populations. Exp. Biol. 47, 195 –207. Collin, S. P. & Collin, H. B. 2001 The fish cornea: adaptation for different aquatic environments. In Sensory biology of jawed fishes. New insights (eds B. G. Kapoor & T. J. Hara). pp. 57–96. Enfield (NH), USA: Science Publishers Inc. Collin, S. P. & Potter, I. C. 2000 The ocular morphology of the southern hemisphere lamprey Mordacia mordax Richardson with special reference to a single class of photoreceptor and a retinal tapetum. Brain Behav. Evol. 55, 120 –138. (doi:10.1159/000006647) Collin, S. P. & Trezise, A. E. O. 2004 The origins of colour vision in vertebrates. Clin. Exp. Optom. 87, 217 –223. (doi:10.1111/j.1444-0938.2004.tb05051.x) Collin, S. P. & Trezise, A. E. O. 2006 Molecular evidence for dim-light vision in the last common ancestor of the vertebrates—response. Curr. Biol. 16, R320. (doi:10. 1016/j.cub.2006.03.091) Collin, S. P., Potter, I. C. & Braekevelt, C. R. 1999 The ocular morphology of the southern hemisphere lamprey Geotria australis Gray, with special reference to optical specialisations and the characterisation and phylogeny of photoreceptor types. Brain Behav. Evol. 54, 96– 118. (doi:10.1159/000006616) Collin, S. P., Knight, M. A., Davies, W. L., Potter, I. C., Hunt, D. M. & Trezise, A. E. O. 2003a Ancient colour vision: multiple opsin genes in the ancestral vertebrates. Curr. Biol. 13, R864–R865. (doi:10.1016/j.cub.2003.10. 044) Collin, S. P., Hart, N. S., Shand, J. & Potter, I. C. 2003b Morphology and spectral absorption characteristics of retinal photoreceptors in the southern hemisphere lamprey (Geotria australis). Vis. Neurosci. 20, 119– 130. (doi:10.1017/S0952523803202030) Collin, S. P., Hart, N. S., Wallace, K. M., Shand, J. & Potter, I. C. 2004 Vision in the southern hemisphere lamprey Mordacia mordax: spatial distribution, spectral absorption characteristics and optical sensitivity of a single class of retinal photoreceptor. Vis. Neurosci. 21, 765– 773. (doi:10.1017/S0952523804215103) Compagno, L. J. V., Dando, M. & Fowler, S. 2005 In A field guide to the sharks of the world. London, UK: Collins. Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 2938 S. P. Collin et al. Review. Vertebrate photoreceptor evolution Crescitelli, F. 1956 The nature of the lamprey visual pigment. J. Gen. Physiol. 39, 423– 435. (doi:10.1085/jgp. 39.3.423) Crescitelli, F. 1972 The visual cells and visual pigments of the vertebrate eye. In Handbook of sensory physiology, VII/1, photochemistry of vision (ed. H. J. A. Dartnall), pp. 245 –363. Berlin, Germany: Springer-Verlag. Davies, W. L., Cowing, J. A., Carvalho, L. S., Potter, I. C., Trezise, A. E. O., Hunt, D. M. & Collin, S. P. 2007 Functional characterization, tuning and regulation of visual pigment gene expression in an anadromous lamprey. FASEB J. 21, 1–12. (doi:10.1096/fj.06-8057com) Davies, W. L., Carvalho, L. S., Tay, B. H., Brenner, S., Hunt, D. M. & Venkatesh, B. 2009 Into the blue: gene duplication and loss underlie color vision adaptations in a deep-sea chimaera, the elephant shark Callorhinus milii. Genome Res. 19, 415 –426. (doi:10.1101/gr. 084509.108) Dickson, D. H. & Collard, T. R. 1979 Retinal development in the lamprey (Petromyzon marinus L): premetamorphic ammocoete eye. Am. J. Anat. 154, 321–336. (doi:10. 1002/aja.1001540303) Dickson, D. H. & Graves, D. A. 1979 Fine structure of the lamprey photoreceptors and retinal pigment epithelium (Petromyzon marinus L.). Exp. Eye Res. 29, 45– 60. (doi:10.1016/0014-4835(79)90165-9) Dickson, D. H. & Graves, D. A. 1982 The ultrastructure and development of the eye. In The biology of lampreys, vol. 3 (eds M. W. Hardisty & I. C. Potter), pp. 43– 94. London, UK: Academic Press. Douglas, R. H. & Marshall, N. J. 1999 A review of vertebrae and invertebrate ocular filters. In Adaptive mechanisms in the ecology of vision. (eds S. N. Archer, M. B. A. Djamgoz, E. R. Loew, J. C. Partridge & S. Vallerga), pp. 95–162. Dordrecht, The Netherlands: Kluwer Academic Press. Dowling, J. E. & Ripps, H. 1972 Adaptation in skate photoreceptors. J. Gen. Physiol. 60, 698 –719. (doi:10.1085/jgp. 60.6.698) Fernholm, B. 1998 Hagfish systematics. In The biology of hagfishes (eds J. M. Jorgensen, J. P. Lomholt, R. E. Weber & H. Malte), pp. 33–44. London, UK: Chapman and Hall. Gill, H. S., Renaud, C. B., Chapleau, F., Mayden, R. L. & Potter, I. C. 2003 Phylogeny of living parasitic lampreys (Petromyzontiformes) based on morphological data. Copeia 2003, 687–703. (doi:10.1643/IA02-085.1) Govardovskii, V. I. & Lychakov, L. V. 1977 Visual cells and visual pigments in the Black Sea elasmobranchs (In Russian). Zh. Evol. Biokhim. Fiziol. 13, 162–166. Govardovskii, V. I. & Lychakov, D. V. 1984 Visual cells and visual pigments of the lamprey, Lampetra fluviatilis L. J. Comp. Physiol. A. 154, 279 –286. (doi:10.1007/ BF00604994) Govardovskii, V. I., Fyhrquist, N., Reuter, T., Kuzmin, D. G. & Donner, K. 2000 In search of the visual pigment template. Vis. Neurosci. 17, 509 –528. (doi:10. 1017/S0952523800174036) Grillner, S., Wallén, P. & Brodin, L. 1991 Neuronal network generating locomotor behaviour in lamprey: circuitry, transmitters, membrane properties, and stimulation. Annu. Rev. Neurosci. 14, 169– 199. (doi:10.1146/ annurev.ne.14.030191.001125) Grogan, E. D. & Lund, R. 2004 The origin and relationships of early Chondrichthyes. In Biology of sharks and their relatives (eds J. C. Carrier, J. A. Musick & M. R. Heithaus), pp. 3–32. Boca Raton, FL: CRC Press. Gruber, L. J. & Cohen, J. L. 1978 Visual system of the elasmobranchs: state of the art. In Sensory biology of sharks skates and rays (eds E. S. Hodgson & R. F. Mathewson). Arlington, VA: Office of Naval Research, Department of the Navy. Phil. Trans. R. Soc. B (2009) Gustafsson, O. S. E., Collin, S. P. & Kröger, R. H. H. 2008 Early evolution of multifocal eye optics for acute colour vision in vertebrates. J. Exp. Biol. 211, 1559– 1564. (doi:10.1242/jeb.016048) Hardisty, M. W. 1982 Lampreys and hagfishes: analysis of cyclostome relationships. In The biology of lampreys, vol. 4B (eds M. W. Hardisty & I. C. Potter), pp. 165 –259. London, UK: Academic Press. Hárosi, F. I. 1994 An analysis of two spectral properties of vertebrate visual pigments. Vis. Res. 34, 1359 –1367. (doi:10.1016/0042-6989(94)90134-1) Hárosi, F. I. & Kleinschmidt, J. 1993 Visual pigments in the sea lamprey, Petromyzon marinus. Vis. Neurosci. 10, 711 –715. (doi:10.1017/S0952523800005411) Hart, N. S. 2001 The visual ecology of avian photoreceptors. Prog. Ret. Eye Res. 20, 675 –703. (doi:10.1016/S13509462(01)00009-X) Hart, N. S. & Vorobyev, M. 2005 Modelling oil droplet absorption spectra and spectral sensitivities of bird cone photoreceptors. J. Comp. Physiol. A 191, 381–392. (doi:10.1007/s00359-004-0595-3) Hart, N. S., Lisney, T. J., Marshall, N. J. & Collin, S. P. 2004 Multiple cone visual pigments and the potential for trichromatic colour vision in two species of elasmobranch. J. Exp. Biol. 207, 4587–4594. (doi:10. 1242/jeb.01314) Hart, N. S., Lisney, T. J. & Collin, S. P. 2006a Visual communication in elasmobranchs. In Communication in fishes (eds F. Ladich, S. P. Collin, P. Moller & B. G. Kapoor), pp. 337 –372. Plymouth: Science Publishers. Hart, N. S., Lisney, T. J. & Collin, S. P. 2006b Cone photoreceptor oil droplet pigmentation is affected by ambient light intensity. J. Exp. Biol. 209, 4776–4787. (doi:10. 1242/jeb.02568) Hart, N. S., Bailes, H. J., Vorobyev, M., Marshall, N. J. & Collin, S. P. 2008 Visual ecology of the Australian lungfish (Neoceratodus forsteri). BMC Ecol. See http:// www.biomedcentral.com/1472-6785/8/21/abstract. Hawryshyn, C. W. 1998 Vision. In The physiology of fishes (ed. D. H. Evans), pp. 345 –374, 2nd edn. Boca Raton, FL: CRC Press. Heinermann, P. H. 1985 Yellow intraocular filters in fishes. J. Exp. Biol. 43, 127–147. Hisatomi, O. & Tokunaga, F. 2002 Molecular evolution of proteins involved in vertebrate phototransduction. J. Comp. Biochem. Physiol. B 133, 509– 522. (doi:10. 1016/S1096-4959(02)00127-6) Hisatomi, O., Iwasa, T., Tokunaga, F. & Yasui, A. 1991 Isolation and characterization of lamprey rhodopsin cDNA. Biochem. Biophys. Res. Comm. 174, 1125 –1132. (doi:10.1016/0006-291X(91)91537-M) Hisatomi, O., Ishikawa, M., Tonosaki, A. & Tokunaga, F. 1997 Characterisation of lamprey rhodopsin by isolation from lamprey retina and expression in mammalian cells. Photochem. Photobiol. 66, 792–795. (doi:10.1111/j.17511097.1997.tb03226.x) Hodgson, E. S. & Mathewson, R. F. 1978 In Sensory biology of sharks, skates and rays. Arlington, VA: Office of Naval Research Department of the Navy. Holmberg, K. 1970 The hagfish retina: fine structure of retinal cells in Myxine glutinosa L., with special reference to receptor and epithelial cells. Zeitschrift für Zellforschung 111, 519 –538. (doi:10.1007/BF00330929) Holmberg, K. 1971 The hagfish retina: electron microscopic study comparing receptor and epithelial cells in the Pacific hagfish, Polistotrema stoutii, with those in the Atlantic hagfish Myxine glutinosa. Zeitschrift für Zellforschung 121, 249 –269. (doi:10.1007/BF00340676) Holmberg, K. & Öhman, P. 1976 Fine structure of retinal synaptic organelles in lamprey and hagfish Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 Review. Vertebrate photoreceptor evolution photoreceptors. Vis. Res. 16, 237 –239. (doi:10.1016/ 0042-6989(76)90105-X) Hubbs, C. L. & Potter, I. C. 1971 Distribution, phylogeny and taxonomy. In The biology of lampreys, vol. 1 (eds M. W. Hardisty & I. C. Potter), pp. 1– 65. London, UK: Academic Press. Hueter, E. R., Mann, D. A., Maruska, K. P., Sisneros, J. A. & Demski, L. S. 2004 Sensory biology of elasmobranchs. In Biology of sharks and their relatives (eds J. C. Carrier, J. A. Musick & M. R. Heithaus), pp. 325–368. London, UK: CRC Press. Hunt, D. M., Davies, W. L., Pan, L., Qin, M., Cowing, J. A., Potter, I. C., Trezise, A. E. O. & Collin, S. P. 2007 The evolution of vertebrate vision: the phototransduction genes of lamprey, Geotria australis. Vision Downunder: eye to brain. Satellite Meeting of the 7th Int. Brain Research Organization (IBRO) Congress of Neuroscience, Cairns, Australia, 19–22 July 2007. Imai, H., Kuwayama, S., Onishi, A., Morizumi, T., Chisaka, O. & Scichida, Y. 2005 Molecular properties of rod and cone visual pigments from purified chicken cone pigments to mouse rhodopsin in situ. Photochem. Photobiol. Sci. 4, 667–674. (doi:10.1039/b416731g) Ishikawa, M., Takao, M., Washioka, H., Tokunaga, F., Watanabe, H. & Tonosaki, A. 1987 Demonstration of rod and cone photoreceptors in the lamprey retina by freeze replication and immunofluorescence. Cell Tissue Res. 249, 241 –246. (doi:10.1007/BF00215506) Jacobs, G. H. & Rowe, M. P. 2004 Evolution of vertebrate colour vision. Clin. Exp. Optom. 87, 206–216. (doi:10. 1111/j.1444-0938.2004.tb05050.x) Kelber, A., Vorobyev, M. & Osorio, D. 2003 Animal colour vision—behavioural tests and physiological concepts. Biol. Rev. 78, 81–118. (doi:10.1017/S1464793102005985) Kemp, A. 1986 The biology of the australian lungfish, Neoceratodus forsteri (Krefft 1870). J. Morphol. Supp. 1, 181 –198. (doi:10.1002/jmor.1051900413) Kemp, N. E. 1999 Integumentary system and teeth. In Sharks, skates and rays: the biology of elasmobranch fishes (ed. W. C. Hamlett), pp. 43–68. Baltimore, MD: The Johns Hopkins University Press. Kemp, A. & Molnar, R. E. 1981 Neoceratodus forsteri from the lower Cretaceous of New South Wales, Australia. J. Paleontol. 55, 211 –217. Lamb, T. D. 2009 Evolution of vertebrate retinal photoreception. Phil. Trans. R. Soc. B 364, 2911–2924. (doi:10. 1098/rstb.2009.0102). Lamb, T. D., Collin, S. P. & Pugh Jr, E. N. 2007 Evolution of the vertebrate eye: opsins, photoreceptors, retina and eye cup. Nature Rev. Neurosci. 8, 960–975. (doi:10.1038/nrn2283) Lamb, T. D., Pugh Jr, E. N. & Collin, S. P. 2008 The origin of the vertebrate eye. Evol. Educ. Outreach 1, 415–426. (doi:10.1007/s12052-008-0091-2) Litherland, L. & Collin, S. P. 2008 Comparative visual function ion elasmobranchs: spatial arrangement and ecological correlates of photoreceptor and ganglion cell distributions. Vis. Neurosci. 25, 549–561. (doi:10.1017/ S0952523808080693) Lythgoe, J. N. 1980 Vision in fishes. In Environmental physiology of fishes (ed. M. A. Ali), pp. 431 –445. New York, NY: Plenum Publishing Corporation. Marshall, N. J., Collin, S. P., Hart, N. S. & Bailes, H. J. In press. Vision in lungfish. In Biology of lungfishes (eds J. M. Jorgensen, J. Joss & T. Wang). NJ, USA: Science Publishers Inc. Maximov, V. V. 2000 Environmental factors which may have led to the appearance of colour vision. Phil. Trans. R. Soc. Lond. B 355, 1239 –1242. (doi:10.1098/rstb.2000.0675) Meyer, A. & Zardoya, R. 2003 Recent advances in the (molecular) phylogeny of vertebrates. Ann. Rev. Ecol. Evol. Phil. Trans. R. Soc. B (2009) S. P. Collin et al. 2939 System. 34, 311 –338. (doi:10.1146/annurev.ecolsys.34. 011802.132351) Meyer-Rochow, B. & Stewart, D. 1996 Review of larval and postlarval eye ultrastructure in the lamprey (Cyclostomata) with special emphasis on Geotria australis (Gray). Micro. Res. Tech. 35, 431 –444. (doi:10. 1002/(SICI)1097-0029(19961215)35:6,431::AIDJEMT3.3.0.CO;2-L) Nag, T. C. & Battacharjee, J. 1995 Retinal ellipsosomes: morphology, development, identification, and comparison with oil droplets. Cell Tissue Res. 279, 633– 637. (doi:10.1007/BF00318176) Nathans, J., Thomas, D. & Hogness, D. S. 1986 Molecular genetics of human colour vision: the genes encoding blue, green, and red pigments. Science 232, 193 –202. (doi:10.1126/science.2937147) Negishi, K., Teranishi, T., Kuo, C.-H. & Miki, N. 1987 Two types of lamprey retina photoreceptors immunoreactive to rod- or cone-specific antibodies. Vis. Res. 27, 1237–1241. (doi:10.1016/0042-6989(87)90199-4) Newth, D. R. & Ross, D. M. 1955 On the reaction to light of Myxine glutinosa L. J. Exp. Biol. 32, 4– 21. Nikol’skii, G. V. 1956 Some data on the period of marine life of the Pacific Lampetra japonica. Zool. Zh. 35, 588– 591. O’Brien, J., Ripps, H. & Al-Ubaldi, M. R. 1997 Molecular cloning of a rod opsin cDNA from the skate retina. Gene 193, 141–150. (doi:10.1016/S03781119(97)00079-6) Öhman, P. 1971 The photoreceptor outer segments of the river lamprey (Lampetra fluviatilis). An electron-fluorescence and light microscopic study. Acta. Zool. Stockh. 52, 287 –297. Öhman, P. 1976 Fine structure of photoreceptors and associated neurons of Lampetra fluviatilis (Cyclostomi). Vis. Res. 16, 659 –662. (doi:10.1016/0042-6989(76)90014-6) Okano, T., Fukada, Y., Artamonov, I. D. & Yoshizawa, T. 1989 Purification of cone visual pigments from chicken retina. Biochemistry 28, 8848–8856. (doi:10.1021/ bi00448a025) Oota, S. & Saitou, N. 1999 Phylogenetic relationship of muscle tissues deduced from superimposition of gene trees. Mol. Biol. Evol. 16, 856–867. Parker, A. R. 1998 Colour in Burgess shale animals and the effect of light on evolution in the Cambrian. Proc. R. Soc. Lond. B 265, 967– 972. (doi:10.1098/rspb.1998.0385) Parry, J. W. L. & Bowmaker, J. K. 2000 Visual pigment in intact goldfish retina using synthetic retinaldehyde isomers. Vis. Res. 40, 2241–2247. (doi:10.1016/S00426989(00)00101-2) Parry, J. W. L., Carleton, K. L., Spady, T., Carboo, A., Hunt, D. M. & Bowmaker, J. K. 2005 Mix and match colour vision: tuning spectral sensitivity by differential opsin gene expression in Lake Malawi cichlids. Curr. Biol. 15, 1734 –1739. (doi:10.1016/j.cub.2005.08. 010) Partridge, J. C. 1989 The visual ecology of avian cone oil droplets. J. Comp. Physiol. A. 165, 415 –426. (doi:10. 1007/BF00619360) Partridge, J. C. 1995 The colour sensitivity and vision of fishes. In Light and life in the sea (eds P. J. Herring, A. K. Campbell, M. Whitfield & L. Maddock), pp. 167 – 184. Cambridge, UK: Cambridge University Press. Pisani, D., Mohun, S. M., Harris, S. R., McInerney, J. O. & Wilkinson, M. 2006 Molecular evidence for dim-light vision in the last common ancestor of the vertebrates. Curr. Biol. 16, R318–R319. (doi:10.1016/j.cub.2006.03. 090) Potter, I. C. 1980 The Petromyzoniformes with particular reference to paired species. Can. J. Fish. Aquat. Sci. 37, 1595–1615. (doi:10.1139/f80-207) Downloaded from http://rstb.royalsocietypublishing.org/ on May 12, 2017 2940 S. P. Collin et al. Review. Vertebrate photoreceptor evolution Potter, I. C. & Hilliard, R. W. 1987 A proposal for the functional and phylogenetic significance of differences in the dentition of lampreys (Agnatha: Petromyzontiformes). J. Zool. Lond. 212, 713 –737. (doi:10.1111/j.1469-7998. 1987.tb05966.x) Potter, I. C. & Strahan, R. 1968 The taxonomy of the lampreys Geotria and Mordacia and their distribution in Australia. Proc. Linn. Soc. Lond 179, 229– 240. Potter, I. C., Prince, P. A. & Croxall, J. P. 1978 Data on the adult marine and migratory phases in the lifecycle of the southern hemisphere lamprey, Geotria australis Gray. Environ Biol. Fish. 4, 65–69 (doi:10.1007/BF00005929) Pratt Jr, H. L. & Carrier, J. C. 2001 A review of elasmobranch reproductive behaviour with a case study on the nurse shark, Ginglymostoma cirratum. Environ. Biol. Fish. 60, 157 –188. (doi:10.1023/A:1007656126281) Renaud, C. B. 1997 Conservation status of Northern Hemisphere lampreys (Petromyzontidae). J. Appl. Ichthyol. 13, 143 –148. (doi:10.1111/j.1439-0426.1997.tb00114.x) Robinson, S. R. 1994 Early vertebrate colour vision. Nature 367, 121. (doi:10.1038/367121a0) Rubinson, K. 1990 The developing visual system and metamorphosis in the lamprey. J. Neurobiol. 21, 1123– 1135. (doi:10.1002/neu.480210715) Rubinson, K. & Cain, H. 1989 Neural differentiation in the retina of the larval sea lamprey (Petromyzon marinus L). Vis. Neurosci. 3, 241–248. (doi:10.1017/S09525238 00009998) Schultze, M. 1866 Zur Anatomie und Physiologie der Retina. Arch. f. mikr. Anat. 2, 175– 286. (doi:10.1007/ BF02962033) Schultze, M. 1867 Über Stäbchen und Zapfen der Retina. Arch. mikroskop. Anat. 3, 215–247. (doi:10.1007/ BF02960456) Shand, S. et al. 2008 The influence of ontogeny and light environment on the expression of visual pigment opsins in the retina of the black bream, Acanthopagrus butcheri. J. Exp. Biol. 211, 1495–1503. (doi:10.1242/jeb.012047) Shu, D. G. et al. 2003 Head and backbone of the early Cambrian vertebrate. Haikouichthys. Nature 421, 526 –529. (doi:10.1038/nature01264) Siebeck, U. E. & Marshall, N. J. 2001 Ocular media transmission of coral reef fish—can coral reef fish see ultraviolet light? Vis. Res. 41, 133–149. (doi:10.1016/ S0042-6989(00)00240-6) Siebeck, U. E., Collin, S. P., Ghoddussi, M. & Marshall, N. J. 2003 Occlusable corneas in toadfishes: light transmission, movement and ultrastructure of pigment during light and dark adaptation. J. Exp. Biol. 206, 2177–2190. (doi:10.1242/jeb.00401) Sillman, A. J., Sorsky, M. E. & Loew, E. R. 1995 The visual pigments of wild white sturgeon (Acipenser transmontanus). Can. J. Zool. 73, 805 –809. (doi:10.1139/z95-093) Steven, D. M. 1955 Experiments on the light sense of the hag, Myxine glutinosa L. J. Exp. Biol. 32, 22– 38. Takezaki, N., Figueroa, F., Zaleska-Rutczynska, Z. & Takahata, N. 2004 The phylogenetic relationship of tetrapod, coelacanth, and lungfish revealed by the sequences of 44 nuclear genes. Mol. Biol. Evol. 21, 1512– 1524. (doi:10.1093/molbev/msh150) Temple, S. E., Veldhoen, K. M., Phelan, J. T., Veldhoen, N. J. & Hawryshyn, C. W. 2008 Ontogenetic changes in opsin gene expression in coho salmon (Oncorhynchus kisutch, Walbaum). J. Exp. Biol. 211, 3879–3888. (doi:10.1242/jeb.020289) Theiss, S. M., Lisney, T. J., Collin, S. P. & Hart, N. S. 2007 Colour vision and visual ecology of the blue-spotted maskray, Dasyatis kuhlii Müller & Henle, 1814. J. Comp. Physiol. A 193, 67–79. (doi:10.1007/s00359006-0171-0) Phil. Trans. R. Soc. B (2009) Tonosaki, A., Washioka, H., Hara, M., Ishikawa, M. & Watanabe, H. 1989 Photoreceptor disc membranes of Lampetra japonica. Neurosci. Res. 6, 340 –349. (doi:10. 1016/0168-0102(89)90026-6) Trezise, A. E. O. & Collin, S. P. 2005 Opsins: evolution in waiting. Curr. Biol. 15, R794–R796. (doi:10.1016/j.cub. 2005.09.025) Ullén, F., Deliagina, T. G., Orlovsky, G. N. & Grillner, S. 1995 Spatial orientation in the lamprey. II. Visual influence on orientation during locomotion and in the attached state. J. Exp. Biol. 198, 675 –681. Vigh-Teichmann, I., Vigh, B., Olsson, R. & van Veen, T. 1984 Opsin-immunoreactive outer segments of photoreceptors in the eye and in the lumen of the optic nerve of the hagfish, Myxine glutinosa. Cell Tissue Res. 238, 515 –522. (doi:10.1007/BF00219867) Vorobyev, M. 1997 Costs and benefits of increasing the dimensionality of colour vision system. In Biophysics of photoreception: molecular and phototransductive events (ed. C. Taddei-Ferretti), pp. 280–289. Singapore: World Scientific. Vorobyev, M. 2003 Coloured oil droplets enhance colour discrimination. Proc. R. Soc. Lond. B. 270, 1255– 1261. (doi:10.1098/rspb.2003.2381) Walker, K. R. & Laporte, L. F. 1970 Congruent fossil communities from Ordovician and Devonian carbonates of New York. J. Paleontol. 44, 928–944. Walls, G. L. 1942 In The vertebrate eye and its adaptive radiation, Bloomfield Hills, MI: Cranbrook Press. Walls, G. L. & Judd, H. D. 1933 The intraocular colour filters of vertebrates. Br. J. Ophthalmol. 17, 641 –725. (doi:10.1136/bjo.17.11.641) Whitmore, A. V. & Bowmaker, J. K. 1989 Seasonal variation in cone sensitivity and short-wave-absorbing visual pigments in the rudd Scardinius erythrophthalmus. J. Comp. Physiol. A 166, 103 –115. (doi:10.1007/BF00190215) Xian-Guang, H., Aldridge, R. J., Siveter, D. J. & XiangHong, F. 2002 New evidence on the anatomy and phylogeny of the earliest vertebrates. Proc. R. Soc. Lond. B 269, 1865–1869. (doi:10.1098/rspb.2002.2104) Yokoyama, S. 1997 Molecular genetic basis of adaptive selection: examples from colour vision in vertebrates. Annu. Rev. Genet. 31, 315–336. (doi:10.1146/annurev. genet.31.1.315) Yokoyama, S. & Yokoyama, R. 1990 Molecular evolution of visual pigment genes and other G-protein coupled receptor genes. In Population biology of genes and molecules. (eds N. Takahata & J. F. Crow), pp. 307 –322. Tokyo, Japan: Baifukan. Yokoyama, S. & Yokoyama, R. 1996 Adaptive evolution of photoreceptors and visual pigments in vertebrates. Annu. Rev. Ecol. Syst. 27, 543–567. (doi:10.1146/ annurev.ecolsys.27.1.543) Yoshida, M. & Tonosaki, A. 1994 Anti-lamprey (Lampetra japonica) retinal antibodies which distinguish between short and long photoreceptor cells. Comp. Biochem. Physiol. B 107, 183 –188. (doi:10.1016/0305-0491(94) 90038-8) Young, G. C. 2008 Number and arrangement of extraocular muscles in primitive gnathostomes: evidence from extinct placoderm fishes. Biol. Lett. 4, 110– 114. (doi:10.1098/ rsbl.2007.0545) Young, R. W. & Martin, G. R. 1984 Optics of retinal oil droplets: a model of light collection and polarization detection in the avian retina. Vis. Res. 24, 129 –137. (doi:10.1016/0042-6989(84)90098-1) Zhang, H. & Yokoyama, S. 1997 Molecular evolution of the rhodopsin gene of marine lamprey, Petromyzon marinus. Gene 191, 1 –6. (doi:10.1016/S0378-1119(96) 00864-5)