Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Anatomical Demonstration of Orientation Columns in
Macaque Monkey
DAVID H. HUBEL, TORSTEN N. WIESEL A N D MICHAEL P. STRYKER
Department ofNeurobiology, Harvard Medical School, Boston, Massachusetts 02115
ABSTRACT
In the macaque monkey striate (primary visual) cortex, the
grouping of cells into ocular dominance and orientation columns leads to the prediction of highly specific spatial patterns of cellular activity in response to stimulation by lines through one or both eyes. In t h e present paper these patterns have
been examined by the 2-deoxyglucose autoradiographic method developed by
Sokoloff and his group (Kennedy e t al, '76). An anesthetized monkey was given
a n injection of I4C 2-deoxyglucose and then visually stimulated for 45 minutes
with a large array of moving vertical stripes, with both eyes open. The 14Cautoradiographs of striate cortex showed vertical bands of label extending through the
full cortical thickness. Layer I was a t most only lightly labelled, and layers IV b
and VI were t h e most dense. Layer IV c (the site of terminations of most geniculate afferents) was labelled uniformly along its length, a s expected from the lack
of orientation specificity of units recorded in t h a t layer. In the other layers the
pattern seen in tangential sections was complex, consisting of swirling stripes
with many bifurcations and blind endings, but with occasional more regular regions where t h e stripes were roughly parallel. Interstripe distance was rather
constant, a t 570 pm. Ocular dominance columns were examined in this same
monkey, in t h e same region, by injecting one eye with H-proline two weeks
before t h e deoxyglucose experiment, and preparing a second set of autoradiographs of the sections after prolonged washing to remove the 14C-deoxyglucose.
As seen in tangential sections through layer IV c, these columns had the usual
stripe-like form, with a period of 770 p m , but were simpler in their pattern than
the orientation stripes, with fewer bifurcations and less swirling. A comparison
of the two sets of columns in t h e same area showed many intersections, but no
strict or consistent relationships: angles of intersection showed a distribution
t h a t was not obviously different from t h a t expected for any two randomly
superimposed sets of lines.
Another monkey was stimulated with vertical stripes, but with only one eye
open. Deoxyglucose autoradiographs of tangential sections showed regular uniform rows of label in layer IV c, with all t h e characteristic features of eye dominance columns. In the layers above and below IV c the rows in tangential view
were broken up into regularly spaced patches of label, presumably representing
aggregations of cells responsive to vertically oriented stimuli. The patches
showed no consistent alignment across the ocular dominance rows, and indeed no
such tendency would be expected, considering t h e complexity of the orientation
columns. This pattern of labelling is again predicted from and confirms t h e previous physiological studies.
Two functions of t h e primary visual cortex
of higher mammals have now been established
(Hubel and Wiesel, '62, '68). The information
from t h e lateral geniculate body is reorJ. COMP. NEUR. (1978)177: 361-380.
ganized in such a way t h a t cells after t h e initial stage respond best to specifically oriented
straight-line segments, rather t h a n to spots of
light; and t h e cortex is for all practical pur-
361
362
D. H. HUBEL. T. N. WIESEL A N D M. P. STRYKER
poses the first site in the retino-geniculo-cortical pathway at which signals from the two
eyes converge upon single cells.
Related to and subserving these two functions are two independent sets of vertical subdivisions, the orientation columns and t h e
ocular dominance columns (Hubel and Wiesel,
'62, '63, '68, '74a). When a micro-electrode
penetrates through the cortex in a direction
perpendicular to the surface, recording from
many cells in sequence, t h e optimal orientation of a stationary or moving short-line stimulus tends to be virtually constant, and t h e
same eye remains dominant, though many
cells are influenced (usually unequally) by the
two eyes. In a tangential or oblique penetration the optimal orientation and the ocular
dominance both vary. Orientation changes
systematically in a clockwise or counterclockwise direction, in small steps or perhaps
continuously, at rates such t h a t 180' is
covered in about 600 p m or more. Reversals in
the direction of rotation occur irregularly,
roughly one or two to a centimeter, and occasionally the sequence may be broken by a n
abrupt shift of up to go", which we term a fracture. Ocular dominance meanwhile shifts
back and forth, apparently quite independently of orientation: first one eye dominates, then
the two become roughly equal, and finally the
other dominates, a complete cycle occurring
roughly every 800 p m . Thus while the individual subdivisions in the two systems a r e very
different in size (25-50 p m for t h e orientation
columns a s opposed to 400 p m for the ocular
dominance) a complete set of either type, leftplus-right eye or a cycle of 180" of orientation,
occupies about the same distance on t h e cortex, 0.5-1 mm. One complete set of columns, of
either type, has been termed a hypercolumn
(Hubel and Wiesel, '74b). This is illustrated in
figure 1.
This description applies to all layers from I1
to VI, except for IV c , the layer in which the
bulk of t h e geniculo-cortical inputs terminate.
In IV c the cells show no hint of orientation selectivity, but seem to have circularly symmetric center-surround receptive fields resembling the fields of geniculate cells. Moreover here the cells a r e virtually all monocular,
so t h a t on proceeding tangenitially along IV c,
crossing from one ocular dominance column to
the next, the electrode passes from a region in
which the cells respond only to one eye to a region monopolized by t h e other. There is thus a
strict alternation of eyes, with abrupt transi-
tions a t the column borders. A cell above or
below IV c is presumed to receive convergent
input, relayed directly or over several synapses, from several of these monocular regions
in IV c, and is consequently likely to be binocularly influenced, but dominated by the eye
t h a t corresponds to the region in I V c lying
below or above along the same radial line, i.e.,
in the same column.
The cortex is thus subdivided by two independent sets of partitions t h a t a r e perpendicular to t h e surface and the layers. These
subdivisions are not visible by ordinary histological staining methods. Several lines of evidence nevertheless suggest t h a t both systems
of grouping have the form of parallel vertically disposed sheets. For the eye dominance system the sheet-like geometry has been made
evident by three independent anatomical
methods: the Nauta/Fink-Heimer/Wiitanen
stain (Hubel and Wiesel, '721, transneuronal
autoradiography after eye injection (Wiesel e t
al., '74; Hubel et al., '771, and a reduced silver
stain (LeVay et al., '75). I n tangential sections
through layer IV c the ocular dominance columns appear as a set of parallel stripes.
Though on t h e whole t h e stripes are remarkably regular, in places they show bifurcations
and blind endings, and often form loops and
whorls. The evidence t h a t the orientation columns a r e arranged in parallel slabs stems
from physiological recordings, and is deduced
from reconstructions of multiple parallel penetrations (Hubel and Wiesel, '63, '74a) and
from t h e fact t h a t the orientation shifts in
any single oblique or tangential penetration
are small and regular. While so far there has
been no anatomical method for demonstrating
t h e orientation sheets, t h e recordings, and in
particular t h e reversals and fractures, suggested t h a t they might swirl and branch extensively.
Figure 1shows in schematic form a model of
t h e monkey striate cortex (Hubel and Wiesel,
'77). The diagram represents two orientation
hypercolumns a n d two ocular dominance
hypercolumns. I t should be kept in mind t h a t
t h e orientation columns can be far from flat,
t h a t t h e choice of vertical orientation to begin
a n d e n d t h e orientation hypercolumn is
arbitrary, t h a t nothing is known about t h e relationship between the two types of columns
- the decision to draw them as orthogonal is
again arbitrary - and t h a t whether or not the
orientation columns are discrete is still unsettled (Albus, '75; Hubel and Wiesel, '74a). I n
ORIENTATION COLUMNS IN MACAQUE MONKEY
this figure t h e orientation slabs represent regions of cortex over which optimal orientation
is assumed to be constant, and the shifts in
orientation from one slab to t h e next are represented a s 10". I t is worth stressing, however,
t h a t t h e region of cortex activated by a line in
a particular orientation would be much wider
than these slabs, because for most cells the
range of orientations over which responses are
evoked is several times greater than 10".
Moreover, such regions of activation would
not be precisely defined, since for each cell the
responses vary with orientation according to a
tuning curve, and since these tuning curves
themselves vary in width from cell to cell.
(See, for example, Schiller e t al., '76).
Given these columnar groupings of cells
according to response preferences, i t is possible to predict t h e cortical activity patterns
produced by various visual stimuli. Figure 2A
represents the activity pattern produced in
the upper or lower layers Le., 11-111, IV b, V,
or VI) by a set of vertical stimulus stripes
covering a large part of t h e visual field and
viewed by both eyes. Each shaded strip in the
figure is of course maximally activated along
a narrow center line: as discussed above the
width of the shaded strips will depend on tuning curve widths, and their borders will not be
sharp but will shade off just as tuning curves
do.
Figure 2B shows t h e pattern predicted, for
the same layers, when only one eye views the
vertical stripes. And finally 2C shows t h e pattern for one stimulus orientation (e.g., vertical) and one eye in layer IV c; here t h e dominance columns corresponding to the stimulated eye a r e activated along their entire
lengths, since cells in IV c respond equally to
all line orientations. Needless to say, a fourth
diagram to illustrate a combination of vertical
lines, for both eyes, in layer I V c , would
require shading throughout.
Until now these patterns had been inferred
entirely from physiological recordings, except
for the case of dominance stripes in layer IV c
(fig. 2C), which had been seen anatomically by
the methods already listed. Recently, however, a more direct approach has become possible. Over the past few years Sokoloff and his
group have developed a method by which recently active regions of nervous tissue can be
differentially marked (Sokoloff, '75; Sokoloff
et al., '77). The technique rests on t h e fact
t h a t nerve cells use glucose as their main
energy source and on t h e assumption t h a t
363
heightened activity leads to a n increase in
glucose consumption. An animal is injected
intravenously with a single dose of "C 2-deoxyglucose, which is taken up by nerve cells
and is phosphorylated as if i t were normal
glucose, to t h e 2-deoxyglucose-6-phosphate,
but not further metabolized. The cell membrane is relatively impermeable to this compound, so t h a t the label is effectively trapped
inside the cell in concentrations proportional
to the integrated uptake of glucose. If the
brain is quickly frozen and sectioned in a
frozen state to prevent diffusion of the watersoluble label, then regions with increased
metabolic rates during the period in which t h e
deoxyglucose was available will be visible on
autoradiographs.
As one demonstration of the potential of
this tool in neurobiology, Sokoloffs group has
used i t to demonstrate t h e ocular dominance
columns in macaque monkeys by stimulating
one eye only (Kennedy e t al., '76). The method
has been used in several other systems in the
r a t by Sharp and his colleagues (Sharp e t al.,
'75, Sharp, '76b), and by Durham and Woolsey
('77) to demonstrate whisker barrels in mouse
cortex. We hoped t h a t by stimulating both
eyes with lines in one orientation we might
reveal the corresponding subset of orientation
columns and thus obtain anatomical evidence
for t h e columnar organization. It would then
a t last be possible to see the 3-dimensional
arrangement of t h e orientation columns, and
compare this with the arrangement of ocular
dominance columns obtained by amino acid
eye injections in the same animal.
METHOD
The present study is based on three experiments done in macaque monkeys about six
months old. I n each animal we followed in
most respects the procedure described by
Sokoloff (Kennedy et al., '761, to whom we are
indebted for first-hand instruction in the
method. The animal, lightly anesthetized with
thiopental and paralyzed with a continuous
intravenous infusion of curare and gallamine,
was stimulated (as described below) for 45
minutes following a rapid intravenous injection of I4C 2-deoxyglucose (New England Nuclear, 150 pCi/kg in 1.5 ml/kg of 0.9%saline).
At 5-minute intervals during t h e stimulation
period samples of blood were drawn and 14C
levels determined to be sure t h a t at t h e end
they had fallen to a few percent of the initial
level. The animal was then given intravenous-
364
D. H. HUBEL. T. N. WIESEL AND M. P. STRYKER
\
\
B
h h Q$
@
4
R
/
/
\
L
R
f
L
/
\
\
\
\
\
Fig. 1 Idealized model of the monkey striate cortex, showing two orientation hypercolumns each covering a
full 180",and two ocular dominance hypercolumns. Here t h e columnar walls are represented as flat, and the two
sets are shown intersecting a t right angles, but the present paper indicates that neither set of walls is flat, and
that the intersections are probably random. R, right eye; L, left eye. (From Hubel and Wiesel, '77: fig. 27)
ly 50 mg/kg of thiopental followed by a lethal
dose of KC1 (4 ml saturated solution) and decapitated. The head was immediately cleaned
of skin, frozen by gradual immersion in Freon22 at -125°C over a period of four minutes,
a n d stored at -80°C. The skull was later removed and small blocks of brain sectioned at
20 g m in a cryostat at -22" to -26°C. The
sections were immediately dried on a cover
slip heated to 98°C and pressed against X-ray
film for two to three weeks, after which t h e
film was developed. Every fifth section was
mounted and stained for Nissl substance
(cresyl violet); in addition some of the sections
used for autoradiography were later stained
for Nissl substance.
RESULTS
Ocular dominance columns
I n the first monkey we wished to examine
t h e ocular dominance columns. The left eye
was occluded with a n opaque cover and the
right eye stimulated with a large brightly
illuminated screen containing white stripes
' I a - ' h 0 wide and spaced 1 s - 2 "apart, on a black
Fig. 2 Drawings to indicate the patterns of cortical activity expected from three different visual stimuli. A patch
of monkey striate cortex several millimeters in length and
width is viewed face-on. The horizontal lines represent
boundaries between orientation columns, the vertical lines,
boundaries between ocular dominance columns. H and V
stand for horizontal and vertical stimulus orientations; L
and R, t h e left and right eyes. As in figure 1, the column
boundaries are very schematic. Shaded areas indicate the
regions in which cells are expected to be activated. (Little is
known about the response properties of cells in layers I or
IV a,)
A The pattern expected in layers 11,111,IV b, V and
VI when both eyes are stimulated by vertical lines.
B Activity pattern corresponding to vertical lines
stimulating one eye only, in 11, 111, IV b, V, and VI.
C Activity in layer IV c, in response to vertical-line
stimuli to the left eye only.
ORIENTATION COLUMNS IN MACAQUE MONKEY
A
B
C
Vertical lines
Both eyes
Layers II, Ill, IVb,V. VI
Vertical lines
Left eye
Layers II, 111, I V b . V , V I
Vertical lines
Left eye
Layer IV c
365
366
D. H. HUBEL, T. N. WIESEL AND M. P. STRYKER
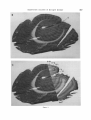
Fig. 3A, B Deoxyglucose autoradiographs of coronal sections through the right occipital pole of monkey No.
1 to show ocular dominance columns. This animal was stimulated with stripes in all orientations through the
right eye only. A is 5 mm in front of the occipital pole, B is 0.5mm in front of A. Down and right in the diagram is
superior; up and right is medial. In the upper part of the figure (medial aspect of occipital lobe) t h e columns are
seen in transverse section; two folds below this in the figure (in t h e superior bank of t h e calcarine fissure, labelled S) they a r e c u t very obliquely.
C The left portion is from the same section a s B, the right is a Nissl-stained section 60 pm distant,
matched to show t h a t the highest density of label is in layers IV b and VI.
background. The screen, which covered most
of t h e visual field, was held 1 m away (the distance at which the eye had been focused) and
moved in a direction perpendicular to the
stripe orientation a t 2-5"/sec. Meanwhile t h e
screen was rotated slowly and t h e direction of
movement was changed so as to stimulate in
all orientations a t least once every minute.
Autoradiographs from coronal sections
through two regions close to the right occipital pole of this monkey are shown in
figures 3A and B. B is taken a t a level 0.5 mm
anterior to A. Bands representing ocular
dominance columns cut in a plane roughly
perpendicular to the surface a r e best seen on
t h e medial surface (uppermost, in the figures),
while more oblique, almost tangential sections show the stripe-like pattern in the superior bank ( S ) of the calcarine fissure. The columns extend from almost the surface to t h e
white matter, and a r e most densely labelled at
two levels, one about midway down and one in
the deepest part of the cortex. When a n adjacent Nissl-stained section is matched to the
autoradiograph of B, t h e two densely labelled
levels can be seen to correspond to layers IV b
(the line of Gennari) and VI. In layer IV b
there is some faint higher-than-background
labelling between columns; this is not nearly
so dense as the label in the columns at t h a t
level, but i t does stand out against the otherwise relatively label-free gaps between columns.
In these sections layer I was very thin, and
we were not convinced t h a t columns had been
labelled there.
The distance between bands averaged 760
p m , giving 380 p m for t h e columnar width, a
figure close to t h a t obtained with other methods.
These sections closely resemble those produced in similar experiments by Kennedy e t
ORIENTATION COLUMNS IN MACAQUE MONKEY
Figure 3
367
368
D. H. HUBEL. T. N. WIESEL AND M. P. STRYKER
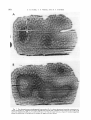
Fig. 4 Deoxyglucose autoradiograph of a section through left area 17 (occipital operculum),in monkey No. 2.
Section passes perpendicular t o the surface. cutting orientation columns transversely. The stimulus consisted
of moving vertical stripes; both eyes were open. Note the continuous label in layer IV c, about half way down. In
t he columns there is a particularly high density of label in layer IV b. a n d in layer VI j u s t above t h e white
matter
al. ('76) (see also Sokoloff 1'751). In some of
their experiments one eye was enucleated
rather than occluded, and during the stimulation t h e i r a n i m a l s were a l e r t r a t h e r
t h a n anesthetized. Previous work (Hubel,
'59; Wurtz, '69) has similarly indicated t h a t
light barbiturate anesthesia does not seriously interfere with specific responses to visual
stimuli in striate cortex. Any general lessening of background neuronal activity produced
by the anesthetic may indeed be a n advantage
for these kinds of studies.
Orientation columns
In the second monkey our main object was
to reveal the orientation columns in area 17
and compare them with the eye dominance
columns. To label t h e dominance columns in
the same monkey we used the method of
transneuronal autoradiography (Wiesel et al.,
'74), injecting 2.0 mCi of tritiated proline into
the vitreous of the right eye two weeks before
the final experiment. At the time of the experiment the animal was anesthetized and set
up for physiological recording from the striate
cortex. Paralysis was induced with a n intravenous infusion of gallamine and curare. The
eyes were both kept open and focused a t 1 m a s
before, and were aligned with a variable prism
so t h a t the projections of t h e foveas, and also
the receptive fields of binocular cells a s
mapped on the screen, were precisely superimposed. I t was necessary to do this to be sure
t h a t the eyes would work in synergy for the
cells in area 1 7 (Hubel and Wiesel, '70) rather
than compete, as might happen if they were
out of alignment. The animal was injected as
before with "C 2-deoxyglucose and stimulated
with the moving stripes for 45 minutes, but
this time both eyes were open and the stripes
were oriented vertically throughout. The recording allowed us to be sure t h a t the animal
was in good condition and t h a t our stimuli
were actually evoking responses in cortical
cells.
An autoradiograph of this monkey's left occipital lobe, from a section perpendicular to
t h e exposed outer part of area 17, is shown in
figure 4.At first glance t h e result might seem
similar to t h a t just described for ocular dominance columns: the periodicity of t h e labelled
regions is not too different and their width in
places approaches t h a t of dominance columns.
One feature, nevertheless, is quite different namely the continuous and dense labelling of
layer I V c , which appears as a conspicuous
horizontal band placed about half-way down
in the cortex, and occupies about one-fifth of
t h e thickness. This is, of course, just what is
expected from the lack of orientation specificity of the units that are recorded in layer
IV c. Each column in addition shows two re-
ORIENTATION COLUMNS IN MACAQUE MONKEY
369
Fig. 5 Relationship between cortical layering as seen in Nissl stain, and laminar differences in labelling. Deoxyglucose autoradiograph of tangential section through the right striate cortex of monkey No. 2. The section
just grazes layer V and cuts the more superficial layers very obliquely. To the right is a neighboring Nisslstained section to identify the layers. The main features are the high denaity of label in IV b, the continuous la.
be1 in 1V c, and the paucity of label in layer I. (Pattern of orientation columns in this section is obscured by microtome-knife artifacts.)
gions of especially high density, one a t a level
just above IV c, the other in the very depths,
presumably in layer VI.
The relationship between the cortical layers
and these differences in density and labelling
pattern is best seen in oblique or tangential
sections, where the layers are more spread out.
In a n example taken from the right occipital
370
D. H . HUBEL, T. N. WIESEL AND M. P. STRYKER
Fig. 6 Deoxyglucose autoradiograph of a tangential section through the left striate cortex of monkey No. 2.
Same region as in figure 7. To the left, the same section as was used to make the autoradiograph has been stained
for Nissl substance and matched, to identify the layers. Note the high density of label in layers I V b and VI. and
the confluence of label in IV c
lobe, shown in figure 5, neighboring autoradiographs and Nissl-stained sections were
cut and spliced. The sections are tangential to
layer V, which appears as an oval, surrounded
in turn by IV c, IV b, 11-111-IVa (these three
subdivisions cannot be sharply distinguished)
and I. The face-on pattern of the labelled regions is not particularly well seen in this section, which was chosen because the layering is
well defined. The columnar pattern seems to
extend up so as to include layer I, though the
labelling is not dense enough to make this absolutely certain. What this figure shows is (1)
the continuous label in IVc, both in its
deepest, most densely cell-packed part and in
its upper, more sparsely populated part; (2)
the clear correspondence between the densely
labelled portions of the columns and layer IV b
(roughly equivalent to the Gennari line), and
(3) the relatively lower density of label in II111.
An autoradiograph of a deeper tangential
section through the opposite (left) occipital
lobe is shown in figure 6. As in figure 5, it is
spliced to a Nissl section, in this case a cresylviolet stain of the section from which the
autoradiograph itself was made. Again one
can see the high density of label in IV b and
the confluence of label in IV c; in addition the
figure shows a high density of label in layer
VI, and a relatively weak labelling in V.
Figure 7 shows autoradiographs from tangential sections a t four different levels
through this region - D is the same as the
right-hand portion of figure 6. These sections
were chosen because the pattern formed by
the labelled columns is reasonably clear. Seen
face-on, they form a complex network of interconnected bands. In places the bands run parallel for short distances, for example in part
of the upper right quadrant in sections A and
C, but more often they branch or end blindly,
or form swirls or irregular rings. The spacing, nevertheless, is remarkably regular, as
ORIENTATION COLUMNS IN MACAQUE MONKEY
though there were strong constraints on the
frequency with which a given orientation recurs.
By making prints of these four sections on
film and superimposing them it was easy to
show t h a t t h e patterns closely coincide. Like
the transverse sections of figure 4, this confirms t h e physiological observation that the
columns are vertical and extend through the
full cortical thickness (at least through layers
11-VI), interrupted only by the uniformly labelled layer IV c.
The overall pattern was reconstructed from
a number of sections parallel to those of figure
7 by cutting out from each the part containing
layer VI and matching them. This is shown in
figure 8A.
Comparison of orientation and ocular
dominance column patterns
From t h e block whose sections a r e shown in
figures 6 and 7 every third section was set
aside to examine t h e ocular dominance columns, using t h e transneuronal 3H label transported from t h e eye t h a t had been injected.
The sections were fixed, washed i n water for
four hours to remove t h e I4C label, dehydrated
and defatted, dipped in photographic emulsion, exposed in darkness for ten weeks, and
then developed. When viewed in dark-field
illumination t h e sections showed banding in
layer IV c typical of ocular dominance columns. The montage of figure 8B was prepared
by cutting out layer IV c from several sections
and assembling them. The stripes form a
rather irregular pattern, as is usual for regions close to t h e representation of t h e
horizontal meridian (as this was). On the
other hand t h e pattern is simpler than t h a t of
the orientation columns in the same region
and certainly t h e two a r e very different in
their details. I n fact, from inspection of t h e
two, separately, there is no hint of any relationship between them. I n figure 9 we superimposed tracings of the two sets of columns of
figure 8, with the dominance columns drawn
in thin lines and the orientation columns in
thick. There is in places some suggestion t h a t
the two patterns may be orthogonal; to test
this we measured the angles a t each of t h e 154
points of intersection and compared the resulting distribution of angles with that expected if t h e two sets were randomly related
(a sine function). From t h e result one can only
say t h a t in this region there was no clear rela-
371
tionship between the two, and in particular no
marked tendency for them to run parallel or to
be orthogonal. We do not consider this question settled, however, and plan to examine
more macaque brains. The widths of the two
sets of hypercolumns were determined by
tracing the two sets of boundaries, in figures
8A and B, and for each set selecting a number
of small, regular regions, determining the
areas of several lengths of columns, and dividing by the lengths. This gave 770 p m for ocular dominance hypercolumns, divided equally
(to within 10%) between t h e two eyes. This is
almost identical to t h e figure obtained from
the first monkey, and is similar to that determined previously from Nauta, reduced silver,
and eye-injection studies (Hubel et al., '77).
For t h e orientation hypercolumns (fig. 8A) the
hypercolumn width, obtained by the same
method, was 570 p m . This may be compared to
a minimum width of 450 p m obtained in physiological recordings from a maximum slope of
about 400"/mm in graphs of orientation vs.
electrode track distance (Hubel and Wiesel,
'74a). From these rough figures we conclude
t h a t t h e orientation and ocular dominance
hypercolumns are of t h e same order of magnitude, with the orientation hypercolumns
slightly smaller.
Ocular dominance plus orientation
In a third monkey we combined the procedures used in the first two deoxyglucose animals by stimulating the left eye only, with
vertical stripes. The expected distribution of
cells activated by such a stimulus has already
been shown schematically in figure 2B, for the
upper and lower layers and, in figure 2C, for
layer IV c. The results strongly supported this
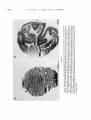
expectation. Figure 10A shows a section running tangential to the right occipital convexity, passing through layers I1 and 111. The label is clearly i n patches, which tend to line up
in rows t h a t run, in t h e figure, from lower left
to upper right.
A much deeper section, B, parallel to A, cuts
t h e calcarine fissure a t right angles over
much of i t s stem, and again shows how regular
and vertical t h e columns are. The patches are
shown almost in tangential section just above
and below the mouth of t h e fissure, and here
their arrangement in rows is even more evident.
Figure 11 compares tangential sections
from t h e left hemisphere a t various depths, to
372
D H HUBEL. T N W I E S E L A N D M P STRYKER
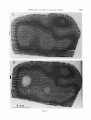
Fig. 7 Four deoxyglucose autoradiographs from monkey No. 2, t o show t h e pattern formed by orientation columns viewed face-on. Tangential sections through left occipital lobe, exposed surface. Sections A and B and B
and C a r e separated by 250 p m , C and D by 200 p m . D is from t h e same section as figure 6. Careful inspection
shows t h e similarity of the pattern of columns in upper a n d lower layers.
ORIENTATION COLUMNS I N MACAQUE MONKEY
Figure 7
373
374
D. H. HUBEL. T. N. WIESEL AND M. P. STRYKER
Fig. 8A Reconstruction of orientation columns from monkey No. 2, left occipital lobe, in t h e same series a s
t h a t of figure 7, made by cutting and mounting the parts of each section passing through layer VI.
B Reconstruction of ocular dominance columns in t h e same region a s A, made from autoradiographs of
H-proline sections following injection of t h e right eye; dark.field photographs, Pa r t s of each autoradiograph
passing through layer IV c were c ut a nd mounted.
375
ORIENTATION COLUMNS IN MACAQUE MONKEY
5 mm
Fig. 9 The vertical-orientation columns, from figure 8A, are traced as thick lines, the left-eye ocular dominance columns from figure 8B, a s thin lines. The average widths of the hypercolumns are 770 pm for the ocular
dominance, 570 wm for the orientation. The two sets are certainly not parallel, but neither are they strictly
orthogonal,
test the prediction of figure 2C, that an oriented stimulus to one eye would give confluent ocular-dominance stripes in IV c. The
sections, three of which are illustrated, were
taken just behind the lunate sulcus. Each section was first used for autoradiography and
then stained with cresyl violet to identify the
layers with certainty: a t each level the cresyl
violet stains are shown in the lower half of the
figure (posterior, in each of the 6 photographs,
is up).
The lunate sulcus (L) runs just in front of
and parallel to the 17-18 border, and from a
previous study (LeVay a t al., '75) it is known
that the ocular dominance stripes are at right
angles to this boundary, running roughly in an
anteroposterior direction. A t each level in
figure 11 one can see the parallel stripes of label, corresponding to the left, stimulated eye,
all stopping short right a t the 17-18 border. In
the superficial (11, 111, IV a) and deep (V, VI)
layers, seen best in A and C, the stripes are
broken up into patches, but in both A and B, in
just the regions that pass through IV c, the
patches coalesce to form continuous stripes, as
predicted in figure 2.
DISCUSSION
This study partly confirms previous work
and partly supplies new information that
would have been very difficult to obtain
by conventional physiological or anatomical
methods.
The evidence for ocular dominance columns
was already massive. In the macaque monkey, microelectrode recordings monitored by
electrode track reconstructions and several
independent anatomical techniques (Nauta/
Fink-Heimer/Wiitanen stain; 3H-proline eye
injection; and reduced silver staining) had all
demonstrated the subdivision of layer IV c
into left-eye and right-eye regions, as well as
revealing the crispness of the segregation.
Until the deoxyglucose method was applied,
the evidence that these columns extended
through all layers (perhaps excepting layer I)
was purely physiological, but was nevertheless compelling. The present study agrees with
the earlier work of the Sokoloff group in providing a confirmation of the extension of the
ocular dominance columns to the upper and
lower layers. Outside layer IV many cells, perhaps about half, are binocular, and the column
376
D. H. HUBEL, T N. WIESEL AND M. P. STRYKER
ORIENTATION COLUMNS IN MACAQUE MONKEY
377
378
D. H. HUBEL, T. N. WIESEL AND M. P. STRYKER
walls as revealed by the deoxyglucose method
would therefore not necessarily be expected to
be a s sharp as in IV c. Moreover, the apparent
widths of the columns as determined by this
method might be expected to vary from layer
to layer depending on responsiveness of cells,
the amount of label uptake, and so on. In fact
the walls turn out in our autoradiographs to
be fairly straight. That the columns in figure
3 are widest in their deepest parts is perhaps
related to the high density of label in layer VI.
The present study provides t h e first a n a tomical demonstration of the orientation columns. The morphological evidence t h a t these
structures are vertically organized and span
all layers except IV c and possibly I is especially welcome, since the physiological evidence,
while strong, was not very direct: It depended
on a few penetrations t h a t were so nearly perpendicular t h a t orientation was constant
throughout (Hubel and Wiesel, '681, on the reconstruction of multiple parallel penetrations
(fig. 3 of Hubel and Wiesel, '631, and on t h e
finding t h a t in oblique penetrations t h e
graphs of orientation plotted against track
distance were often virtually straight lines
through the full cortical thickness, interrupted only in the part of t h e penetration
passing through layer IV c (Hubel and Wiesel,
'74a). That t h e columns in the present deoxyglucose study a r e perpendicular to the surface
is clear not only from their appearance in sections perpendicular to the surface (fig. 4) but
also from the similarity of the patterns in
tangential sections taken at different levels
(fig. 7).
The lack of even a slight suggestion of orientation columns in IV c is itself strong indication t h a t the columns a r e orientation columns, since physiologically we see no trace of
orientation preference in t h a t layer. It is gratifying to find such a good fit between a n a t omy and physiology, especially since the lack
of orientation specificity in IV c has not been
a fact in which one could place extreme confidence. The geniculate inputs to this layer a r e
not orientation selective, and given the difficulty in recording large and clearly defined
spikes in this layer there was always t h e possibility t h a t we were recording only from the afferents, a s might be so if t h e cells themselves
did not fire impulses. The present result does
not completely settle t h e question, however,
since as suggested by Sharp ('76a) uptake by
nerve endings may account for a significant
fraction of the total deoxyglucose uptake. In
this context it is probably worth noting t h a t in
t h e cat the cells are mainly orientation selective whereas t h e geniculate inputs, as in
t h e monkey, are not. The cat's orientation columns are clearly defined even a t t h e base of
layer IV (Stryker e t al., '77).
The marked variation in label density from
layer to layer is at present completely unexplained. In layer IV b, which is sparsely populated with cell bodies, t h e especially high density of label seems to support the idea that
neuropil and cell bodies take up about the
same amount of deoxyglucose, per unit cross
sectional area (Sharp, '76a). But there is certainly no clear inverse correlation between label uptake and cell density: layer V is also
sparsely populated, and is lightly labelled; VI
is both cell-rich and heavily labelled; I has
almost no cell bodies, is rich in nerve terminals, but is hardly labelled at all. These laminar variations in uptake a r e intriguing and
will probably be understood only when the
method is increased in resolution by several
orders of magnitude.
The main new finding in this study is obviously the pattern formed by the sheets of cons t a n t orientation. That this pattern is considerably more complex than the orientationcolumn pattern was already suspected from
t h e high frequency of reversals and fractures
encountered in oblique penetrations. So far
t h e pattern appears not only complex, but also
irregular and unpredictable, but we would
have made a similar conclusion for the eye
dominance columns had we only looked a t a
few regions such as t h e foveal (cf. Hubel and
Wiesel, '72: fig. 17; LeVay et al., '75). A larger
number of regions must still be examined in
more animals before one can say whether or
not there is any consistent or orderly pattern.
Our results a t least seem to rule out any simple order, as well a s any strict relationship between the two sets of columns. Apparently
random intersection is enough to guarantee
t h a t t h e two sets intersect frequently and
never remain parallel over long distances, and
t h a t is probably what is important (Hubel and
Wiesel, '771.
Finally, t h e constancy of the distance separating one column representing vertical from
the next is striking, just as was t h e constancy
of spacing of t h e dominance columns. This
would seem to uphold t h e notion (Hubel and
Wiesel, '74b, '77) t h a t contained in each small
ORIENTATION COLUMNS IN MACAQUE MONKEY
block of cortex, roughly 1 mm X 1 mm, is the
machinery needed to subserve both eyes in all
orientations.
ACKNOWLEDGMENTS
We are grateful to Birthe Storai, Karen Larson, and Jean Thompson for histological assistance and to Carolyn Yoshikami for photography.
Supported by grants from The Rowland
Foundation, Inc., The Esther and Joseph
Klingenstein Foundation, Inc., a n d NIH
Grants E Y 00605 and EY 00606. Doctor
Stryker is supported by a n NIH training
Grant EY 00082.
LITERATURE CITED
Albus, K. 1975 A quantitative study of the projection
area of t he central and paracentral visual field in area 17
of t he cat. 11. The spatial organization of the orientation
domain. Exp. Brain Res., 24: 181-202.
Durham, D., and T. A. Woolsey 1977 Barrels and columnar
cortical organization: Evidence from 2-deoxyglucose
(2DG) experiments. Brain Res., 235.
Hubel, D. H. 1959 Single unit activity in striate cortex of
unrestrained cats. J. Physiol., 147: 226-238.
Hubel, D. H.. and T. N. Wiesel 1962 Receptive fields, binocular interaction and functional architecture in the
cat’s visual cortex. J. Physiol., 160: 106-154.
1963 Shape an d arrangement of columns in cat’s
striate cortex. J . Physiol., 165: 559-568.
1965 Binocular interaction in striate cortex of
kittens reared with artificial squint. J. Neurophysiol., 28:
1041-1059.
1968 Receptive fields and functional architecture of monkey striate cortex. J. Physiol., 295: 215-243.
1970 Cells sensitive t o binocular depth in area
18 of the macaque monkey cortex. Nature, 225: 41-42.
1972 Lam i na r a n d columnar distribution of
geniculo-cortical fibers in t h e macaque monkey. J. Comp.
Neur., 146: 421-450.
1974a Sequence regularity a nd geometry of orientation columns in t h e monkey striate cortex. J. Comp.
Neur., 158: 267-294.
1974b Uniformity of monkey striate cortex: a
parallel relationship between field size, scatter, and magnification factor. J. Comp. Neur., 158: 295-306.
379
1977 Functional architecture of macaque mon
key visual cortex. Proc. Roy. Soc. Lond. B., 298- 1-59.
Hubel, D. H., T. N. Wiesel and S. LeVay 1977 Plasticity of
ocular dominance columns in monkey striate cortex. Phil.
Trans. Roy. Soc. Lond. B., 278: 377-410.
Kennedy. C.. M. H. Des Rosiers, 0. Sakurada, M. Shinohara,
M. Reivich, H. W. Jehle and L. Sokoloff 1976 Metabolic
mapping of t h e primary visual system of t h e monkey by
means of t h e autoradiographic I’T-deoxyglucose terhnique. Proc. Natl. Acad. Sci. (U.S.A.).73: 4230-4234.
D. H. Hubel and T. N. Wiesel 1975 The pattern of
LeVay, S.,
ocular dominance columns in macaque visual cortex
revealed by a reduced silver stain. J . Comp. Neur 259.
559-576.
Schiller, P. H.. B. L. Finlay and S. F. Volman 1976 Quan
titative studies of single-cell properties in monkey striate
cortex. 11. Orientation specificity and ocular dominance.
J . Neurophysiol., 39: 1320.1333.
Sharp, F. R. 1976a Relative cerebral glucose uptake of
neuronal perikarya and neuropil determined with 2-deoxyglucose in resting and swimming r a t Brain Hes.. I 20:
127-139.
1976b Rotation induced increases of glucose
uptake in r a t vestibular nuclei and vestibulocerebellum.
Brain Res., 110: 141.151.
Sharp, F. R., J . S. Kauer a n d G . M. Sheperd 1975 Local sites
of activity-related glucose metabolism in r a t olfactory
bulb d u r i n g olfactory stimulation Brain Res.. 98
596-600.
Sokoloff. L. 1975 Influence of functional activity on
local cerebral glucose utilization. In. Brain Work. The
Coupling of Function, Metabolism and Blood Flow in t h e
Brain. Ingvar, D. H and N. A. Lassen, eds. Academic
Press, New York, pp. 385-388.
Sokoloff. L.. M. Reivich, C. Kennedy, M. H. Des Rosiers, C. S.
Patlak, K. D. Pettigrew, 0. Sakurada and M. Shinohara
1977 The 1’4Cldeoxyglucosemethod for the measurement
of local cerebral glucose utilization: theory, procedure,
a n d normal values in the conscious and anesthetized
albino rat. J . Neurochem., 28: 897-916.
Stryker, M. P., D. H. Hubel and T. N. Wiesel 1977 Orientation columns in t h e cat’s visual cortex. Neurosci. Abstr..
3: 1852.
Wiesel. T. N., D. H. Hubel and D. M. K. Lam 1974 Autoradiographic demonstration of ocular-dominance columns in
t h e monkey striate cortex by means of transneuronal
transport. Brain Res., 79: 273-279.
Wurtz, R. H. 1969 Visual receptive fields of striate cortex neurons in awake monkeys. J . Neurophysiol., 32.
727-742.
~
.