Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
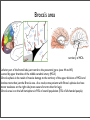
Cerebral Cortex 3 Language and brain Hemispheric difference Yasushi Nakagawa Department of Neuroscience University of Minnesota 1 Speech and Language Vocal communication mediated by fully-developed speech and language is a uniquely human trait. Speech: mechanical aspects of verbal communication (articulation, voice, fluency) Language: a higher- order function, based on accepted rules that govern what words mean, how to make new words, how to put words together, and what word combinations are appropriate in specific situations useful website to check: http://www.asha.org/public/speech/development/language_speech.htm Speech and language disorders expressive: problems in expressing speech or language receptive: problems understanding speech or language mixed: both comprehension and production are impaired Speech disorders: lisp (articulation), stuttering (fluency), dyspraxia (generation and proper sequencing of speech sounds), dysarthria (voice) Language disorders: deficits in encoding or decoding information in phrases and sentences according to accepted rules, such as those of grammar. Language disorders include specific language impairment (SLI) and dyslexia. 2 What is aphasia? a disorder that results from damage to the parts of the brain that contain language -affects one million people in the U.S. -caused by: stroke (most common) 25-40% of patients with stroke survivors have aphasia head trauma...about 1/3 of severe head injuries are accompanied by aphasia brain tumors infections causes problems with any or all of the following: speaking, listening, reading, and writing Individuals with aphasia may also have other problems, such as dysarthria, apraxia or swallowing problems. A classical model of brain and language was developed in the 19th century by neurologists studying patients with aphasia. 3 Specific cortical regions are involved in 1 language processing C H A P T E R Pierre Paulof Broca (1824-1880) Neurobiology Language -French surgeon, neurologist, 1 2 neuroanatomist ven L. Small andhe Gregory Hickoka patient who had severe impairment of language. This patient -In 1861, came across sity of California, Irvine, CA, USA; 2Department of Cognitive Sciences, Center for spoke only “tan, tan”, although heCA, could r for Cognitive Neuroscience, University of California, Irvine, USA understand the language pretty well. After he died, Broca did an autopsy and found a large lesion in the left frontal lobe. RY gical basis of human us of attention in medibrain basis of language Pierre Paul Broca in the he patient Louis Victor ented to the Hôpital lty speaking, purport,” sometimes as a pair by gestures (Domanski, r until autopsy, when hat some neurological lection of serous fluid) osterior inferior frontal A subsequent patient, f speech output (five n not dissimilar to that he ongoing debates at of language, including uage” to the frontal aud, 1825; Gall & oca to investigate this FIGURE 1.1 The exterior surface of the brain of LeBorgne (“tan”). -Broca saw about(1874), ten the more patients over the next several years and found that everyone Wernicke diagram-making of Lichtheim (1885)symptoms (Figure 1.2) and Grashey (1885),in thethe anatomy with similar had a lesion left offrontal lobe. Déjerine (1895), and of course many other contributors. In the past century, Norman Geschwind recapitulated and added to the language “center” models that preceded him and presented a reconceptualized “connectionist” view of the brain mechanisms of language 4 Broca’s aphasia -non-fluent, telegraphic, poorly articulated verbal output -meaning is conveyed by content or information-carrying words such as nouns and verbs (nouns are named more accurately than verbs) -function words (e.g., articles, conjunctions, prepositions, auxiliary verbs, pronouns) are often omitted (agrammatic speech production) -An example of Broca’s aphasia: http://www.youtube.com/watch?v=f2IiMEbMnPM 5 6 Broca’s area 3. THE VENTROLATERAL FRONTAL REGION 68 2 Vascularization of the Brain and Spinal Cord a 5 6 b 4 4 3 3 7 8 2 5 6 7 2 6 7 1 5 4 8 1 9 3 11 2 10 12 Fig. 2.5 (a) Vascularization of the medial surface of the cerebral cortex by the anterior (ACA in light red) and posterior (PCA in red) cerebral arteries. The vascular territory of the middle cerebral artery (MCA) is uncoloured. The ACA gives off the following branches: 1 orbitofrontal artery; 2 frontopolar artery; 3–5 anterior, middle and posterior frontal arteries; 6 paracentral artery; 7, 8 superior and inferior parietal arteries. The PCA divides into: 1 hippocampal arteries (not shown); 2–4 anterior, middle and posterior temporal arteries; 5 calcarine artery; 6 parieto-occipital artery; 7 splenial artery (after ten Donkelaar et al. 2007). (b): Vascularization of the lateral surface of the cerebral cortex, largely by the middle cerebral artery (MCA; uncoloured). The vascular territories of the ACA and PCA are indicated in light red and red, respectively. The MCA gives off the following branches: 1 orbitofrontal artery; 2 prefrontal artery; 3 precentral artery; 4 central artery; 5, 6 anterior and posterior parietal arteries; 7 angular artery; 8 temporo-occipital artery; 9–11 posterior, middle and anterior temporal arteries; 12 temporopolar artery (based on Nieuwenhuys et al. 1988; after ten Donkelaar et al. 2007). (3D). 3D-TOF is usually better for vessels with high flow velocity such as the cervical arteries and the circle of Willis, whereas 2D-TOF is more appropriate for assessing vertebral veins and sinuses. Phase-contrast MRA is based on signals coming from phase shifts of protons in flowing blood. With this technique, it is possible to combine imaging with measurement of flow velocities and flow directions. and extending to the superior frontal sulcus on the superior lateral surface; posteriorly, the arterial territory extends to the parieto-occipital sulcus; and (2) callosal branches for the rostrum, genu, corpus and splenium of the corpus callosum. The terminal pericallosal branches are joined posteriorly by the splenial branches of the PCA. Cortical arteries of the M4 segment of the MCA extend over the lateral surface of the hemisphere, usually to the superior frontal sulcus, the intraparietal sulcus and the inferior temporal gyrus. On the orbitofrontal surface, the arterial territory of this artery includes the lateral orbital gyri. Leptomeningeal branches of the PCA include the hippocampal arteries, the splenial artery for the splenium of the corpus callosum, and cortical branches to the inferomedial surfaces of the temporal and occipital lobes, extending to the parieto-occipital fissure (for hippocampal arteries, see Erdem et al. 1993; Duvernoy 1998; Huther et al. 1998). Perforating branches of the cerebral arteries are the hypophysial arteries, the anterior choroidal artery, the anterior communicating artery, perforating branches from the anterior, middle and posterior cerebral arteries, thalamoperforating and thalamogeniculate branches from the PCA, and the posterior choroidal arteries (see Sect. 2.6). The variability of the territories of the major cerebral arteries has extensively been studied by van der Zwan (1991) and van der Zwan et al. (1992). van der Zwan simultaneously injected the six major cerebral arteries (under the same pressure) with different-coloured Araldite F mixtures under standardized conditions to obtain the most realistic territorial distribution. The variability of the territories of the major cerebral arteries appeared to be much larger than generally is described in the literature (Fig. 2.6): territory of MCA FIGURE 3.2 The sulcal and gyral morphology of the ventrolateral frontal region in the human brain. The shaded region represents the orbitofrontal cortex that is continuous with the pars orbitalis of the inferior frontal gyrus. Abbreviations: aalf, anterior ascending ramus of the lateral fissure (ascending sulcus, vertical sulcus); ascs, anterior subcentral sulcus; cs, central sulcus; ds, diagonal sulcus; half, horizontal anterior ramus of the lateral fissure (horizontal sulcus); IFG, inferior frontal gyrus; ifs, inferior frontal sulcus; iprs, inferior precentral sulcus; los-p, posterior ramus of the lateral orbital sulcus; MFG, middle frontal gyrus; Op, pars opercularis of the inferior frontal gyrus; Or, pars orbitalis of the inferior frontal gyrus; PrG, precentral gyrus; prts, pretriangular sulcus; ScG, subcentral gyrus; STG, superior temporal gyrus; sts, superior temporal sulcus; Tr, pars triangularis of the inferior frontal gyrus; ts, triangular sulcus (incisura capitis). -inferior part of the frontal lobe just rostral to the precentral gyrus (area 44 and 45) -served by upper branches of the middle cerebral artery (MCA) -Broca’s aphasia is the result of massive damage to the territory ofSupply the divisions of MCA and 2.5 Arterial of theupper Cerebral Cortex involves more than just the Broca’a area. As a result, some patients with Broca’s aphasia also have motor weakness on the right side (more severe for arms than for legs). subdivisions of the inferior frontal region (Mohr, 1976; et al., 1978). The syndrome aphasia, -Broca’s area is on the left Mohr hemisphere in 90%of Broca’s of overall population (70% of left-handed people) which is characterized by severe impairment in lan- GURE 3.1 Cytoarchitectonic map of the lateral surface of the man and the macaque monkey frontal lobe by Petrides and Pandya 94). The white region on the precentral gyrus is the primary motor tex (area 4) and the various subdivisions of the premotor region ea 6). The inset shows the location of area 44 in the macaque monkey the fundus of the inferior limb (ramus) of the arcuate sulcus. ontal cortical region that plays a critical role in cerin aspects of language production (Friederici, 2011; eschwind, 1970; Grodzinsky, 2000). Several attempts guage production (including impaired syntactic processing), is the result of massive damage to the territory of the upper division of the middle cerebral artery and involves not only the cortical structures in the posterior part of the inferior frontal gyrus (i.e., areas 44 and 45) but also the adjacent frontoparietal opercular region and the anterior parts of the insula (Ackermann & Riecker, 2004; Baldo, Wilkins, Ogar, Willock, & Dronkers, 2011; Dronkers, 1996; Mohr, 1976). The best evidence thus far linking specific parts of the inferior frontal gyrus to language production has been obtained from electrical stimulation of the cerebral cortex under local anesthesia during brain sur- The arterial cerebral circulation can be divided into two systems: (1) the leptomeningeal arteries, consisting of the terminal branches of the anterior, middle and posterior cerebral arteries (for collateral circulation, see Vander Eecken 1959; Duvernoy et al. 1981); and (2) the perforating branches that perforate the brain parenchyma as direct penetrators. They arise from the circle of Willis, from its immediate branches and also from the leptomeningeal arteries, and supply the basal ganglia, the internal capsule and the diencephalon. Leptomeningeal branches of the three cerebral arteries and their territories are shown in Fig. 2.5. The leptomeningeal anastomoses in the subarachnoid space between the arterial boundary zones represent connections between distal branches of major cerebral arteries (Duvernoy et al. 1981). These arterial boundary zones, also termed “watershed zones”, are especially susceptible to damage following any generalized decrease in blood flow as is the case in severe systemic hypotension. The distal segment of the ACA, the pericallosal artery, usually gives rise to: (1) cortical branches to the medial surface of the hemisphere including the medial orbital gyri on the orbitofrontal surface of the hemisphere 6 Wernicke’s aphasia -first reported by Karl Wernicke (1848-1905) -fluent, effortless, but relatively meaningless, spontaneous speech and repetition -impaired comprehension at the word, sentence, and discourse levels -Spoken language may be limited to jargon comprising either real words or neologisms (nonwords such as “klimorata”) or a combination of the two. -usually caused by neural dysfunction in regions supplied by the lower branches of the left MCA -Wernicke’s area includes posterior, superior part of the temporal lobe as well as inferior part of the parietal lobe Wernicke’s area -An example of Wernicke’s aphasia: http://www.youtube.com/watch?v=aVhYN7NTIKU&feature=relmfu 7 Specific cortical regions are involved in language processing -Wernicke proposed that complex cognitive functions result from interconnections between several regions of the brain Wernicke postulated that language involves separate motor and sensory programs: -sensory functions: angular gyrus integrates visual and auditory information, and spoken or written words are transformed into neural code in Wernicke’s area, it is recognized as language and associated with meaning -motor functions: Broca’s area contains the rules or grammar for transforming sensory information into a motor representation 8 Conduction aphasia Wernicke’s area and Broca’s area are connected by a robust fiber bundle named arcuate fasciculus. Wernicke found patients who show a type of aphasia different from Broca’s and Wernicke’s aphasia. Conduction aphasia -intact understanding of written and spoken words -no motor difficulty when they speak -lack of coherency (omit parts of words or substitute incorrect sounds) Traditionally, conduction aphasia is thought to be caused by a lesion in the arcuate fasciculus and thus is considered a “disconnection syndrome”. However, this hypothesis has been recently challenged. 9 Functional imaging of language areas PET (positron emission tomography) A. looking at words activates primary visual area and visual association area B. listening the same words as used for A activates temporal cortex including Wernicke’s area C. speaking words (repeating a word presented through either earphone or on a screen) activates Broca’s area D. subjects were asked to respond to the word “brain” with an appropriate verb (e.g., to think); many areas are activated (frontal lobe, Broca’s, Wernicke’s) 10 Does Broca’s area contain grammatical rules of language? -in 1960s, linguist Noam Chomsky proposed that all natural languages share a common design (universal grammar) -is there a innate system in the human brain that evolved to mediate the grammatical design of language? -if so, where in the brain does such a system reside? 11 Does Broca’s area contain grammatical rules of language? -If a second language is learned early, both the native and the second languages are represented in a common region in Broca’s area (A). -If a second language is learned in adulthood, it is represented in a separate region in Broca’s area (B). -Wernicke’s area does not show separation between languages. -Learning an artificial languages (violating the rules of “universal grammar”) as a second language does not increase the activity of Broca’s area. -Broca’s area must contain some kind of constraints that determine the structure of all natural languages. 12 Sign language -Deaf people use ASL (American Sign Language) that uses hand gestures instead of sound but has the same structural complexity as spoken languages. -Signing is also localized to the left hemisphere. -Damage to the left hemisphere can have specific consequences for signing just as for spoken languages (right hemisphere damages rarely have consequences). -damage to Wernicke’s area....comprehension is affected -damage to Broca’s area...grammar and fluency are affected -Emergence and operation of language capability in the left hemisphere does not need functional auditory system. 13 Chimpanzee’s language areas? -Chimps intentionally and referentially communicate via manual gestures and, like humans, preferentially use their right hand for communicative gestures. -They also use vocal signals to communicate with humans. -Portions of the left inferior frontal gyrus (Broca’s area in humans) as well as other cortical and subcortical regions in chimpanzees are active during the production of communicative signals. -Neurological substrates underlying language production in the human brain may have been present in the common ancestor of humans and chimpanzees. -However, connection between Wernicke’s and Broca’s area (dorsal pathway: arcuate fasciculus) was much less robust in non-human primates. 766 15 The Cerebral Cortex and Complex Cerebral Functions a cs ips 7b 22 6 b cs 7a sts ips 8 40 22 44 6 45 45 44 c 47 37 sts cs ips 39 46 40 8 39 10 44 45 47 22 6 sts 37 a: macaque b: chimpanzee c: human 14 New models of language and brain 14126 • J. Neurosci., October 10, 2012 • 32(41):14125–14131 Poeppel et al. • Towards a New Neurobiology of Language Figure 1. A, The classical brain language model, ubiquitous but no longer viable. From Geschwind (1979). With permission of Scientific American. B, The dorsal and ventral stream model of speech sound processing. From Hickok and Poeppel (2007). With permission from Nature Publishing Group. Recent advances of non-invasive functional neuroimaging (fMRI, etc.) and other research new maps of the functional anatomy of language. Speaking spasemantics,” and so on. From a neurobiological perspective, the goal tially,have local regions, processing hemispheres, is tomodel. develop mechanistic (and ultimately explanatory) linking hyfields identified a streams, need the totwo revise the and classic distributed global networks are now implicated in language funcpotheses that connect well defined linguistic primitives with equally tion in unanticipated ways. For example, the canonical language well defined neurobiological mechanisms. It is this next phase of region, Broca’s is now known, of based innovativeof cytoneurolinguistic research that both is now beginning, developing com-Broca’s area area, is composed a on number subregions, and serve language and anonarchitectural and immunocytochemical data, to be composed of putational neurobiology of language, and there are grounds for oplanguage a numberfunctions. of subregions (on the order of 10, ignoring possible timism that genuine linking hypotheses between the neurosciences laminar specializations), plausibly implicating a much greater and the cognitive sciences are being crafted (Poeppel, 2012). number of different functions than previously assumed (Amunts Is the classical model salvageable? Are the new approaches and -The languages are into at leastinsights twomerely processing (dorsal et al., 2010; Fig. 2), areas supporting bothorganized language and non-language expansionsstreams and adaptations to the and traditional processing. Moreover, there is emerging consensus that regions view? We submit that the model is incorrect along too many ventral) are organized into at least two (and likely more) processing dimensions for new research to be considered mere updates. streams, dorsal and ventral streams (Hickok and Poeppel, 2004; While these issues have been discussed in more detail, we point to Saur et al., 2008), that underpin different processing subroutines, just two serious shortcomings. First, due to its underspecification for example, mediating aspects of lexical recognition and lexical both biologically and linguistically, the classical model makes 15 Hemispheric differences asymmetry in the human body -mostly symmetric -preferential use of a specific hand for skilled actions (most people use the right hand) -hand preference can be observed already in the first week of life -are hand preference and left hemisphere dominance of language associated? -95% of right-handers are left-dominant for language -70% of left-handers are also left-dominant for language Broca: “we speak with the left hemisphere” (1865) right-hand preference and left hemisphere dominance of language are very clear 16 Laterality or asymmetry? Jackson: complementary role for the right hemisphere in visuospatial functions (1874) 1960s: Roger Sperry’s hemisphere specialization theory “indeed a conscious system in its own right, perceiving, thinking, remembering, reasoning, willing, and emoting, all at a characteristically human level, and . . . both the left and the right hemisphere may be conscious simultaneously in different, even in mutually conflicting, mental experiences that run along in parallel” Hemispheric Differences 757 15.36 Lateralized tions revealed by psychocal tests of commissurotomy ents (after Sperry 1967) -Although it is generally accepted that right hemisphere is better equipped with visuo-spatial information, this form of asymmetry is much less clear. Left visual field Right visual field Image of right visual field Image of left visual field Left hemisphere Right hemisphere -Correlation between measures of laterality for different functions are generally very low or nonsignificant. Memory for shapes Verbal memory Left hand manipulation and feeling shapes Articulating speech Right-hand skill (writing, etc) -Asymmetry is a better term than laterality. Hearing speech (right ear advantage) Superior comprehension of language Hearing environmental sounds (left ear advantage) -Asymmetry is not even specific to humans. Superior recognition of topological forms, faces, etc. Body image Right visual field ding actions. For more abstract representations of objects scenes, for instance for reasoning or problem solving, metric information is not necessary (see also Jager and Left visual field from Sperry (1967) indicating damage to the PPC, more in particular to the right IPL. Later on reports were published on patients with typical left hemineglect and lesions in the region of the FEF, the 17 What is special about right hemisphere? Comparison of stroke due to right and left MCA occlusion -Both cause contralateral hemiparesis (motor weakness) and loss of sensory function. -Left lesion can also involve severe language deficit. -In right hemisphere lesion, patients are often unaware of their functional loss and pay no attention to their disability (“anosognosia”). -Anosognosia is often accompanied by left hemispatial neglect (caused by lesions in the posterior parietal cortex). -Similar lesions in the left hemisphere cause much less severe hemispatial neglect than the right-side lesion. -Prosopagnosia (problem in face recognition) is caused by bilateral lesion of the temporal lobe, and functional imaging shows both hemispheres are active during face recognition. -However, unilateral anterior temporal lesions have different patterns of impairment depending on the side that is affected: right lesion--loss of familiarity feeling, person-specific information retrieval left lesion--impairment in name finding 18 y impaired. be lesions, e pattern of s, there was fic informaa prevalent cognition is s have been e, the interselectively sing materials, the aphasia ugh someamusia was mplex funcelodies and gnitive proPeretz and o be a spey, suggestmay involve so Chap. 7). ence and as ody is recpitch from here capacsical stimual musician n a normal music is a way matey. Expertise rned to lisZatorre and Anatomical, functional and genetic differences between right and left hemispheres 759 a Structural analysis HG HG -size of gyri in temporal cortex: L>R -similar in chimpanzee cortex -size difference already found in 5 months-old fetuses b si TG1 TG2 sh TG PT PT pm pm c HG Functional analysis -Connectional asymmetry also found early in development -DTI studies found that connections between Broca’s and Wernicke’s areas are much more robust on the left side. -People with symmetrical distribution had higher performance in verbal memory tasks HG Gene expression analysis L R Fig. 15.38 Anatomical differences between the left and right hemispheres. In (a), von Economo and Horn’s data on asymmetry of Heschl’s gyrus (HG) are shown; in (b) Geschwind and Levitsky’s data; and in (c) Galaburda’s data on asymmetry of the planum temporale. pm posterior margin of planum temporale, PT planum temporale, sh sulcus of Heschl, si sulcus intermedius, TG, TG1, TG2 transverse gyri of Heschl (a after von Economo and Horn 1930, (b) after Geschwind and Levitsky 1968, and (c) after Galaburda et al. 1978b) -identified 27 genes whose levels of expression is already different between right and left cortex of 14-weeks old human fetuses ences right cered Levitsky he areas of ). The left he primary mal brains, asymmetry and Horn (1930). It appeared that the auditory association cortex is larger in the left hemisphere of the human brain (Galaburda et al. 1978a, b; for recent data see Dorsaint-Pierre et al. 2006; Toga and Thompson 2007). Comparative studies between human and non-human primates showed that chimpanzees, similarly to humans, show an asymmetry in the planum temporale (Gannon et al. 1998; Hopkins et al. 1998). However, only humans have a robust asymmetry -Intrinsic, genetic program early in development may contribute to the later anatomical and functional differences between right and left hemispheres. 19 Genetic factors contribute to susceptibility to speech and language impairments Speech disorders: lisp (articulation), stuttering (fluency), dyspraxia (generation and proper sequencing of speech sounds), dysarthria (voice) Language disorders: deficits in encoding or decoding information in phrases and sentences according to accepted rules, such as those of grammar. Language disorders include specific language impairment (SLI) and dyslexia. Speech and language disorders are: -heritable and show strong familial aggregation -an increase in monozygotic twin concordance rates over that of dizygotic twins ➔Much of this aggregation can be attributed to genetic influences. Researchers have recently begun to identify genetic factors that may play a role in the etiology of speech and language disorders. 20 Methods for Identifying Contributory Genetic Variants For some genetic disorders, it is possible to select candidate genes on the basis of their function alone. However, for speech and language disorders, in which the underlying biological mechanism is unclear, the identification of susceptibility genes usually starts with an unbiased screening approach. This step allows the identification of a candidate region in the human genome and thus acts to reduce the number of possible contributory genes to a manageable size prior to a more in-depth investigation. These screening approaches usually take the form of genome-wide linkage or association studies. One investigates families with members affected by the disorder under study. Linkage studies look for regions of the human genome in which there is a correlation between the level of genetic identity and the level of phenotypic similarity for any given sib pair. . 21 KE family and verbal dyspraxia Verbal dyspraxia: difficulties in the control of orofacial muscles leading to a deficit in the production of fluent speech. http://www.youtube.com/watch?v=tYmm23EPXjU In 1980’s, a large family was found to be affected by this disorder. In addition to their speech problems, affected members of this family also had expressive and receptive language deficits and, in some cases, written language problems and nonverbal cognitive impairment. The pattern of inheritance indicated that they may be caused by a mutation in a single gene. 22 KE family and verbal dyspraxia Genome-wide linkage analysis identified linkage to chromosome 7q (Fisher et al., 1998) and fine mapping of the locus indeed identified a mutation in the FOXP2 gene. The relevance of FOXP2 mutations to other cases of verbal dyspraxia was supported by the identification of an unrelated child with a similar form of speech impairment who was found to have a chromosome rearrangement that disrupted the FOXP2 gene The FOXP2 gene encodes a DNA-binding protein from the FOX family. This protein acts as a transcriptional repressor. The identification of FOXP2 opened up a whole field of research encompassing a wide range of disciplines including neuroimaging, animal models (primarily mouse and songbird), molecular studies of gene function and expression, and population and evolutionary studies 23