Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
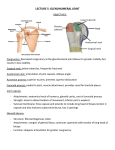
PC 6/7 ABSTRACT NO. 0934 PAPER NO. 0372 Corresponding Author: John First Name Novotny Last Name Presenting Author: John First Name Novotny Last Name I prefer a Poster Presentation Please consider for New Investigator Recognition Awards Please consider for the American Geriatrics Society Award REQUIRED - Supply first keyword from one of these lists: Supply 4 remaining keywords from the list in the Instructions: None Upper Extremity Glenohumeral Joint Anatomy Animal Model COMPARATIVE ANATOMY OF THE HUMAN GLENOHUMERAL JOINT TO THREE ANIMAL MODELS +*Novotny, J.E., Macy, J.C., Nichols, C.E. III, Beynnon, B.D.; +*The University of Vermont, McClure Musculoskeletal Research Center, Dept. of Orthopaedics and Rehab., Stafford Hall, Burlington, Vermont, USA 05405; PH (802)656-2250; FAX:(802)656-4247; [email protected] Introduction: Study of injuries to the glenohumeral joint, its capsule and articular surfaces and their healing responses requires the development of an adequate animal model. Knowledge of the differences in the anatomy compared to the human is necessary. There is little known about injury and healing response of the glenohumeral joint capsule after dislocation. Animal models have been developed to address these issues in other joint systems. The rat has been identified as a model of rotator cuff injury [Soslowsky, 1996]. Rabbits have been used in studies of the knee ligaments [Woo, 1992, Walsh, 1992]. Primate models might be more anatomically accurate for studying the glenohumeral joint. The objective of this work is to describe the differences in the glenohumeral joint anatomy of the rat, rabbit and monkey compared to human. An understanding of the bony, muscular and capsular anatomy of these animal models will aid in assessing their future usefulness in the study of human glenohumeral pathology. Methods: Three common animals used in medical research were studied. These were the Sprague-Dawley rat (Rattus norvegicus), white New Zealand rabbit (Oryctolagus cuniculus) and Rhesus monkey (Macaca mullata). Three other animals were excluded earlier due to growth or gross morphological differences: chicken, pig and cow. Four specimens of each animal were dissected with detailed examination of the muscles and soft tissues of the glenohumeral joint. Photographs were taken at each step of the dissection for documentation and comparison. The outer layers of skin and fat were removed. The layers of muscle over the shoulder girdle were dissected from those near the surface to the rotator cuff. An attempt to identify all human muscles was made and others were noted. The tendons of the rotator cuff muscles were separated from the glenohumeral capsule, and thickenings within the capsule were observed. The glenoid labrum was examined. The bony and articular surfaces were finally noted after the capsule was removed. The results note differences of the animal model compared to the human anatomy. Results: Rat: Bone: The greater tuberosity projects from the shaft by more than the shaft’s diameter and extends distally. The scapula is more tear-drop shaped than triangular. The coracoid projects from the scapula adjacent to the anterior rim of the glenoid. The acromion extends laterally from the scapular spine. Muscle: The muscles include all those in the human and also a dorso-epitrochlearis. This runs from the latissimus dorsi tendon to the distal humerus. Teres minor is very small in comparison to the other rotator cuff muscles. Capsule: The capsule is very thin, almost entirely translucent. There is a thin anterior band, running from the superior glenoid to the inferior-medial humeral head similar to the SGHL. There are also thickenings posterior-inferiorly, perhaps representing an IGHL complex. No coracohumeral ligament was identified An axillary recess is present. A labrum is robust and attached firmly to the glenoid rim. Rabbit: Bone: The humerus is thick with a prominent greater tuberosity and deep bicipital groove. The scapula is triangular and the medial-lateral dimension is twice the superior-inferior. The glenoid face has distinct superior and inferior facets with the superior facet directed inferiorly. The facets are not continuous with an obvious transition between them. The acromion extends inferiorly from the spine of the scapula. The clavicle is not attached to the acromion. Muscle: All human muscles are present. Capsule: There were no real thickenings visible within the capsule. There was a dense thick intra-articular ligament from the anterior- superior glenoid to the anterior-medial humeral head distinct from the capsule consistent with the coracohumeral ligament in the human. There was no axillary recess or labrum. A rotator interval was present. No coracoacromial ligament was identified. Monkey: Bone: There is less retroversion of the humeral head with respect to the epicondylar line. The scapula is triangular with the medial-lateral dimension one and a half that of the superior-inferior. The glenoid is concave in the supero-inferior dimension with the superior facet angled inferiorly. There is a prominent supraglenoid tubercle. The coracoid projects inferolaterally. The acromion extends laterally from the scapular spine. Muscle: All human muscles are present along with three others. The intraspinatus runs along the distal scapular spine to the greater tuberosity between the supraspinatus and infraspinatus. The dorso-epitrochlearis is present as in the rat. The coracobrachialis profundus runs from the tip of the coracoid to the anterior humerus distal to the lesser tubercle. Capsule: There is a prominent intra-capsular thickening from the superior tubercle of the glenoid to the anterior-medial humeral head similar to the SGHL. Also there is a thickening extending from the superior glenoid to the medial humeral head, or IGHL. A distinct extra-capsular thickening from the base of the coracoid to the anterior humeral head is consistent with the coracohumeral ligament. An axillary recess and rotator interval were also present, as well as a firmly attached labrum. There is no coracoacromial ligament. Discussion: None of these three models is an ideal representation of the anatomy of the human glenohumeral joint. The primate and rat have distinct capsular and glenoid geometries that are nearer to human than the rabbit. None of the animals have similar scapular geometries and differences in the humerus are evident. Differences in the acromion and coracoid are also present. Muscular anatomies are consistent in the rotator cuff for the rat and rabbit, with the monkey having the additional intraspinatus and coracobrachialis profundus deeply, and the monkey and rat having the dorso-epitrochlearis nearer to the surface. Consideration of the more bipedial biomechanics and greater size of the joint may make the primate model more useful, although practical, monetary concerns may lead to use of the rat. The many anatomical differences in the rabbit make it less useful. Acknowledgments: Bristol-Meyers Zimmer Squibb Excellence in Orthopaedics Award. References: Soslowsky et al.: Development and use of an animal model for investigations on rotator cuff disease. J Should Elbow Surg 5: 383-392, 1996. Walsh and Frank: Two methods of ligament injury: a morphological comparison in a rabbit model. J Surg Res 45: 159-166, 1988. Woo et al.: Biology, healing and repair of ligaments. In Biology and Biomechanics of the Traumatized Synovial Joint: the Knee as a Model.: Editors: Finerman and Noyes. AAOS, Rosemont, IL 1992. One or more of the authors have received something of value from a commercial or other party related directly or indirectly to the subject of my presentation. The authors have not received anything of value from a commercial or other party related directly or indirectly to the subject of my presentation. 45th Annual Meeting, Orthopaedic Research Society, February 1-4, 1999, Anaheim, California 372