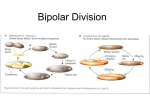
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Plant, Cell and Environment (2009) 32, 666–681 doi: 10.1111/j.1365-3040.2009.01926.x Regulation and function of root exudates DAYAKAR V. BADRI & JORGE M. VIVANCO Centre for Rhizosphere Biology and Department of Horticulture and LA, Colorado State University, Fort Collins, CO 80523, USA ABSTRACT Root-secreted chemicals mediate multi-partite interactions in the rhizosphere, where plant roots continually respond to and alter their immediate environment. Increasing evidence suggests that root exudates initiate and modulate dialogue between roots and soil microbes. For example, root exudates serve as signals that initiate symbiosis with rhizobia and mycorrhizal fungi. In addition, root exudates maintain and support a highly specific diversity of microbes in the rhizosphere of a given particular plant species, thus suggesting a close evolutionary link. In this review, we focus mainly on compiling the information available on the regulation and mechanisms of root exudation processes, and provide some ideas related to the evolutionary role of root exudates in shaping soil microbial communities. Key-words: ABC transporters; elicitors; mechanism of secretion; root system architecture; tritrophic interactions. INTRODUCTION The main functions of the ‘hidden’ part of the plant, its root system, have traditionally been thought to be anchorage and uptake of nutrients and water. However, roots secrete an enormous range of compounds into the surrounding soil. This area, called the rhizosphere, can be divided into three zones: endorhizosphere (root tissue, including the endodermis and cortical layers), rhizoplane (the root surface with the epidermis and mucilage) and ectorhizosphere (the soil nearby the root) (Lynch 1987). The first observation that microbes are more abundant in the rhizosphere than in distant soil was made by Hiltner (1904), and the first indication concerning root exudation and microbe abundance was provided by Knudson (1920) and Lyon & Wilson (1921). In recent years, the field of rhizosphere biology has explored the relative importance of root exudates in mediating interactions with neighbouring plants and microbes (Bais et al. 2004, 2006; Weir, Park & Vivanco 2004; Broeckling et al. 2008). Root exudation is part of the rhizodeposition process, which is a major source of soil organic carbon released by plant roots (Hutsch, Augustin & Merbach 2000; Nguyen 2003).The quantity and quality of root exudates are determined by plant species, the age of an individual plant and external factors like biotic and abiotic stressors. Root Correspondence: J. M. Vivanco. Fax: +1 970 491 7745; e-mail: [email protected] 666 exudation clearly represents a significant carbon cost to the plant (Marschner 1995), with young seedlings typically exuding about 30–40% of their fixed carbon as root exudates (Whipps 1990). Root exudates contain released ions (i.e. H+), inorganic acids, oxygen and water, but mainly consist of carbon-based compounds (Uren 2000; Bais et al. 2006). These organic compounds can often be separated into two classes: low-molecular weight compounds, which include amino acids, organic acids, sugars, phenolics and an array of secondary metabolites, and high-molecular weight compounds like mucilage and proteins. The classes of compounds secreted by roots are listed in Table 1. Despite the technical difficulties inherent in the study of plant roots, significant advances in root research have been made using molecular and genetic tools. In addition, the knowledge gained by studying the root system of the model plant Arabidopsis thaliana (L.) Heynh. has been indispensable in advancing our understanding of the impact of agricultural practices on root development and the impact of roots (and their exudates) on the soil environment (Bucher 2002). Molecular tools, such as cloning of root-specific genes, differential and subtractive hybridization techniques (Conkling et al. 1990; Rodriguez & Chader 1992), differential display (Liang & Pardee 1992), root-specific cDNA libraries (Bucher et al. 1997) and analyzing root cell-specific gene expression using a combination of molecular tools (Birnbaum et al. 2003), have resulted in the rapid compilation of new information on root development, physiology and biochemistry. In addition, the development of ‘composite plants’ (transgenic ‘hairy root’ systems with nontransgenic shoot systems) by employing Agrobacterium rhizogenes-mediated transformation methods has enabled studies of root-specific biochemistry, endosymbiosis, production of secondary metabolites and root-specific interactions (Boisson-Dernier et al. 2001; Choi et al. 2004; Lee et al. 2004; Limpens et al. 2004). Root exudates mediate both positive and negative interactions in the rhizosphere. The positive interactions include symbiotic associations with beneficial microbes, such as mycorrhizae, rhizobia and plant growth-promoting rhizobacteria (PGPR). Negative interactions include association with parasitic plants, pathogenic microbes and invertebrate herbivores. In this review, we do not intend to provide exhaustive coverage of the large body of literature that has been published on root exudates and their interaction with soil organisms. The reader is referred to other reviews for further information on these areas (Bertin, Yang & Weston © 2009 Blackwell Publishing Ltd Regulation and function of root exudates 667 Table 1. Classes of compounds released in plant root exudates Class of compounds Single componentsa Carbohydrates Amino acids Organic acids Arabinose, glucose, galactose, fructose, sucrose, pentose, rhamnose, raffinose, ribose, xylose and mannitol All 20 proteinogenic amino acids, l-hydroxyproline, homoserine, mugineic acid, aminobutyric acid Acetic acid, succinic acid, l-aspartic acid, malic acid, l-glutamic acid, salicylic acid, shikimic acid, isocitric acid, chorismic acid, sinapic acid, caffeic acid, p-hydroxybenzoic acid, gallic acid, tartaric acid, ferulic acid, protocatacheuic acid, p-coumaric acid, mugineic acid, oxalic acid, citric acid, piscidic acid Naringenin, kaempferol, quercitin, myricetin, naringin, rutin, genistein, strigolactone and their substitutes with sugars Catechol, benzoic acid, nicotinic acid, phloroglucinol, cinnamic acid, gallic acid, ferulic acid, syringic acid, sinapoyl aldehyde, chlorogenic acid, coumaric acid, vanillin, sinapyl alcohol, quinic acid, pyroglutamic acid Umbelliferone Benzyl aurones synapates, sinapoyl choline Cyclobrassinone, desuphoguconapin, desulphoprogoitrin, desulphonapoleiferin, desulphoglucoalyssin Cyanidin, delphinidin, pelargonidin and their substitutes with sugar molecules Indole-3-acetic acid, brassitin, sinalexin, brassilexin, methyl indole carboxylate, camalexin glucoside Linoleic acid, oleic acid, palmitic acid, stearic acid Campestrol, sitosterol, stigmasterol Jugulone, sorgoleone, 5,7,4′-trihydroxy-3′, 5′-dimethoxyflavone, DIMBOA, DIBOA PR proteins, lectins, proteases, acid phosphatases, peroxidases, hydrolases, lipase Flavonols Lignins Coumarins Aurones Glucosinolates Anthocyanins Indole compounds Fatty acids Sterols Allomones Proteins and enzymes a List of single components presented in this table is reported mainly from the model plant Arabidopsis (see Narasimhan et al. 2003), and the represented list is partial. 2003; Karthikeyan & Kulakow 2003; Bais et al. 2004; 2006; Singh et al. 2004, Weir et al. 2004, Barea et al. 2005; Morgan, Bending & White 2005; Prithiviraj, Paschke & Vivanco 2007; Bais, Broeckling & Vivanco 2008). Instead, this review focuses on exploring the current knowledge relating to the regulation and mechanism of root exudation, and how these processes impact the plant at both the individual and ecosystem level. Furthermore, in this review, we argue that the above-ground and below-ground diversities are linked by plants, and that plant root-secreted compounds act as signals to modulate underground microbe diversity and vice versa. ROOT ARCHITECTURE AND EXUDATION Plant adaptation and survival in a given environment are primarily determined by the ability of an individual to acquire resources (Aerts 1999). The root system plays a big role in acquisition of resources in a natural heterogeneous soil environment (Lynch & Brown 2001). Root system architecture (RSA) changes in nutrient-rich patches of soil such as that found under conditions of high nitrate and phosphorus (Ho et al. 2005; Paterson et al. 2006). In addition, the release of organic compounds from roots is a key factor in mineralizing acquired nutrients and in mediating plant–microbe interactions (Pierret et al. 2007). Therefore, modulating growth and root branching in regions of nutrient-rich patches may be expected to be coincident with increased root exudation that could affect the nutrient dynamics and microbial community (Paterson et al. 2006). Hence, the authors believe that it is relevant to review the information about RSA and its impact on exudation as one of the focus points of this article. Root system development is an important agronomic trait for plant growth and survival because of its role in water and nutrient uptake. RSA determines a plant’s survival in a given environment, allowing for the uptake of resources and in turn the RSA is determined by the soil environment. A wide variation in RSA of different plant species suggests that it is determined by inherent genetic factors. Basically, there are two well-known root system structures: one is typically found in dicotyledonous species (Arabidopsis, tomato, pea, etc.) and is usually comprised of a primary (tap) root and lateral roots; the second one is typically found in monocotyledonous species (rice, maize) and is characterized by the development of many adventitious roots in parallel to the primary root (Esau 1965). RSA is influenced by several biotic and abiotic factors. Thus, the RSA is a plastic trait in which even genotypically identical plants can differ depending on their macro- and micro-environment.The molecular mechanisms responsible for this plasticity are still poorly understood. A recent report demonstrated that the lateral root primordial emergence is repressed by limiting water supply, and this response is associated with abscisic acid and the LATERAL ROOT DEVELOPMENT2 gene (Deak & Malamy 2005). Apart from water, there are other exogenous abiotic factors such as nitrogen (Lopez-Bucio, Cruz-Ramirez & Herreraestrella 2003), phosphorus (Linkhor et al. 2002), iron (Moog et al. 1995) and light (Cluis, Mouchel & Hardtke 2004; Sorin et al. 2005) that modulate root branching and RSA. Plants under phosphorus starvation accumulate sugars and starch in their leaves, and this increasing load of sucrose to the phloem functions to relocate carbon resources to the roots, which increase their size relative to the shoot by initializing sugar signalling cascades that alter the RSA (Hammond & White 2008). Under suboptimal nutrient levels of phosphorus, nitrogen and iron, plant species in the family of Proteaceae develop hairy rootlets that are aggregated in longitudinal rows to form distinct clusters called proteoid © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 668 D. V. Badri & J. M. Vivanco roots. These roots solubilize the minerals and organic nutrients, and facilitate uptake of inorganic nutrients by enhanced secretion of carboxylates, phenolics and water (Lamont 2003). There are other endogenous factors, called phytohormones, that modulate root branching and RSA; these include auxin, cytokinin, ethylene, gibberellins, abscisic acid and brassinosteroids (Malamy 2005; Osmont, Sibout & Hardtke 2007). RSA is also influenced by biotic factors, because in nature plant roots are in contact with saprophytic and pathogenic microbes. For example, after infection with a specific Ralstonia solanacearum strain, the lateral root elongation in petunia plants is inhibited, but new lateral roots with abnormal morphology are induced (Zolobowska & Van Gijsegem 2006). Similarly, the rhizobacterium Bacillus megaterium also alters the RSA by inhibiting primary root growth and increasing lateral root growth in A. thaliana (Lopez-Bucio et al. 2007). RSA in some plants also changes upon symbiotic interactions such as those with N-fixing bacteria in legumes (de Billy et al. 2001; Hirsch, Lum & Downie 2001) and mycorrhizae (Hetrick 1991; Paszkowski et al. 2002). It is possible that root branching and RSA play a significant role in determining the composition of exudates both quantitatively and qualitatively. In addition, the knowledge of the involvement of particular root cells (i.e. root cap, epidermal cells and root hairs) in secretion of compounds from roots is not clearly understood. In general, the zone immediately behind the root tip is considered to be a major site of exudation (Pearson & Parkinson 1961; Schroth & Snyder 1962). However, it has been observed that the older parts of the roots also exude organic compounds, and different sites have been recorded for different plant species (Pearson & Parkinson 1961; Bowen 1968; McDougall 1968; Rovira 1969). Van Egeraat (1975) demonstrated that the root tips of the primary and lateral roots were sites of exudation by spraying ninhydrin on the filter paper where the plant roots grew. Ninhydrin is a chemical compound that interacts specifically with amine groups to produce a purple colour. Since the discovery of this compound in 1910, it has become a powerful analytical tool in the fields of chemistry, biochemistry and forensic science. Frenzel (1957, 1960) reported that different compounds were released from different parts of root system: asparagine and threonine from the meristem and root elongation zone; glutamic acid, valine, leucine and phenylalanine from root hair zone; and aspartic acid from the whole root. McDougall & Rovira (1970) used 14C-labelled compounds to identify the sites of exudation from wheat roots, and noticed that non-diffusible material released from both primary and lateral root tips, and diffusible material released from the whole length of roots. In addition,Van Egeraat (1975) found that the release of compounds was evident at the point of lateral root emergence in primary roots; this emergence leaves open wounds that are not readily healed by the plant and seep compounds into the soil. Additionally, there are a few recent reports indicating that root cap and root hair cells are involved in secretion of compounds (Pineros et al. 2002; Czarnota et al. 2003; Nguyen 2003). Generally, the apical meristem of plant roots is covered by a group of cells arranged in layers called the root cap, which sloughs off as the root tip wends its way through the soil (Barlow 1975). It has been proposed that these sloughed-off cap cells play a significant role in determining the rhizosphere ecology, and therefore the term ‘border cells’ was proposed (Hawes 1990). Border cells are involved in several functions: they decrease frictional resistance experienced by root tips (Bengough & McKenzie 1997), they regulate microbial interactions through avoidance of harmful microbes (pathogens) and favouring associations with beneficial microbes (PGPR) (Hawes 1990; Hawes et al. 1998, 2000) and they protect against heavy metal toxicity such as aluminium (Morel, Mench & Guckert 1986; Miyasaka & Hawes 2001). A mucilaginous layer has been observed on the surface of roots, particularly at the root tip, where it can form a droplet in the presence of water (Samsevitch 1965). This mucilage is secreted from the outer layers of root cap cells and has been observed in most plant species (Paull & Jones 1975; Miki, Clarke & McCully 1980; Rougier 1981). However, small drops of mucilage secretion have also been observed from root hairs or epidermal cells (Werker & Kislev 1978). This secretion may derive from the root cap or from the degradation of epidermal cell walls; alternately, it may be synthesized by rhizoplane microorganisms (Rovira, Foster & Martin 1979; Foster 1982; Vermeer & McCully 1982). Root hairs are the extensions of single epidermal cells and comprise as much as 77% of the total root surface area of cultivated crops, forming the major point of contact between the plant and the rhizosphere (Parker et al. 2000). They play a pivotal role in rhizosphere processes including anchorage and uptake of water and nutrients (Fan et al. 2001; Grierson, Parker & Kemp 2001; Michael 2001). Apart from these functions, root hair cells are involved in root secretion of compounds. Head (1964) observed spherical droplets of liquid secreted from the tips of root hairs in young apple roots growing behind a glass plate by using time-lapse cine-photomicrography. Using light, cryoscanning electron and transmission electron microscopy, it has also been observed that Sorghum species secrete sorgoleone solely from root hairs (Czarnota et al. 2003). Besides the root cap and root hair cells, other root cells are also involved in root secretion of compounds. For example, maize roots secrete citrates in response to aluminium toxicity. Using the patch clamp technique, it was determined that citrate secretion was predominant 5 cm above the root cap, and involved cortex and stellar cells (Pineros et al. 2002). In addition, comparison of citrate exudation rates in de-capped and capped roots indicated that the root cap was not playing a role in citrate secretion. Despite these findings, there is still a need to focus our attention on root cells involved in root exudation both at biochemical and molecular levels. Using ninhydrin is still considered an important tool to identify the sites of exudation on roots, but the limitation of using it is that it is able to detect only amino acids or ninhydrin-positive compounds in the exudates. Further, identification of specific root cells © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 Regulation and function of root exudates 669 involved in exudation is now possible, thanks to new molecular tools, such as the creation and study of Arabidopsis mutants impaired in the formation of specific root cells. REGULATION OF ROOT EXCRETING PHYTOCHEMICALS Roots are constantly exposed to a range of biotic and abiotic stresses at the root–soil interface, and they respond to these stresses by secreting a different blend of chemicals to protect against negative influences and encourage positive interactions. Mechanical impedance affecting root morphology (Groleau-Renaud, Plantureux & Guckert 1998), soil compaction and mild drought conditions (Brimecombe, De Leij & Lynch 2000) have been shown to result in increased root secretion. In vitro, different growth media can change the composition of root exudates of a particular plant species, thus indicating that specific nutrition plays a role in root exudation. It has also been reported that nutrient deficiency enhances exudation of certain metabolites, particularly those that increase the availability of nutrients for uptake by plant roots (Jones 1998). Environmental factors like temperature, light and soil moisture also modulate root exudation processes. For example, exudation of tannins and phenolic compounds in Vicia faba was greatly reduced at 4 °C compared to the amounts at 30 °C (Bekkara et al. 1998). Similarly, light intensity alters the exudation of secondary metabolites because of changes in photosynthesis. Watt & Evans (1999) reported that the root exudation process follows diurnal rhythms, with exudation increasing during light periods. For example, Almus glutinosa (L.) root exudates have increased flavonoid content under light conditions (Hughes et al. 1999). High soil moisture also regulates the root secretion of compounds because of the limited availability of oxygen, which leads to hypoxia. Hypoxia causes a respiration shift from aerobic to anaerobic, resulting in the accumulation of ethanol, lactic acid and alanine at phytotoxic levels (Rivoal & Hanson 1994). Xia & Roberts (1994) reported that plants escape the toxic effects of accumulated ethanol and lactic acid by secreting these metabolites from their roots. The presence (or absence) of particular minerals and toxic metals in the soil can also alter the composition of root exudates. It has been shown that plant roots secrete citric, oxalic and malic acids to detoxify aluminium in the soils, and the secretions of these organic acids are highly specific to aluminium stress. In addition, the aluminium-induced secretion pattern of organic acids varies with plant species (Ma 2000; Liao et al. 2006; Wang et al. 2006). Phosphorus deficiency also results in enhanced root secretion of phenolic compounds in certain tree and legume species, and the specificity of organic acid secretion in response to P deficiency varies with plant species (Dinkelaker, Hengeler & Marschner 1995; Chishaki & Horiguchi 1997; Dinkelaker et al. 1997; Neumann, George & Romheld 1998; Neumann & Romheld 1999). Plant species that normally co-occupy an ecological niche are likely to have developed mechanisms of detoxifying toxins produced by one another through oxidation, carbohydrate conjugation or sequestration (Inderjit & Duke 2003). Recent evidence suggests that some plant species can better withstand assault by (⫾) catechin, a potential allelotoxin produced by spotted knapweed through increased secretion of oxalic acid, which protects the roots against damage incurred by reactive oxygen species (ROS) resulting from interactions with the allelochemical (Weir et al. 2006). Exudation rates also vary with plant developmental stage and between genotypes within a single species. Seedlings produce the lowest amounts of root exudates; this gradually increases until flowering and decreases again at maturity (Aulakh et al. 2001). Garcia et al. (2001) showed that root exudation is positively correlated with root growth; it means that actively growing root systems secrete more exudates. Similarly, they observed variations in exudation patterns between genotypes of the same plant species. For example, the A. thaliana ecotypes, Col-0 and Ler, differ in the levels of malate present in their root exudates (Hoekenga et al. 2003). Both qualitative and quantitative differences were observed in the root exudation of different plant species (Cieslinski et al. 1997), and the differences are greater if they are less phyologenetically related (Fletcher & Hegde 1995). Elicitors are molecules that stimulate defence or stressinduced responses in plants. Gleba et al. (1999) postulated that chemical and physical elicitors stimulate roots of various plants to secrete phytochemicals in much higher quantities than non-elicited plants. Moreover, roots of elicited plants exude an array of compounds not detected in the exudates of non-elicited plants. In addition, a single elicitor can trigger induction of different compounds in different plant species. Exogenous application of defencesignalling molecules, such as salicylic acid (SA), methyl jasmonate (MeJA) and nitric oxide (NO) induces the accumulation of a wide range of secondary metabolites (see review of Zhao, Davis & Verpoorte 2005). Mineral deficiencies also induce the intrinsic production of elicitors that mediated signalling responses; for example, potassium deprivation induces the jasmonic acid-mediated defence responses (Schachtman & Shin 2007). It has been demonstrated that SA and MeJA are found in the medium of cultured plant cells (Parchmann, Gundlach & Mueller 1997; Chen et al. 2001), and NO is found in the apoplastic space of plant roots (Stohr & Ullrich 2002). Noritake, Kawakita & Doke (1996) reported that NO induces phytoalexin (rishitin) accumulation in potato tuber tissues as well as in the exudates. Kneer et al. (1999) showed that roots of hydroponically cultivated Lupinus luteus secrete genistein, which was induced 10-fold by SA treatment. Recent evidence suggests that treatment of Arabidopsis plant roots with SA, MeJA and NO increased the root exudation of phytochemicals compared with control plant roots (Badri et al. 2008a). The list of secondary metabolites shown to be induced by these defence-signalling molecules is shown in Table 2. © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 670 D. V. Badri & J. M. Vivanco Plants Signalling molecule Targeted secondary metabolites Arabidopsis thaliana Avena sativa Catharanthus roseus Centella asiatica Coleus blumei Cupressus lusitanica Daucus carota MeJA, SA NO JA, NO MeJA MeJA MeJA JA Echinacea pallida Eschscholtiza californica Glycine max MeJA MeJA, SA MeJA Glycyrrhiza glabra Hyoscyamus niger Hypericum perforatum Lithospermum erythrorthion Lupinus luteus Lycopersicon esculentum Oryza sativa Ocimum basilicum Panax ginseng Portulaca Rauvolfia canescens Sanguinaria canadensis Saussurea medusa Silybum marianum Solanum tuberosum Sophora flavescens Taxus chinensis MeJA MeJA, SA MeJA, SA MeJA SA MeJA MeJA MeJA MeJA, SA MeJA MeJA MeJA MeJA, SA MeJA MeJA, NO JA, NO Synthetic MeJA (HEJA) MeJA Indole glucosinolates, camalexin Avenanthramides Ajmalicine, catharanthine, vindoline Asiatocoside, triterpenes Rosmarinic acid b-Thujaplicin 6-Methoxymellein, 4-hydroxybenzoic acid Alkamides, ketoalkene/ynes Benzphenanthridines, sanguinarine Glyceollins, apigenin, daidzein, genistein, luteolin Soyasaponin Hyoscyamine, rishitin, scopolamine Hypericin Rosmarinic acid Genistein Scopoletin Momilactones, sakuranetin Rosmarinic acid, caffeic acid Ginsenoside Betacyanin Raucaffricine Sanguinarine Jaceosidin, hispidulin Silymarin Rishitin, lubimin, phytuberin Matrine Taxoid Vitis vinifera Table 2. List of secondary metabolites induced by defence signalling molecules in plantsa Stilbene, resveratrol a This list is modified and updated from Zhao et al. (2005). MeJA, methyl jasmonate; JA, jasmonic acid; SA, salicylic acid; NO, nitric oxide; HEJA, 2-hydroxyethyl jasmonate. CAN NEIGHBOURS ALTER PLANT ROOT EXUDATION PROFILES? The plant root–soil interface is an environment with high microbial inoculum, composed of both pathogenic and beneficial microbes (Rouatt & Katznelson 1960; Rouatt, Katznelson & Payne 1960). Thus, plant roots are constantly exposed to an array of microbes, and must interact and defend according to the type of biotic stress (Bais et al. 2004, 2006). How do these interactions affect root exudation? It has also been demonstrated that plants release host-specific flavonoids in response to compatible rhizobia strains (Dakora, Joseph & Phillips 1993; Pueppke et al. 1998). Recent evidence shows that microbes can modulate plant root exudation of proteins (De-la-Pena et al. 2008). The study of De-la-Pena et al. (2008) clearly demonstrated that the compositions of proteins present in the root exudates change upon the presence of a given microbial neighbour and that the exudation of proteins by a given bacterium is modulated by the presence of a specific plant neighbour. This study used two model plants, Arabidopsis and Medicago, and the microbes Pseudomonas syringae (DC3000) as a bonafide pathogen of Arabidopsis and Sinorhizobium meliloti (RM1021) as a symbiont of Medicago. The availability of the genome sequences for all four organisms allowed tracking down the origin of a given protein to determine if it was produced by a plant or by a bacterium. It was found that the interaction between Medicago–S. meliloti increased the secretion of seven plant proteins such as hydrolases, peptidases and peroxidases, but these proteins were not induced in Medicago–P. syringae interaction. Similarly, the Arabidopsis–P. syringae interaction induced the secretion of several plant defence-related proteins, but these proteins were not induced in the Arabidopsis–S. meliloti interaction. Additionally, it was found that S. meliloti secreted four proteins in high levels [superoxide dismutase, putative glycine-betain-binding ATP-binding cassette (ABC) transporter protein, outer membrane lipoprotein and hypothetical protein SMc02156] in the presence of Medicago. But in the presence of Arabidopsis, S. meliloti secreted different proteins. Similarly, P. syringae secreted a different array of proteins in the presence of Arabidopsis or Medicago. These data provide concrete evidence that both plant root and bacterial protein secretion profiles change in response to the identity of the neighbour. © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 Regulation and function of root exudates 671 At this point, it is appropriate to speculate that if plants can sense and respond to the presence of a given neighbouring microbe by altering the secretion of proteins for a particular purpose, it is then possible that plants can sense and respond to the presence of particular plant neighbours. If a microscopic organism is sensed by the roots possibly because of specific receptors, then the presence of a macroscopic neighbouring root might be better perceived by the plant. This sort of perception mechanism suggests a highly sophisticated behavioural pattern by which plants could alter their physiology and biochemistry based on the presence of a particular neighbour. The case of invasive weeds is worth highlighting as a possible example of this kind of behaviour. For the most part, invasive weeds are not invasive in their native range where they are kept in check supposedly by the presence of specialized insects and pathogens among other factors, whereas the weeds become highly invasive in the introduced range where those insects or pathogens are not present. It could be possible to infer that the native surrounding plants and the overall interaction of the weed with its native plant community might keep it in check by modulating certain physiological or biochemical parameters like favouring defence as compared to growth. When a weed arrives to a new location where those evolutionary interactions with plant neighbours are not present, its biochemistry and physiology could be altered to favour growth over defence. Further, the presence of the same species as a neighbour (monoculture) as compared to the presence of a different individual (polyculture) could be part of the response and could also alter the balance between defence and growth among other physiological characteristics. If this sort of response happens to be true, plants could be exhibiting some sort of social response. A recent paper has shown that the outcome of glucosinolates activation in Arabidopsis was regulated by the presence of neighbouring plants and found that the production of glucosinolates is increased in a high-density enviornment (large number of Arabidopsis plants grown together) compared with a low-density range (few number of Arabidopsis plants grown together) (Wentzell & Kliebenstein 2008). MECHANISM OF ROOT SECRETION In recent years, researchers have made enormous progress in analyzing the composition of root exudates and their interactions with neighbours in rhizosphere soil. However, the mechanism of secretion of root exudates is still poorly understood. The production and release of root-derived compounds are commonly constitutive, but may be induced by biotic or abiotic stress as described previously in this article. The mechanism by which plant roots secrete compounds is primarily thought to be a passive process mediated through three separate pathways: diffusion, ion channels and vesicle transport (Fig. 1) (Neumann & Romheld 2000; Bertin et al. 2003). During diffusion, small polar molecules and uncharged molecules are transported through permeability nature of lipid membranes (Sanders & Bethke 2000). This passive diffusion process depends on membrane permeability (Guern, Renaudin & Brown 1987) and cytosolic pH (Marschner 1995). Other compounds like sugars, amino acids and carboxylate anions are transported across membranes by the aid of proteins, and their direction of movement is dependent on their electrochemical gradient that allows them to pass from the cytoplasm of intact root cells (millimolar range) to the soil (micromolar range). Samuel, Fernando & Glass (1992) showed that the large cytosolic K+ diffusion potential and the extrusion of protons through ATPase generate a positively charged gradient that Cytosol ABC transporter Fatty acids/ Flavonoids Vesicular trafficking Anion channel Carbohydrates PM TMD1 NBD1 TMD2 Diffusion High-molecular weight compounds Low-molecular weight compounds Metal transporters Silica Aquaporins Uncharged molecules NBD2 2ATP 2ADP + 2Pi Apoplast Figure 1. Mechanisms of root exudation of compounds through the plant cell membrane (modified from Bertin et al. 2003). PM, plasma membrane; TMD, transmembrane domain; NBD, nucleotide-binding domain. © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 672 D. V. Badri & J. M. Vivanco promotes the release of carboxylate anions. Factors that affect membrane integrity could also promote the release of organic acids (Jones & Darrah 1995). In these cases, anion channels in the roots mediate the controlled release of these compounds (Ryan, Delhaize & Randall 1995; Zheng, Ma & Matsumoto 1998; Neumann et al. 1999; Sakaguchi et al. 1999). There are specific transporters for sugars, amino acids and metals that are involved in the secretion of specific compounds from root cells (Williams, Pittman & Hall 2000; Hussain et al. 2004; Colangelo & Guerinot 2006; Hirner et al. 2006; Hoekenga et al. 2006; Grabov 2007; Lee et al. 2007; Svennerstam et al. 2007). Plants have mechanisms of metal homeostasis to avoid excess concentrations of free metal ions (e.g. Fe, Zn, Mn and Cu); these mechanisms involve coordination of metal ion transporters for uptake, translocation and compartmentalization (see review by Hayden & Cobbett 2007). For example, graminaceous species secrete mugineic acid, a metal-binding ligand secreted from roots into the rhizosphere, and form Fe(III)-MA ligand to reduce the Fe toxicity and then enter into the root cells via a specific transporter YSL identified in maize (Curie et al. 2001; Curie & Briat 2003). Recent evidence demonstrated that the silicon efflux transporter from rice is involved in efflux of silicon from root cells (Ma et al. 2007; Ma & Yamaji 2008). Similarly, it has been demonstrated that malate transporter (AtALMT1) plays a critical role in releasing malate from root cells under aluminium toxicity in Arabidopsis (Kobayashi et al. 2007). There are other transporters like monosaccharide transporters that are involved in transporting hexoses, pentoses including ribose and polyols such as myo-inositol and glycerol (Klepek et al. 2005; Buttner 2007). Excretion of high-molecular weight compounds by roots generally involves vesicular transport (Battey & Blackbourn 1993). Knowledge of the vesicle-mediated trafficking of proteins is well understood (see review by Field, Jordan & Osbourn 2006), but the mechanism of vesiclemediated transport of phytochemicals is not fully characterized (Lin, Irani & Grotewold 2003; Grotewold 2004). There are reports demonstrating that plant defence responses are accompanied by trafficking of antimicrobial compounds to the site of pathogen infection. For example, pigmented vesicles (such as the antimicrobial flavonoids, 3-deoxyanthocyanidins) accumulate on sorghum leaves at sites of attempted fungal infection (Snyder & Nicholson 1990; Snyder et al. 1991). Similarly, the pigmented antimicrobial napthoquinones are secreted into the apoplast of the boraginaceous plant, Lithospermum erythrorhizon by a vesicle-mediated mechanism in response to fungal elicitation (Tabata 1996; Yazaki et al. 2001, 2002). However, these studies only demonstrate the vesicle-mediated transport of phytochemicals in leaf cells, and there is still no clear evidence for the mechanism of phytochemical secretion from root cells, except for Golgi-mediated transport of mucilage polysaccharides across the root cap (Neumann & Romheld 2000). ARE ABC TRANSPORTERS INVOLVED IN ROOT EXUDATION PROCESSES? Other mechanisms of transport of defence-related phytochemicals within plant cells include ABC transporters and multidrug and toxic compound extrusion (MATE) transporters (Yazaki 2005). The former mechanism involves directly energized primary transport by ATP hydrolysis, and the latter an H+ gradient-dependent secondary transport. Both groups have been implicated in transport of flavonoids to the vacuole (Yazaki 2005). ABC transporters encompass a large protein family found in all phyla (Higgins 1992), and the number of these transporters reported in Arabidopsis exceeds those reported in yeast or humans (Decottignies & Goffeau 1997; Dean, Hamon & Chimini 2001; Rea 2007). Plants are sessile and thus require many adaptive strategies to interact with the environment, and this suggests that the high number of potential chemicals produced by plants compared to other organisms may need a higher number of transporters (Dixon 2001). In bacteria, ABC transporters function as importers and exporters of compounds from the cell. In eukaryotes, recent evidence suggests that some plant ABC transporters also have import functions (Saurin, Hofung & Dassa 1999; Shitan et al. 2003; Santelia et al. 2005; Terasaka et al. 2005). Plant ABC proteins are classified into 13 subfamilies on the basis of protein size (full or half), orientation (forward or reverse), presence or absence of idiotypic transmembrane/ linker domains and overall sequence similarity (SanchezFernandez et al. 2001). ABC transporters are involved in diverse cellular processes, such as the excretion of potential toxic compounds, lipid translocation, heavy metal tolerance, nutrient transport, salt stress and in disease resistance (Balzi & Goffeau 1994; Szczypka et al. 1994; Maathuis et al. 2003; Kobae et al. 2006; Stein et al. 2006). In plants, the best-characterized ABC transporters are the full molecules belonging to three subfamilies, multidrug resistance-related protein (MRP), pleiotropic drug resistance protein (PDR) and multidrug resistance P-glycoproteins (PGP). Recent studies, using a pharmacological approach, demonstrated that root secretion of certain plant secondary metabolites is an ATP-dependent process, suggesting that ABC transporters are involved in root secretion processes (Loyola-Vargas et al. 2007;Sugiyama,Shitan &Yazaki 2007). Loyola-Vargas et al. (2007) demonstrated that Arabidopsis root secretion profiles showed differences (quantitatively and qualitatively) compared with control in presence of inhibitors like potassium cyanide, sodium orthovanadate, verapamil, nifedipine, glibenclamide and quinidine. This clearly indicates that different active transporting systems are involved in root secretion process that includes ABC transporters and P-type ATPases, because all these inhibitors used in this study would deplete the ATP pool in the cell. In addition, another study reported that the secretion of genistein, a signal flavonoid involved in rhizobium symbiosis secreted from soy bean roots, was mediated by an ABC transporter by an ATP-dependent manner, which was demonstrated by using the specific ABC transporter © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 Regulation and function of root exudates 673 inhibitor sodium orthovanadate, but the secretion of genistein was not inhibited by other inhibitors like nigericin, valinomycin and gramicidin D that inhibit plasma membrane ionophores (Sugiyama et al. 2007). The two previous evidences describe the involvement of ABC transporters in root secretion processes indirectly by using inhibitors. Recently, Badri et al. (2008b) showed direct evidence of the involvement of ABC transporters in root secretion process by using Arabidopsis knock-out mutants lacking expression of specificABC transporter proteins highly expressed in root cells (lateral root cap, epidermal cells, endodermis). This study identified three different compounds transported by distinct ABC transporters, and among those one compound was tentatively identified as 3-hydroxy-4(Z), 6(Z), 8(Z), 10(Z)-tetraenoic acid. Further, this study showed that one ABC transporter can transport structurally different compounds, or one compound could be transported by different transporters. Further investigation to identify other transporting systems (such as MATE) and their substrates involved in root secretion of phytochemicals is necessary to reap agricultural and biotechnological benefits from root exudation. NOVEL FUNCTIONS OF ROOT EXUDATES Root exudates engage in several types of interactions both positive and negative such as plant–plant, plant–microbe and tritrophic (plant–microbe–nematode) interactions in the complex rhizosphere. There is exhaustive literature available on root exudates mediating plant–plant and plant–microbe interactions (Bertin et al. 2003; Bais et al. 2004, 2006, 2008; Weir et al. 2004; Prithiviraj et al. 2007). In this review, we focus on two aspects with a few examples in the literature: tritrophic interactions and self/non-self recognition. Unlike plants and microbes, rhizospheric nematodes are highly mobile and may respond to the chemical communication that occurs between microbes and plants. Tritrophic (plant, microbe and nematode) interactions are best described in the context of research with rhizobia, mycorrhizal fungi and plant pathogens (Khan 1993; Khan et al. 2000). The outcome of these studies has shown that tritrophic interactions in the rhizosphere occur when nematodes and microbes act synergistically to influence plant growth. A different study shows that the soil-dwelling nematode Caenorhabditis elegans could also mediate interactions between roots and rhizobia in a positive way leading to increased nodulation (Horiuchi et al. 2005). This study demonstrated that the nematode acts as vector by carrying S. meliloti to the roots of the legume plant in response to plant root-released volatiles resulting in the initiation of plant–microbe symbiosis. A similar study reported the attraction of entomopathogenic nematodes to insectdamaged corn roots in a field setting (Rasmann et al. 2005). However, the knowledge of the influence of nematodes and their interactions with plant roots and root secreting compounds and microbes is very limited. An understanding of the underlying signalling in the rhizosphere related to these tritrophic interactions could greatly contribute to the improvement of more ecologically friendly agricultural practices. Another well-studied example of tritrophic interactions is the communication mediated by plant root-secreted compounds with parasitic plants and arbuscular mycorrhizal fungi (AMF). Plants use a wide range of compounds to attract beneficial organisms and deter harmful organisms. However, the attraction of beneficial organisms could also lead to abuse by malevolent organisms. The classical example of such a case is the relationship between plants, beneficial mutualistic AMF and harmful parasitic plants which is mediated by the root-secreted compound strigolactone and its derivatives (Bouwmeester et al. 2007). Strigolactones are detected in low quantities in the exudates of a range of plant species including maize, pearl millet, red clover, tomato, Lotus japonicus and Menispermum dauricum (Sugimato 2000; Sato et al. 2003; Awad et al. 2006). Numerous previously unrelated facts from past research about this complex interaction can now be integrated into a schematic way: (1) low phosphate in soil is conducive to AMF symbiosis (Smith & Read 1997); (2) phosphate starvation induces secretion of strigolactone from plant roots (Yoneyama et al. 2007); (3) strigolactones promote AMF colonization of host plants (Gomez-Roldan et al. 2007); (4) strigolactones also promote parasitic plant infection (Bouwmeester et al. 2003); and (5) AMF colonization prevents parasitic plant infection through the downregulation of germination stimulant production (Lendzemo et al. 2007). Besides those functions of strigolactones and their related compounds, two recent studies demonstrate that they are also involved in the inhibition of shoot branching (Gomez-Roldan et al. 2008; Umehara et al. 2008). There is communication between above-ground and below-ground parts of the plant to prevent or limit pathogen-causing diseases. A corresponding intra-plant signalling between roots and shoots was demonstrated in herbivory (Rasmann et al. 2005). In this study, it was demonstrated that western corn rootworm (WRC) larvae feeding on maize leaves induce the secretion of the belowground plant signal (E)-b-caryophyllene by maize roots to recruit an entomopathogenic nematode. Certain beneficial rhizobacteria activate the plant defence responses to prevent foliar diseases (Ryu et al. 2004). Similarly, plant roots secrete signalling compounds to attract symbionts such as rhizobium and AMF (Kent Peters & Long 1988; Besserer et al. 2006), but what plant root signals are involved in recruiting beneficial rhizobacteria has been poorly studied. Recent evidence (Rudrappa et al. 2008) demonstrated that l-malic acid, an intermediate of the tricarboxylic acid (TCA) cycle secreted from plant roots, is involved in recruiting the beneficial rhizobacteria Bacillus subtilis FB17 in a dose-dependent manner. This study further demonstrated that the secretion of l-malic acid from roots was enhanced by foliar inoculation of the pathogen P. syringae pv. tomato (Pst DC3000). This study should pave the way for the understanding of signals that tie above- and below-ground responses in plants. © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 674 D. V. Badri & J. M. Vivanco The below-ground competition driven by plant roots is a ubiquitous phenomenon in many natural and semi-natural types of vegetation. Several studies have confirmed that roots respond to neighbouring roots in a very specific manner that depends on the identity of the neighbour (Maina, Brown & Gersani 2002; de Kroon, Mommer & Nishiwaki 2003; Falik, de Kroon & Novoplansky 2006). Recent evidence suggests that plants can recognize kin, thus influencing competitive conditions in root interactions, allowing greater root allocation when grown with strangers rather than with siblings (Dudley & File 2007). The mechanism behind self/non-self root discrimination still remains obscure, but it has been suggested that root exudates could play a role in this response (de Kroon 2007). Ariel Novoplansky’s article in this issue, ‘Behaviour Under Competitive Conditions, Including Self Recognition’, provides more detailed information on this aspect. ARE ROOT EXUDATES INVOLVED IN CO-EVOLUTIONARY RELATIONSHIPS IN THE RHIZOSPHERE? Some of the examples given in the previous section pertain to artificial environmental or highly controlled greenhouse conditions, so the reader might ask how root exudation is playing a role in affecting or contributing to biodiversity under natural conditions. The diversity of the microbial (bacterial and fungal) communities in soil is extraordinary, and 1 g of soil could contain more than 10 billion microorganisms belonging to thousands of different species (Roselló-Mora & Amann 2001). Soil microbial populations are involved in a framework of interactions known to affect key environmental processes, like biogeochemical cycling of nutrients, plant health and soil quality (Barea et al. 2005; Giri et al. 2005). Most of the dynamic microbial interactions happen near the plant roots and in the root–soil interface, an area called the rhizosphere (Lynch 1987; Barea et al. 2005; Bais et al. 2006; Prithiviraj et al. 2007). There is variation in microbial communities in rhizosphere soil that supposedly depends on the age of the plant, crop species and soil type (Wieland, Neumann & Backhaus 2001; Buyer, Roberts & Russek-Cohen 2002; Kowalchuk et al. 2002; Hogberg, Hogberg & Myrold 2007). Most importantly, recent evidence suggests that specific plant species cultivate their own soil fungal community composition and diversity, and that this ‘cultivation’ is mediated by root secreting compounds (Broeckling et al. 2008). In this study, two model plant species (A. thaliana and Medicago truncatula) were grown in their native soil (collected in natural communities) and in the other plant’s soil under greenhouse conditions. It was found that Arabidopsis maintained its own fungal community in resident soil, but not in the non-resident soil (Medicago-grown soil). When the plants were grown in a third soil that did not support Arabidopsis or Medicago plants, the microbial communities in those soils declined dramatically as in the non-resident soil treatments. The same response was observed when root exudates were added to the soil rather than growing plants, thus indicating that plants drive these responses by the release of root exudates and that this interaction has a co-evolutionary component. Another study (Broz, Manter & Vivanco 2008) showed a similar observation but under field conditions. This study found that soils collected in Montana from high-density stands of the invasive plant spotted knapweed had significant declines in fungal community composition and diversity compared with soil collected from low-density stands of the weed. The fact that invasive weeds decrease the diversity and relative numbers of microbes (Broz et al. 2008) in the soil strengthens our belief that there is a co-evolutionary link between roots and soil microbes that is mediated by the release of root exudates. In the case of the weed, when it arrives in a new place, it simply wipes out the soil microbes in that location through the release of root exudates. These exudates could either have negative effects on the microbes (antimicrobials) or are simply not the correct carbon source required to support the growth of the microbes. It is possible that after a certain period, the weeds might start to culture a microbial community resembling those of their native range; however, this scenario is unlikely because the full ecological community of the native range would be nearly impossible to duplicate. Such a community is composed of a variety of other plants, each releasing their own root exudates and modifying the soil microbial community for the benefit of their ecosystem. When this ecosystem is affected such as by the introduction of a new plant with no co-evolutionary history, the decline in co-evolved soil microbes might affect physiological processes in some of the plants. In an invaded range, this could lead to some of the plants being made less competitive and ultimately vulnerable to the weed’s invasion. A similar correlation could be made with monocultures of crops: it could be hypothesized that such crops are drastically altering the microbial composition of a given soil. It is known that plant crops in their native habitats do not produce as much as the same crop in a non-native habitat, and this has usually been attributed to inputs (soil properties and nutrients) and weather. For instance, potatoes grow very small with low yield in the Andes of South America, its centre of origin and domestication. Based on the ideas provided earlier, one could speculate that crops in their native land are exposed to a significant number of co-evolved soil microbes, including beneficial and pathogenic ones, and that these microbes have co-evolutionary interactions that maintain an overall balance, preventing any particular species from becoming dominant. On the other hand, a crop grown in a nonnative habitat can have high yields because of the low number of soil pathogens; however, if a particular inoculum of a pathogen were to arrive to the system, it could easily flourish and become dominant because of the lack of other competing and co-evolved microbes (Fig. 2). A possible example of this situation is the potato famine of Ireland (circa 1845–1852) where Phytopthora infestans decimated most of the potato fields within a few years of being introduced. © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 Regulation and function of root exudates 675 Native habitat (a) Introduced habitat (recent) (b) Introduced habitat (old) (c) Figure 2. Co-evolutionary link of a given plant with soil microbes. A plant in its native habitat (a) survives relatively well even in the presence of diverse set of microbes, some of which are pathogenic. However, because of co-evolved interactions between rhizosphere microbes, all organisms are kept in check and none of them become dominant. When a plant is taken to a new habitat (b), it grows better than in native habitat (a) because of the lack of co-evolved pathogens present in the new habitat and could eventually decrease the diversity of the microbes in the soil. However, if a pathogen were to arrive in the new habitat (b), it could become dominant in the rhizosphere because of the lack of other co-evolved microbes to keep the potential pathogen in check, resulting in scenario (c). As a result, the plant could die (c). FURTHER LINKS BETWEEN ABOVE- AND BELOW-GROUND RESPONSES An increasing number of studies have probed the ecological significance of changes in root exudation patterns, but the clearest evidence of significance comes from the studies that seek a mechanistic basis for plant responses to herbivory. For example, the root herbivore Heterodera trifolii causes changes in the root exudation patterns of white clover, stimulating the biomass and activity of soil microbes. These positive effects subsequently create a positive feedback with the plant, ultimately benefiting plant growth in longer term (Hamilton & Frank 2001; Ayres et al. 2004). Another example is AMF that enhances plant species diversity in early successional communities (Van der Heijden et al. 1998), because they promote subordinate herb species relative to the dominant graminoids and also help to distribute the soil resources evenly, reducing the ability of certain species to monopolize resources (Van der Heijden 2004). It is obvious that the association of roots and AMF is initiated by root secreting compounds called strigolactones. Recently, several studies have determined that the rootand shoot-induced responses in different plant species show that below-ground root-induced responses to different pathogenic microbes alter several direct (production of toxins) or indirect (volatiles) defence responses in the plant above-ground that can affect the above-ground multitrophic interactions (Bezemer & Van Dam 2005). Positive and negative feedback mechanisms operate between plant and soil biota through root secreting compounds, which strongly influence rates of nutrient cycling and vegetation change. CONCLUSIONS AND FUTURE DIRECTIONS The secretion of phytochemicals and proteins from roots is an important way for plants to respond to and alter their environment. Over the last several years, research and technical advances have provided a better understanding of how root exudates mediate communication between plants and other organisms. These advances could be applied to agricultural systems to enhance production by increasing defence responses against soil-borne pathogens and/or favouring association with beneficial soil microbes. In addition, this knowledge could be applied to develop better methods of reclaiming land infested with invasive weeds, heavy metals or toxic compounds. Exciting trends are emerging from different but interconnected strands of research in the field of rhizosphere biology. However, efforts should now focus on decoding the chemical dialogues between organisms in the multifarious rhizosphere. In addition, there is a need to understand the mechanism and regulation of root exudation to better utilize phytochemical production for enhanced agricultural benefit. A major challenge for researchers is to characterize new transport systems and regulatory mechanisms involved in the root secretion process. This will lead to a greater understanding of root-secreted phytochemicals and their role in the rhizosphere. Another major challenge is absolute characterization of the chemical components of root exudates involved in favouring disease resistance and facilitating more beneficial associations with microbes in the rhizosphere. Enormous knowledge of the secondary metabolites of root exudates is currently available, but it is important to tease out the role of proteins secreted as root exudates to © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 676 D. V. Badri & J. M. Vivanco complete the scenario. Finally, there is a need for a deeper understanding of the role of root secreting compounds in determining the link between below-ground and aboveground interactions and vice versa, and also the abiotic factors that interact with biotic interactions to drive ecosystem properties. This is a major challenge that can be addressed most effectively by interdisciplinary teams of scientists: plant ecologists working with soil chemists, soil physicists, chemists and plant pathologists. In addition, it is very clear that research on root behaviour is needed to provide ecological and evolutionary data to understand multitrophic interactions and the link between belowground and above-ground diversity for the benefit of balanced ecosystem. ACKNOWLEDGMENTS The work in JMV laboratory was supported by the National Science Foundation (MCB-0542642) and US Department of Defense SERDP (SI 1388). We acknowledge the journal Plant, Cell & Environment for inviting us to write this article. Lastly, we apologize to those authors whose work could not be discussed because of the space limitation. REFERENCES Aerts R. (1999) Interspecific competition in natural plant communities: mechanisms, trade-offs and plant–soil feedbacks. Journal of Experimental Botany 50, 29–37. Aulakh M.S., Wassmann R., Bueno C., Kreuzwieser J. & Rennenberg H. (2001) Characterization of root exudates at different growth stages of ten rice (Oryza sativa L.) cultivars. Plant Biology 3, 139–148. Awad A.A., Sato D., Kusumoto D., Kamioka H., Takeuchi Y. & Yoneyama K. (2006) Characterization of strigolactones, germination stimulants for root parasitic plants Striga and Orobanche, produced by maize, millet and sorghum. Plant Growth Regulation 48, 221–227. Ayres E., Heath J., Possell M., Black H.I.J., Kerstiens G. & Bardgett R.D. (2004) Tree physiological responses to above-ground herbivory directly modify belowground processes of soil carbon and nitrogen cycling. Ecology Letters 7, 469–479. Badri D.V., Loyola-Vargas V.M., Du J., Stermitz F.R., Broeckling C.D., Iglesias-Andreu L. & Vivanco J.M. (2008a) Transcriptome analysis of Arabidopsis roots treated with signaling compunds: a focus on signal transduction, metabolic regulation and secretion. New Phytologist 179, 209–223. Badri D.V., Loyola-Vargas V.M., Broeckling C.D., et al. (2008b) Altered profile of secondary metabolites in the root exudates of Arabidopsis ATP-binding cassette transporter mutants. Plant Physiology 146, 762–771. Bais H.P., Park S.W., Weir T.L., Callaway R.M. & Vivanco J.M. (2004) How plants communicate using the underground information superhighway. Trends in Plant Science 9, 26–32. Bais H.P., Weir T.L., Perry L.G., Gilroy S. & Vivanco J.M. (2006) The role of root exudates in rhizosphere interactions with plants and other organisms. Annual Reviews of Plant Biology 57, 233–266. Bais H.P., Broeckling C.D. & Vivanco J.M. (2008) Root exudates modulate plant–microbe interactions in the rhizosphere. In Secondary Metabolites in Soil Ecology, Soil Biology 14 (ed. P. Karlovsky), pp. 241–252. Springer-Verlag, Berlin, Heidelberg, Germany. Balzi E. & Goffeau A. (1994) Genetics and biochemistry of yeast multidrug resistance. Biochimica et Biophysica Acta 1187, 152– 162. Barea J.M., Pozo M.J., Azcon R. & Azcon-Aguilar C. (2005) Microbial co-operation in the rhizosphere. Journal of Experimental Botany 56, 1761–1778. Barlow P.W. (1975) The root cap. In The Development and Function of Roots (eds J.G. Torrey & D.T. Clarkson), pp. 21–54. Academic Press, London, UK. Battey N.H. & Blackbourn H.D. (1993) The control of exocitosis in plant cells. New Phytologist 125, 307–308. Bekkara F., Jay M., Viricel M.R. & Rome S. (1998) Distribution of phenolic compounds within seed and seedlings of two Vicia faba cvs differing in their see tannin content and study of their seed and root phenolic exudations. Plant and Soil 203, 27–36. Bengough A.G. & McKenzie B.M. (1997) Sloughing of root cap cells decreases the frictional resistance to maize (Zea mays L.) root growth. Journal of Experimental Botany 48, 885–893. Bertin C., Yang X. & Weston L.A. (2003) The role of root exudates and allelochemicals in the rhizosphere. Plant and Soil 256, 67–83. Besserer A., Puech-Pagès V., Kiefer P., Gomez-Roldan V., Jauneau A., Roy S., Portais J.C., Roux C., Bécard G. & Séjalon-Delmas N. (2006) Strigolactones stimulate arbuscular mycorrhizal fungi by activating mitochondria. PLoS Biology 4, e226. Bezemer T.M. & Van Dam N.M. (2005) Aboveground– belowground interactions via plant defenses. Trends in Ecology Evolution 20, 617–624. de Billy F., Grosjean C., May S., Bennett M. & Cullimore J.V. (2001) Expression studies on AUX1-like genes in Medicago truncatula suggest that auxin is required at two steps in early nodule development. Molecular Plant–Microbe Interactions 14, 267–277. Birnbaum K., Shasha D.E., Wang J.Y., Jung J.W., Lambert G.M., Galbraith D.W. & Benfey P.N. (2003) A gene expression map of the Arabidopsis root. Science 302, 1956–1960. Boisson-Dernier A., Chabaud M., Garcia F., Becard G., Rosenberg C. & Barker D.G. (2001) Agrobacterium rhizogenes-transformed roots of Medicago truncatula for the study of nitrogen-fixing and endomycorrhizal symbiotic associations. Molecular Plant– Microbe Interactions 14, 695–700. Bouwmeester H.J., Matusova R., Zhongkui S. & Beale M.H. (2003) Secondary metabolites signaling in host–parasitic plant interactions. Current Opinion in Plant Biology 6, 358–364. Bouwmeester H.J., Roux C., Lopez-Raez J.A. & Becard G. (2007) Rhizosphere communication of plants, parasitic plants and AM fungi. Trends in Plant Science 12, 224–230. Bowen G.D. (1968) Chloride efflux along Pinus radiata roots. Nature 218, 686–687. Brimecombe M.J., De Leij F.A. & Lynch J.M. (2000) The effect of root exudates on rhizosphere microbial populations. In The Rhizosphere, Biochemistry and Organic Substances at the Soil– Plant Interface (eds R. Pinton, Z. Varini & P. Nannipieri), pp. 95–140. Marcel Dekker, New York, NY, USA. Broeckling C.D., Broz A.K., Bergelson J., Manter D.K. & Vivanco J.M. (2008) Root exudates regulate soil fungal community composition and diversity. Applied Environmental Microbiology 74, 738–744. Broz A.K., Manter D.K. & Vivanco J.M. (2008) Soil fungal abundance and diversity: another victim of the invasive plant Centaurea maculosa. The ISME Journal 1, 763–765. Bucher M. (2002) Molecular root bioengineering. In Plant Roots – The Hidden Half (eds Y. Waisel, A. Eishel & U. Kafkafi), pp. 279–294. Marcel Dekker, New York, NY, USA. Bucher M., Schroeer B., Willmitzer L. & Riesmeier J.W. (1997) Two genes encoding extensin-like proteins are predominantly expressed in tomato root hair cells. Plant Molecular Biology 35, 497–508. © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 Regulation and function of root exudates 677 Buttner M. (2007) The monossacharide transporter (-like) gene family in Arabidopsis. FEBS Letters 581, 2318–2324. Buyer J.S., Roberts D.P. & Russek-Cohen E. (2002) Soil and plant effects on microbial community structure. Canadian Journal of Microbiology 48, 955–964. Chen H.J., Hou W.C., Kuc J. & Lin Y.H. (2001) Ca2+-dependent and Ca2+-independent excretion modes of salicylic acid in tobacco cell suspension culture. Journal of Experimental Botany 52, 1219–1226. Chishaki N. & Horiguchi T. (1997) Responses of secondary metabolism in plants to nutrient deficiency. Soil Science & Plant Nutrition 43, 987–991. Choi P.S., Kim Y.D., Choi K.M., Chung H.J., Choi D.W. & Liu J.R. (2004) Plant regeneration from hairy-root cultures transformed by infection Agrobacterium rhizogenes in Catharanthus roseus. Plant Cell Reports 22, 828–831. Cieslinski G., Rees K.C.J.V., Szmigielska A.M. & Huang P.M. (1997) Low molecular weight organic acids released from roots of durum wheat and flax into sterile nutrient solutions. Journal of Plant Nutrition 20, 753–764. Cluis C.P., Mouchel C.F. & Hardtke J. (2004) The Arabidopsis transcription factor HY5 integrates light and hormone signaling pathways. The Plant Journal 38, 332–347. Colangelo E.P. & Guerinot M.L. (2006) Put the metal to the petal: metal uptake and transport throughout plants. Current Opinion in Plant Biology 9, 322–330. Conkling M., Cheng C., Yamamoto Y. & Goodman H. (1990) Isolation of transcriptionally regulated root-specific genes from tobacco. Plant Physiology 93, 1203–1211. Curie C. & Briat J.F. (2003) Iron transport and signaling in plants. Annual Review of Plant Biology 54, 183–206. Curie C., Panaviene Z., Loulergue C., Dellaporte S.L., Briat J.F. & Walker E.L. (2001) Maize yellow stripe1 encodes a membrane protein directly involved in Fe(III) uptake. Nature 409, 346– 349. Czarnota M.A., Paul R.N., Weston L.A. & Duke S.O. (2003) Anatomy of sorgoleone-secreting root hairs of Sorghum species. International Journal of Plant Sciences 164, 861–866. Dakora F.D., Joseph C.M. & Phillips D.A. (1993) Alfalfa (Medicago sativa L.) root exudates contain isoflavonoids in the presence of Rhizobium meliloti. Plant Physiology 101, 819–824. Deak K.I. & Malamy J. (2005) Osmotic regulation of root system architecture. The Plant Journal 43, 17–28. Dean M., Hamon Y. & Chimini G. (2001) The human ATP-binding cassette (ABC) transporters superfamily. Journal of Lipid Research 42, 1007–1017. Decottignies A. & Goffeau A. (1997) Complete inventory of the yeast ABC proteins. Nature Genetics 15, 137–145. De-la-Pena C., Lei Z., Watson B.S., Sumner L.W. & Vivanco J.M. (2008) Root–microbe communication through protein secretion. Journal of Biological Chemistry 283, 25247–25255. Dinkelaker B., Hengeler C. & Marschner H. (1995) Distribution and function of proteoid roots and other root clusters. Botanica Acta 108, 183–200. Dinkelaker B., Hengeler C., Neumann G., Eltrop L. & Marschner H. (1997) Root exudates and mobilization of nutrients. In Trees – Contributions to Modern Tree Physiology (eds H. Rennenberg, W. Eschrich & H. Ziegler), pp. 441–452. Backhuys Publishers, Leiden, the Netherlands. Dixon R.A. (2001) Natural products and plant disease resistance. Nature 411, 843–847. Dudley S.A. & File A.L. (2007) Kin recognition in an annual plant. Biology Letters 3, 435–438. Esau K. (1965) Plant Anatomy. Wiley, New York, NY, USA. Falik O., de Kroon H. & Novoplansky A. (2006) Physiologicallymediated self/nonself root discrimination in Trifolium repens has mixed effects on plant performance. Plant Signaling & Behavior 1, 116–121. Fan T.W.M., Lane A.N., Shenkar M., Bartley J.P., Crowley D. & Higashi R.M. (2001) Comprehensive chemical profiling of gramineous plant root exudates using high-resolution NMR and MS. Phytochemistry 57, 209–221. Field B., Jordan F. & Osbourn A. (2006) First encounters – deployment of defence-related natural products by plants. New Phytologist 172, 193–207. Fletcher J.S. & Hegde R.S. (1995) Release of phenols by perennial plant roots and their potential importance in bioremediation. Chemosphere 31, 3009–3016. Foster R.C. (1982) The fine structure of epidermal cell mucilages of roots. New Phytologist 91, 727–740. Frenzel B. (1957) Zur abgabe von aminosauren und amiden an das nahrmedium durch die wurzeln von Helianthus annus. Planta 49, 210–234. Frenzel B. (1960) Zur atiologie der anreicherung von aminosauren und amiden in wurzelraum von Helianthes annus. Ein Beitrag zur Klarung der Probleme der Rhizosphare. Planta 55, 169–207. Garcia J.A.L., Barbas C., Probanza A., Barrientos M.L. & Manero F.J.G. (2001) Low molecular weight organic acids and fatty acids in root exudates of two Lupinus cultivars at flowering and fruiting stages. Phytochemical Analysis 12, 305–311. Giri B., Giang P.H., Kumari R., Prasad R. & Varma A. (2005) Microbial diversity in soils. In Microorganisms in Soils: Roles in Genesis and Functions (eds F. Buscot & S. Varma), pp. 195–212. Springer-Verlag, Heidelberg, Germany. Gleba D., Borisjuk N.V., Borisjuk L.G., Kneer R., Poulev A., Skarzhinskaya M., Dushenkov S., Logendra S., Gleba Y.Y. & Raskin I. (1999) Use of plant roots for phytoremediation and molecular farming. Proceedings of the National Academy of Sciences of the United States of America 96, 5973–5977. Gomez-Roldan V., Roux C., Girard D., Becard G. & Puech V. (2007) Strigolactones: promising plant signals. Plant Signaling & Behavior 2, 163–164. Gomez-Roldan V., Fermas S., Brewer P.B., et al. (2008) Strigolactone inhibition of shoot branching. Nature 455, 189–195. Grabov A. (2007) Plant KT/KUP/HAK potassium transporters: a single family – multiple functions. Annals of Botany (London) 99, 1035–1041. Grierson C.S., Parker J.S. & Kemp A.C. (2001) Arabidopsis genes with roles in root hair development. Journal of Plant Nutrition and Soil Science 164, 131–140. Groleau-Renaud V., Plantureux S. & Guckert A. (1998) Influence of plant morphology on root exudation of maize subjected to mechanical impedance in hydroponic conditions. Plant and Soil 201, 231–239. Grotewold E. (2004) The challenges of moving chemicals within and out of cells: insights into the transport of plant natural products. Planta 219, 906–909. Guern J., Renaudin J.P. & Brown S.C. (1987) The compartimentation of secondary metabolites in plant cell cultures. In Cell Culture and Somatic Cell Genetics of Plants (eds F. Constabel & I.K. Vasil), pp. 43–76. Academic Press, San Diego, CA, USA. Hamilton E.W. & Frank D.A. (2001) Can plants stimulate soil microbes and their own nutrient supply? Evidence from a grazing tolerant grass. Ecology 82, 2397–2402. Hammond J.P. & White P.J. (2008) Sucrose transport in the phloem: integrating root responses to phosphorus starvation. Journal of Experimental Botany 59, 93–109. Hawes M.C. (1990) Living plant cells released from the root cap: a regulator of microbial populations in the rhizosphere. Plant and Soil 129, 19–27. Hawes M.C., Brigham L.A., Wen F., Woo H.H. & Zhu Z. (1998) © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 678 D. V. Badri & J. M. Vivanco Function of root border cells in plant health: pioneers in the rhizosphere. Annual Reviews of Phytopathology 36, 311–327. Hawes M.C., Gunawardena U., Miyasaka S. & Zhao X. (2000) The role of root border cells in plant defense. Trends in Plant Science 5, 128–133. Hayden M.J. & Cobbett C.S. (2007) Transporters of ligands for essential metal ions in plants. New Phytologist 174, 499–506. Head G.C. (1964) A study of exudation from the root hairs of apple roots by time-lapse cine-photomicrography. Annals of Botany 28, 495–498. Hetrick B.A.D. (1991) Mycorrhizas and root architecture. Experientia 47, 355–362. Higgins C.F. (1992) ABC transporters: from microorganisms to man. Annual Reviews of Cell Biology 8, 67–113. Hiltner L. (1904) Uber neure erfahrungen und probleme auf dem Gebiet der Boden-bakteriologie und unter besondere Berucksichtigung der grundungung und Bracke. Arbeiten der Deutschen Landwirtschaft Gesellschaft 98, 59–78. Hirner A., Ladwig F., Stransky H., Okumoto S., Keinath M., Harms A., Frommer W.B. & Koch W. (2006) Arabidopsis LHT1 is a high-affinity transporter for cellular amino acid uptake in both root epidermis and leaf mesophyll. The Plant Cell 18, 1931– 1946. Hirsch A.M., Lum M.R. & Downie J.A. (2001) What makes the rhizobia–legume symbiosis so special? Plant Physiology 127, 1484–1492. Ho M.D., Rosas J.C., Brown K.M. & Lynch J.P. (2005) Root architectural tradeoffs for water and phosphorus acquisition. Functional Plant Biology 32, 737–748. Hoekenga O.A., Vision T.J., Shaff J.E., Monforte A.J., Lee G.P., Howell S.H. & Kochian L.V. (2003) Identification and characterization of aluminum tolerance in Arabidopsis (Landsberg erecta ¥ Columbia) by quantitative trait locus mapping. A physiologically simple but genetically complex trait. Plant Physiology 132, 936–948. Hoekenga O.A., Maron L.G., Pineros M.A., et al. (2006) AtALMT1, which encodes a malate transporter, is identified as one of several genes critical for aluminum tolerance in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America 103, 9749–9750. Hogberg M.N., Hogberg P. & Myrold D.D. (2007) Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the tress, or all three? Oecologia 150, 590–601. Horiuchi J., Prithiviraj B., Bais H.P., Kimball B.A. & Vivanco J.M. (2005) Soil nematodes mediate positive interactions between legume plants and rhizobium bacteria. Planta 222, 848–857. Hughes M., Donnelly C., Crozier A. & Wheeler C.T. (1999) Effects of the exposure of roots Almus glutinosa to light on flavonoid and nodulation. Canadian Journal of Botany 77, 1311–1315. Hussain D., Haydon M.J., Wang Y., Wong E., Sherson S.M., Young J., Camakaris J., Harper J.F. & Cobbett C.S. (2004) P-type ATPase heavy metal transporters with roles in essential zinc homeostasis in Arabidopsis. The Plant Cell 16, 1327–1339. Hutsch B.W., Augustin J. & Merbach W. (2000) Plant rhizodeposition an important source for carbon turnover in soils. Journal of Plant Nutrition and Soil Science 165, 397–407. Inderjit & Duke S.O. (2003) Ecophysiological aspects of allelopathy. Planta 217, 529–539. Jones D.L. (1998) Organic acids in the rhizosphere – a critical review. Plant and Soil 205, 25–44. Jones D.L. & Darrah P.R. (1995) Influx and efflux of organic acids across the soil–root interface of Zea mays L. and its implications in rhizosphere C flow. Plant and Soil 173, 103–109. Karthikeyan R. & Kulakow P.A. (2003) Soil plant microbe interactions in phytoremediation. Advances in Biochemical Engineering and Biotechnology 78, 51–74. Kent Peters N. & Long S.R. (1988) Alfalfa root exudates and compounds which promote or inhibit induction of Rhizobium meliloti nodulation genes. Plant Physiology 88, 396–400. Khan A.G., Kuek C., Chaudhry T.M., Khoo C.S. & Hayes W.J. (2000) Role of plants, mycorrhizae and phytochelators in heavy metal contaminated land remediation. Chemosphere 41, 197– 207. Khan M.W. (1993) Nematode Interactions. Chapman & Hall, London, UK. Klepek Y.S., Geiger D., Stadler R., Klebl F., Landour-Arsivaud L., Lemoine R., Hedrich R. & Sauer N. (2005) Arabidopsis POLYOL TRANSPORTERS, a new member of the monosaccharide transporter-like superfamily, mediates H+-symport of numerous substrates including myo-inositol, glycerol and ribose. The Plant Cell 17, 204–218. Kneer R., Poulev A., Olesinski A. & Raskin I. (1999) Characterization of the elicitor-induced biosynthesis and secretion of genistein from roots of Lupinus luteus L. Journal of Experimental Botany 50, 1553–1559. Knudson L. (1920) The secretion of invertase by plant roots. American Journal of Botany 7, 371–379. Kobae Y., Sekino T., Yoshioka H., Nakagawa T., Martinoia E. & Maeshima M. (2006) Loss of AtPDR8, a plasma membrane ABC transporter of Arabidopsis thaliana, causes hypersensitive cell death upon pathogen infection. Plant Cell Physiology 47, 309– 318. Kobayashi Y., Hoekenga O.A., Itoh H., Nakashima M., Saito S., Shaff J.E., Maron L.G., Pineros M.A., Kochian L.V. & Koyama H. (2007) Characterization of AtALMT1 expression in aluminum-inducible malate release and its role for rhizotoxic stress tolerance in Arabidopsis. Plant Physiology 145, 843– 852. Kowalchuk G.A., Buma D.S., de Boer W., Klinkhamer P.G. & van Veen J.A. (2002) Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms. Antonie van Leeuwenhoek 81, 509–520. de Kroon H. (2007) How do roots interact? Science 318, 1562–1563. de Kroon H., Mommer L. & Nishiwaki A. (2003) Root competition: towards a mechanistic understanding. In Ecological Studies, Root Ecology (eds H. de Kroon & E.J.W. Visser), pp. 215–234. Springer-Verlag, Berlin, Heidelberg, Germany. Lamont B.B. (2003) Structure, ecology and physiology of root clusters – a review. Plant and Soil 248, 1–19. Lee M.H., Yoon E.S., Jeong J.H. & Choi Y.E. (2004) Agrobacterium rhizogenes-mediated transformation of Taraxacum platycarpum and changes of morphological characters. Plant Cell Reports 22, 822–827. Lee Y.H., Foster J., Chen J., Voll L.M., Weber A.P. & Tegeder M. (2007) AAP1 transports uncharged amino acids into roots of Arabidopsis. The Plant Journal 50, 305–319. Lendzemo V.W., Kuyper T.W., Matusova R., Bouwmeester H.J. & Ast A.V. (2007) Colonization by arbuscular mycorrhizal fungi of sorghum leads to reduced germination and subsequent attachment and emergence of Striga hermonthica. Plant Signaling & Behavior 2, 58–62. Liang P. & Pardee A.B. (1992) Differential display of eukaryotic messenger RNA by means of the polymerase chain reaction. Science 257, 967–971. Liao H., Wan H., Shaff J., Wang X., Yan X. & Kochian L.V. (2006) Phosphorus and aluminum interactions in soybean in relation to aluminum tolerance. Exudation of specific organic acids from different regions of the intact root system. Plant Physiology 141, 674–684. Limpens E., Ramos J., Franken C., Raz V., Compann B., Franssen H., Bisseling T. & Geurts R. (2004) RNA interference in Agrobacterium rhizogenes-transformed roots of Arabidopsis and © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 Regulation and function of root exudates 679 Medicago truncatula. Journal of Experimental Botany 55, 983– 992. Lin Y., Irani N.G. & Grotewold E. (2003) Sub-cellular trafficking of phytochemicals explored using auto-fluorescent compounds in maize cells. BMC Plant Biology 3, e10. Linkhor B.I., Williamson L.C., Fitter A.H. & Leyser H.M. (2002) Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis. The Plant Journal 29, 751–760. Lopez-Bucio J., Cruz-Ramirez A. & Herrera-estrella L. (2003) The role of nutrient availability in regulating root architecture. Current Opinion in Plant Biology 6, 280–287. Lopez-Bucio J., Campos-Cuevas J.C., Hernandez-Calderon E., Velasquez-Becerra C., Farias-Rodriguez R., Macias-Rodriguez L.I. & Valencia-Cantero E. (2007) Bacillus megaterium rhizobacteria promote growth and alter root-system architecture through an auxin- and ethylene-independent signaling mechanism in Arabidopsis thaliana. Molecular Plant–Microbe Interactions 20, 207–217. Loyola-Vargas V.M., Broeckling C.D., Dayakar B.V. & Vivanco J.M. (2007) Effect of transporters on the secretion of phytochemicals by the roots of Arabidopsis thaliana. Planta 225, 301–310. Lynch J.M. (1987) The Rhizosphere. Wiley Interscience, Chichester, UK. Lynch J.P. & Brown K.M. (2001) Topsoil foraging: an architectural adaptation to low phosphorus availability. Plant and Soil 237, 225–237. Lyon T.L. & Wilson J.K. (1921) Liberation of organic matter by roots of growing plants. New York Agricultural Experimental Station Bulletin 40, 1–44. Ma J.F. (2000) Role of organic acids in detoxification of aluminum in higher plants. Plant Cell Physiology 41, 383–390. Ma J.F. & Yamaji N. (2008) Functions and transport of silicon in plants. Cellular and Molecular Life Sciences 65, 3049–3057. Ma J.F., Yamaji N., Mitani N., Tamai K., Konishi S., Fujiwara T., Katsuhara M. & Yano M. (2007) An efflux transporter of silicon in rice. Nature 448, 209–212. Maathuis F.J.M., Filatov V., Herzyz P., et al. (2003) Transcriptome analysis of root transporters reveals participation of multiple gene families in the response to cation stress. The Plant Journal 35, 675–692. McDougall B.M. (1968) The exudation of C14-lebelled substances from roots of wheat seedlings. Transactions of the 9th Congress International Soil Science, Adelaide 3, 647–655. McDougall B.M. & Rovira A.D. (1970) Sites of exudation of 14Clabelled compounds from wheat roots. New Phytologist 69, 999– 1003. Maina G.G., Brown J.S. & Gersani M. (2002) Intra-plant versus inter-plant root competition in beans: avoidance, resource matching or tragedy of the commons. Plant Ecology 160, 235– 247. Malamy J.E. (2005) Intrinsic and environmental response pathways that regulate root system architecture. Plant, Cell & Environment 28, 67–77. Marschner H. (1995) Mineral Nutrition of Higher Plants. Academic Press, London, UK. Michael G. (2001) The control of root hair formation: suggested mechanisms. Journal of Plant Nutrition and Soil Science 164, 111–119. Miki N.K., Clarke K.J. & McCully M.E. (1980) A histological and histochemical comparison of the mucilages on the root tips of several grasses. Canadian Journal of Botany 58, 2581–2593. Miyasaka S.C. & Hawes M.C. (2001) Possible role of root border cells in detection and avoidance of aluminum toxicity. Plant Physiology 125, 1978–1987. Moog P.R., van der Kooij T.A., Bruggemann W., Schiefelbein J.W. & Kuiper P.J. (1995) Responses to iron deficiency in Arabidopsis thaliana: the turbo iron reductase does not depend on the formation of root hair cells and transfer cells. Planta 195, 505–513. Morel J.L., Mench M. & Guckert A. (1986) Measurement of Pb2+, Cu2+ and Cd2+ binding with mucilage exudates from maize (Zea mays L.) roots. Biological Fertility Soils 2, 29–34. Morgan J.A.W., Bending G.D. & White P.J. (2005) Biological costs and benefits to plant–microbe interactions in the rhizosphere. Journal of Experimental Botany 56, 1729–1739. Narasimhan K., Basheer C., Bajic V. & Swarup S. (2003) Enhancement of plant–microbe interactions using a rhizosphere metabolomics-driven approach and its application in the removal of polychlorinated biphenyls. Plant Physiology 132, 146–153. Neumann G. & Romheld V. (1999) Root excretion of carboxylic acids and protons in phosphorus-deficient plants. Plant and Soil 211, 121–130. Neumann G. & Romheld V. (2000) The release of root exudates as affected by the plant’s physiological status. In The Rhizosphere, Biochemistry and Organic Substances at the Soil–Plant Interface (eds R. Pinton, Z. Varanini & P. Nannipieri), pp. 41–93. Marcel Dekker, New York, NY, USA. Neumann G., George E. & Romheld V. (1998) White lupin – a model plant to study mechanisms involved in root-induced mobilization of sparingly available P-sources. In International Workshop on Role of Environmental and Biological Factors of Toxic and Essential Elements by Plants (ed. Research Institute of Pomology and Floriculture), pp. 27–33. Skierniewice, Poland. Neumann G., Massonneau A., Martinoia E. & Romheld V. (1999) Physiological adoptation to phosphorus deficiency during proteoid root development in white lupin. Planta 208, 373–382. Nguyen C. (2003) Rhizodeposition of organic C by plants: mechanisms and controls. Agronomoie 23, 375–396. Noritake T., Kawakita K. & Doke N. (1996) Nitric oxide induces phytoalexin accumulation in potato tuber tissues. Plant Cell Physiology 37, 113–116. Osmont K.S., Sibout R. & Hardtke C.S. (2007) Hidden branches: developments in root system architecture. Annual Reviews of Plant Biology 58, 93–113. Parchmann S., Gundlach H. & Mueller M.J. (1997) Induction of 12-oxo-phytodienoic acid in wounded plants and elicited plant cell cultures. Plant Physiology 115, 1057–1064. Parker J.S., Cavell A.C., Dolan L., Roberts K. & Grierson C.S. (2000) Genetic interactions during root hair morphogenesis in Arabidopsis. The Plant Cell 12, 1961–1974. Paszkowski U., Kroken S., Roux C. & Briggs S.P. (2002) Rice phosphate transporters include an evolutionarily divergent gene specifically activated in arbuscular mycorrhizal symbiosis. Proceedings of the National Academy of Sciences of the United States of America 99, 13324–13329. Paterson E., Sim A., Standing D., Dorward M. & McDonald J.S. (2006) Root exudation from Hordeum vulgare in response to localized nitrate supply. Journal of Experimental Botany 57, 2413–2420. Paull R.E. & Jones R.L. (1975) Studies on the secretion of maize root cap slime. II. Localization of slime production. Plant Physiology 56, 307–312. Pearson R. & Parkinson D. (1961) The sites of excretion of ninhydrin-positive substances by broad bean seedlings. Plant and Soil 13, 391–396. Pierret A., Doussan C., Capowiez Y., Bastardie F. & Pages L. (2007) Root functional architecture: a framework for modeling the interplay between roots and soil. Vadose Zone Journal 6, 269–281. Pineros M.A., Magalhaes J.V., Alves V.M.C. & Kochian L.V. (2002) The physiology and biophysics of an aluminum tolerance © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 680 D. V. Badri & J. M. Vivanco mechanism based on root citrate exudation in maize. Plant Physiology 129, 1194–1206. Prithiviraj B., Paschke M.W. & Vivanco J.M. (2007) Root communication: the role of root exudates. Encyclopedia of Plant and Crop Science 1, 1–4. Pueppke S.G., Bolanos-Vasquez M.C., Werner D., Bec-ferte M.-P., Prome J.-C. & Krishnan H.B. (1998) Release of flavonoids by the soybean cultivars McCall and Peking and their perception as signals by the nitrogen-fixing symbiont Sinorhizobium fredii. Plant Physiology 117, 599–606. Rasmann S., Kollner T.G., Degenhardt J., Hiltpold I., Toepfer S., Kuhlmann U., Gershenzon J. & Turlings T.C.J. (2005) Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 434, 732–737. Rea P.A. (2007) Plant ATP-binding cassette transporters. Annual Reviews of Plant Biology 58, 347–375. Rivoal J. & Hanson A.D. (1994) Metabolic control of anaerobic glycolysis – overexpression of lactate dehydrogenase in transgenic tomato roots supports the Davies–Roberts hypothesis and points to a critical role for lactate secretion. Plant Physiology 106, 1179–1185. Rodriguez I.R. & Chader G.J. (1992) A novel method for isolation of tissue-specific genes. Nucleic Acids Research 20, 3528. Roselló-Mora R. & Amann R. (2001) The species concept for prokaryotes. FEMS Microbiology Reviews 25, 39–67. Rouatt J.W. & Katznelson H. (1960) Influence of light on bacterial flora of roots. Nature 186, 659–660. Rouatt J.W., Katznelson H. & Payne T.M.B. (1960) Statistical evaluation of the rhizosphere effect. Soil Science Society of America Journal 24, 271–273. Rougier M. (1981) Secretory activity of the root cap. In Extracellular Carbohydrates, Encyclopedia of Plant Physiology, New Series, Plant Carbohydrates II (eds B.W. Tanner & F.A. Loewus), pp. 542–574. Springer-Verlag, Berlin, Germany. Rovira A.D. (1969) Plant root exudates. Botanical Reviews 35, 35–57. Rovira A.D., Foster R.C. & Martin J.K. (1979) Note on terminology: origin, nature and nomenclature of the organic materials in the rhizosphere. In The Soil Root Interface (eds J.L. Harley & R.S. Russell), pp. 1–4. Academic Press, London, UK. Rudrappa T., Czymmek K.J., Paré P.W. & Bais H.P. (2008) Rootsecreted malic acid recruits beneficial soil bacteria. Plant Physiology 148, 1547–1556. Ryan P.R., Delhaize E. & Randall P.J. (1995) Characterization of Al-stimulated efflux of malate from the apices of Al-tolerant wheat roots. Planta 196, 103–110. Ryu C.M., Farag M.A., Hu C.H., Reddy M.S., Kloepper J.W. & Pare P.W. (2004) Bacterial volatiles induce systemic resistance in Arabidopsis. Plant Physiology 134, 1–10. Sakaguchi T., Nishizawa N.K., Nakanishi H. & Yoshimura E. (1999) The role of potassium in the secretion of megineic acids family phytosiderophores from iron-deficient barley roots. Plant and Soil 215, 221–227. Samsevitch S.A. (1965) Active excretions of plant roots and their significance, translated from Fiz. Rastenii 12, 837–846. Samuel A.L., Fernando M. & Glass A.D.M. (1992) Immunofluorescent localization of plasma membrane H+-ATPase in barley roots and effects of K nutrition. Plant Physiology 99, 1509–1514. Sanchez-Fernandez R., Davies T.G.E., Coleman O.D. & Rea P.A. (2001) The Arabidopsis thaliana ABC protein superfamily, a complete inventory. Journal of Biological Chemistry 276, 30231– 30244. Sanders D. & Bethke P. (2000) Membrane transport. In Biochemistry and Molecular Biology of Plants (eds B.B. Buchanan, W. Gruisham & R.L. Jones), pp. 110–158. ASPP, Rockville, MD, USA. Santelia D., Vincenzetti V., Azzarello E., Bovet L., Fukao Y., Duchtig P., Mancuso S., Martinoia E. & Geisler M. (2005) MDR-like ABC transporter AtPGP4 is involved in auxinmediated lateral root and root hair development. FEBS Letters 579, 5399–5406. Sato D., Awad A.A., Chae S.H., Yokota T., Sugimoto Y., Takeuchi T. & Yoneyama K. (2003) Analysis of strigolactones, germination stimulants for Striga and Orobanche by high-performance liquid chromatography/tandem mass spectrometry. Journal of Agricultural Food Chemistry 51, 1162–1168. Saurin W., Hofung M. & Dassa E. (1999) Getting in or out: early segregation between importers and exporters in the evolution of ATP-binding cassette (ABC) transporters. Journal of Molecular Evolution 48, 22–24. Schachtman D.P. & Shin R. (2007) Nutrient sensing and signaling: NPKS. Annual Reviews of Plant Biology 58, 47–69. Schroth M.N. & Snyder W.C. (1962) Exudation patterns from bean seeds and hypocotyls and their effects on Fusarium solanii f. phaseoli (Abstr.). Phytopathology 52, 751. Shitan N., Bazin I., Dan K., Obata K., Kigawa K., Ueda K., Sato F., Forestier C. & Yazaki K. (2003) Involvement of CjMDR1, a plant multidrug-resistance-type ATP-binding cassette protein, in alkaloid transport in Coptis japonica. Proceedings of the National Academy of Sciences of the United States of America 100, 751– 756. Singh B.K., Millard P., Whiteley A.S. & Murrell J.C. (2004) Unravelling rhizosphere–microbial interactions: opportunities and limitations. Trends in Microbiology 12, 386–393. Smith S.E. & Read D.J. (1997) Mycorrhizal Symbiosis. Academic Publisher, London, UK. Snyder B.A. & Nicholson R.L. (1990) Synthesis of phytoalexins in sorghum as a site-specific response to fungal ingress. Science 248, 1637–1639. Snyder B.A., Leite B., Hipskind J., Butler L.G. & Nicholson R.L. (1991) Accumulation of sorghum phytoalexins induced by Colletotrichum graminicola at the infection site. Physiological and Molecular Plant Pathology 39, 463–470. Sorin C., Bussell J.D., Camus I., et al. (2005) Auxin and light control of adventitious rooting in Arabidopsis require ARGONAUTE1. The Plant Cell 17, 1343–1359. Stein M., Dittgen J., Sanchez-Rodriguez C., Hou B.H., Molina A., Schulze-Lefert P., Lipka V. & Somerville S. (2006) Arabidopsis PEN3/PDR8, an ATP binding cassette transporter, contributes to nonhost resistance to inappropriate pathogens that enter by direct penetration. The Plant Cell 18, 731–746. Stohr C. & Ullrich W.R. (2002) Generation and possible roles of NO in plant roots and their apoplastic space. Journal of Experimental Botany 53, 2293–2303. Sugimato Y. (2000) Germination stimulants for the seeds of root parasitic weeds. Journal of Pesticide Science 25, 438–440. Sugiyama A., Shitan N. & Yazaki K. (2007) Involvement of a soybean ATP-binding cassette-type transporter in the secretion of genistein, a signal flavanoid in legume–Rhizobium symbiosis. Plant Physiology 144, 2000–2008. Svennerstam H., Ganeteg U., Bellini C. & Nasholm T. (2007) Comprehensive screening of Arabidopsis mutants suggests the lysine histidine transporter 1 to be involved in plant uptake of amino acids. Plant Physiology 143, 1853–1860. Szczypka M.S., Wemmie J.A., Moye-Rowley W.S. & Thiele D.S. (1994) A yeast metal resistance protein similar to human cystic fibrosis transmembrane conductance regulator (CFTR) and multidrug resistance-associated protein. Journal of Biological Chemistry 269, 22853–22857. Tabata M. (1996) The mechanism of shikonin biosynthesis in cell Lithospermum cultures. Plant Tissue Culture Letters 13, 117– 125. © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681 Regulation and function of root exudates 681 Terasaka K., Blakeslee J.J., Titapiwatanakun B., et al. (2005) PGP4, an ATP binding cassette p-glycoprotein, catalyzes auxin transport in Arabidopsis thaliana roots. The Plant Cell 17, 2922–2939. Umehara M., Hanada A., Yoshida S., et al. (2008) Inhibition of shoot branching by new terpenoid plant hormones. Nature 455, 195–200. Uren N.C. (2000) Types, amounts and possible functions of compounds released into the rhizosphere by soil-grown plants. In The Rhizosphere, Biochemistry and Organic Substances at the Soil–Plant Interface (eds R. Pinton, Z. Varanini & P. Nannipieri), pp. 19–40. Marcel Dekker, New York, NY, USA. Van Egeraat A.W.S.M. (1975) The growth of Rhizobium leguminosarum on the root surface and in the rhizosphere of pea seedlings in relation to root exudates. Plant and Soil 42, 367–379. Van der Heijden M.G.A. (2004) Arbuscular mycorrhizal fungi as support systems for seedling establishment in grassland. Ecology Letters 7, 293–303. Van der Heijden M.G.A., Klironomos J.N., Moutoglis P., StreitwolfEngel R., Boller T., Wienken A. & Sanders R. (1998) Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396, 69–72. Vermeer J. & McCully M.E. (1982) The rhizosphere in Zea: new insight into its structure and development. Planta 156, 45–61. Wang P., Bi S., Wang S. & Ding Q. (2006) Variation of wheat root exudates under aluminum stress. Journal of Agricultural Food Chemistry 54, 10040–10046. Watt M. & Evans J.R. (1999) Linking development and determinacy with organic acid efflux from proteoid roots of white lupin grown with low phosphorus and ambient or elevated atmospheric CO2 concentration. Plant Physiology 120, 705–716. Weir T.L., Park S.W. & Vivanco J.M. (2004) Biochemical and physiological mechanisms mediated by allelochemicals. Current Opinion in Plant Biology 7, 472–479. Weir T.L., Bais H.P., Stull V.J., Callaway R.M., Thelen G.C., Bhamidi S., Stermitz F.R. & Vivanco J.M. (2006) Oxalate contributes to the resistance of Gaillardia grandiflora and Lupinus sericeus to a phytotoxin produced by Centaurea maculosa. Planta 223, 785–795. Wentzell A.M. & Kliebenstein D.J. (2008) Genotype, age, tissue, and environmental regulate the structural outcome of glucosinolate activation. Plant Physiology 147, 415–428. Werker E. & Kislev M. (1978) Mucilage on the root surface and root hairs of sorghum: heterogeneity in structure, manner of production and site of accumulation. Annals of Botany 42, 809– 816. Whipps J.M. (1990) Carbon economy. In The Rhizosphere (ed. J.M. Lynch), pp. 59–97. John Wiley & Sons Ltd, Essex, UK. Wieland G., Neumann R. & Backhaus H. (2001) Variation of microbial communities in soil, rhizosphere, and rhizoplane in response to crop species, soil type, and crop development. Applied Environmental Microbiology 67, 5849–5854. Williams L.E., Pittman J.K. & Hall J.L. (2000) Emerging mechanisms for heavy metal transport in plants. Biochimica et Biophysica Acta 1465, 104–126. Xia J.H. & Roberts J.K.M. (1994) Improved cytoplasmic pH regulation, increased lactate efflux, and reduced cytoplasmic lactate levels are biochemical traits expressed in root tips of whole maize seedlings acclimated to a low oxygen environment. Plant Physiology 105, 651–657. Yazaki K. (2005) Transporters of secondary metabolites. Current Opinion in Plant Biology 8, 301–307. Yazaki K., Matsuoka H., Shimomura K., Bechthold A. & Sato F. (2001) A novel dark-inducible protein LeDI-2 and its involvement in root-specific secondary metabolism in Lithospermum erythrorhizon. Plant Physiology 125, 1831–1841. Yazaki K., Kunihisa M., Fujisaki T. & Sato F. (2002) Geranyl diphosphate: 4-hydroxybenzoate geranyltransferase from Lithospermum erythrorhizon. Journal of Biological Chemistry 277, 6240–6246. Yoneyama K., Yoneyama K., Takeuchi Y. & Sekimoto H. (2007) Phosphorus deficiency in red clover promotes exudation of orobanchol, the signal for mycorrhizal symbionts and germination stimulant for root parasites. Planta 225, 1031–1038. Zhao J., Davis L.C. & Verpoorte R. (2005) Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnology Advances 23, 283–333. Zheng S.J., Ma J.F. & Matsumoto H. (1998) High aluminum resistance in buckwheat. I. Aluminum-induced specific secretion of oxalic acid from root tips. Plant Physiology 117, 745–751. Zolobowska L. & Van Gijsegem F. (2006) Induction of lateral root structure formation on petunia roots: a novel effect of GMI100 Ralstonia solanacearum infection impaired in Hrp mutants. Molecular Plant–Microbe Interactions 19, 597–606. Received 20 August 2008; received in revised form 3 December 2008; accepted for publication 4 December 2008 © 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 666–681