Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
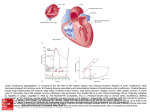
Lesson 31 Lesson Outline: • The Circulatory System Phylogenetic Trends - Heart and Aortic Arches • Chondrichthyes • Actinopterygians • Birds and Mammals Evolution of the Completely Divided Ventricle • Air-breathing Actinopterygians • Lungfish • Amphibians • Reptiles Objectives: At the end of this lesson you should be able to: Describe the phylogenetic trends that are seen in the design of vertebrate hearts and the aortic arches that arise from them. Describe the evolution of the completely divided ventricle References: Chapter 14: 314-350 Reading for Next Lesson: Chapter 14: 314-350 The Circulatory System Phylogenetic Trends - Heart and Aortic Arches (In most of the diagrams that accompany this lecture, the arterioles, venules and capillaries have been omitted for simplicity. Also note that the chambers of the heart are drawn in a straight line from posterior to anterior, again for simplicity. Finally, note that all figures are viewed looking into the body cavity with the animal lying on its back and so its right hand side is on your left, and vice versa) The hypothetical protochordate had a primitive heart and six gill arches. The heart only consists of three chambers in hagfish and early vertebrates. It has one-way valves and a simple tubular structure. The ventral aorta gives rise to the afferent branchial arteries, which enter the gills and divide down into the capillary beds. The capillaries then reanastomose into the efferent branchial arteries that join together dorsally to form the dorsal aorta. Chondrichthyes In sharks and rays, the heart now consists of four chambers, with one way valves and is twisted into an "S" shape. - the ventral portion of the first aortic arch is lost. The spiracle remains but is supplied with blood from a branch of the efferent branchial artery of the second gill arch. I II VI CA – cardiac muscle - with valves - the remaining aortic arches give rise to efferent branchial arteries that initially form paired dorsal aortae which fuse into one common dorsal aorta to supply oxygenated blood to the body. - Thus the major change is the loss of one half of the first aortic arch. This reflects the fact that the spiracle no longer contributes to gas exchange but appears to become a sensory organ (sensing water composition) - and requires oxygenated blood. Actinopterygians The hearts of bony fishes are almost identical to that of the shark. III VI The most notable difference is that the conus arteriosus is now a bulbus arteriosus (smooth muscle and not cardiac muscle and no valves). BA (smooth muscle) - no valves In most Actinopterygian fishes, the spiracle is lost and so too are both of the first two efferent branchial arteries (i.e. the first two aortic arches). Birds and Mammals The hearts of birds and mammals also have four chambers but a different four chambers than the fish heart: EC SC The sinus venosus is reduced to a small patch of tissue where the vena cavae enter the right atrium - the sinoatrial node. It retains the fastest rhythm of all the myogenic tissue and hence still serves the role as the pacemaker for the heart. IV IC III VI PA Aorta (Systemic Arch) Bird RV LV RA LA SA Node The conus or bulbus arteriosus disappears during embryonic development. It splits during development giving rise to two major arteries and these are the pulmonary artery and the aortic trunk. The two remaining chambers, the atrium and the ventricle each become divided into paired chambers. Thus there is a right and left atrium and a right and left ventricle. The venous blood from the body returns to the right atrium, passes on to the right ventricle, which pumps it to the lungs. The blood returning from the lungs in the pulmonary veins returns to the left atrium, passes to the left ventricle, which pumps it to the body. This gives rise to a dual circulation. In both birds and mammals, up to six aortic arches develop in the embryo but only three remain in the newborn animal. The VIth aortic arch develops into the pulmonary artery, the IVth aortic arch develops into the systemic arch (aorta) and the IIIrd aortic arch develops into the major vessels supplying the head. There are two major differences in this development between birds and mammals. In birds it is the right aortic arch (IV) that gives rise to the systemic arch while in mammals it is the left arch. In birds, the blood vessels to the forelimb (subclavian arteries) arise from the anterior arteries supplying the head (brachiocephalic, common carotid, internal carotid arteries). SC Mammal SC In mammals, the blood vessels to the forelimbs (subclavian arteries) arise from the dorsal aorta and left systemic arch. All of the difference in the anatomy of the heart and the aortic arches that we have described so far should make logical sense. In fishes we have a single circulation that makes sure that blood is oxygenated first and then distributed to all of the tissues. In the birds and mammals, gills are no longer present and we now have a dual system where blood goes to the lungs to be oxygenated and then returns to the other side of the heart to be pumped to the tissues. We have in essence two pumps now (that work synchronously) but contained within the same structure. There are two questions that we need to consider next: How did we get from the fish system to the bird/mammalian system? In the process (i.e. in the lungfish, amphibians and reptiles) we see a variety of partially divided hearts. Why have these evolved? Why aren't all of these other hearts completely divided too? Evolution of the Completely Divided Ventricle Air-Breathing Actinopterygians In those fish with accessory air-breathing organs, blood supply to the organ arises from either the dorsal aorta, or from the efferent branch of the VIth branchial artery. III either/or VI Oxygenated blood leaving these organs enters the general venous circulation and mixes with deoxygenated venous blood returning to the heart. Lungfish With lungfish, we begin to see the start of major change. II – reduced, has gill As with the teleosts, the first afferent branchial artery fails to develop and the second aortic arch is reduced (but present). Aortic arches III to VI do develop. In the Australian lungfish (Neoceratodus) they all deliver blood to fully functional gills. In the African lungfish (Protopterus), however, the III and IVth arches are intact but there are no gills associated with these arches. ductus arteriosus VI Finally, in all lungfish, the VIth arch gives rise to a branch that services the lungs. It also maintains a connection to the dorsal aorta via the ductus arteriosus. This should give rise to a very inefficient system. While blood entering the dorsal aorta from aortic arches II, V and VI will carry oxygenated blood from the gills, it appears as though blood from aortic arches III and IV can also enter the dorsal aorta carrying deoxygenated blood straight from the heart. This is misleading, however as we will see when we look at the heart. There are several changes to the lungfish heart: - In all three genera of lungfish, the single atrium is partially divided by an interatrial septum into a larger right and smaller left atrial chamber. - In the African and South American lungfish, the blood returning from the lungs enters into this smaller left atrial chamber while blood returning from the body, via the sinus venosus, enters into the larger right atrial chamber. - In place of the atrioventricular valve is an atrioventricular plug which functions very much like the atrioventricular valve in other vertebrates. - The ventricle is also partially divided by an interventricular septum and the conus arteriosus contains a spiral valve. The spiral valve divides the aorta into two separate channels. The net result of these partial divisions is to create internal channels within the heart. Oxygenated blood returning from the lungs is kept relatively separate from deoxygenated blood returning from the body. It passes through the left atrium, the left side of the ventricle and enters one side of the spiral valve in the conus arteriosus and emerges to enter into the anterior gill arches. Thus the blood that passes through arches 3 and 4, which have no functional gills, is already oxygenated. Deoxygenated blood enters the sinus venosus, the right atrium, the right side of the ventricle and the opposite side of the spiral valve that delivers it to the posterior two gill arches. This blood then passes through the functional gills and can then either enter the dorsal aorta or be diverted to the lungs. When oxygen levels in water are high, most of the blood passes from the gills into the dorsal aorta but when the levels are low, much of this blood bypasses the capillaries in the gills and is then shunted to the lungs and not into the dorsal aorta, as a result of constriction of the ductus arteriosus. While this explains why it is not deleterious to not have gills on arches 3 and 4, it does not really explain why this is an advantage. It is an advantage because it prevents the loss of oxygen from the body into the water under conditions when O2 levels in water are low. At this time, the blood returning from the lungs contains more O2 than the water and if these arches had gills with a good exchange surface, O2 obtained from the lungs would be lost to the water before it could be delivered to the body. (The reasons for the retention of the 2nd aortic arch are not clear but again, this arch may serve a chemosensory function). This story had been a generalized one and primarily fits the situation in the African lungfish. The Australian lungfish is not as highly evolved, while the South American lungfish is even more highly evolved. Amphibians The situation we find in amphibians is slightly different between groups (anurans versus urodeles) as well as within groups. For simplicity we will consider only the general adult anuran scheme. (Remember that the larvae (i.e. tadpoles) are aquatic and have gills). Note: no connection III IV cutaneous VI As with the teleosts and lungfishes, the first afferent branchial artery fails to develop. In the Adult anurans, the second and fifth aortic arches also fail to develop. Aortic arches III, IV and VI do develop. The IIIrd aortic arch now only supplies the head (it is not connected to the systemic arch) and the IVth aortic arch becomes the systemic arch. Finally, the VIth arch primarily services the lungs although it does have a branch that still supplies part of the systemic circulation - the skin (the pulmocutaneous artery). It does not maintain a connection to the dorsal aorta (the ductus arteriosus disappears during metamorphosis). All arches are bilateral. The heart is also different from that of the lungfish. The atrium is now completely divided into a right and left atrium. The ventricle is completely undivided with no septum. It is, however, very trabeculate (spongy). The conus arteriosus has a spiral valve just like the lungfish. bilateral systemic arches Now, oxygenated blood returning from the lungs is kept completely separate from deoxygenated blood returning from the body in the atria. While there is the opportunity for blood to mix within the ventricle, this is largely prevented by the spongy nature of the cavity, which limits (but does not prevent) mixing. The blood on the two sides of the ventricle then enters different sides of the spiral valve in the conus arteriosus. The oxygenated blood emerges to enter into the anterior aortic arches and be distributed to the body, while the deoxygenated blood enters the VIth aortic arch to go to either the lungs and/or skin to be oxygenated. Note that blood returning from the two sites will not return to the same place. To allow this to occur, it is essential that the ventricle not be divided - otherwise, not enough blood would return to the left side of the heart to be pumped to the body. Reptiles The primary difference seen in the aortic arches of reptiles is that the ventral aorta divides into three channels embryonically. III LIV RIV As a result, the base of the VIth aortic arch retreats and now arises separately from the ventricle. VI The base of the IVth aortic arch also retreats and now the left branch of the IVth arch arises from the ventricle independent of the right branch of the IVth arch, which arises along with the remainder of the ventral aorta, and the IIIrd arches. Thus there is one pulmonary circuit and two systemic circuits (one of which gives rise to all the anterior arteries) that arise independently from the ventricle. (Note that this is much akin to the situation seen in birds (right arch predominates - and unlike that seen in mammals (left arch predominates)). The hearts in reptiles all possess both right and left atria but retain varying degrees of incomplete separation of the ventricles. We don't have time to go into these in detail other than to point out that the hearts of the chelonia, as well as those of the snakes and lizards (squamates) are incompletely divided within the ventricle while the hearts of the crocodilians have a completely divided ventricle but, the left aorta arises from the right ventricle along with the pulmonary artery and has a connection just outside the heart to the right aorta. All reptile hearts are capable of functionally separating oxygenated from deoxygenated blood but retain the ability to mix blood from the two circuits. The functional separation is critically important for it allows the two sides of the heart to develop differently. The left side provides blood to the entire body. As animals increase their levels of activity, they also increase the blood supply to their tissues. The rise in the number of capillary beds leads to a rise in resistance to blood flow and requires that the heart pump harder to push the blood through all the tubes. It needs to be large and strong. The right side of the heart, on the other hand, only supplies the lungs and the resistance of this circuit is low. Indeed, if blood pressure in this circuit rises, fluid exudes out of the capillaries into the tissues (pulmonary edema). The pressure to this circuit must be kept low. Thus there is a conflict and the only solution is to separate the two sides of the heart. So why not separate them anatomically as well as functionally? This is a question that has tantalized anatomists and physiologists for over a century and the answer is still not clear. Almost all textbooks tell us that the reason for retaining the ability to shunt blood between the two circuits has to do with diving and the ability to redistribute blood when animals are unable to breathe. This is not a very satisfying explanation since most of the squamate reptiles do not dive. It also does not conserve O2 or reduce the work of the heart - two common explanations given for the presence of these shunts. It is clear that the amount of shunt (mixing between the two sides) is reduced whenever the demand for O2 for the tissues increases. It is not clear why mixing occurs when the demand is reduced. One current theory is that it allows some oxygenated blood returning from the lungs to reach the inside of the ventricle, which would otherwise only receive deoxygenated blood, which is a good thing. When the animals become active, however, and the heart must generate high levels of flow to the tissues, the shunt must be reduced.