Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
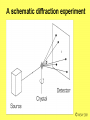
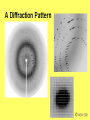
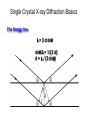
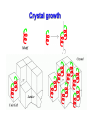
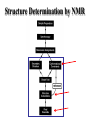
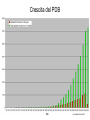
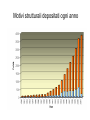
Metodi per la determinazione della struttura di biomolecole Che informazioni offre la struttura? • • • • • • Conformazione dei siti attivi e di legame Orientazione dei residui conservati Interpretazione di meccanismi Visualizzazione di cavità Calcolo di potenziale elettrostatico … The paper The model Maurice Wilkins M.H.F. Wilkins, A.R. Stokes, H.R. Wilson: Molecular Structure of Deoxypentose Nucleic Acids. Nature 171, 738 (1953) Rosalind Franklin R.E. Franklin and R.G. Gosling Molecular Configuration in Sodium Thymonucleate, Nature 171, 740 (1953) Nobel laureates 1962 Wilkins, Perutz, Crick, Steinbeck, Watson, Kendrew Cristallografia a raggi X • Ottenere cristalli della proteina – 0.3-1.0 mm – Le singole molecole sono ordinate in modo periodico, ripetitivo. • La struttura è determinata dai dati di diffrazione. Diffrazione di raggi X Schmid, M. Trends in Microbiology, 10:s27-s31. X-ray emission Instrumentation or synchrotron X-rays Instrumentation SR beam Instrumentation ESRF - Grenoble Instrumentation Injection process Instrumentation SR tunnel Instrumentation Focusing magnet Instrumentation Front - end Instrumentation Beamline Single Crystal X-ray Diffraction Basics The Bragg law l = 2 d sinq sinq/l = 1/(2 d) d = l / (2 sinq) Single Crystal X-ray Diffraction Basics The Bragg law l = 2 d sinq sinq/l = 1/(2 d) d = l / (2 sinq) Single Crystal X-ray Diffraction Basics The Bragg law l = 2 d sinq sinq/l = 1/(2 d) d = l / (2 sinq) Cristallografia a raggi X • Le proteine devono cristallizzare – Grande quantità – Solubili • Accesso a radiazione adatta • Tempo di calcolo per risolvere la struttura Forza Ionica Esercizio: calcolare la differenza di forza ionica tra le soluzioni: 0.01 M NaCl 0.01 M CaCl2 0.01 M CaSO4 Used for crystallization experiments Le solubilità della proteina è determinata dalle interazioni soluto solvente dovute alle cariche superficiali della proteina Lys, Arg, Asp, Glu L’aumento della forza ionica della soluzione favorisce la interazione tra le cariche superficiali della proteina ed il solvente polare Un eccesso di forza ionica della soluzione favorisce le solvatazione degli ioni stessi a scapito della solvatazione della proteina. Il sale compete con la proteina e la solvatazione diminuisce all’aumentare della forza ionica Crystal growth Crystal growth Crystal growth Supersaturation Nucleation + growth Supersolubility curve Solubility curve Growth only Unsaturation Protein concentration Risoluzione Campo magnetico NMR NOE (Nuclear Overhauser Effect) Il campo magnetico Magnete a 900 MHz Magnete a 400 MHz Costo commerciale di uno spettrometro a 900 MHz: ca. 5 M€ n. di spettrometri 900 MHz operativi al mondo < 10 The Magnet History First magnets were built using ferromagnetic material= permanent magnet Then Electromagnets: i.e. field was generated by wiring of conducting material Now: cyomagnets: i.e. electromagnets made of superconducting wire. A “cutted” magnet L’energia della E= -m•B 0 transizione NMR DE=(h/2p)B0 NUCLEUS m=-1/2 DE=h0 • Sensitivity E m=+1/2 B0 B0 MAGNETIC FIELD Larmor Frequency I due livelli energetici sono Il campo magnetico B0 serve per creare la degeneri se B0=0 separazione di energia tra i 2 livelli L’energia della E= -m•B 0 transizione NMR DE=(h/2p)B0 Devo applicare una radiofrequenza alla frequenza DE=h0 Per stimolare la transizione! m=-1/2 DE=h0 E m=+1/2 B0 Il campo di radiofrequenza utilizzato per eccitare la transizione si chiama anche B1 Larmor Frequency B1 deve emettere una radiofrequenza in corrispondenza della frequenza di Larmor Some paradigmatic examples Why? NMR is a unique method to obtain information in solution AT THE ATOMIC LEVEL. Each individual atom has a peculiar resonance frequency. Because the resonance frequency depends on the environment of the atom, EACH atom has a different resonance frequency and can, therefore be identified CH3-CH2-OH equivalent Sensibilità dell’Esperimento NMR S/N ≈ N 5/2 B3/2 N = Numero di spins che contribuiscono al segnale rapporto giromagnetico del nuclide studiato B Camp magnetico utilizizzato NMR Spectrum to 3D Structure H H Interactions Spectrum H H H H H H H H H Structure Challenges For Determining Protein Structures Using NMR • Proteins have thousands of signals • Assign the specific signal for each atom • Thousands of interactions between atomsalso need to be assigned • Need to transform from NMR spectrum through interpretation of scalar and dipolar interactions to generate 3D coordinates Resonance Assignment CH3-CH2-OH OH CH2 CH3 Which signal from which H atoms? The key attribute: use the scalar and dipolar couplings to match the set of signals with the molecular structure Proteins Have Many Signals 1H NMR Spectrum of Ubiquitin ~500 resonances A large number of signals are overlapped A Critical Feature of Protein NMR Spectra • Only some nuclei are coupled Each amino acid gives rise to an independent NMR sub-spectrum, which is much simpler than the complete protein spectrum Methods have been developed to extract each sub-spectrum from the whole Basic Strategy to Assign Resonances in a Protein 1. Identify resonances for each amino acid T L S G S R G 2. Put amino acids in order - Sequential assignment (R-G-S,T-L-G-S) - Sequence-specific assignment 1 2 3 4 5 6 7 R-G-S-T-L-G-S Critical Features of Protein NMR Spectra • The nuclei are not all mutually coupled • Regions of the spectrum correspond to different parts of the amino acid • Tertiary structure leads to increased dispersion of resonances Regions of the 1H NMR Spectrum are Further Dispersed by the 3D Fold What would the unfolded protein look like? Proteins Have Overlapped Signals 1H NMR Spectrum of Ubiquitin Resolve resonances by multi-dimensional experiments Resolve Peaks By Multi-D NMR A BONUSregions in 2D spectra provide protein fingerprints If 2D cross peaks overlap go to 3D or 4D ….. Solution to the Protein Challenge 1. Increase dimensionality of spectra to better resolve signals: 1234 2. Detect signals from heteronuclei (13C,15N) t2 t1 t3 Heteronuclear nD NMR 1. Increase dimensionality of spectra to better resolve signals: 1234 2. Detect signals from heteronuclei (13C,15N) Better resolution of signals/chemical shifts not correlated between nuclei More information to identify signals Lower sensitivity to MW of protein Structure Determination by NMR NMR Experimental Observables Providing Structural Information • Backbone conformation from chemical shifts (Chemical Shift Index- CSI) • Distance constraints from NOEs • Hydrogen bond constraints • Backbone and side chain dihedral angle constraints from scalar couplings • Orientation constraints from residual dipolar couplings 1H-1H Distances From NOEs Long-range (tertiary structure) Sequential Intraresidue A B C D •••• Z Medium-range (helices) Challenge is to assign all peaks in NOESY spectra Determining Protein Fold Before Structure Calculations 1. Determine secondary structure •CSI directly from assignments •Medium-range NOEs 2. Add key long-range NOEs to fold Approaches to Identifying NOEs • 1H-1H (homonuclear) 2D 3D • 15N- or 13C-dispersed 3D (heteronuclear) 4D 1H 1H 1H 1H 1H Identifying Unique NOEs • Filtered, edited NOE: based on selection of NOEs from two molecules with unique labeling patterns. Labeled protein Unlabeled peptide Only NOEs at the interface • Transferred NOE: H H based on: 1) faster build-up H kon of NOEs in large versus small molecules; 2) signal koff H of free state when in excess and exchanging quickly Only NOEs from bound state Hydrogen Bonds C=O H-N • NH chemical shift to low field • Slow rate of NH exchange with solvent • Characteristic pattern of NOEs • Scalar couplings across the H-bond When H-bonding atoms are known can impose a series of distance/angle constraints to enforce standard H-bond geometries Angles From Scalar (J) Couplings Must accommodate multiple solutions multiple J values Orientational Constraints From Dipolar (D) Couplings Ho Reports angle of internuclear vector relative to magnetic field Ho F2 F3 F1 Must accommodate multiple solutions multiple orientations NMR Structure Calculations • Objective is to determine all conformations consistent with the experimental observables • In contrast to X-ray crystallography, NMR observes the atoms indirectly • NMR data is not perfect: noise, incomplete Multiple solutions: caused by uncertainties in the experimental constraints Conformational Ensemble Representing an NMR Structure C N Precision: RMSD of the ensemble NMR Structure Calculations: Two Primary Approaches 1. Start with polymer and fold Distance Geometry Hybrid Simulated Annealing 2. Start with all conformations and restrict Systematic Grid Search Filters NMR Structures: The Challenges • Distances alone are not sufficient • Interiors good, but surfaces can be poor because of few distances/high dynamics- variable resolution • Poor electrostatics – Simplified treatment (scaled charges) – No explicit solvent, dielectric Variable Resolution Interiors well defined, surfaces more variable Regular secondary structures well defined, loops more variable Backbone and side chain trends are similar Stereopairs Linewidth is Dependent on MW A B A 15N B Linewidth determined by size of particle 15N 15N Fragments have narrower linewidths 1H 1H 1H Dynamics/Constraints Regions with higher flexibility will exhibit fewer H-H NOEs Dynamics/Uncertainty in Structures Weak correlation Strong correlation High uncertainty MAY be correlated with a high degree of flexibility Risonanza Magnetica Nucleare (NMR) • • • • • • Proteine in soluzione Limite di dimensione ~ 40 kDa Proteine stabili a lungo Marcatura con 15N, 13C, 2H. Strumentazione molto costosa Tempo per assegnare le risonanze Pro e contro X-ray NMR • Richiede cristalli, problematico • • Non ha limiti (teorici) di grandezza Possibile in soluzione, più semplice • Limitato a proteine fino a circa 300 residui • Piú preciso • Meno preciso • Risoluzione • Numero di vincoli • Struttura può essere deformata dai cristalli, rigida • Struttura nativa in soluzione, flessibile • Una “soluzione“ • Molti modelli X-ray NMR Protein Data Bank (PDB) • URL: http://www.rcsb.org/pdb/ • Coordinate 3-D di strutture proteiche • Formato unico • Tutte le strutture risolte con i raggi X e NMR • Più vecchia della maggior parte degli altri database • Strutturata male a causa dello sviluppo storico Il Protein Data Bank Crescita del PDB Motivi strutturali depositati ogni anno Percentuale di nuovi motivi strutturali Formato PDB I HEADER COMPND COMPND SOURCE SOURCE AUTHOR AUTHOR ... REMARK REMARK REMARK REMARK REMARK REMARK REMARK REMARK REMARK ... SEQRES SEQRES SEQRES ... ONCOGENE PROTEIN 06-JUN-91 121P H-RAS P21 PROTEIN COMPLEX WITH GUANOSINE-5'-[B,G-METHYLENE] 2 TRIPHOSPHATE HUMAN (HOMO SAPIENS) CELLULAR HARVEY-RAS GENE TRUNCATED AND 2 EXPRESSED IN (ESCHERICHIA COLI) U.KRENGEL,K.SCHEFFZEK,A.SCHERER,W.KABSCH,A.WITTINGHOFER, 2 E.F.PAI 121P 121P 121P 121P 121P 121P 121P 2 3 4 5 6 7 8 1 1 REFERENCE 1 1 AUTH U.KRENGEL,I.SCHLICHTING,A.SCHEIDIG,M.FRECH,J.JOHN, 1 AUTH 2 A.LAUTWEIN,F.WITTINGHOFER,W.KABSCH,E.F.PAI 1 TITL THE THREE-DIMENSIONAL STRUCTURE OF P21 IN THE 1 TITL 2 CATALYTICALLY ACTIVE CONFORMATION AND ANALYSIS OF 1 TITL 3 ONCOGENIC MUTANTS 1 REF NATO ASI SER.,SER.A V. 220 183 1991 1 REFN ASTM NALSDJ US ISSN 0161-0449 2002 121P 121P 121P 121P 121P 121P 121P 121P 121P 17 18 19 20 21 22 23 24 25 1 2 3 121P 121P 121P 56 57 58 166 166 166 MET THR GLU TYR LYS LEU VAL VAL VAL GLY ALA GLY GLY VAL GLY LYS SER ALA LEU THR ILE GLN LEU ILE GLN ASN HIS PHE VAL ASP GLU TYR ASP PRO THR ILE GLU ASP SER Formato PDB II HELIX HELIX ... SHEET SHEET SHEET ... TURN TURN TURN ... ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ... HETATM HETATM HETATM HETATM ... 1 A1 2 A2 LYS SER 1 S 2 S 3 S 6 GLU 6 GLU 6 THR 1 T1 2 T2 3 T3 ALA ILE ALA 1 2 3 4 5 6 7 8 9 10 11 1324 1325 1326 1327 16 65 GLN THR 37 49 2 11 46 83 25 74 ILE THR VAL VAL GLU ASN 1 1 46 0 58 -1 9 1 O N LEU LEU 53 6 N O LYS ASP 42 54 14 49 86 121P 121P 80 81 121P 121P 121P 85 86 87 121P 121P 121P 91 92 93 104 105 106 107 108 109 110 111 112 113 114 N CA C O CB CG SD CE N CA C MET MET MET MET MET MET MET MET THR THR THR 1 1 1 1 1 1 1 1 2 2 2 -7.176 -5.913 -5.903 -6.703 -4.712 -4.594 -3.193 -4.325 -4.966 -4.759 -4.312 32.630 31.928 30.860 30.881 32.869 33.420 34.558 35.886 29.934 28.930 29.597 -6.655 -6.676 -5.600 -4.654 -6.415 -4.990 -4.899 -4.618 -5.760 -4.751 -3.441 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 14.06 17.27 16.41 16.12 17.94 19.41 21.82 23.68 15.08 16.71 16.63 121P 121P 121P 121P 121P 121P 121P 121P 121P 121P 121P PG O1G O2G O3G GTO GTO GTO GTO 167 167 167 167 5.150 4.768 4.164 4.834 32.173 32.597 32.683 30.641 22.030 23.390 21.069 22.025 1.00 1.00 1.00 1.00 11.69 13.29 12.61 13.18 121P1427 121P1428 121P1429 121P1430 X Y Z B-factor ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM ATOM 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 CA C O N CA C O CB OG N CA C O CB OG1 CG2 GLU GLU GLU SER SER SER SER SER SER THR THR THR THR THR THR THR 225 225 225 226 226 226 226 226 226 227 227 227 227 227 227 227 -0.900 -0.185 -0.514 0.788 1.534 2.231 1.883 2.572 3.237 3.242 3.989 4.274 4.179 5.354 5.114 6.256 -1.002 0.146 1.329 -0.203 0.805 1.806 1.952 0.130 -0.941 2.478 3.417 2.705 3.296 3.797 4.682 4.492 39.233 39.970 39.758 40.823 41.594 40.681 39.514 42.515 41.848 41.223 40.410 39.080 38.022 41.074 42.172 40.065 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00 70.00 70.00 70.00 70.00 70.00 68.89 70.00 70.00 70.00 65.51 70.00 56.25 44.63 70.00 70.00 70.00 1HXN 1HXN 1HXN 1HXN 1HXN 1HXN 1HXN 1HXN 1HXN 1HXN 1HXN 1HXN 1HXN 1HXN 1HXN 1HXN 170 171 172 173 174 175 176 177 178 179 180 181 182 183 184 185