Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
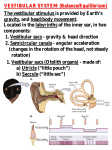
NIH Public Access Author Manuscript Curr Opin Otolaryngol Head Neck Surg. Author manuscript; available in PMC 2012 October 01. NIH-PA Author Manuscript Published in final edited form as: Curr Opin Otolaryngol Head Neck Surg. 2011 October ; 19(5): 376–381. doi:10.1097/MOO. 0b013e32834a5be1. EFFERENT MODULATION OF HAIR CELL FUNCTION RICHARD D. RABBITT1,2 and WILLIAM E. BROWNELL3 1Department of Bioengineering, University of Utah, Salt Lake City, UT, USA 2Marine Biological Laboratory, Woods Hole, MA, USA 3Department of Otolaryngology, Head and Neck Surgery, Baylor College of Medicine, Houston, TX, USA Abstract Purpose of review—This review covers papers published between 2010 and early 2011 that presented new findings on inner ear-efferents and their ability to modulate hair cell function. NIH-PA Author Manuscript Recent findings—Studies published within the review period have increased our understanding of efferent mechanisms on hair cells in the cochlear and vestibular sensory epithelium and provide insights on efferent contributions to the plasticity of bilateral auditory processing. The central nervous system controls the sensitivity of hair cells to physiological stimuli by regulating the gain of hair cell electromechanical amplification and modulating the efficiency of hair cell-8th nerve transmission. A notable advance in the past year has been animal and human studies that have examined the contribution of the olivocochlear efferents to sound localization particularly in a noisy environment. Summary—Acoustic activation of olivocochlear fibers provides a clinical test for the integrity of the peripheral auditory system and has provided new understanding about the function and limitations of the cochlear amplifier. While similar tests may be possible in the efferent vestibular system they have not yet been developed. The structural and functional similarities of the sensory epithelia in the inner ear offer hope that testing procedures may be developed that will allow reliable testing of the vestibular hair cell function. Keywords NIH-PA Author Manuscript olivocochlear; hearing; efferent vestibular system; balance; descending control; bilateral integration INNER EAR EFFERENTS INTRODUCTION This review covers papers that enhance our understanding of how the brain modulates inner ear mechanoreception. Hair cell activity can be modulated by altering the magnitude of the mechanical stimulus that reaches the inner ear either by activation of middle ear muscles to attenuate sound or by changing the position or angular acceleration of the head with skeletal muscles. Inner ear hair cells differ from most sensory cells in receiving efferent innervation Correspondence to William E. Brownell, Oto-HNS, Baylor College of Medicine, One Baylor Plaza, Houston, TX 77030. [email protected]; tel: 713-798-8540; fax: 713-798-8553.. This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. RABBITT and BROWNELL Page 2 NIH-PA Author Manuscript from neurons located in the brain. There are two innervation patterns, one is directly onto the hair cell body and the other is onto eighth nerve dendrites. The former has been called presynaptic and the latter postsynaptic in relation to their presumed impact on the neurotransmission at the hair cell-eighth nerve synapse. Presynaptic efferent activity can affect neurotransmitter release but it can also modulate the gain of mechanical force generation by the hair cell. Two types of hair cell electromechanical force generation have been identified; one is somatic outer hair cell (OHC) electromotility [1, 2] and the other is a stereocilia bundle motor [3, 4]. Medial olivocochlear OC (MOC) efferents are myelinated fibers that innervate OHCs in mature animals. They modulate cochlear mechanics and otoacoustic emissions (OAE, which include: distortion product, DPOAE; spontaneous, SOAE; click evoked, COAE; etc). Lateral OC (LOC) efferents innervate auditory-nerve fibers under inner hair cells (IHCs) and directly modulate the firing of the auditory eighth nerve fibers. There is a direct efferent innervation onto Type II hair cells in the vestibular sensory epithelia. Calyceal afferent endings interfere with a somatic contact onto Type I hair cells but a single efferent can innervate both the Type II cell body presynaptically and a calyceal terminal postsynaptically. The MOC and LOC cell bodies of origin and the anatomical trajectory of their axons to the inner ear have been established, but efferent vestibular system (EVS) neurons are scattered in clusters that do not divide cleanly into two systems. NIH-PA Author Manuscript This review builds on an excellent review of auditory olivocochlear efferents that appeared last year in this journal [5]. We identify new findings and encourage the reader to look to last year's review for background. The past year has seen a new review of the EVS [6] that provides a context for several new fundamental findings including the first observation of efferent modulation of hair cell force production in the semicircular canals [4]. Another excellent review of physiological principles underlying efferent inhibition in both vestibular and auditory hair cells appeared this year [7]*. Additional background information may also be found in a new book entitled “Auditory and Vestibular Efferents” that has just been published [8]. NIH-PA Author Manuscript MOC activation alters outer hair cell electromotility—We have previously shown how current shunting caused by MOC efferent activity in a piezoelectric model of the OHC can result in reduced electromotility and thereby alter cochlear amplification [9]. The model results were consistent with and improved on earlier models that offered an explanation for the suppression of cochlear amplification observed in physiological, behavioral and human experiments [5]. The past year has seen a number of experimental studies that confirm and extend the fundamental finding that MOC activity alters cochlear amplification by acting on OHC electromotility. Behavioral studies with chinchillas demonstrated a reduction of cochlear sensitivity during selective attention to visual stimuli [10]. The investigators found a decrease of cochlear sensitivity during the period of attention to visual stimuli but not when attending to an auditory task, demonstrating that the decrease is related to selective attention to visual stimuli rather than to arousal level. Since the effect is present early in sensory transduction the authors suggested it was mediated by activation of olivocochlear efferent fibers. The authors have tested this hypothesis by stimulating MOC efferent axons in chinchillas with sectioned middle-ear muscles, while recording cochlear potentials [11]. MOC stimulation produced CAP amplitude suppression of and CM amplitude increments similar (but somewhat smaller) than those found in studies on other animals, supporting their suggestion that the visual attention effects could have been mediated by olivocochlear activation. This finding is consistent with the broader hypothesis that efferent innervation of the inner ear Curr Opin Otolaryngol Head Neck Surg. Author manuscript; available in PMC 2012 October 01. RABBITT and BROWNELL Page 3 plays an important role in sensory integration and modulation inner ear afferent inputs to meet the behavioral needs of the organism. NIH-PA Author Manuscript Fast and slow olivocochlear effects in humans—Physiological studies have shown that the MOC system modulates basilar membrane and auditory nerve activity on both a fast (10–100 ms) and slow (10–100 s) time scale. The fast effect is thought to result from current shunting while the slow effect may result from a G-protein mediated change in OHC lateral wall mechanics. The slow effect has been postulated to aide in protection against acoustic trauma. A recent study explored the slow effect in humans by looking at the effect of contralateral acoustic stimulation on spontaneous otoacoustic emissions [12, 13]. Fast and slow effects were observed but with a high threshold and small effect magnitude for the slow effect. NIH-PA Author Manuscript Olivocochlear effects in the processing of auditory localization cues—There is a rich history of experimental and model data supporting a contribution of the MOC system in detecting relevant signals in a noisy background [5]. A systematic study of the response of cat single auditory nerve fibers suggests that the MOC system has a greater effect on low spontaneous rate fibers than on high spontaneous rate fibers [14]. Broad band noise that was shaped by one of ten head related transfer functions (HRTF) was presented to the ear in the presence and absence of noise. HRTF capture the effect of the cat's head shape on the sound arriving at the tympanic membrane and contains cues that are relevant for sound localization. There were population differences between fibers having a spontaneous firing rate of ≤ 1 spike/sec and those ≥ 18 spikes/sec. A reasonable interpretation of the differences was that activation of the MOC system expanded the intensity range over which the low spontaneous range fibers encode the HRTF information. Low and high spontaneous fibers arise from inner hair cells and why activation of the outer hair cell by the MOC results in different coding behavior at synapses on the same cell remains a mystery. This might reflect an MOC dependent nonlinearity, or it is also possible that LOC activation might combine systematically with MOC to shape the afferent discharge. NIH-PA Author Manuscript The membrane properties and synaptic responses of neurons in the mouse lateral superior olivary nucleus (LSO) were measured in a brain slice preparation [15]. The neurons could be divided into two populations based on their response latencies, ion channels and pharmacology. The authors concluded the short response latency neurons were the principal cells of the LSO involved in processing interaural intensity differences and projecting to the inferior colliculus. They postulated the slower population consisted of the cell bodies of origin for the LOC neurons that project to the afferent fibers in the IHCs in the ipsilateral cochlea. If the identity of the slower population is unequivocally identified their observations could be useful in modeling the effect of LOC activity on hearing. Olivocochlear effects on sound localization—Two notable auditory localization studies were published during the past year that examined OC effects on sound localization, one in ferrets [16]* and the other in humans [17]**. An earlier animal study [18] examined the effect of OC lesions on the ability of cats to localize a sound source in the presence of noise. Bilateral OC lesions were made at locations that should cut most of the crossed and uncrossed MOC efferent fibers. Performance in noise was worse immediately after the lesions and all three lesioned animals showed some compensation with one returning to prelesion values after 6 sessions. The study was consistent with earlier studies on the MOC mediated improvement in detection of a signal in a noisy background but the improvement provoked questions as to the basis of the neural plasticity that allowed for compensation. A behavioral study [16] looked at the effect of lesions on the ability of ferrets to compensate for the introduction of an earplug in a sound localization task. The animals received a Curr Opin Otolaryngol Head Neck Surg. Author manuscript; available in PMC 2012 October 01. RABBITT and BROWNELL Page 4 NIH-PA Author Manuscript parasaggital lesion of the OC either at the midline or to one side of the midline. Histological examination of the cochlea confirmed that LOC innervation was greatly diminished and the MOC partially spared in the ear on the same side as the lateral lesion (OC innervation in the contralateral ear appeared normal). The midline lesions were assumed to remove MOC input in both ears while preserving LOC input. All the lesioned animals learned to localize a one second sound (without noise) and localization was compromised following the introduction of an earplug. The control and the lateral lesion groups showed a modest improvement in their ability to localize over a 10 day plug period and both returned to preplug performance when the plug was removed. The lateral lesion group showed a similar loss of localization on plugging but did not compensate and they also returned to normal performance when the plug was removed. The results indicate that intact MOC innervation is required for the relearning of sound localization cues when input from one ear is compromised and that a single ear, with intact efferent innervations, is sufficient for rebalancing the processing of the binaural cues involved in localization. NIH-PA Author Manuscript Subjects in the human study identified which of 8 speakers emitted a short, band-pass burst embedded in a longer noise stimulus that was played on all eight speakers. Signal to noise ratio and the location of the sound were randomly varied. Sound localization results were compared with a variety of OAE measures including the magnitude of the OC-reflex. The ability to localize in noise as well as in the magnitude of the OC-reflex suppression were highly variable as has been previously reported. However, the magnitude of each measure covaried, with individuals having the greatest OC-reflex suppression tending to perform best in detecting the speaker in a noisy environment. This is the first study to demonstrate a relation between OC-activity and sound localization in humans and lends further support to the view that the OC-reflex facilitates sound-localization in the presence of background noise particularly at low signal to noise ratios. A study in women [19] identified one of the factors that may contribute to OC-reflex variability. The investigators looked at the time the test was performed during the ovarian cycle (as defined by oestradiol and progesterone serum levels and menstrual cycle dating). OAEs including SOAEs and TEOAEs, ABRs and MOC suppression of TEOAEs were measured. There were correlations with the cycle including a significant negative correlation of MOC suppression with oestradiol levels in the follicular phase. The results of the study show small changes in auditory function during the ovarian cycle but they are suggestive of an increased hearing sensitivity including MOC suppression around the time of ovulation. NIH-PA Author Manuscript Olivocochlear effects in overshoot—Overshoot is a psychophysical effect where a brief sound has a lower threshold when presented 100 ms or more after the start of a noise burst compared to its threshold near the start of the noise burst. Last year's review [5] described a study in which a behavioral measure of overshoot and an OAE-based measure that resembled overshoot in all but one of the same subject population [20]. Another study [21] by the same investigators provided further support for the role of the MOC system in overshoot. Very short (10 ms) tones were masked by a 400 ms noise burst. Detection improvement reached a temporary plateau between 10–30 ms, after which a steady improvement with time occurred. Similar plateaus have been reported in physiological studies of MOC effects. A computational model was used to evaluate two possible mechanisms for psychophysical overshoot, adaptation and MOC efferent feedback [22]. The influence of model variables for the detection of a tone-pip in a broadband noise revealed the MOC feedback was essential to produce overshoot. The analysis supports the hypothesis that the noise burst elicits MOC activity leading to a decrease in cochlear-amplifier gain that reduces the response to the lowlevel noise more than it reduces the response to the brief, high-level tone. Curr Opin Otolaryngol Head Neck Surg. Author manuscript; available in PMC 2012 October 01. RABBITT and BROWNELL Page 5 NIH-PA Author Manuscript Olivocochlear effects on auditory system development in a noisy environment —The possible involvement of the MOC system in auditory system development was discussed in last year's review [5]. A recent study examined its involvement in mediating the deleterious effects of noise on cochlear development [23]. A population of knock-out mice with a homozygous null mutation of the α9 nicotinic acetylcholine receptor subunit (α9KO) were raised in a noisy environment and their ability to process auditory stimuli was assessed behaviorally (startle response) and physiologically (ABR). Their results were compared with another population of the knock-out mice raised in a normal acoustic environment as well as a population of control mice raised in both the noisy and quite environments. The α9 nicotinic acetylcholine receptor subunit is found on OHCs postsynaptic to the MOC terminals and hair cells that do not have it are unable to respond to MOC activation. The knock-out mice raised in the presence of noise were less able to process rapid temporal information and showed elevated ABR thresholds in the middle frequencies. DPOAE generation was not affected but a change in wave 2 of the ABR was noted suggesting a modification of synaptic activation in the cochlear nucleus. The results suggest that the MOC system is required for the development of normal connections in the CNS in an abnormal noisy environment. NIH-PA Author Manuscript Efferent Vestibular System (EVS)—Anatomical evidence of the extensive efferent innervation of the inner ear began to appear in the early 20th century [24–26]. Phylogenetic observations highlight the ancient origin of efferent of neural pathways and draws attention to the fundamental importance of efferent action in both auditory and vestibular sensation [27–29]. Mechanisms of efferent action in the more primitive vestibular organs therefore are likely to have relevance to efferent action in the cochlea. The anatomy and physiology of the efferent vestibular system are reviewed this year in extensive articles by Holt et al. [6] and Wersinger & Fuchs [7]. NIH-PA Author Manuscript Historical evidence in-vivo suggests that a primary function of the EVS is to tune vestibular sensation to the interest and needs of the organism, for example by decreasing the sensitivity during large self-generated movements [30, 31], and adjusting the background discharge characteristics of afferents [6]. The decrease in sensitivity appears to have principal origins in efferent inhibition of hair cell electrical responses through inhibitory post-synaptic potentials [3], opening of basolateral ion channels [6], and a concomitant decrease in hair cell receptor potential modulation [3, 32]. Although EVS effects on vestibular signal encoding are profound in situ, and have been observed in some species in vivo, the same level of EVS control has not been demonstrated in primates [33]. It is not yet known if there are fundamental interspecies differences in EVS action, or if experimental conditions such as the level of attention or relevance of the stimulus to the needs of the animal might be at play. There remains a paucity of single unit recordings from efferent vestibular neurons and a lack of information regarding specific sensory stimuli or states of attention that evoke changes in EVS activity. Clearly electrical activation of the brainstem efferent vestibular nucleus has substantial effects on vestibular sensation and neural coding, but when and how the system is activated under normal physiological conditions remains speculative. Sensitivity modulation prior to self-induced movements has been shown, but very little is known about dynamic responses of efferent vestibular neurons to traditional vestibular movement stimuli, let alone responses driven by multisensory integration, bi-lateral balancing, or dynamic optimization of signal to noise analogous to that achieved in the mammalian cochlea. The primary mechanisms of EVS activation alter the electrical excitability of hair cells and afferent neurons, acting primarily through nicotinic cholinergic receptors (nAChRs), secondarily through muscarinic receptors (mAChRs) [6, 34, 35], and several apparently less understood transmitters and receptors [6]. EVS activation also decreases semicircular canal Curr Opin Otolaryngol Head Neck Surg. Author manuscript; available in PMC 2012 October 01. RABBITT and BROWNELL Page 6 NIH-PA Author Manuscript hair bundle motion in response to low strength mechanical stimuli [4], but this mechanical effect is quite small relative to the electrical effects. Nevertheless, the observation of EVS inhibition of active hair bundle movements in a teleost vestibular organ suggests that neural control of hair cell mechanical amplification predates the appearance of outer hair cells in the mammalian cochlea. Efferent control of bundle-based amplification in non-mammalian hearing organs might be a general principle, such as control of short hair cell bundle-based amplification in the avian auditory papilla. The biophysics underlying efferent control of hair bundle mechanical amplification is not entirely clear [3], but has been speculated to be controlled by somatic electrical shunting [36]. Both electrical and mechanical actions of efferent innervation in the vestibular system may have particular relevance to efferent action in the low-frequency apical region of the mammalian cochlea. NIH-PA Author Manuscript Kinetics of efferent action on hair cells—Inner ear hair cell organs transduce signals with frequencies ranging from zero (gravity) to nearly 100kHz in some mammals. The kinetics of efferent action on hair cells follows a wide range of time courses, presumably reflecting needs of the animal and specializations of specific hair cell sensory organs [3, 32, 37]. Recent evidence suggests timing differences might partially involve the spatial distribution of channels/receptors and intracellular organelles that alter reaction-diffusion kinetics and temporal properties of intracellular calcium signaling [7]*. Pulsed infrared laser stimuli has been shown to evoke transient mitochondrial calcium currents [38], thus modulating synaptic transmission from vestibular hair cells with a delay of ~7ms [39]. This form of stimulation might prove useful to investigate temporal responses of hair cells to optically controlled calcium transients. NIH-PA Author Manuscript Efferent involvement in tuning bilateral sensory integration—Both the auditory and vestibular systems integrate bilateral information at brainstem nuclei. Both systems are characterized by the convergence of topographically organized input onto topographically organized neurons (similar frequency inputs from both ears to the superior olivary nuclei and input from mirror end organs to the vestibular nuclei). The conduction time for the information to arrive at the midline nuclei must be regulated within close tolerances [40] and a peripheral insult would be expected to alter timing. The total conduction time includes the time for action potentials to traverse the conduction pathways as well as for dendritic integration and synaptic delays. Axonal conduction velocity is determined by fiber diameter and involves axonal-glial signaling to adjust the internodal length between nodes of Ranvier and the thickness of the myelin sheath [40, 41]. Both sensory modalities have sufficient plasticity to compensate for either temporary or permanent insult to the inner ear. Compensation for damage to the end organ is most likely an extreme example of a normal tuning of the conduction time that must occur to optimize bilateral sensory integration. The efferent innervation to the periphery originates from neurons located in and around the very nuclei that process bilateral information so they are ideally poised to monitor the timing and activate the requisite feedback mechanisms. The OC and EVS efferent systems release acetylcholine and the comparison with motor systems has often been made. Efferents to the lateral line organs fire with motor neuron activation of the muscles involved in swimming [42] and our middle ear muscles are activated by speech. Other agents are also released at efferent terminals that could be part of second messenger signaling either for activating maturation or maintaining system tolerances. Inner ear efferent activation may be associated with motor activity such as bruxism and myoclonic jerks for which there is no known function. Bruxism generates loud bone conducted sounds that would essentially activate both ears simultaneously. Myoclonic jerks will activate the vestibular epithelia. Speculatively, the midline auditory and vestibular nuclei could assess whether the conduction time is appropriate. If not, the efferent fibers would signal the afferent fibers to adjust conduction velocity appropriately. Curr Opin Otolaryngol Head Neck Surg. Author manuscript; available in PMC 2012 October 01. RABBITT and BROWNELL Page 7 CONCLUSION NIH-PA Author Manuscript Research on the effects of efferent activity on hair cells has provided insight on the function and feedback control of inner ear mechanoreceptor sensory organs by the brain. Efferent fiber activity modulates the gain of electromechanical motors in both the auditory and vestibular system. The ability of olivocochlear fiber activity to alter the gain of the cochlear amplifier is well established with strong evidence that it facilitates the detection of acoustic signals in a noisy environment. New findings suggest the OC system may play a role in sensory plasticity but the roles of the MOC and LOC in these processes are as yet unclear. Acoustic activation of olivocochlear fibers is used in the clinic to test the integrity of the outer hair cell function. The structural and functional similarities of efferent stimulation of the auditory and vestibular portions of the inner ear offer hope that testing procedures may be developed that will allow reliable testing of the vestibular hair cell function. Acknowledgments We thank Dr. JJ Guinan for providing the outline and structure for this review in his 2010 review of olivocochlear efferents in Current Opinion in Otolaryngology & Head and Neck Surgery. Supported by NIH NIDCD RO1s DC004928 (RDR) and DC000384/DC002775 (WEB) Supported by NIDCD R01s DC004928/DC006685 (RDR) and DC000384/DC002775 (WEB) NIH-PA Author Manuscript References and recommended reading Papers of particular interest, published within the annual period of review, have been highlighted as: * of special interest ** of outstanding interest NIH-PA Author Manuscript [1]. Brownell WE, Bader CR, Bertrand D, de Ribaupierre Y. Evoked mechanical responses of isolated cochlear outer hair cells. Science. 1985; 227:194–196. [PubMed: 3966153] [2]. Brownell WE, Lue AJC. Hair cell motility. Current Opinion in Otolaryngology & Head & Neck Surgery. 1996; 4:289–293. [3]. Castellano-Munoz M, Israel SH, Hudspeth AJ. Efferent control of the electrical and mechanical properties of hair cells in the bullfrog's sacculus. PLoS One. 2010; 5:e13777. [PubMed: 21048944] [4]. Rabbitt RD, Boyle R, Highstein SM. Mechanical amplification by hair cells in the semicircular canals. Proc Natl Acad Sci U S A. 2010; 107:3864–3869. [PubMed: 20133682] [5]. Guinan JJ Jr. Cochlear efferent innervation and function. Curr Opin Otolaryngol Head Neck Surg. 2010; 18:447–453. [PubMed: 20717032] [6]. Holt, JC.; Lysakowski, A.; Goldberg, JM. The efferent vestibular system. In: Auditory and Vestibular Efferents. In: DKRe. , editor. Springer Handbook of Auditory Research. Springer; 2011. [7]. Wersinger E, Fuchs PA. Modulation of hair cell efferents. Hear Res. 2010*Review examining the importance of calcium signaling on temporal properties of hair cell responses to efferent synaptic inputs. [8]. Ryugo, DK.; Fay, RR.; Popper, AN. Auditory and Vestibular Efferents. In: Fay, RR.; Popper, AN., editors. Springer Handbook of Auditory Research. 1st. Springer Science+Business Media, LLC; New York: 2011. p. 360 [9]. Rabbitt RD, Clifford S, Breneman KD, et al. Power efficiency of outer hair cell somatic electromotility. PLoS Comput Biol. 2009; 5:e1000444. [PubMed: 19629162] [10]. Delano PH, Elgueda D, Hamame CM, Robles L. Selective attention to visual stimuli reduces cochlear sensitivity in chinchillas. J Neurosci. 2007; 27:4146–4153. [PubMed: 17428992] Curr Opin Otolaryngol Head Neck Surg. Author manuscript; available in PMC 2012 October 01. RABBITT and BROWNELL Page 8 NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript [11]. Delano PH, Elgueda D, Ramirez F, et al. A visual cue modulates the firing rate and latency of auditory-cortex neurons in the chinchilla. J Physiol Paris. 2010; 104:190–196. [PubMed: 19958832] [12]. Zhao W, Dhar S. The effect of contralateral acoustic stimulation on spontaneous otoacoustic emissions. J Assoc Res Otolaryngol. 2010; 11:53–67. [PubMed: 19798532] [13]. Zhao W, Dhar S. Fast and slow effects of medial olivocochlear efferent activity in humans. PLoS One. 2011; 6:e18725. [PubMed: 21494578] [14]. Reiss LA, Ramachandran R, May BJ. Effects of signal level and background noise on spectral representations in the auditory nerve of the domestic cat. J Assoc Res Otolaryngol. 2011; 12:71– 88. [PubMed: 20824483] [15]. Sterenborg JC, Pilati N, Sheridan CJ, et al. Lateral olivocochlear (LOC) neurons of the mouse LSO receive excitatory and inhibitory synaptic inputs with slower kinetics than LSO principal neurons. Hear Res. 2010; 270:119–126. [PubMed: 20813177] [16]. Irving S, Moore DR, Liberman MC, Sumner CJ. Olivocochlear efferent control in sound localization and experience-dependent learning. J Neurosci. 2011; 31:2493–2501. [PubMed: 21325517] *The effect of OC lesions on the ability of ferrets to compensate for a unilateral earplug in a sound localization task. The results indicate that intact MOC innervation is required for the relearning of sound localization cues and that a single ear, with intact efferent innervations, is sufficient for rebalancing the processing of the binaural cues involved in localization. [17]. Andéol G, Guillaume A, Micheyl C, et al. Auditory Efferents Facilitate Sound Localization in Noise in Humans. The Journal of Neuroscience. 2011; 31:6759–6763. [PubMed: 21543605] **It is difficult to use conventional contralateral acoustic stimulation to evoke an MOC response when performing a sound localization task in a noisy environment. Using a clever experimental design, these investigators demonstrated that a measure of sound localization covaried with OCreflex suppression. This is the first study to demonstrate a relation between OC-activity and sound localization in humans and, while it is not a definitive demonstration, it lends further support to the view that the OC-reflex facilitates sound-localization in the presence of background noise. [18]. May BJ, Budelis J, Niparko JK. Behavioral studies of the olivocochlear efferent system: learning to listen in noise. Arch Otolaryngol Head Neck Surg. 2004; 130:660–664. [PubMed: 15148194] [19]. Al-Mana D, Ceranic B, Djahanbakhch O, Luxon LM. Alteration in auditory function during the ovarian cycle. Hear Res. 2010; 268:114–122. [PubMed: 20685243] [20]. Walsh KP, Pasanen EG, McFadden D. Overshoot measured physiologically and psychophysically in the same human ears. Hear Res. 2010; 268:22–37. [PubMed: 20430072] [21]. McFadden D, Walsh KP, Pasanen EG, Grenwelge EM. Overshoot using very short signal delays. J Acoust Soc Am. 2010; 128:1915–1921. [PubMed: 20968363] [22]. Jennings SG, Heinz MG, Strickland EA. Evaluating adaptation and olivocochlear efferent feedback as potential explanations of psychophysical overshoot. J Assoc Res Otolaryngol. 2011; 12:345–360. [PubMed: 21267622] [23]. Lauer AM, May BJ. The medial olivocochlear system attenuates the developmental impact of early noise exposure. J Assoc Res Otolaryngol. 2011; 12:329–343. [PubMed: 21347798] [24]. Engström H, Wersäll J. The ultrastructructural organization of the organ of Corti and of the vestibular sensory epithelia. Exp Cell Res. 1958; 5:460–492. [PubMed: 13586306] [25]. Lorente de Nó R. Anatomy of the eighth nerve: The central projection of the nerve endings of the inner ear. Laryngoscope. 1933; 43:1–38. [26]. Rasmussen GL. The olivary peduncle and other fiber projections of the superior olivary complex. J. Comp. Neurol. 1946; 84:141–219. [PubMed: 20982804] [27]. Elliott KL, Fritzsch B. Transplantation of Xenopus laevis ears reveals the ability to form afferent and efferent connections with the spinal cord. Int J Dev Biol. 2011; 54:1443–1451. [PubMed: 21302254] [28]. Gilland E, Baker R. Evolutionary patterns of cranial nerve efferent nuclei in vertebrates. Brain Behav Evol. 2005; 66:234–254. [PubMed: 16254413] Curr Opin Otolaryngol Head Neck Surg. Author manuscript; available in PMC 2012 October 01. RABBITT and BROWNELL Page 9 NIH-PA Author Manuscript NIH-PA Author Manuscript [29]. Muller F, O'Rahilly R. The initial appearance of the cranial nerves and related neuronal migration in staged human embryos. Cells Tissues Organs. 2010; 193:215–238. [PubMed: 20980719] [30]. Tricas TC, Highstein SM. Visually mediated inhibition of lateral line primary afferent activity by the octavolateralis efferent system during predation in the free-swimming toadfish, Opsanus tau. Exp Brain Res. 1990; 83:233–236. [PubMed: 2073946] [31]. Tricas TC, Highstein SM. Action of the octavolateralis efferent system upon the lateral line of free-swimming toadfish, Opsanus tau. J Comp Physiol [A]. 1991; 169:25–37. [32]. Boyle R, Rabbitt RD, Highstein SM. Efferent control of hair cell and afferent responses in the semicircular canals. J Neurophysiol. 2009; 102:1513–1525. [PubMed: 19571186] [33]. Sadeghi SG, Goldberg JM, Minor LB, Cullen KE. Efferent-mediated responses in vestibular nerve afferents of the alert macaque. J Neurophysiol. 2009; 101:988–1001. [PubMed: 19091917] [34]. Li GQ, Correia MJ. Responses of pigeon vestibular hair cells to cholinergic agonists and antagonists. Brain Res. 2011; 1373:25–38. [PubMed: 21147073] [35]. Perez C, Limon A, Vega R, Soto E. The muscarinic inhibition of the potassium M-current modulates the action-potential discharge in the vestibular primary-afferent neurons of the rat. Neuroscience. 2009; 158:1662–1674. [PubMed: 19095045] [36]. Breneman KD, Brownell WE, Rabbitt RD. Hair cell bundles: flexoelectric motors of the inner ear. PLoS One. 2009; 4:e5201. [PubMed: 19384413] [37]. Wersinger E, McLean WJ, Fuchs PA, Pyott SJ. BK channels mediate cholinergic inhibition of high frequency cochlear hair cells. PLoS One. 2010; 5:e13836. [PubMed: 21079807] [38]. Dittami GM, Rajguru SM, Lasher RA, et al. Intracellular calcium transients evoked by pulsed infrared radiation in neonatal cardiomyocytes. J Physiol. 2011; 589:1295–1306. [PubMed: 21242257] [39]. Rajguru SM, Richter CP, Matic AI, et al. Infrared photostimulation of the crista ampullaris. J Physiol. 2011; 589:1283–1294. [PubMed: 21242259] [40]. Seidl AH, Rubel EW, Harris DM. Mechanisms for adjusting interaural time differences to achieve binaural coincidence detection. J Neurosci. 2010; 30:70–80. [PubMed: 20053889] [41]. Rushton WA. A theory of the effects of fibre size in medullated nerve. J Physiol. 1951; 115:101– 122. [PubMed: 14889433] [42]. Russell IJ. The role of the lateral-line efferent system in Xenopus laevis. J Exp Biol. 1971; 54:621–641. [PubMed: 5090097] NIH-PA Author Manuscript Curr Opin Otolaryngol Head Neck Surg. Author manuscript; available in PMC 2012 October 01. RABBITT and BROWNELL Page 10 KEY POINTS NIH-PA Author Manuscript • Efferent fiber activity modulates the gain of electromechanical motors in both the auditory and vestibular system. • Olivocochlear (OC) fiber activity alters the gain of the cochlear amplifier and improves the detection of acoustic signals in a noisy environment. • New findings suggest the OC system may play a role in sensory plasticity but the roles of the medial OC and lateral OC in these processes are as yet unclear. • Acoustic activation of olivocochlear fibers provides an objective clinical test for the functional integrity of outer hair cells. • The structural and functional similarities of inner ear sensory epithelia offer hope that testing procedures based on activation of the efferent vestibular system (EVS) may be developed that would allow testing of vestibular hair cell function. NIH-PA Author Manuscript NIH-PA Author Manuscript Curr Opin Otolaryngol Head Neck Surg. Author manuscript; available in PMC 2012 October 01.