Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
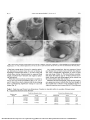
Investigative Ophthalmology & Visual Science, Vol. 30, No. 5, May 1989 Copyright © Association for Research in Vision and Ophthalmology Optical Constancy of the Chick Lens During Pre- and Post-Hatching Ocular Development J. G. Sivok, L. A. Ryall, J. Weerheim, and M. C. W. Campbell Embryonic and post-embryonic development of the ocular lens is associated with the continual production of new secondary lens fibers by the mitotic activity of equatorial epithelial cells. This continual development affects lens size and shape and refractive index distribution. Study of embryonic lens optical function has been largely ignored. The optical characteristics of the developing chick lens, including paraxial and eccentric focal lengths, were measured during the embryonic period of development and up to 15 days after hatching. Measurements were made with an automated scanning laser system in which the video image of a helium-neon laser beam refracted by an excised lens in solution is digitized. Focal length is measured for beams moving in small steps on either side of that center. Measurements were made on excised lenses as well as with the lens in situ within the anterior segment of the eye. The results, collected from a study of a total of 80 lenses, indicates that embryo lenses at 6-7 days of incubation have long and very variable focal lengths. At the tenth embryo day, focal length drops by more than one-half and focal variations, between lenses and for different beam positions within a single lens, is reduced. Further measures for 14- and 17-day embryo lenses, as well as for lenses from hatchling and 5-, 10- and 15-day-old chicks, indicate that there is little change in focal length, either paraxially or for eccentric beam positions. Measures of lens size and shape from frozen eye sections and from freshly excised lenses indicate that this focal constancy is accompanied by major changes in lens size and shape. These results indicate that the developing chick lens may be a static refractive feature of the developing eye. It is suggested that this focal stability simplifies the process of emmetropization in that ocular refractive state is governed by manipulating the size and shape of only the ocular globe. Invest Ophthalmol Vis Sci 30:967-974, 1989 The lens of the eye is a cellular structure that develops from the surface ectoderm.1'2 In all vertebrates, the lens initially forms a hollow vesicle which is filled by the elongation of the cells of the posterior hemisphere. The newer cells, the primary lens fibers, are joined by new elongating cells formed around the equator. These cells, the secondary lens fibers, produce successive layers or shells around the primary ones. The continual growth of the lens and the compression of older tissue toward the center results in the formation of a lens of variable refractive index, the center being higher than the periphery.3 This factor has the important optical consequence of affecting the ability of the lens to control spherical aberration.4-5 While the above developmental sequence is common to all vertebrate lenses, the size, shape and refractive index distribution of the adult lens of each species will depend on the overall refractive plan of the eye. Thus in aquatic species such as fish, where the lens is the only refractive element of the eye, the lens is usually spherical in shape and has an elevated refractive index. The lens of terrestrial animals is often elliptical in shape, softer in consistency and lower in refractive index.5'6 However, among terrestrials, lens relative size and shape and consistency may vary considerably, depending on whether the eye is used in nocturnal or diurnal light conditions.67 Major variations in lens shape have been noted even among fish.8 These variations in lens shape may have dramatic effects on lens focal properties.5'9 A recent preliminary study of a limited number of human embryo lenses suggests that the developing human lens has an approximately constant focal length and a minimal amount of spherical aberration.10 In view of the extensive change in size and shape of the lens during embryo development, this result is very surprising. The study which follows is an effort to examine focal variation during lens development in greater detail. From the School of Optometry, University of Waterloo, Waterloo, Ontario, Canada. The support of the Medical Research Council of Canada and of the Natural Sciences and Engineering Research Council of Canada is gratefully acknowledged. Submitted for publication: May 9, 1988; accepted October 25, 1988. Reprint requests: J. O. Sivak, School of Optometry, University of Waterloo, Waterloo, Ontario, Canada N2L 3G1. 967 Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933377/ on 05/05/2017 968 Vol. 30 INVESTIGATIVE OPHTHALMOLOGY & VISUAL 5CIENCE / May 1989 0.0 2.0 4.0 6.0 8.0 10.0 Fig. 1. Sample focal length measurements for single lens from a 15-dayold chick. Focal length (mm) on abcissa, eccentricity (mm) from optic axis (0.0) on ordinate. Note characteristic negative spherical aberration. Data points (+) indicate axial crossover point for each eccentricity. FOCAL LENGTH (mm) Materials and Methods Chick embryo lenses were obtained from eggs (Cobb Arbor Acres, New Hamburg, Ontario, Canada) incubated for specific numbers of days at a local hatchery. The eggs were opened and the embryos transected close to the anterior aspect of the spinal column. Lenses were carefully removed from the embryo eyes and placed in physiological saline. All animal tissue used in this study was obtained in a manner in keeping with the ARVO Resolution on the Use of Animals in Research. Chick lenses were also obtained from hatchling chicks (Cobb Arbor Acres) of various ages. These chicks were collected on the day of hatching from the same hatchery. The chicks were housed in stainless steel chick brooders for varying periods, up to 15 days. Brooder temperatures were kept at 32°C for the first week and 20°C for the second week, as necessary. The chicks were fed commercial chick starter and water ad libitum. A fluorescent light schedule of 14 hr light and 10 hr dark was maintained. Chicks were sacrificed at 5, 10 and 15 days after hatching by CO2 asphyxiation. Lenses were carefully removed from the enucleated eyes and placed in physiological saline. The optical (focal) properties of the excised lenses (embryo and post-hatching) were determined with an automated laser scanner." The scanner consists of a low-power helium-neon laser mounted on a computer-controlled X-Y table, and a television camera with video frame digitizer. The laser is programmed to scan across the chick lens in predetermined steps while the digitizer determines the position of the beam after it is refracted by the lens. In practice, the device first locates the optical center of the lens by locating the position of zero or minimal refraction for both the X and Y direction. It then determines focal lengths for beam positions on either side of the center. Scanning step sizes of 0.05 to 0.10 mm were used in this experiment (Fig. 1). The principal plane is the surface of unit magnification defined by the intercept of the path of the incoming ray with the path of the existing ray. The focal length is measured from this intercept to the intercept of the ray with the optic axis along a path parallel to the optic axis. Changes in this distance with ray eccentricity are influenced by the presence of coma and spherical aberration, but spherical aberration is the dominant factor. Focal variations are therefore referred to as spherical aberration. Each lens was placed in a special lens cell on a ring of Duxseal® (Johns-Manville, Asbestos, Quebec, Canada), a soft pliant material used to minimize lens damage or deformation. Each lens was maintained in physiological saline (0.9% NaCl) in the cell i for the duration of the scan. Lenses were scanned within 3 hr after excision. Tests indicate little or no change in lens focal properties or transparency for 12 hr after being placed in saline. To provide an overview of change in lens shape and size during development, frozen sections were made of two eyes of each incubation stage studied. This involved freezing the eyes rapidly in acetone and dry ice (-80°C) and mounting each eye in a position appropriate for axial sectioning on a freezing microtome head. As thin sections (10-20 /urn) of the central portion of the eye were removed, photographs were taken of the remaining block of tissue with a singlelens reflex camera mounted above the preparation. The photograph showing the greatest lens dimensions (to an accuracy of ±0.025 mm) was assumed to represent an axial section of the lens. A photograph of a millimeter rule provided the control for magnification. As an indication of the possible effect of freezing on lens shape and size, as well as the effect of le|ns removal from the eye on its shape, the axial and equatorial diameters of a small number of freshly excised embryo lenses of various ages were measured with a vernier caliper to an accuracy of ±0.05 mm. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933377/ on 05/05/2017 CHICK LENS DEVELOPMENT / Sivok er a\ No. 5 969 7 DAY CHICK EMBRYOS Fig. 2. Average focal lengths for 7-day-old chick embryo lenses. Focal length (mm) on ordinate, eccentricity from (mm) optic axis (0.0) on abcissa. Bars denote standard error of the mean. -1.0 -0.3 -O'.O' 015' DISTANCE FROM OPTIC CENTRE (mm) For a more sensitive indication of the effect of removal of the lens on its optical properties, a limited number of laser scans were made with the lens in place in its suspensory apparatus. In this case the posterior one-third of the globe was removed. The rest of the eye was placed in a lens cell and immersed in physiological solution. Since the refractive power of the cornea is virtually eliminated when its external surface is in water the refractive effect measured by the scanner may be assumed to be that of the lens alone.12 This procedure was used with 5- and 15-day chicks and 17-day embryos. The procedure was not possible for earlier embryos because of the excessive flexibility of the developing globe. Results The results are based on the study of a total of 39 chick embryo lenses (6-21 days of incubation) and 22 post-hatching chick lenses (0-15 days). The lower limit of embryo incubation age, 6 days, resulted from difficulty experienced in handling and orienting smaller lenses (<0.7 mm diameter) as well as to mechanical and software limits of the scanning laser system. At 6 and 7 days of incubation chick embryo lenses have long and variable focal lengths (Fig. 2). This is true both for axial and eccentric measures of focal length. Spherical aberration, as indicated by the dif- ference in focal length between paraxial and eccentric lens positions, varies non-monotonically between positive (undercorrected) and negative (overcorrected) values. This finding is probably not the result of handling deformation since the lens at this age has a more rigid consistency than lenses from older embryos (eg, 21 days) which show monotonic focal length variation even though they are softer and more difficult to handle. After a few more days of incubation (10th day of incubation) the chick embryo lens demonstrates a precipitous drop in focal length (from 30-35 mm to 14 mm) as well as a much smaller variation in focal length between lenses (Fig. 3). The variations between paraxial and eccentric focal lengths (spherical aberration) is reduced. The aberration that exists increases continuously in the direction of negative (overcorrected) spherical aberration. Thus eccentric focal lengths, particularly those near the edge of the scan are longer (about 20 mm) than near axial ones (about 10 mm). • • • Lenses from 14 day, 17 day and 21 day embryos demonstrate optical characteristics very similar to those which have been incubated for 10 days (Fig. 4). This is true of axial focal length, interlens focal variation and the appearance of negative spherical aberration. Paraxial focal lengths average between 12 to 14 mm while focal lengths for rays refracted by the edge Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933377/ on 05/05/2017 970 Vol. 30 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / May 1989 10 DAY CHICK EMBRYOS N=6 Fig. 3. As Figure 2for10day-old chick embryo lenses. -0.5 -O'.O' 0l5' DISTANCE FROM OPTIC CENTRE (mm) 21 DAY CHICK EMBRYOS N=7 Fig. 4. As Figure 2 for 21 day-old chick embryo lenses. -1.0 -0.5 -0.0 0.5 1.0 DISTANCE FROM OPTIC CENTRE (mm) Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933377/ on 05/05/2017 071 CHICK LENS DEVELOPMENT / Sivok er ol No. 5 Fig. 5. Frozen hemi-sectionsofchick embryo eyes at (A) day 7, (B)day 10, (C)day 14, (D) day 21. Note changing size and shape of the lens. Magnification X9. The unilateral appearance of a lens in (A) is a result of the fact that the two eyes were not sectioned in the same plane. of the lens average about 20 mm (ie, negative spherical aberration). Lenses from the eyes of chicks at hatching as well as lenses from 5-, 10- and 15-day-old chicks show similar characteristics (ie, paraxial focal length, interlens focal variation and negative spherical aberration). While the frozen sections are limited in number to one or two eyes of each incubation age (7, 10, 14, 17 and 21 days), they clearly indicate that the developing lens is undergoing significant change in size and shape (Fig. 5). As a rough comparison, the two measures, frozen eye (frozen lens in frozen eye) and fresh lens diameters, show considerable agreement in terms of lens size and shape (Table 1). The lens flattens considerably from the embryo age of 10 days onward. Curiously, frozen and fresh lenses appear to be rounder on the tenth embryo day than on day 7. Measures of lens focal length, with the lens suspensory apparatus intact, indicate that the focal length of the excised lens is shorter in the periphery than it is when the anterior eye is intact (Fig. 6). The difference Table 1. Fresh lens and frozen lens dimensions. Number in brackets refers to number of lenses tested. R = equatorial diameter/axial diameter Fresh lens diameter (mm) Embryo age (days) 7 10 14 17 19 21 ' Frozen lens diameter (mm.) Equatorial Axial R Equatorial Axial R 1.34(3) 1.88(4) 2.25 (4) 0.62 (3) 1.24 (4) 1.40(4) 2.16 1.52 1.61 1.35(2) 1.68(1) 2.45 (2) 2.87 (2) 0.83 (2) 1.36(1) 1.45(2) 1.50(2) 1.63 1.24 1.69 1.91 3.16(4) 1.56(4) 2.02 3.30 (2) 1.73(2) 1.90 Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933377/ on 05/05/2017 972 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / May 1989 Vol. 30 15 DAY POST HATCHING N=g oenudeated lens • non-enucleated lens E X o -2.0 -1.0 -0.0 Fig. 6. Focal length measurements on either side of the optic axis for enucleated and nonenucleated 15-dayold chick lenses. In the latter case the eye is intact except for the removal of the posterior and sclera, choroid and retina. Corneal refractive power is eliminated by immersion in water. Bars denote standard error of the mean. 1.0 DISTANCE FROM OPTIC CENTRE (mm) is minimal paraxially and larger (to 3 mm) at more eccentric locations. In other words, the difference between paraxial and eccentric focal lengths is decreased by lens excision. These differences are presumed to be the result of changes in shape of the excised lens. However, this assumes that the immersed cornea does not contribute to spherical aberration. Since the difference was found at the three ages studied (5- and 15-day embryos and 17-day embryos), it may also be assumed that the constancy of optical properties referred to in this study is true of both conditions. Discussion The results of this study are consistent with earlier work on human embryo lenses in that they suggest that lens focal length, and therefore lens refractive power, is relatively constant (Fig. 7) during most of the period of lens embryonic development.10 The wide range in focal length between lenses and within a single lens at the 6 to 7 day incubation period indicates that lens ultrastructure and molecular organization is irregular until after this point. The lack of focal constancy may be related to local variations in refractive index within the matrix of lens tissue. The significant drop in focal length from the 6th and 7th day of incubation to the 10th day indicates that the overall refractive index of the developing lens is in- creasing at a rate beyond the tendency for an increase in focal length due to the marked increase in anterior and posterior lens radii of curvature. In this context it is of interest to note that the presence of various classes of lens crystallins varies with age and location in the embryo and post-hatching chick lens13 and in the mammalian lens.14 From the 1 Oth day of incubation onwards, change in lens size and shape and/or refractive index distribution, does not affect its axial focal characteristics. This is true for both paraxial and eccentric lens positions, indicating that spherical aberration (the (difference in focal length for paraxial and eccentric positions) does not increase with development, a finding opposite to that of the developing rat lens.15 The fact that this constancy is carried over into the post-hatching period of lens development supports the existence of a mechanism which predetermines lens refractive power in isolation from other ocular refractive developments and regardless of whether light rays enter the eye or not. In third-order aberration theory, minimum blur will occur three-quarters of the distance between the paraxial and marginal focal points.16 Since the^ lens paraxial focal points are constant and marginal focal points are similar, the position of minimum blujr will not change with age. Of course total ocular refractive power will depend on both the cornea and the lens. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933377/ on 05/05/2017 970 CHICK LENS DEVELOPMENT / Sivok er ol No. 5 CHICK DEVELOPMENT AVERAGED VALUES Fig. 7. Summary plot of average focal lengths for all laser beam positions for preand post-hatching chick lenses indicating fairly constant focal lengths from day 10 of the developing embryo. Bars denote standard error of the mean. (PRE- AND POST-HATCHING) Z LU < O o 10 15 20 25 AGE OF EMBRYO/CHICK The developmental isolation of the lens, in the refractive sense, is suggested by a variety of other evidence. For example, recent studies, in which myopia is produced in hatchling chicks by depriving the eye of form vision, show that the gross anatomy, biochemistry and focal characteristics of the lens are not altered.16"19 These findings are contrary to those of Coulombre and Coulombre who note that lens growth and development is largely tied in with globe development.20 However, the possible independence of lens development is not ruled out entirely, even in the latter work. Efforts to analyze age-related change of the refractive components of the living human eye yield conflicting data. Gordon and Donzis use corneal and axial measurements of the eye to calculate lens refractive power.21 Their results show that the refractive power of the human lens varies from 34.4 diopters in the newborn to 18.8 in the adult eye. Grosvenor,22 on the other hand, used Sorsby's data23 to show that age related change in human lens refractive power is small (1.0 to 1.5 diopters). While the data presented here and earlier have raised the possibility that the lens maintains a constant focal profile,10 it is clear that this cannot be a universal property of the vertebrate lens. For example, the typical fish lens, the only refractive element of the fish eye has a never-changing spherical shape.6 In 30 35 (DAYS) Haplochromis burtoni, a cichlid fish, focal length has been shown to scale with lens size.24 However, the control of lens spherical aberration can be shown to vary with age and with species in accord with visual need.5 The shape and focal nature of the rat lens has also been shown to vary considerably during the immediate (2 week) post-natal period.14 In this case, the lens of the newborn, which is elliptical in shape and more or less free of spherical aberration, rapidly assumes the large and nearly spherical proportions of the adult rat lens. Large and characteristic amounts of negative spherical aberration develop as well. In the avian eye, the ciliary body (and muscle) is in direct contact with the lens by way of ciliary folds.6 It is, therefore, important to note that the shape of the excised bird lens may be influenced by the absence of contact with the ciliary body. It is noted that this study shows relatively little difference between the excised lens and the lens in situ. However, little is known regarding the development of the avian accommodative mechanism nor is it even certain whether accommodation in chickens involves lens change only or change in curvature of the lens and the cornea. 122526 Despite the uncertainty regarding the effect of excision, it is clear from the frozen sections and measures of fresh lenses representing the various developmental stages of the eye that lens size is changing Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933377/ on 05/05/2017 974 INVESTIGATIVE OPHTHALMOLOGY G VISUAL SCIENCE / May 1989 dramatically. Despite the rounder appearance of the 10-day embryo lens (Table 1) the change in shape of the chick lens is more or less in accordance with that expected from the study of lens development. The pre- and post-natal human lens is commonly described as changing from a more spherical to a more elliptical shape.1-2-27 This change is inferred to result from the continued equatorial development of new lens cells and their tapered anterior and posterior growth. Thus it appears that the chick lens (and possibly the human embryo lens) may be a static refractive feature in an otherwise complex and changing optical system. Whether this constancy is a coincidental development resulting from a balance between size and shape of the lens and refractive index distribution or whether it is a genetically programmed characteristic may be difficult to ascertain. A spherical distribution of refractive index within a lens can give rise to the same axial optics as an ellipsoidal refractive index distribution.28 Thus, it is important to note that even if the lens changes in shape and refractive index in such a way as to keep focal length and spherical aberration constant, the off-axis optical properties of the lens may be expected to change. Nevertheless it is tempting to suggest that the focal stability of the lens simplifies the process of emmetropization; for rather than coping with two independent variables, lens and globe, the refractive state of the eye is controlled by manipulating the size and shape of the globe (sclera and cornea) alone. Key words: chick, lens, focal length, embryo, post-hatching References 1. Mann I: The Development of the Human Eye, 3rd edition. New York, Grune and Stratton, 1969. 2. Arey LB: Development of the lens: A classical approach. In Cataract and Abnormalities of the Lens, Bellows JG, editor. New York, Grune and Stratton, 1975, pp. 42-48. 3. Pumphrey RJ: Concerning vision. In The Cell and the Organism, Ramsay JG and Wigglesworth VB, editors. Cambridge, Cambridge University Press, 1961, pp. 193-208. • 4. Campbell MCW and Hughes A: An analytic, gradient index schematic lens and eye for the rat which predicts aberrations for finite pupils. Vision Res 21:1129, 1981. 5. Sivak JG and Kreuzer RO: Spherical aberration of the crystalline lens. Vision Res 23:59, 1983. 6. Walls GL: The Vertebrate Eye and its Adaptive Radiation. Bloomfield Hills, Michigan, Cranbrook Institute of Science, 1942. Vol. 30 7. Gur M and Sivak JG: Refractive state of the eye of a small diurnal tnannal: The ground squirrel. Am J Optom Physiol Optics 56:689, 1979. 8. Sivak JG: Optical variability of the fish lens. In The Visual System of Fish, Douglas RH and Djamgoz MBA, editors. Buckingham, Kent, Croom-Helm Ltd., 1988, in press. 9. Sivak JG, Gur M, and Dovrat A: Spherical aberration of the lens of the ground squirrel (Spermophilis Lridecemlineatus). Ophthalmic Physiol Opt 3:261, 1983. 10. Sivak JG and Dovrat A: Embryonic lens of the human eye as an optical structure. Am J Optom Physiol Optics 64:559,1987. 11. Sivak JG, Gershon D, Dovrat A, and Weerheim J: Computer assisted scanning laser monitor of optical quality of the excised crystalline lens. Vision Res 26:1873, 1986. 12. Sivak JG, Hildebrand TE, Lebert CG, Myshak LM, and Ryall LA: Ocular accommodation in chickens: Corneal vs lenticular accommodation and effect of age. Vision Res 11:186;5, 1986. 13. Genis-Galvez JM, Maisel H, and Castro J: Changes in chick lens proteins with aging. Exp Eye Res 7:593, 1968. 14. Breitman ML, Lok S, Wistow G, Piatigorsky J, Tijeton JA, Gold RJM, and Tsui L: •y-crystalline family of the mouse lens: Structural and evolutionary relationships. Proc Natl Acad Sci USA 81:7762, 1984. 15. Sivak JG and Dovrat A: Aging and the optical quality of the rat lens. Invest Ophthalmol Vis Sci 24:1162, 1983. 16. Fincham WHA and Freeman MH: Optics. London, Butterworths, 1980. 17. Hayes BP, Fitzke FW, Hodos W, and Holden AL: A morphological analysis of experimental myopia in young chickens. Invest Ophthalmol Vis Sci 27:981, 1986. 18. Wallman J and Adams JI: Developmental aspects of experimental myopia in chicks: Susceptibility, recovery and relation to emmetropization. Vision Res 27:1139, 1987. 19. Pickett-Seltner RL, Weerheim J, Sivak JG, and Pasternak J: Experimentally induced myopia does not affect post-hatching development of the chick lens. Vision Res 27:1779, 1987. 20. Coulombre JL and Coulombre AJ: Lens development: IV. Size, shape and orientation. Invest Ophthalmol 8:251, 1969. 21. Gordon RA and Donzis PB: Refractive development of the human eye. Arch Ophthalmol 103:785, 1985. 22. Grosvenor T: Reduction in axial length with age: An emmetropizing mechanism for the adult eye. Am J Optomi Physiol Optics 64:657, 1987. 23. Sorsby A, Benjamin JB, Davey M, Sheridan M, and! Tanner JM: Emmetropia and its Aberrations. London, Her Majesty's Stationery Office, 1957. 24. Fernald RD and Wright SE: Maintenance of optical! quality during crystalline lens growth. Nature 301:618, 1983. 25. Troilo D and Wallman J: Changes in corneal curvature during accommodation in chicks. Vision Res 27:241, 1987. 26. Schaeffel F and Howland HC: Corneal accommodation in chick and pigeon. J Comp Physiol A 160:375, 1987. 27. Worgul BV: Lens. In Ocular Anatomy, Embryology and Teratology, Jakobiec FA, editor. Philadelphia, Harper antf Row, 1982, p. 355. 28. Campbell MCW: Measurement of refractive index in an intact crystalline lens. Vision Res 24:409, 1984. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933377/ on 05/05/2017