Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
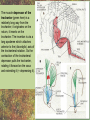
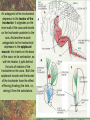
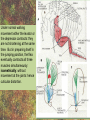
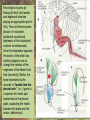
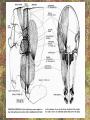
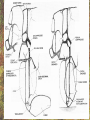
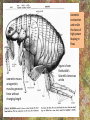
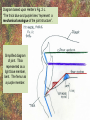
POWER AND SPRINGS Patek, S.N. et al. From bouncy legs to poisoned arrows: elastic movements in invertebrates. J. exp. Biol. 214: 1973-1980. Rothschild M. et al. 1973. The flying leap of the flea. Scientific American: 92Sutton G.P., Burrows M. 2011. Biomechanics of jumping in the flea. J. of exp. Biol. 214: 836-847. Montealegre-Z F. Et al. Generation of extreme ultrasonics in rainforest katydids. J. Exp. Biol. 209: 4923- Springs (materials storing energy by distortion) can be critical in animal locomotion and a much better way to catch prey. “… springs can provide outstanding efficiency and stability for continuous, long-term movements. These rhythmic movements include mechanisms ranging from flying fruit flies to singing katydids …(Montealegre-Z et al. 2006). Power ultrasonics achieved by a cuticlular amplifier scraper file mirror See Montealegre-Z 2006 Sphagniana sphagnorum Katydid wings are used to stridulate. A raised edge of the right wing (scraper) engages from below with a row of teeth lining the transversely running file vein of the left wing. The wings are moved against each other and the scraper moves tooth to tooth, shocking the glassy wing cells (mirror) into motion making sound. This species powers its way along a few teeth at a time using scraper elasticity. The scraper lodges behind a tooth the cuticle behind it bends storing energy; as the scraper slips free it strikes a number of teeth much more quickly than the overall wing movement. Power: rate of doing work • • • When we say a muscle’s effect is more powerful, we mean it is doing work faster. Work is force times distance [W=FS] . Force is mass times acceleration [F= mA] . Power is the rate at which work is done. A stridulating katydid accelerates its scraper mass through the distance between a number of file teeth, doing a certain amount of work: the faster it jumps between teeth the higher the power. There is a limit to the speed at which wing muscles can move the scraper. But higher tooth contact rates – more power -- can be achieved by using the springiness of the scraper cuticle. Forces arising in muscles can be stored in materials to be released at a later time. And at that later time they may give back their action faster, i.e., are more powerful. Arthropod cuticle is a composite material: chitin nanofibres (sugar chains) embedded in a protein matrix. Sometimes the protein is almost completely dominant giving resilin or ‘insect rubber’. • • • Resilin is a protein; it is protein chains joined by covalent bonds crosslinking between tyrosine amino acids (residues). “this amino acid lacks a side chain and is nonpolar, characteristics that prevent the formation of the sort of electrostatic bonds that would otherwise constrain the shape of the molecule. Freedom from constraint allows formation of random-coil chains.” This is the topographical basis of great elasticity; one gets a material that is a cord-like tangle of highly variable topography which can be easily distorted. Resilin is 50% water in its natural state. Young’s modulus • Young’s modulus is an index of the stiffness/elasticity of a material. Stiffness and elasticity are opposite ends of a ‘spectrum’. • E = change in stress/change in strain • Stiff materials have a high Young’s modulus and rubbery deformable materials have a low Young’s modulus. • Coral skeleton YM= 60000 very stiff • Mollusc nacre YM= 30000 very stiff • Abductin YM= 4 • Resilin YM=1.8 remarkably elastic. • See Vogel for a table of values: p. 298, 2nd edition Comparative Biomechanics Power amplification (Patek 2011 et al.) • Archer uses muscles slowly to bend a bow, storing elastic energy in the material of the bow and the bow string. When ready to shoot the tension in the bow and string is suddenly released.* “The arrow is shot far more rapidly than would have been possible if the archer had simply thrown the arrow.” • “The basic building blocks of archery and any fast biological system are an engine (the archer’s muscles) an amplifier (the springy bow and latch-like fingers) and tool (arrow). The unifying principle is called power amplification: the amplifier reduces the time to perform the engine’s work.” • Resilin is an amplifier. But it is not the exclusive basis of elasticity in cuticle. Cuticle can be relatively stiff and carry out amplification. The basic building blocks of …any fast biological system are an engine (horse’s muscles), an amplifier (the pole) and a ‘tool’, in this case the horse is the (inadvertent) tool or perhaps ‘payload’. Dark Side of the Horse by Samson • A bow should never be ‘shot’ without an arrow (Gordon 1976, p.92): this is because there is no way of getting rid of stored strain (elastic) energy. It is possible to shatter a bow in this way. The strain energy stored in the bent bow can no longer be dissipated in the kinetic energy of the arrow and is used to make cracks in the substance of the bow. • A makes a nice example of tension and compression surfaces in a ‘beam’ (Vogel 1988, p. 202). Bending an object of thickness has the effect of creating a gradient of tension on the outside of the curve and one of compression on the inside. In the middle there will be a neutral plane where there is no stress in either tension or compression (but not in shear). From this middle plane toward both surfaces, stresses increase. This means that central regions of structures contribute less strength; it is also the reason why bones can be (nearly) just as strong when hollow. Rothschild or Sutton & Burrows Sources Morphological features form of the flea: no wings, it’s flightless (its ancestors had wings); body extremely laterally compressed; greatly enlarged metathoracic legs; unidirectional body spines. Apply the course theme to this insect: thinki about where and how the animal lives. The cat flea Ctenocephalides felis A flea only 2 mm long can jump 200 mm, 100 times its own body length, the equivalent by a 6-foot human would be 600 feet! Accelerates from rest to 1 metre/sec in a distance of 0.4 mm; by fully extending its legs in about 8 milliseconds. Jumping is by power amplification. Energy is loaded (relatively slowly by isometric contraction of antagonistic muscles) into a pleural arch [amplifier] (the site of the winghinge in its flying insect ancestors?) and stored there in the rubbery protein. Once loaded the energy is held there as potential energy by latching sclerites, so no ongoing effort is needed by the flea. Release is by body width change. The leg segments extend, pushing down on the substrate and because of the stored energy they do this very very fast. So the ‘engine amplifier tool’ arrangement of Patek is: leg muscles as engine, resilin of pleural arch as amplifier, flea as tool (payload). The muscle depressor of the trochanter (green here) is a relatively long way from the trochanter; it originates on the notum, it inserts on the trochanter. The insertion is via a long apodeme which attaches anterior to the (dicondylic) axis of the trochanteral rotation. So the contraction of the trochanteral depressor pulls the trochanter, rotating it forward on the coxa and extending it (= depressing it). An antagonist of the trochanteral depressor is the levator of the trochanter. It originates on the inner wall of the coxa and inserts on the trochanter posterior to the axis. And another muscle antagonistic to the trochanteral depressor is the epipleural muscle: this inserts on the base of the coxa; on its contraction, as with the levator, it pulls behind the axis of rotation of the trochanter on the coxa. Both the epipleural muscle and the levator of the trochanter have the effect of flexing (levating) the limb, i.e., raising it from the substratum. Under normal walking movement either the levator or the depressor contracts: they are not shortening at the same time. But in preparing itself in the jumping position, the flea eventually contracts all three muscles simultaneously: isometrically: without movement at the joints: hence cuticular distortion. Flea begins its jump by flexing the limb (the levator and epipleural muscles playing an appropriate part in this). Then all three muscles [levator of trochanter, epipleural muscle and depressor of the trochanter] contract simultaneously. Since the depressor opposes the action of the other two, nothing happens now to change the relation of the segments of the flexed hind limb [isometric]. Rather the force expended by the muscles is "loaded into the pleural arch", i.e., it goes to compress the resilin pad located above the pleural plate, squeezing the resilin between the plate and the notum, deforming it. Some of the wonderful Illustrations by Rothschild explaining the same jumping process isometric contraction and resilin the basis of high-power leaping in fleas isometric means antagonistic muscles generate force without changing length figure is from Rothschild’s Scientific American article Sutton G.P., Burrows M. 2011. Biomechanics of jumping in the flea. J. of exp. Biol. 214: 836-847. In this paper the authors evaluate two hypotheses as to how the flea jump works: 1) Rothschild’s Hypothesis: ‘trochanters driven into ground’ 2) Bennet-Clark’s Hypothesis: ‘overall extension of leg speeded up’. They decide in favour of the latter: the trochanters do not touch the ground, rather the “expansion of the spring applied a torque about the coxo-trochanteral joint”; this torque is “carried through the femur and tibia” and finally resulted in a force applied to the ground by the hind tibia and tarsus. In other words the whole chain of leg segments extends with speed enhanced by the resilin at the leg base. Driving down the trochanters into the ground has some arguments against: the flea would be propelled more vertically and could have trouble making horizontal distance (though of course to reach from beside a standing dog vertical might be rather good). • Saltatorial: animal modified for leaping and jumping. Forelimbs small, hindlimbs enlarged, powerful muscles Sources • Heitler, W.J. 1974. The locust jump, specialisations of the metathoracic femoral-tibial joint. Journal of comparative Physiology 89: 93-104. • Burrows M., Sutton G.P. 2012. Locusts use a composite of resilin and hard cuticle as an energy store for jumping and kicking. J. exp. Biol. 215: 3501-3512. • Bayley T.G., Sutton G.P., Burrows M. 2012. A buckling region in locust hindlegs contains resilin and absorbs energy when jumping or kicking goes wrong. J. exp. Biol. 215: 1151-1161. • See : JEB highlight by Kathryn Knight same issue: Buckling zone protects locust legs Orthoptera Species File The migratory locust from Wikkipedia Video of jumping locust Anatomy and leverage of locust metathoracic leg femorotibial joint • • • • • • Flexion/extension: joint angle goes from 0 to 150⁰ Dicondylic joint: condyles [= pivot pegs] are part of femur, the sockets part of tibia. Lump [apodeme] (dome-shaped inflection) can lock via forked ‘pocket’, part of flexor apodeme. Extensor of tibia, flexor of tibia are antagonists, yet their muscles pull in nearly the same direction. The extensor works as a 1st class lever; the flexor is 3rd class. Lever arm has a crooked shape which also affects force direction. muscles The angles of ‘force in’ change as not shown, just apodemes the tibia moves from completely flexed to maximally extended (see below). Geometry/anatomy of the joint see Heitler’s Fig. 1: find the parts on his figure. • • • a) Fully flexed locked joint: bifurcate pocket of apodeme of flexor sits astride the lump. Note apodeme of extensor and two accessory muscles. b) Lock is disengaged and joint extended midway; flexor apodeme is now riding pulleylike on top of the lump. c) The two condyles of the dicondylic joint (sockets in tibia) seen dorsally along with the lump. Diagram based upon Heitler’s Fig. 2 c. “The thick blue and purple lines “represent a mechanical analogue of the joint structure”. Simplified diagram of joint. Tibia represented as a light blue member, bent. The femur as a purple member. Paradox [Logical statement that apparently contradicts itself.] For cuticular amplification of the jump of a grasshopper there must be isometric muscle contraction: a smaller flexor muscle must hold the tibia flexed against the force of a much larger extensor muscle.” Levers are the basis for solving this paradox • • A moment of force about an axis is the product of force magnitude and the perpendicular distance to the axis (Newton metres). To keep the two leg muscles in isometric contraction at 5⁰, the moment of force of the weaker muscle must be made equal to the moment of force of the stronger. Paradox resolved: The force advantage of the flexor muscle is different at different angles of flexion of the femorotibial joint. When the angle of flexion is less than 5⁰ (top) the moment of the flexor muscle is better than that of the extensor. It is better for two reasons Better distance from axis Better pulling angle • With joint angle at 5⁰ (top), the apodeme of the flexor makes an angle with the effort arm of the lever of almost 90⁰; because it rides up over the 'lump'. The lump functions as a pulley and changes the ‘force-in’ direction, making it nearly 90⁰ to the tibia; by contrast, at 5⁰, the stronger extensor has a force-in direction at a very poor angle of 6⁰. Perpendicular distances (lever arms) and angle of force application are in favour of the smaller muscle, so the moments of force for the two antagonists can be equal when leg is flexed. • So isometric contraction becomes possible as a way for both muscles together to store energy in cuticle, i.e., elastic distortion gets involved. Resilin regions of exoskeleton in the neighbourhood of the joint store elastic energy: they are called semilunar processes. Just like the flea this stored energy can be stored relatively slowly then made available much more quickly so achieving power amplification. Think of the bow shot without an arrow that fractures: shock aborbers in the cuticle of the grasshopper’s tibia are material adaptations to dissipate energy of fracture. • Energy of a bow shot without an arrow or a kick that misses its target is dissipated by a specialized proximal region of the tibia. There is resilin in this region, revealed as a band that fluoresces blue under UV illumination (with appropriate filters to confirm resilin’s identity). There are also special campaniform sensilla in the cuticle (proprioceptors, mechanoreceptors) that monitor the buckling. “The features of the buckling region show that it can act as a shock absorber as proposed previously [by Heitler] when jumping and kicking movements go wrong.” Photographs of the distal femur of the right hind leg taken under white and UV epi-illumination, and then combined. Burrows M , Sutton G P J Exp Biol 2012;215:3501-3512 ©2012 by The Company of Biologists Ltd Locking to minimize energy cost while waiting to jump • Quoting Heitler: “Once the tibia is fully flexed a lock is engaged which can hold the tibia in this position against the developing extensor tension. Just proximal to its insertion onto the tibia the strap-like flexor tendon effectively bifurcates into strands which insert on either side of the tibia, leaving a strengthened pocket of connective tissue in the middle. At any angle of extension greater than about 5 degrees the flexor tendon rides on top of the femoral lump, which thus acts as … [a] pulley…. As the tibia approaches the fully flexed position …, the two arms of the tendon slide down on either side of the lump, fitting into grooves at its base, while the lump fits snugly into the connective tissue pocket in the middle (Fig. 1a). In this position the tibia is now locked against the femur, and considerable extensor tension can be developed without the tibia moving.” • With the lock in place the muscles can bend the exoskeleton and resilin can begin to contribute its power amplification ability. Special shock aborbers for kicks gone astray • Gordon’s comments about bows needing arrows apply readily to locust kicks that go wrong . • From the abstract of Bayley et al. 2012. “If a hindleg of a locust slips during jumping, or misses its target during kicking, energy generated by the two extensor tibiae muscles is no longer expended in raising the body [jumping] or striking a target.” • Bayley found a special region of the proximal hind tibia that is adapted to buckle under these conditions through the agency of resilin. • Both muscles work with a poor mechanical advantage due to constraints of body shape* but a good speed-distance advantage – its good to have speed and distance working for you when trying to jump. • “Myograms show that there is co-activation of the extensor and flexor muscles during the pre-jump crouch...” • “Complete extension of the tibia takes some 20 ms... to develop peak power ...in the jump the extensor muscle must first build up tension isometrically.” The two antagonists simultaneously contract, but there is no movement at the joint. • “The extensor muscle is much larger and occupies the greater part of the femoral volume.” Its pinnate [angled like a feather] fibres are many more and short; an arrangement which enables the muscle to develop a very large force at its tendon [apodeme], though moving through a shorter distance. The flexor muscle, by contrast, is composed of long thin parallel fibres, and is of comparatively small cross-sectional area. “This weak muscle must hold the tibia flexed against the full force of the powerful extensor muscle.” This is an apparently contradictory statement. • How is a stalemate achieved during isometric contraction? The answer: special adaptations of leverage at the joint and the flexor apodeme lock. Burrows M. & Sutton G.P. 2012. Locusts use a composite of resilin and hard cuticle as an energy store for jumping and kicking. J. exp. Biol. 215: 3501-3512. • Burrows & Sutton have explained where the energy of isometric contraction is stored. It goes into paired semilunar processes of the femur, located at the sides of its distal extremity, lateral to where the condyles protrude into the sockets of the tibia. semilunar process Photographs of the femoro-tibial joint of a right hind leg of an adult locust “Externally visible resilin was compressed and wrinkled as a semi-lunar process was bent. It then spr[a]ng back to restore the semi-lunar process to its original shape. “It is suggested that composite storage devices that combine the elastic properties of resilin with the stiffness of hard cuticle allow energy to be stored by bending hard cuticle over only a small distance and without fracturing. In this way all the stored energy is returned and the natural shape of the femur is restored rapidly so that a jump or kick can be repeated.” ©2012 by The Company of Biologists Ltd Burrows M , Sutton G P J Exp Biol 2012;215:3501-3512 Burrows & Sutton 2012 Photographs of the femoro-tibial joint of a right hind leg of an adult locust. • • “The inside surface of a semi-lunar process consists of a layer of resilin, particularly thick along an inwardly pointing ridge and tightly bonded to the external, black cuticle.” There is (shown by imaging [movie]) distortion/bending in all three dimensions during the isometric contraction. Wikki: A recombinant form of the resilin protein of… Drosophila melanogaster, pro-resilin, was synthesized in 2005 by expressing a part of the fly gene in the bacterium Escherichia coli. It is expected to have many applications in the athletic footwear, medical, microelectronics and other industries. Burrows M , Sutton G P J Exp Biol 2012;215:3501-3512 ©2012 by The Company of Biologists Ltd The buckling region of the right hind-tibia viewed under white and UV illumination. Bayley T G et al. J Exp Biol 2012;215:1151-1161 ©2012 by The Company of Biologists Ltd