Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
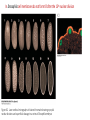
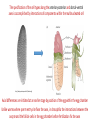
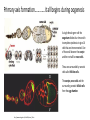
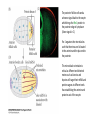
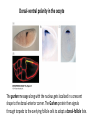
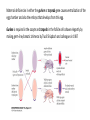
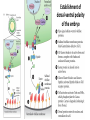
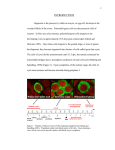
Axis Specification and Patterning I Syncytial specification in the Drosophila embryo We know more about the genetics of Drosophila melanogaster than any other multicellular organism thanks to the pioneering studies initiated by Thomas Hunt Morgan But Drosophila proved to be a difficult organism to study embryology. The Drosophila embryo was neither transparent so that it could be observed microscopically nor was it large enough to be manipulated experimentally. It was not until the advent of molecular biology, which facilitated the identification and modification of genes and RNAs did it become possible to relate the genetics of Drosophila to its development Fertilization Cleavage http://www.wisegeek.com/what-is-the-fertilization-process.htm Specification of early embryos happen by cytoplasmic determinants stored in the oocyte. Cell membranes formed during cleavage partition the cytoplasm into blastomeres. In Drosophila cell membranes do not form till after the 13th nuclear division Figure 6.2 Laser confocal micrographs of stained chromatin showing syncytial nuclear divisions and superficial cleavage in a series of Drosophila embryos The specification of the cell types along the anterior-posterior and dorsal-ventral axes is accomplished by interactions of components within the multinucleated cell http://willy.supatto.perso.sfr.fr/3dentier.gif Axial differences are initiated at an earlier stage by position of the egg within the egg chamber Unlike worms where sperm entry site fixes the axes, in drosophila the interactions between the oocyte and the follicle cells in the egg chamber before fertilization fix the axes Genes the pattern the Drosophila body plan A powerful “forward genetics” approach in the early 1990s spearheaded by Christiane Nϋsslein-Volhard and Eric Weischaus was employed to identify the genes involved in shaping the larval and adult Drosophila. Randomly mutagenized flies were screened for mutations that disrupted the normal body plan. http://biology.kenyon.edu/courses/biol114/Chap13/Chapter_13A.html The genes involved in these mutant phenotypes were cloned and characterized by many different groups worldwide Primary axis formation………….it all begins during oogenesis A single female germ cell the oogonium divides four times with incomplete cytokinesis to give 16 cells that are interconnected. One of these will become the oocyte and the rest will be nurse cells. These are surrounded by somatic cells called follicles cells. The oocyte, nurse cells and the surrounding somatic follicle cells form the egg chamber. http://www.zoology.ubc.ca/~bio463/lecture_13.htm Anterior-posterior polarity in the oocyte The follicle cells surrounding the oocyte are initially uniform but this is broken by two signals organized by the oocyte nucleus involving the same gene gurken The gurken RNA is made in nurse cells but transported to the oocyte and localized specifically between the posteriorly localized oocyte nucleus and the follicle cells. The Gurken protein is translated and activates the Torpedo receptors on the follicle cells giving them a posterior identity. The posterior follicle cells send a unknown signal back to the oocyte which brings the Par-1 protein to the posterior edge of cytoplasm (Green signal in C). Par-1 organizes the microtubules such that the minus end is located to the anterior and the plus end to the posterior. This microtubule orientation is critical as different end-directed motors such as kinesins and dyneins will target their mRNA and protein cargoes to different ends thus establishing the anterior and posterior axis of the oocyte. The cytoskeletal rearrangements lead to localization of maternal messages such as bicoid and nanos mRNAs. Also the oocyte volume increases and the oocyte nucleus is pushed by growing microtubules to an anterior-dorsal location. Dorsal-ventral polarity in the oocyte The gurken message along with the nucleus gets localized in a crescent shape to the dorsal-anterior corner. The Gurken protein then signals through torpedo to the overlying follicle cells to adopt a dorsal-follicle fate. Maternal deficiencies in either the gurken or torpedo gene causes ventralization of the egg chamber and also the embryo that develops from this egg. Gurken is required in the oocyte and torpedo in the follicle cells shown elegantly by making germ-line/somatic chimeras by Trudi Shϋpbach and colleagues in 1987 Reciprocal interactions between the oocyte and egg chamber (follicle cells) specify the dorsal-ventral polarity of the embryo Establishment of dorsal-ventral polarity of the egg chamber (follicle cells) Establishment of dorsal-ventral polarity of the embryo Cell. 1992 Feb 7;68(3):429-40. Multiple extracellular activities in Drosophila egg perivitelline fluid are required for establishment of embryonic dorsal-ventral polarity. Stein D1, Nüsslein-Volhard C. Embryonic dorsal-ventral polarity is defined within the perivitelline compartment surrounding the embryo by the ventral formation of a ligand for the Toll receptor. Here (as demonstrated by the transplantation of perivitelline fluid) are found three separate activities present in the perivitelline fluid that can restore dorsal-ventral polarity to mutant easter, snake, and spatzle embryos, respectively. These activities are not capable of defining the polarity of the dorsal-ventral axis; instead they restore structures according to the intrinsic dorsal-ventral polarity of the mutant embryos. They appear to be involved in the ventral formation of a ligand for the Toll protein. This process requires serine proteolytic activity; the injection of serine protease inhibitors into the perivitelline space of wild-type embryos results in the formation of dorsalized embryos (Stein, 1992). Injection experiments involving the use of dominant active Easter (Chasan, 1992) and Snake, as well as injection of perivitelline (PV) fluid from dorsal mutant embryos into gd mutant embryos, have lead to production of ventral elements at the site of injection, rather than in the normal ventral region (Stein, 1992). These data suggest that D/V polarity is established by asymmetric presentation of the Toll ligand to the oocyte. PV fluid from dorsal mutant embryos (thought to be depleted of Spätzle ligand because of the presence of the Toll receptor) can rescue D/V polarity in snake and easter mutant embryos. This same PV fluid cannot restore normal ventral structures to gd embryos. In contrast, injection of PV fluid from Toll mutant embryos (thought to contain active Spätzle ligand) into gd embryos produces ventral structures at the site of injection (Stein, 1992). The same fluid injected into snake or easter embryos produces embryos with normal polarity, independent of the site of injection (Stein, 1992). Generation of Dorsal-ventral pattern in the embryo by Dorsal the morphogen 16 cells with the highest concentration of dorsal are the ones that generate the mesoderm.