Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
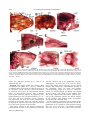
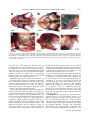
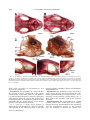
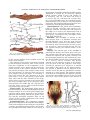
JOURNAL OF MORPHOLOGY 271:1352–1365 (2010) Comparative Cranial Osteology of Fossorial Lizards From the Tribe Gymnophthalmini (Squamata, Gymnophthalmidae) Juliana G. Roscito* and Miguel T. Rodrigues Departamento de Zoologia, Instituto de Biociências, Universidade de São Paulo, São Paulo-SP, Brazil ABSTRACT Squamates (lizards, snakes and amphisbaenians) are represented by a large number of species distributed among a wide variety of habitats. Changes in body plan related to a fossorial habit are a frequent trend within the group and many morphological adaptations to this particular lifestyle evolved convergently in nonrelated species, reflecting adaptations to a similar habitat. The fossorial lifestyle requires an optimal morphological organization for an effective use of the available resources. Skeleton arrangement in fossorial squamates reflects adaptations to the burrowing activity, and different degrees of fossoriality can be inferred through an analysis of skull morphology. Here, we provide a detailed description of the skull morphology of three fossorial gymnophthalmid species: Calyptommatus nicterus, Scriptosaura catimbau, and Nothobachia ablephara. J. Morphol. 271: 1352–1365, 2010. Ó 2010 Wiley-Liss, Inc. KEY WORDS: body plan; fossoriality; gymnophthalmidae; skull morphology INTRODUCTION Squamates comprise approximately 8,400 species and represent a diverse group of animals, which exhibit several adaptative phenotypes related to the biogeographic context in which each group evolved. These adaptations range from differences in habitat use and diet to differences in size and body plans (Vitt et al., 2003). Convergent evolution of morphological traits is an often observed phenomena within the group (Wiens and Slingluff, 2001), the most obvious being the repeated evolution of a snake-like body plan related to fossorial habits, involving body lengthening and limb reduction (Brandley et al., 2008; Gans, 1975; Greer, 1991; Wiens et al., 2006). According to Wiens et al. (2006), a snake-like body plan has evolved independently about 25 times, and even in the same family, (e.g., anguids, gymnophthalmids, and scincids) intermediate or extreme cases of body elongation and limb reduction are found among closely related species or within the same genus (Pellegrino et al., 2001; Shapiro, 2002; Skinner et al., 2008; Wiens and Slingluff, 2001). The transition to fossoriality reflects an adaptative complex, which involves morphological modifiÓ 2010 WILEY-LISS, INC. cations in skeletal patterns related to elongation of the body, reduction/loss of limb bones, and skull modifications, such as loss of elements and excessive growth and robustness of others, associated with skull consolidation (Greer, 1991; Lee, 1998; Rieppel, 1996; Tarazona et al., 2008). Gymnophthalmidae (Estes et al., 1988) comprises an assemblage of 42 genera of small lizards distributed throughout Central and South America (Rodrigues et al., 2007), many of which are fossorial burrowers with elongated trunks and reduced or lost limbs. These modifications toward the evolution of snake-like body plans have originated independently several times during the evolutionary diversification of the family (Pellegrino et al., 2001). Within the Gymnophthalmidae, a monophyletic group of nine genera which is taxonomically known as the tribe Gymnophthalmini (Pellegrino et al., 2001; Rodrigues, 1991, 1995; Rodrigues and dos Santos, 2008), shows a clear and noticeable evolutionary trend related to a transition from a lacertiform to a serpentiform body plan. The basal genera (Tretioscincus, Micrablepharus, Gymnophthalmus, Procellosaurinus, Vanzosaura, and Psilophthalmus) are diurnal, lizard-like in shape, and have well-developed limbs and digits and tails longer than body length, while the derived genera (Nothobachia, Scriptosaura, and Calyptommatus) are fossorial, show adaptations to life in sand, and are characterized by a snake-like shape, with extremely elongated body, tails shorter than body, Contract grant sponsors: Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) and Conselho Nacional de Desenvolvimento Cientı́fico e Tecnológico (CNPq). *Correspondence to: Juliana G. Roscito, Departamento de Zoologia, Instituto de Biociências-Universidade de São Paulo, Cidade Universitária, Rua do Matão, Trav. 14, no. 321, São Paulo-SP, Brazil CEP 05508-090. E-mail: [email protected] Received 10 March 2010; Revised 21 April 2010; Accepted 5 May 2010 Published online 26 August 2010 in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/jmor.10878 CRANIAL OSTEOLOGY OF FOSSORIAL GYMNOPHTHALMIDS and limbs reduced or absent; Calyptommatus is the only nocturnal genus of this radiation. The digit pattern in the Gymnophthalmini is represented by a progressive reduction/loss of phalanges and digits along its evolutionary history. The six lizard-like genera all have four well-developed limbs. The forelimbs of Tretioscincus, Micrablepharus, Gymnophthalmus, Procellosaurinus, Vanzosaura, and Psilophthalmus, all have three, four, five, and three phalanges in Digits II, III, IV, and V respectively, and in Digit I is observed a progressive reduction in the number of phalanges, with two small phalanges in Tretioscincus, one vestigial phalange in Micrablepharus and Gymnophthalmus, and no phalanges in the three latter genera. The hind limbs have five digits with a phalangeal formula of 2:3:4:5:4 (Rodrigues, 1995). In contrast, the three snake-like genera show more severe degrees of limb reduction: Nothobachia has a styliform forelimb and a hind limb with two small digits and Scriptosaura and Calyptommatus have no visible forelimb and a styliform hind limb. These three genera have bodies much more elongated than those of the other six lacertiform genera, showing a snout-vent length (SVL) larger than tail length (due to a increase in the number of presacral vertebrae; Rodrigues, 1995). Also, in the fossorial and snake-like genera, the eyes are reduced, there is no external ear opening, the snout is sharp, and the skull is more compact than that of the diurnal and nonfossorial genera, these characters reflecting the burrowing lifestyle. Furthermore, Calyptommatus shows unique features of some skull bones not present in any of the other fossorial genera (Rodrigues, 1995). In this article, we provide a detailed comparative description of the adult skeletal morphology of the skull and axial skeleton in Calyptommatus nicterus, Scriptosaura catimbau, and Nothobachia ablephara and discuss the morphological adaptations that reflect the different degrees of fossoriality observed in these species. MATERIAL AND METHODS The description is based on eight adult specimens of Calyptommatus nicterus (Rodrigues, 1991) obtained at Vacaria, State of Bahia, Brazil (Museu de Zoologia da Universidade de São Paulo [MZUSP] 79549, MZUSP 79550, MZUSP 79551, MZUSP 79552, MZUSP 79553, MZUSP 79554, Miguel Trefaut Rodrigues [MTR] 886837, and MTR 16767), two adults of Scriptosaura catimbau (Rodrigues and dos Santos, 2008) obtained at Parque Nacional do Catimbau, Buique, State of Pernambuco, Brazil (MTR 16777 and MTR 16778), and five adults of Nothobachia ablephara (Rodrigues, 1984) obtained at Alagoado, State of Bahia, Brazil (MZUSP 79573, MZUSP 79574, MTR 16774, MTR 16775, and MTR 10378). The specimens are deposited in the collection of MTR and the MZUSP. All specimens were cleared and double stained for bone and cartilage following an adaptation of protocols from Potthoff (1984), Taylor and Van Dyke (1985), Song and Parenti (1995), and Springer and Johnson (2000). The material was examined using an Olympus stereomicroscope. Digital pictures, with corresponding scales, were taken with a digital camera attached to the stereo- 1353 microscope. Schematic drawings were made using the digital images as shapes. For all specimens, SVL was measured, and head length was measured from the anterior tip of the snout to the back of the occipital condyle. The anatomical terminology follows Bell et al. (2003), Torres-Carvajal (2003), Tarazona et al. (2008), Evans (2008), and Guerra and Montero (2009). Abbreviations are as follows: 1ctb, first ceratobranchial; 2ctb, second ceratobranchial; 1epb, first epibranchial; 2epb, second epibranchial; aaf, anterior auditory foramen; al.p, alar process; alp.f, anterolateral process of the frontal; alp.m, anterolateral process of the maxilla; amp.crn, anteromedial process of the coronoid; amp.m, anteromedial process of the maxilla; ang, angular; ap.pm, alveolar plate of the premaxilla; asc, anterior semicircular canal; bh, basihyal; bo, basioccipital; bpt.p, basipterygoid process; ch.g, choanal groove; cm, columella; crn, coronoid; d, dentary; dp.p, descending process of the parietal; ecp, ectopterygoid; eph, epihyal; ept, epipterygoid; exc.a, exoccipital area; f, frontal; fcf, facial foramen; fo, fenestra ovalis; fp.m, facial process of the maxilla; ftb, frontoparietal tab ; gh, glossohyal; hc, hyoid cornu; hsc, horizontal semicircular canal; j, jugal; lgc, lagenar cavity; m, maxilla; m.ps, maxillary palatal shelf; mp.pm, maxillary process of the premaxilla; n, nasal; np.pm, nasal process of the premaxilla; obf, orbitosphenoid; oc, occipital condyle; op.a, opisthotic area; or, occipital recess; p, parietal; paa complex, prearticular-articular complex; paf, posterior auditory foramen; pbs, parabasisphenoid; pf, postfrontal; pl, palatine; plp.f, posterolateral process of the frontal; pm, premaxilla; pm.ps, premaxillary palatal shelf; pmp.crn, posteromedial process of the coronoid; po, postorbital; pof, postorbitofrontal; pp, postparietal process; ptf, posttemporal fenestra; prf, prefrontal; prp, paroccipital process; psc, posterior semicircular canal; pss, planum supraseptale; pt, pterygoid; q, quadrate; qp.pt, quadrate process of the pterygoid; so, supraoccipital; spl, splenial; spm, septomaxilla; sq, squamosal; sra, surangular; st, supratemporal; tgn, trigeminal notch; tr, trabecula; v, vomer; vf, vagus foramen; vtc, vestibular cavity; XII, foramina for the hypoglossal nerve roots. RESULTS General Features of the Skull The skulls of Calyptommatus nicterus (Fig. 1), Scriptosaura catimbau (Fig. 2) and Nothobachia ablephara (Fig. 3) measure 6 mm in length, corresponding approximately to 10.5, 11, and 11.5% of the SVL (average SVL of 5.7, 5.5, and 5.2 cm, respectively). All are relatively elongated, but the skull of C. nicterus is much wider than that of the other two species; the snout is sharp in all three. The orbital area of C. nicterus is smaller than that observed in the other two species, and its palate and the lateral wall of the skull posterior to the orbits are completely closed by extensions of the bones that form these regions; in S. catimbau, however, a small ventral projection from the parietal bone is observed anterior to the epipterygoid, in contrast with the almost imperceptible projection in N. ablephara. In S. catimbau and N. ablephara, no complex articulations are observed between elements, whereas the bones of the skull of C. nicterus are tightly articulated with each other, making the skull very compact. Premaxilla. The premaxilla forms the anterior tip of the snout. It comprises an alveolar portion and a nasal process. The alveolar plate is thicker in C. nicterus and thinner in N. ablephara, with that of S. catimbau falling in between; it carries Journal of Morphology 1354 J.G. ROSCITO AND M.T. RODRIGUES the dorsal margin of the nares and to overlap the nasals and the anterior margin of the frontal posteriorly. In C. nicterus, the nasal process of the premaxilla overlaps only a minor portion of the anteromedial margin of the nasals (Fig. 1A), whereas in S. catimbau and N. ablephara, it covers a greater portion of them (Figs. 2A and 3A). Maxilla. The maxilla is a large element forming most of the lateral surface of the skull anterior to the orbit. The anterior end is represented by a bifurcated premaxillary process, divided into a slender anterolateral part that articulates with the maxillary process of the premaxilla and a larger anteromedial part that contacts the dorsal surface of the palatal shelf of the premaxilla and forms the ventral floor of the nares. Ventrally, the anteromedial process is continued by a well-developed maxillary palatal shelf, which is aligned with the lateral margin of the vomer Fig. 1. C. nicterus, drawing of the skull in dorsal (A), lateral (B). and ventral (C) views. Sutures are represented by dashed lines; shaded area in C represents roof of the skull. Scale bar 5 1 mm. 8–9, 9, and 7 unicuspid teeth, respectively, positioned along its ventral margin (Figs. 1C and 4A; 3C and 6B; 2C and 5B, respectively). Dorsally and posterior to the teeth row, the V-shaped palatal shelf contacts the vomer medially and the anteromedial portion of the premaxillary process of the maxilla. Laterally, the maxillary process of the premaxilla articulates with the anterolateral portion of the premaxillary process of the maxilla. In C. nicterus and S. catimbau, this maxillary process is bifurcated into dorsal and ventral rami, supporting the anterior end of the maxilla between them (Fig. 5C); in N. ablephara, the maxillary process is simple and contacts only the internal and ventral surface of the anterior end of the maxilla (Fig. 6G). The broad nasal process of the premaxilla projects from the medial part of the alveolar plate to overlap part of the anterior margin of the frontal. It is slender anteriorly, then widens to form Journal of Morphology Fig. 2. S. catimbau, drawing of the skull in dorsal (A), lateral (B), and ventral (C) views. Shaded area in C represents roof of the skull. Scale bar 5 1 mm. CRANIAL OSTEOLOGY OF FOSSORIAL GYMNOPHTHALMIDS Fig. 3. N. ablephara, drawing of the skull in dorsal (A), lateral (B), and ventral (C) views. Scale bar 5 1 mm. and ends at the region of the choana, being overlapped dorsally by the palatine and contacting the external margin of the ectopterygoid (Figs. 1C, 2C, and 3C); the palatine process of the prefrontal stands on the dorsal surface of the shelf. The maxillary alveolar region bears six unicuspid teeth in C. nicterus (Figs. 1C and 4A), eight in N. ablephara (Figs. 2C and 6B) and 9–10 in S. catimbau (Figs. 2C and 5B). The facial process of the maxilla forms the posterior margin of the naris and contacts anterodorsally and the anterolateral process of the frontal in N. ablephara (Fig. 3B) and S. catimbau (Fig. 5A); in C. nicterus, the anterolateral process of the frontal is completely overlapped by the nasal, and thus the facial process of the maxilla contacts directly the dorsal extremity of the nasal (Fig. 1B). Posterodorsaly, the facial process contacts the prefrontal, overlapping its anterior process, and posteroventrally, the maxillary orbital process underlies the anterior process of the jugal. 1355 Nasal. The nasal is approximately trapezoidal in shape and is smaller in C. nicterus compared with the other two species. It meets the nasal process of the premaxilla medially, which completely separates it from its counterpart at the midline, and posteriorly (Fig. 1A). In N. ablephara and S. catimbau (Figs. 2A and 3A), in which an anterolateral process of the frontal is exposed, the nasal meets this process posteriorly and laterally. The anterior and lateral margins of the nasal contact the facial process of the maxilla and form the dorsal margin of the nares. Prefrontal. The prefrontal forms the anterior and part of the dorsal margins of the orbit, protecting it anteriorly through strong connections with neighboring elements. This element is composed of an anterior process, represented by a broad sheet of bone that is overlapped by the facial process of the maxilla, a posterodorsal process contacting the anterior and lateral margins of the frontal and extending up to the middle of the orbit, and a ventral palatine process contacting the anterior border of the palatine medially and the dorsal surface of the maxillary palatal shelf laterally. A huge lacrimal foramen is observed between the internal wall of the facial process of the maxilla and the ventral process that rests on the maxillary palatal shelf. The ventral end of the prefrontal meets the anterior process of the jugal. The lacrimal flange is represented by a posteriorly directed projection, quite broad in N. ablephara (Fig. 3B) and smaller in S. catimbau (Fig. 2B), when compared with that of the former; in C. nicterus, no lacrimal flange is observed (Figs. 1B and 4G). In C. nicterus, the posterodorsal process of the prefrontal is placed beneath the lateral margin of the frontal and is, thus, not visible in dorsal view. Both this process and the frontal participate in the formation of the dorsal margin of the orbit. The process is clearly seen, though, lateral to the margin of the frontal in the other two species, forming alone the anterior part of the dorsal margin of the orbit (Fig. 6A). Jugal. The jugal is slightly sigmoidal in shape. It forms the ventral and part of the posterior margins of the orbit. Its anterior maxillary process contacts the ventral end of the prefrontal and is supported ventrally by the orbital process of the maxilla; its dorsal temporal process contacts the ventral margin of the postorbital. Medially, the jugal contacts the lateral margin of the ectopterygoid. In N. ablephara, the jugal is a slender element with relatively sharp extremities and a more robust middle portion (Fig. 6E). In C. nicterus, the jugal is stouter and triradiate in shape, with a robust posterior free process (Figs. 4D,G). In S. catimbau, the jugal resembles that of N. ablephara but has a small protuberance at the same place Journal of Morphology 1356 J.G. ROSCITO AND M.T. RODRIGUES Fig. 4. C. nicterus, cleared and stained skulls. (A) Ventral view of the anterior portion of the skull. (B) Posterolateral view of the braincase; anterior to the left. (C) Braincase in lateral view; quadrate not represented. (D) Detail of the jugal in lateral view; anterior to the right. (E) Medial surface of the postorbitofrontal in lateral view; anterior to the left. (F) Descending process of the parietal and alar process of the prootic in lateral view; anterior to the right. (G) Detail of the lateral surface of the skull anterior to the orbit; anterior to the right. (H) Frontal in ventral view showing its well-developed cristae cranii (indicated by asterisk); anterior to the left. Scale bars 5 0.5 mm. where the posterior process of C. nicterus is observed (Fig. 2B). Frontal. The frontal forms the anterior skull roof between the orbits. In C. nicterus, which has a reduced orbit, the frontal is wider than those of N. ablephara and S. catimbau, where the frontal is narrower and longer. Its anterolateral margin bears an anterolateral process with a distinct facet for the nasal, by which it is completely overlapped dorsally in the articulated skull of C. nicterus (Fig. 1A). In N. ablephara and S. catimbau, the nasal does not cover the entire anterolateral process of the frontal (Figs. 2A and 3A), leaving it exposed to separate the nasal from the dorsal tip of the facial process of the maxilla. The lateral margin of the frontal contacts the posterodorsal process of the prefrontal anteriorly Journal of Morphology and the anterior end of the postfrontal (or postorbitofrontal, as is the case in C. nicterus) posteriorly. The cristae cranii originate at each lateral margin and contact the posterodorsal process of the prefrontal. They are short and slightly curved medially, except in C. nicterus, where the cristae are well developed and highly curved (but do not contact each other at the midline; Fig. 4H), being supported by the ventral palatine process of the prefrontal and by the dorsal surface of the palatine. The cristae cranii encapsulate and protect the olfactory bulb. At the posterior end of the lateral margin of the frontal, a small posterolateral process contacts the postfrontal. In N. ablephara, this process is sharp (Fig. 3A), in S. catimbau, it is slightly rounded (Fig. 2A), and in C. nicterus, there is no such pro- CRANIAL OSTEOLOGY OF FOSSORIAL GYMNOPHTHALMIDS 1357 Fig. 5. S. catimbau, cleared and stained skulls. (A) Anterior part of the skull in dorsal view; anterior to the left. (B) Ventral view of the anterior part of the palate; anterior to the left. (C) Articulation between premaxilla and maxilla in lateral view; anterior to the right. (D) Lateral view of the middle portion of the skull, with detail of the descending process of the parietal; anterior to the right. (E) Braincase in lateral view; quadrate not represented. (F) Orbitosphenoid in dorsal view; anterior to the top. Scale bars 5 0.5 mm; except in C in which scale bar 5 0.25 mm. cess (Fig. 1A). This process is distinct from the posteromedial process (frontoparietal tab) next to it. These tabs are longer in N. ablephara and S. catimbau than in C. nicterus and represent broad areas of contact with the corresponding tabs of the parietal, forming an ample area of articulation between these two elements. The frontoparietal suture between the tabs is slightly wavy. Parietal. The parietal forms the posterior roof of the skull and is longer and wider than the frontal. It articulates anteriorly with the frontal and contacts, at its anterolateral margin, the postfrontal (or postorbitofrontal in C. nicterus); the posterolateral margin of the parietal forms the dorsal margin of the supratemporal fenestra. A postparietal process of the parietal extends posteriorly along the anterior margin of the supraoccipital transverse crest, overlapping the otooccipital complex and contacting the supratemporal ventrally. A slender posttemporal fenestra is present in C. nicterus and N. ablephara, separated at the midline by a short anterior projection of the supraoccipital and by the short ascending process of the tectum synoticum (not shown in Figs. 1 and 3), and delimited anteriorly by the posterior margin of the parietal (Figs. 1A and 3A). These fenestra are not present in the two specimens of S. catimbau an- alyzed, where the posterior margin of the parietal overlaps the supraoccipital (Fig. 2A). At the posterior margin of the parietal of N. ablephara and C. nicterus, a small pit is observed for the anchoring of the ascending process of the tectum synoticum. A lateral descending process originates from the midpoint of the lateral margin of the parietal. In N. ablephara it is barely noticeable and does not reach the dorsal tip of the epipterygoid (Figs. 3B and 6F); in S. catimbau, the process is longer and is located at the front of the dorsal tip of the epipterygoid (Figs. 2B and 5D). The highest degree of development of this process is observed in C. nicterus, where it completely closes the lateral wall of the skull from the front of the epipterygoid up to the posterior margin of the orbit (Figs. 1B and 4F), with a ligamentous connection to the pterygoid. Postfrontal. The postfrontal is a short element, located lateral to the frontoparietal suture. It is slightly triangular in shape, with short anteromedial and anterolateral ends and a longer posterior extremity. The postfrontal forms part of the posterodorsal margin of the orbit, contacting the frontal and parietal dorsally (anterior and posteriorly, respectively) and the postorbital ventrally. The bone is not fused in Nothobachia and Scriptosaura Journal of Morphology 1358 J.G. ROSCITO AND M.T. RODRIGUES Fig. 6. N. ablephara, cleared and stained skulls. (A) and (B) Anterior part of the skull in dorsal (A) and ventral (B) views; anterior to the right. (C) Braincase in medial view; anterior to the left. (D) Braincase in external lateral view; anterior to the right. (E) Orbital region in lateral view; anterior to the right. (F) Lateral view of the middle part of the skull; anterior to the right. (G) Articulation between premaxilla and maxilla in lateral view; anterior to the top right. Scale bars from A to F 5 0.5 mm; scale bar for G 5 0.25 mm. (Figs. 2A,B and 3A,B); in Calyptommatus, it is fused to the postorbital. Postorbital. The postorbital is a long and slender element. It forms a small part of the posterodorsal margin of the orbit, ventral to the postfrontal, and extends posteriorly to meet the squamosal, with which it forms the inferior bar of the supratemporal fenestra. It contacts the postfrontal anterodorsally and the dorsal surface of the anterior process of the squamosal posteriorly. In C. nicterus, a single stout element is observed, the postorbitofrontal (Figs. 1B and 4E), corresponding to the embryonic fusion of the postJournal of Morphology frontal with the postorbital (Roscito and Rodrigues, unpublished data). Squamosal. The squamosal is long and slender, with a sharp anterior process contacting the postorbital (postorbitofrontal in Calyptommatus) and a ventrally curved posterior process fitting into a deep notch in the tympanic crest of the quadrate (Fig. 2B). Supratemporal. The supratemporal is a small element, with a wide ventral base and a slightly sharper dorsal tip, inserted between the squamosal and the postparietal process of the parietal (Fig. 1B). Ventrally, it contacts the cephalic con- CRANIAL OSTEOLOGY OF FOSSORIAL GYMNOPHTHALMIDS dyle of the quadrate and the paroccipital process of the otooccipital. Vomer. The vomer (Fig. 3C) is the most anterior component of the skull floor, the palate. It is an elongated and broad bone situated posterior to the premaxillary palatal shelf and attached to a cavity in it (dorsal to the tooth row) through ligaments. The anterior process of the vomer is sharp but expands gradually posteriorly, forming a wider plate that loosely contacts the medial margin of the maxillary palatal shelf. At the tip of the maxillary teeth row, this plate narrows again to end in a sharp posterior medial process, overlain medial and dorsally by the anterior vomerine process of the palatine. The anterior dorsal surface of the vomer has a crest that, together with the septomaxilla, encloses the Jacobson’s organ. Septomaxilla. The septomaxilla is a domeshaped bone, located between the vomer and the nasal process of the premaxilla (Fig. 2A). It encloses the Jacobson’s organ, protecting it laterally and dorsally, and the vomer forms the floor to this capsule. The septomaxilla has a lateral maxillary process that contacts the maxillary palatal shelf dorsally; its dorsal surface has an extensive contact with the nasal process of the premaxilla. Palatine. The palatine forms the middle part of the skull floor (Fig. 3C). It has an approximately triangular shape, being wider anteriorly. It extends from the point of the posterior tip of the vomer, where it contacts the contralateral palatine in the midline, and narrows and diverges posteriorly to meet the pterygoid. Anteromedially, its vomerine process overlaps the posterior end of the vomer; anterolaterally, the maxillary process overlaps the posterior end of the maxillary palatal shelf. Between the medial and lateral extremities of the anterior margin of the palatine, a broad concavity, deeper anteriorly and gradually shallower posteriorly, forms the choanal channel. Posteriorly, the palatine narrows to a pterygoid process that overlaps the medial palatine process of the pterygoid. The lateral margin of the palatine forms the medial margin of the suborbital fenestra. In C. nicterus, the palatine develops a wide extension in the middle portion of the bone that connects to a corresponding expansion of the ectopterygoid and to an anterior extension of the pterygoid to completely close the suborbital fenestra (Figs. 1C and 4A). Pterygoid. The pterygoid forms the posterior portion of the skull floor (Fig. 3C). It is Y-shaped, with the anterior portion divided into a medial palatine process and a lateral transverse process (pterygoid flange), forming the posterior margin of the suborbital fenestra, and a long posterior quadrate process that meets the quadrate. Anteriorly, its palatine process overlaps with the pterygoidal process of the palatine bone, and its transverse process fits into a relatively deep facet between the dorsal and 1359 ventral portions of the posteromedial process of the ectopterygoid. Posteriorly, the quadrate process extends to meet the medial surface of the mandibular condyle of the quadrate. The basipterygoid process contacts the pterygoid at midlength. In C. nicterus, the anterior portion of the pterygoid is widened to form a single plate (rather than two separate processes) that participates in the closure of the suborbital fenestra (Figs. 1C and 4A). Ectopterygoid. The ectopterygoid is a small slender bone that forms the external margin of the suborbital fenestra (Figs. 1C, 2C and 3C). Anteriorly, it contacts the orbital process of the maxilla, laterally the middle portion of the jugal, and posteriorly the transverse process of the pterygoid, articulating with the latter through its posteromedial process. In C. nicterus, the ectopterygoid is wider than those of N. ablephara and S. catimbau, having a medial expansion that partially overlaps with the expansion of the palatine, thereby closing the suborbital fenestra (Figs. 1C and 4A). Quadrate. The robust quadrate is widely concave medially and can be divided into a mandibular condyle, articulating with the articular in the mandible, a lateral concave conch with a slight tympanic crest, and a cephalic condyle articulating with the supratemporal and paroccipital process; the ventral tip of the squamosal fits into a deep notch in the dorsal surface of the conch (Figs. 2B and 3B). Epipterygoid. The epipterygoid is a rod-like element, located between the alar process of the prootic and the descending process of the parietal in a vertical position (Fig. 6C). Its ventral tip is inserted into the dorsal surface of the pterygoid, and its dorsal tip approaches but does not contact the descending process of the parietal. Braincase. The braincase of these three species is relatively prominent compared with the rest of the skull, and most of it is exposed behind the parietal. The semicircular canals are easily observed. In C. nicterus and S. catimbau, a wide occipital recess (Rieppel, 1985; Figs. 1B, 2B, 4C, and 5E) is present, whereas in N. ablephara, no such opening is present (Figs. 3B and 6D). In N. ablephara and S. catimbau, the bones of the braincase are fused without visible sutures, but in C. nicterus, clear sutures can be distinguish between each bone. Thus, the description of each element’s limits will be based on the pattern observed in C. nicterus. Supraoccipital. The roofs of the braincase and of the otic capsules are formed by the wide supraoccipital. In N. ablephara and C. nicterus, the short ascending process of the tectum synoticum, originating at the anterior margin of the supraoccipital, rests in a shallow pit on the posteroventral margin of the parietal bone, delimiting two slender posttemporal fenestrae (Figs. 1A and 3A). In none of the Journal of Morphology 1360 J.G. ROSCITO AND M.T. RODRIGUES Fig. 7. C. nicterus, mandible in labial (A) and lingual (B) views. Scale bar 5 1 mm. S catimbau specimens analyzed was a posttemporal fenestra present; the anterior margin of the supraoccipital is completely overlapped by the parietal in this species. The supraoccipital contacts the dorsal margin of the prootic anterolaterally and the otooccipital complex posterolaterally. The posterior margin forms the dorsal margin of the foramen magnum. The dorsal ends of the anterior and posterior semicircular canals are joined midlaterally. A transverse crest crosses the dorsal surface of the supraoccipital, extending downward to the paroccipital process. This crest is much more prominent in N. ablephara and S. catimbau, whereas in C. nicterus, it is very faint. Prootic. The prootic is the anterior component of the otic capsule. Its anteriorly directed alar process forms part of the lateral wall of the skull, posterior to the eye. In C. nicterus, it expands dorsoventrally to contact part of the descending process of the parietal dorsally and the epipterygoid ventrally (Fig. 4F). In N. ablephara and S. catimbau, no ventral extension of this process is found, and only the most dorsal part approaches the epipterygoid (Figs. 5D and 6D). The anterior ends of the anterior and horizontal semicircular canals are located in this area. The incisura prootica, below the alar process, is an anterior C-shaped exit for the trigeminal nerve (Fig. 6C). The small facial foramen is observed posteriorly and at the same level as the trigeminal notch; a crista prootica begins at the parabasisphenoid and extends up to this foramen (Fig. 6D). Posterior to the facial foramen is the broad fenestra ovalis (Figs. 4C and 6D), marking the posterior limit of the prootic. The fenestra ovalis is broader in C. nicterus and smaller in N. ablephara, with that of S. catimbau of intermediate size. Ventrally, the prootic contacts the parabasisphenoid anteromedially and the basiocciJournal of Morphology pital posteromedially; dorsally, it contacts the supraoccipital. The internal surface of the prootic can be divided into a dorsal vestibular cavity and a ventral lagenar cavity (Fig. 6C), both separated by a constriction delimited by the trigeminal notch anteriorly and the fenestra ovalis posteriorly. The ventral margin of the vestibular cavity is pierced by the anterior and posterior auditory foramina; the internal opening of the facial foramen is observed ventral to the anterior auditory foramen. Columella auris. The columella auris is inserted into the fenestra ovalis. It is composed of a broad foot plate, which filles the entire area of the fenestra, and a shaft projecting laterally. In N. ablephara and S. catimbau, the shaft is relatively long and slender (Fig. 6D), but in C. nicterus, it is short, broad, and stout, being nodular and slightly triangular in shape (Fig. 3C). The posterior end of the cephalic condyle of the quadrate ends proximal to the shaft of the columella; the paroccipital process is located dorsal to the fenestra ovalis. Otooccipital. The compound otooccipital forms the posterior part of the otic capsule. It is formed by the embryonic fusion of the opisthotic and the exoccipital; there is no visible trace of this suture in C. nicterus. It contacts the supraoccipital dorsally and the basioccipital ventrally. The horizontal semicircular canal is located above the fenestra ovalis, and the posterior semicircular canal is located just anterior to the exoccipital region. The anterior region of the otooccipital is formed by the opisthotic, which is wider dorsally and narrower ventrally. Its anterior margin contacts the prootic, forming the posterior margin of the fenestra ovalis, and its anterodorsal margin is thickened to form the paroccipital process. In N. ablephara and S. catimbau the process is well developed, with an anteriorly directed projection (Fig. 6D), whereas in C. nicterus, this process is short and does not have this projection (Fig. 4B). The posterior limit of the opisthotic is determined Fig. 8. S. catimbau, mandible in labial (A) and lingual (B) views. Scale bar 5 1 mm. CRANIAL OSTEOLOGY OF FOSSORIAL GYMNOPHTHALMIDS Fig. 9. N. ablephara, mandible. (A) and (B) Labial views. (C) and (D). Lingual views. Scale bar 5 1 mm. by the anterior borders of the occipital recess and of the vagus foramen. The posterior region of the otooccipital is formed by the triangular exoccipital, with a wide ventral base that contributes to the composition of the occipital condyle and a dorsal end that forms the lateral margin of the foramen magnum. It forms the posterior margins of the occipital recess and of the vagus foramen. N. ablephara does not have an open occipital recess; this region is covered by a thin layer of bone (Fig. 6D). However, in S. catimbau and C. nicterus, this opening is quite broad (Figs. 4B,C and 5E). The foramina for the hypoglossal nerve roots are located in the ventral margin of the exoccipital bone. In N. ablephara, three small foramina are visible (Fig. 6D), whereas in S. catimbau and C. nicterus, only two small foramina are present (Figs. 1B and 2B). Basioccipital. The basioccipital forms most of the braincase floor, from the parabasisphenoid to the occipital condyle. It is a concave plate, slightly pentagonal in shape, fusing anteriorly to the parabasisphenoid and contacting laterally the prootic (anterior) and the otooccipital (posterior) (Figs. 1C, 2C and 3C). It has a minor contribution to the ventral margin of the occipital recess. Parabasisphenoid. The compound parabasisphenoid is composed of the endochondral basisphenoid, which ossifies from the embryonic acrochordal cartilage, the basipterygoid processes, and the dermal parasphenoid, which forms in later stages of 1361 development around the basisphenoid. This parabasisphenoid is formed by an anteriorly directed rostral process, which is long and slender in N. ablephara (Fig. 3C) and short and stout in C. nicterus (Fig. 1C), with that of S. catimbau (Fig. 2C) of an intermediate stage. It contacts the prootic posterolaterally and is fused to the basioccipital posteriorly. The basipterygoid process projects anterolaterally to meet the pterygoid medially. Orbitosphenoid. The paired small orbitosphenoids are located anteriorly with respect to the braincase elements and internally with respect to the orbits, at an angle to the dorsoventral axis of the skull. It is a relatively thin bone with a triangular shape, connecting to the interorbital cartilages (Figs. 3B and 5F). Mandible. The mandible is similar in the three species (Figs. 7–9). Meckel’s cartilage runs through Meckel’s canal, from the most anterior tip of the dentary posterior to the adductor fossa. A relatively short retroarticular process is present, as well as a short coronoid process. The convex splenial shapes the inferior border of the mandible. Dentary. The anterior part of the mandible is formed by the dentary, the largest bone in the jaw. The posterior end of the bone, at the labial surface, contacts the splenial and angular ventrally and the surangular posteriorly and underlies the anterior directed labial process of the coronoid dorsally. In C. nicterus and S. catimbau, this process extends over the posterior margin of the dentary up to the level of the last (posterior) tooth (Figs. 7A and 8A), whereas in N. ablephara, it extends even further (Fig. 9A,B). At the lingual surface, the opening of Meckel’s canal is observed at the anterior end of the dentary, and Meckel’s cartilage extends up to this opening. The posterior margin of the bone is overlapped by the splenial ventrally and by the anteromedial process of the coronoid (Fig. 9C,D). The alveolar portion of the dentary of Fig. 10. S. catimbau, hyoid apparatus in ventral view; (A) Cleared and stained specimen, (B) drawing of A. In both figures, anterior is to the top of the image. Scale bar in A 5 0.5 mm. Journal of Morphology 1362 J.G. ROSCITO AND M.T. RODRIGUES N. ablephara and S. catimbau bears 15 unicuspid teeth (Figs. 8A and 9A), whereas in C. nicterus, only 12 teeth are present (Fig. 7B). Coronoid. The coronoid is triangular with a short dorsal coronoid process. The labial process, relatively thin in C. nicterus and S. catimbau but longer and stouter in N. ablephara, is directed anteriorly and overlaps the posterior dorsal end of the dentary. On the lingual surface of the mandible, the anteromedial process overlaps the dentary and contacts the splenial ventrally, and the prearticular and surangular posteriorly. In C. nicterus, this process is short and broad, with its ventral margin overlapped by the splenial (Fig. 7B). In N. ablephara and S. catimbau, this process is more elongate anteriorly, being wider in the former and slender in the later, but is again overlapped by the splenial extending up to the level of its alveolar foramen (Figs. 8B and 9C,D). The slender posteromedial process of the coronoid overlaps the prearticular, forming the anterior margin of the adductor fossa. Splenial. The splenial (Fig. 9C,D) is a robust bone that forms the midventral portion of the lingual surface of the mandible. It overlaps the dentary anteriorly and the anteromedial process of the coronoid dorsally, and it contacts the prearticular posterodorsally and the angular posteroventrally. The ventral margin of the bone is convex and shapes the inferior margin of the mandible. Two foramina are present: a large alveolar foramen at the anteromedial portion of the splenial and a smaller anterior mylohyoid foramen ventral to the former. The ventral margin of the splenial is found on the labial surface of the mandible, ventral to the dentary. Angular. The small angular is located on the ventral margin of the mandible, posterior to the splenial. It is more exposed on the labial surface, contacting the surangular posterodorsally and having its anterior end overlapped by the dentary (Fig. 8A). On the lingual surface, only a strip of bone is seen. This lies ventral to the posterior end of the splenial, is overlapped by it anteriorly, and contacts the prearticular posteriorly (Fig. 8B). The posterior mylohyoid foramen is visible ventrally. Surangular. The surangular is a large and stout bone that forms the posterior portion of the labial surface of the mandible, posterior to the dentary (Fig. 9). It contacts the coronoid anterodorsally, the dentary anteriorly, and the angular ventrally and forms the external margin of the adductor fossa. A large surangular foramen is observed next to the contact with the coronoid. Posterior to this foramen, another small foramen opens internally next to the anterior part of the prearticular. Posteriorly, the surangular is fused with the prearticular-articular complex. Prearticular-articular complex. This complex is formed by the fusion of a large prearticular, formJournal of Morphology ing most of the posterior end of the lingual surface of the mandible, with the small articular, which provides the articulation to the skull on its dorsal surface (Fig. 9C,D). The prearticular forms the floor and the internal margin of the adductor fossa, while the articular forms its posterior margin. The retroarticular process is formed by the fused surangular and prearticular-articular complex. Hyoid apparatus. The hyoid apparatus (Fig. 10) is a complex of skeletal elements located ventral to the mandible, in the throat region, and supports the muscles associated with this region. The hyoid apparatuses of the three species are similar to each other, thus the description given here applies to all three species. It is composed of a short triradiate basihyal, a long glossohyal (lingual process) extending up to the mandibular symphisis, and three pairs of visceral arches (Fig. 10B). The first visceral arch, the hyoid cornu, originates from the anterior border of the basihyal and is directed anteriorly. From its mid portion originates a posteriorly directed process, the epihyal. The second visceral arch corresponds to the first ceratobranchial, connected to each posterior end of the basihyal and extending posteriorly to connect to the first epibranchial. The third visceral arch corresponds to the rudimentary second ceratobranchial. It is thin and short, and is located between the first ceratobranchial and the basihyal. The second epibranchial (Fig. 10A) is a free sigmoidal element, orientated lateroposteriorly and located anterior to the first ceratobranchial and dorsal to the epihyal; its anterior end is curved to approach the ventral portion of the otooccipital. The first ceratobranchial is the only ossified element of the hyoid apparatus. All other elements consist of calcified cartilage in the adult, except for the free second epibranchial which remains cartilaginous. DISCUSSION The skeleton is intimately associated with life habits and reflects functional adaptations to the style of living. The evolutionary transition from quadrupedal to a snake-like body plan and fossorial and burrowing locomotion requires the evolution of a complex of skeletal adaptations (Hanken and Wake, 1993; Lee, 1998; Rieppel, 1984, 1996). The consequences for skull morphology in fossorial animals are intimately associated with miniaturization and with the development of a solid skeletal covering for the brain and sense organs, protecting them from the impacts of the subterranean limbless locomotion (Rieppel, 1984). Thus, the reduction of body diameter and loss of a neck region (with the uniformization of the body and head diameter), the relative increase in braincase size, the increase of skull stoutness with loss of skull fenestrae, closure of the lateral walls, and more complex connections between the bone elements, especially due to the CRANIAL OSTEOLOGY OF FOSSORIAL GYMNOPHTHALMIDS utilization of the skull as a burrowing tool (Camp, 1923; Gans, 1960, 1969; Lee, 1998; Rieppel, 1984, 1996; Wake, 1993) are frequent and convergent evolutionary events. Therefore, fossoriality requires akinetic and robust skulls that protect the brain and sensorial organs from the mechanical forces of burrowing (Rieppel, 1996). Several features of the skull morphologies of C. nicterus, N. ablephara, and S. catimbau are clearly related to the adaptations to the fossorial and burrowing life style but reflect morphological adaptations to different levels of fossoriality. The skulls are stout and rigid as a result of strong and elaborated articulations and overlap between elements. The degree of fossoriality can be inferred through an analysis of the connections between skull elements: stronger and more complex articulations render a strengthened skull that is capable of supporting a greater stress. In contrast, weaker articulations reflect weaker evolutionary pressures for protection of the brain. Also, some bones show increased growth to contact neighboring elements and close open spaces in the skull, offering further protection to the brain and sense organs from the physical impacts to which the head is subjected. The sharp and projecting snout is formed by the thick horizontal plate of the premaxilla, which forms a strong articulation with the maxilla (in C. nicterus and S. catimbau), and, together with the large nasal process, builds a tool for burrowing. However, in N. ablephara, the articulation between the premaxilla and maxilla is weaker, probably indicating a lesser degree of stress to which the skull is subjected. In the fossorial limbless gymnophthalmid Bachia bicolor (Tarazona et al., 2008), the articulation between premaxilla and maxilla is similar to that observed in N. ablephara, while in Vanzosaura rubricauda (Guerra and Montero, 2009), their close relative, there is no contact between these elements. The frontoparietal contact is tightened through the well-developed frontoparietal tabs and a complex interlocked suture, thus excluding any possibility of cranial mesokinesis (Lee, 1998; Metzger, 2002; Rieppel, 1984, 1996). Also, the loss of the posttemporal fenestrae in S. catimbau indicates an akinetic and rigid skull due to the overlap of the supraoccipital by the parietal (Bell et al., 2003; Evans, 2003; Metzger, 2002; Müller, 2002). The reduced orbit and nasal openings in this fossorial lineage result in wider bones composing the anterior dorsal roof and lateral wall of the snout. The most extreme case of this morphological arrangement is observed in C. nicterus, followed by S. catimbau and by N. ablephara. The well-developed cristae cranii of the frontal of C. nicterus encapsulate the nasal bulbs and establish strong contacts with neighboring bones such as the palatine and prefrontal. Also, the extreme growth of the ventral process of the parie- 1363 tal and of the alar process of the prootic results in a completely closed posterior lateral wall of the skull in C. nicterus. In addition, the growth of bony extensions of the ectopterygoid, pterygoid, and palatine, which contact and overlap each other, in association with the maxillary palatal shelf, completely closes the suborbital fenestra. Together with a near contact between the maxillary palatal shelf and the vomer, this forms a rigid palate, suggesting a greater degree of adaptation to fossoriality in this species. These peramorphic traits (Rieppel, 1996) offer a higher degree of protection for the brain and contrast to the condition observed in the other two species. These traits evolved convergently in other head-first burrowers (Lee, 1998; Rieppel, 1996; Savitzky, 1983; Tarazona et al., 2008), although the different degrees of morphological differentiation suggest different degrees of adaptation to fossoriality. The enlarged otico-occipital region in relation to the dermatocranium results in a shifting in the position of the supraoccipital to directly contact the posterior margin of the parietal. As a consequence, the posttemporal fenestra is closed or reduced in size. The closure of the posttemporal fenestrae is also observed in other serpentiform or miniaturized (Hanken and Wake, 1993) squamates, such as some species of Scincidae, Amphisbaenia, Dibamidae, Serpentes, and Pygopodidae (Lee, 1998; Montero and Gans, 1999; Rieppel, 1981, 1984, 1996; Tarazona et al., 2008). As revised by Rieppel (1984), this relative increase in braincase size observed in small skulls is a consequence of the development of functional semicircular canals, whose curvatures requires a certain minimum radius for proper efficiency. The triradiate jugal of C. nicterus, with a welldeveloped free posterior process, is unique among Gymnophthalmidae, in which the jugal is always semilunar in shape. A free posterior jugal process is also observed in some lacertids, scincids and agamids (Evans, 2008), but it never develops to such large size as found in Calyptommatus. In Sphenodon, the large posterior process of the jugal forms, together with the small quadratojugal, a complete lower temporal bar. The presence of a posterior process of the jugal is now considered a secondary, because in basal lepidosaurians, no such process was present and, hence, no lower temporal bar (Evans, 2003; Müller, 2003). The development of such process in C. nicterus may be a consequence of the peramorphic development of some of the skull elements, such as the ventral process of the parietal or the growth of the bony extensions of the palate elements. Further study is needed to investigate if the presence of this process has a functional significance in feeding forces. A commonly observed pattern in fossorial morphologies is the reduction/loss of elements and/or simplification of their morphology (Hanken and Journal of Morphology 1364 J.G. ROSCITO AND M.T. RODRIGUES Wake, 1993). The skull of miniaturized animals may lack bones that are present in related and nonminiaturized species or show poor development of other elements. This aspect is observed in the reduced number of teeth in the premaxilla, maxilla, and dentary, with the highest degree of reduction being observed in C. nicterus, a feature also seen in miniaturized salamanders (Hanken and Wake, 1993). The poor development of the shaft of the columella as well as that of the paroccipital process in C. nicterus in comparison with those of N. ablephara and S. catimbau and the absence of an ascending process of the tectum synoticum and of posttemporal fenestrae in S. catimbau are features associated with the enlargement of the otic capsule. The increase in size of otic capsule imposes physical limitations on the size and development of other structures, such as observed by Rieppel (1984) for Dibamus species, Lee (1998) for several species of dibamids, snakes, and amphisbaenians, Müller (2002) for Parvilacerta parva, and Tarazona et al. (2008) for B. bicolor and others. Fusion of elements is frequent as a consequence of structural simplification (Lee, 1998). The absence of a lacrimal may indicate either the loss of this element or its fusion with the prefrontal; the analysis of embryonic development of this region may elucidate the condition observed in adult forms. Also, the size of the lacrimal flange (larger in N. ablephara and S. catimbau and absent in C. nicterus) and the presence of a single element representing the fusion of the postorbital with the postfrontal in C. nicterus (while in N. ablephara and S. catimbau both bones are distinct) indicate a greater degree of structural reduction in specific regions of the skull of C. nicterus. The presence of a single element in the postorbital region is frequently observed in species from several taxa (Camp, 1923). Nevertheless, although fossoriality has evolved repeatedly in Gymnophthalmidae, the only other genus presenting this condition is Alopoglossus, as well as the species Dryadosaura nordestina, Anotosaura vanzolinia and Colobosauroides cearensis (Evans, 2008, Rodrigues et al., 2005, 2009). Evans (2008) also reported a postorbitofrontal in Heterodactylus following Presch (1980); this is in contrast to our observations in cleared and stained specimens of H. lundii and H. imbricatus, which have distinct postorbital and postfrontal s. The identification of the postorbitofrontal is quite variable, being represented by either the postorbital or postfrontal alone or by a fusion of both (Camp, 1923). In C. nicterus, the postorbitofrontal is represented by the fusion of the well-developed postorbital with a reduced postfrontal; both elements are clearly identified in late embryonic stages (Roscito and Rodrigues, unpublished data). C. nicterus is also peculiar in having welldefined sutures between the braincase elements as Journal of Morphology an adult, whereas in its closest relatives N. ablephara and S. catimbau, no sutures are observed. The former pattern agrees with that observed in the gymnophthalmid B. bicolor (Tarazona et al., 2008), but the absence of sutures in adults seems to be more common, e.g., in the gymnophthalmids Euspondylus acutirostris (Montero et al., 2002), Potamites ecpleopus (Bell et al., 2003), Dryadosaura nordestina (Rodrigues et al., 2005), Caparaonia itaiquara, Brazil (Rodrigues et al., 2009), Alexandresaurus camacan (Rodrigues et al., 2007), and V. rubricauda (Guerra and Montero, 2009). The presence of a large occipital recess in C. nicterus and S. catimbau contrasts with the completely closed occipital recess in N. ablephara. Although it might be consequence of the small skull adapted to burrowing habits, the closure of the occipital recess is found in many different species and is not always correlated with fossoriality (Rieppel, 1985). However, an open occipital recess is also characteristic of fossorial pygopodids, such as Aprasia and Pletholax (Rieppel, 1985) and of fossorial scincids (Haas, 1936; Greer, 2002; Rieppel, 1985). According to Rieppel (1985), lizards that show a closed occipital recess lack an external tympanic membrane and have a large stapedial footplate, all related structural modifications for the transmission of ground pressure waves rather than airbone vibrations. This is not the case in N. ablephara, which shows a smaller stapedial footplate compared with C. nicterus and S. catimbau. Therefore, the skull of fossorial and burrowing lizards is a complex set of structural modifications related to cranial consolidation and, also of autapomorphic features. The evolution of head-first burrowing and the structural modifications associated with it are convergent features, which, in the Gymnophthalmidae, evolved several times independently (Pellegrino et al., 2001). A complete study of the development of such features, comparing the patterns observed in distantly related species, will probably provide an important evolutionary scenario for their history and may also shed light on the morphological transformations associated with the origin of snakes, as discussed by Rieppel (1984, 1996). ACKNOWLEDGMENTS The authors thank Hussam Zaher and Carolina Castro-Mello, from Museu de Zoologia da Universidade de São Paulo (MZUSP), for access to the stained specimens. LITERATURE CITED Bell CJ, Evans SE, Maisano JA. 2003. The skull of the gymnophthalmid lizard Neusticurus ecpleopus (Reptilia: Squamata). Zool J Linn Soc 139:283–304. Brandley MC, Huelsenbeck JP, Wiens JJ. 2008. Rates and patterns in the evolution of snake-like body form in squamate reptiles: Evidence for repeated re-evolution of lost digits and CRANIAL OSTEOLOGY OF FOSSORIAL GYMNOPHTHALMIDS long-term persistence of intermediate body forms. Evolution 62:2042–2064. Camp CL. 1923. Classification of the lizards. Bull Am Mus Nat Hist 48:289–481. Estes R, de Queiroz K, Gauthier J. 1988. Phylogenetic relationships within Squamata. In: Estes R, Pregill G, editors. Phylogenetic Relationships of the Lizard Families. Stanford: Stanford University Press. pp 119–281. Evans S. 2003. At the feet of the dinosaurs: The early history and radiation of lizards. Bio Rev 78:513–551. Evans S. 2008. The skull of lizards and tuatara. In: Gans C, Gaunt AS, Adler K, editors. Biology of the Reptilia, Vol. 20. Morphology H: The Skull of Lepidosauria. New York: Society for the Study of Amphibians and Reptiles. pp 1–344. Gans C. 1960. Studies on amphisbaenids (Amphisbaenia, Reptilia). 1. A taxonomic revision of the Trogonophinae, and a functional interpretation of the amphisbaenid adaptive pattern. Bull Am Mus Nat Hist 119:129–204. Gans C. 1969. Amphisbaenians—Reptiles specialized for a burrowing existence. Endeavour 28:146–151. Gans C. 1975. Tetrapod limblessness: Evolution and functional corollaries. Am Zool 15:455–467. Greer AE. 1991. Limb reduction in squamates: Identification of the lineages and discussion of the trends. J Herpetol 25:166–173. Greer AE. 2002. The loss of the external ear opening in scincid lizards. J Herpetol 36:544–555. Guerra C, Montero R. 2009. The skull of Vanzosaura rubricauda (Squamata: Gymnophthalmidae). Acta Zool (Stockholm) 90: 359–371. Haas G. 1936. Üeber das Kopfskelett von Chalcides guentheri (Seps monodactylus). Acta Zool 17:55–74. Hanken J, Wake DB. 1993. Miniaturization of body size: Organismal consequences and evolutionary significance. Ann Rev Ecol Syst 24:501–519. Lee MS. 1998. Convergent evolution and character correlation in burrowing reptiles: Towards a resolution of squamate relationships. Biol J Linn Soc 65:369–453. Metzger, KA. 2002. Cranial kinesis in lepidosaurs: Skulls in motion. In: Aerts P, D’Août K, Herrel A, Van Damme R, editors. Topics in Functional and Ecological Vertebrate Morphology. Maastricht, The Netherlands: Shaker Publishing. pp 15–46. Montero R, Gans C. 1999. The head skeleton of Amphisbaena alba Linnaeus. Ann Carnegie Mus 68:15–80. Montero R, Moro SA, Abdala V. 2002. Cranial anatomy of Euspondylus acutirostris (Squamata: Gymnophthalmidae) and its placement in a modern phylogenetic hypothesis. Russ J Herpetol 9:215–228. Müller J. 2002. Skull osteology of Parvilacerta parva, a smallsized lacertid lizard from Asia Minor. J Morphol 253:43–50. Müller J. 2003. Early loss and multiple return of the lower temporal arcade in diapsid reptiles. Naturwissenschaften 90:473–476. Pellegrino KC, Rodrigues MT, Yonenaga-Yassuda Y, Sites JW. 2001. A molecular perspective on the evolution of microteiid lizards (Squamata, Gymnophthalmidae), and a new classification for the family. Biol J Linn Soc 74:315–338. Potthoff T. 1984. Clearing and staining techniques. In: Moser HG, Richards WJ, Cohen DM, Fahay MP, Kendall AW, Richardson SL, editors. Ontogeny and Systematics of Fishes. New York: American Society of Ichthyologists and Herpetologists Spec. Publ. 1. Lawrence: Allen Press, pp 35–37. Presch W. 1980. Evolutionary history of the South American microteiid lizards (Teiidae: Gymnophthalminae). Copeia 1980: 36–56. Rieppel O. 1981. The skull and the jaw adductor musculature in some burrowing scincomorph lizards of the genera Acontias, Typhlosaurus and Feylinia. J Zool 195:493–528. Rieppel O. 1984. Miniaturization of the lizard skull: Its functional and evolutionary implications. In: Ferguson MW, editor. The Structure, Development, and Evolution of Reptiles. Symposia of the Zoological Society of London 52. New York: Oxford University Press. pp 503–520. Rieppel O. 1985. The recessus scalae tympani and its bearing on the classification of reptiles. J Herpetol 19:373–384. 1365 Rieppel O. 1996. Miniaturization in tetrapods: Consequences for skull morphology. In: Miller PJ, editor. Miniature Vertebrates: The Implications of Small Body Size. Symposia of the Zoological Society of London 69. New York: Oxford University Press. pp 47–61. Rodrigues MT. 1984. Nothobachia ablephara: Novo gênero e espécie do nordeste do Brasil (Sauria, Teiidae). Pap Av Zool 35:361–366. Rodrigues MT. 1991. Herpetofauna das dunas interiores do Rio São Francisco, Bahia, Brasil. I. Introdução à área e descrição de um novo gênero de microteiideo (Calyptommatus) com notas sobre sua ecologia, distribuição e especiação (Sauria, Teiidae). Pap Av Zool 37:285–320. Rodrigues MT. 1995. Filogenia e história geográfica de uma radiação de lagartos microteı́deos (Sauria, Teiioidea, Gymnophthalmidae). Unpublished PhD Thesis. Brazil: Universidade de São Paulo. Rodrigues MT, Freire EM, Pellegrino KC, Sites JW. 2005. Phylogenetic relationships of a new genus and species of microteiid lizard from the Atlantic forest of north-eastern Brazil (Squamata, Gymnophthalmidae). Zool J Linn Soc 144:543–557. Rodrigues MT, Pellegrino KC, Dixo M, Verdade VK, Pavan D, Argolo AJ, Sites JW. 2007. A new genus of microteiid lizard from the Atlantic forests of State of Bahia, Brazil, with a new generic name for Colobosaura mentalis, and a discussion of relationships among the Heterodactylini (Squamata, Gymnophthalmidae). Am Mus Novit 3565:1–27. Rodrigues MT, dos Santos EM. 2008. A new genus and species of eyelid-less and limb reduced gymnophthalmid lizard from northeastern Brazil (Squamata, Gymnophthalmidae). Zootaxa 1873:50–60. Rodrigues MT, Cassimiro J, Pavan D, Curcio FF, Verdade VK, Pellegrino KC. 2009. A new genus of microteiid lizard from the Caparó Mountains, southeastern Brazil, with a discussion of relationships among Gymnophthalmidae (Squamata). Am Mus Novit 3673:27. Savitzky AH. 1983. Coadapted character complexes among snakes: Fossoriality, piscivory, and durophagy. Am Zool 23:397–409. Shapiro MD. 2002. Developmental morphology of limb reduction in Hemiergis (Squamata: Scincidae): Chondrogenesis, osteogenesis, and heterochrony. J Morphol 254:211–231. Skinner A, Lee MS, Hutchinson MN. 2008. Rapid and repeated limb loss in a clade of scincid lizards. BMC Evol Biol 8:310. Song J, Parenti LR. 1995. Clearing and staining whole fish specimens for simultaneous demonstration of bone, cartilage and nerves. Copeia 1995:114–118. Springer VG, Johnson GD. 2000. Use and advantages of ethanol solution of alizarin red S dye for staining bone in fishes. Copeia 2000:300–301. Tarazona OA, Fabrezi M, Ramı́rez-Pinilla MP. 2008. Cranial morphology of Bachia bicolor (Squamata: Gymnophthalmidae) and its postnatal development. Zool J Linn Soc 152:775–792. Taylor WR, Van Dyke GC. 1985. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium 9:107–119. Torres-Carvajal O. 2003. Cranial osteology of the Andean lizard Stenocercus guentheri (Squamata: Tropiduridae) and its postembryonic development. J Morphol 255:94–113. Vitt LJ, Pianka ER, Cooper WE, Schwenk K. 2003. History and the global ecology of squamate reptiles. Am Nat 162:44–60. Wake MH. 1993. The skull as a locomotor organ. In: Hanken J, Hall BK, editors. The Skull. Functional and Evolutionary Mechanisms, Vol. 3. Chicago: University of Chicago Press. pp 197–240. Wiens JJ, Slingluff JL. 2001. How lizards turn into snakes: A phylogenetic analysis of body-form evolution in anguid lizards. Evolution 55:2303–2318. Wiens JJ, Brandley MC, Reeder TW. 2006. Why does a trait evolve multiple times within a clade? Repeated evolution of snake-like body form in squamate reptiles. Evolution 60:123– 141. Journal of Morphology