Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
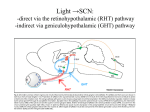
Role of circadian clocks intrinsic to SCN and GnRH neurons in regulating luteinizing hormone surge and other reproductive functions by Xue Wei Lin Integrated Program in Neurosciences, McGill University, Montreal August, 2015 A thesis submitted to McGill University in partial fulfillment of the requirements of degree of Master of Science © Xue Wei Lin, 2015 1 Table of Contents Abstract ....................................................................................................................................... 3 L’abstrait ..................................................................................................................................... 5 Co-‐Authorship Statement ...................................................................................................... 7 Acknowledgements ................................................................................................................. 8 Background ................................................................................................................................ 9 Hypothalamic-pituitary-gonadal (HPG) axis ............................................................................................... 11 Potential role of SCN neurons in HPG axis control .................................................................................... 12 Role of gonadotrope clocks in reproduction ................................................................................................. 14 Potential role of GnRH neuronal clocks in HPG axis function .................................................................. 15 Hypothesis ............................................................................................................................... 17 Methodology ........................................................................................................................... 18 Animal models .................................................................................................................................................. 18 Validation of BMAL1 ablation ........................................................................................................................ 19 Immunohistochemistry and histology ............................................................................................................ 20 Assessment of reproductive phenotypes ........................................................................................................ 21 Results ...................................................................................................................................... 23 GnBmalKO – Validation of BMAL1 Ablation .............................................................................................. 23 GnBmal1KO - Puberty Onset and Body Weight .......................................................................................... 25 GnBmal1KO - Estrus Cyclicity ....................................................................................................................... 27 GnBmal1KO – Gonadotropin assessments ................................................................................................... 28 SCN-Bmal1KO – Validation of BMAL1 Ablation ........................................................................................ 30 SCN-Bmal1KO – Estrus Cyclicity .................................................................................................................. 32 SCN-Bmal1KO – Gonadotropin assessments ............................................................................................... 34 Discussion ................................................................................................................................ 39 Conclusion and Future Directions ................................................................................... 45 References ............................................................................................................................... 47 2 Abstract The circadian clock represents a self-sustaining, entrainable oscillator driving rhythms of ~24 hours. The mammalian circadian system is organized in a hierarchical manner in which the suprachiasmatic nucleus (SCN) acts as the master pacemaker, coordinating the oscillations of peripheral clocks. Desynchrony of these oscillators due to shift work and intercontinental travel may contribute to reproductive dysfunction, warranting the study of the role that circadian clocks play in reproduction. Ovariectomized (OVX) rodents supplemented with surge-permissive estradiol exhibit a daily luteinizing hormone (LH) surge even in constant darkness, arguing that the surge is circadian regulated. While our lab has previously shown that mice deficient of clock function in all tissues (Bmal1-/-) completely lacked a LH surge, the contribution of individual clocks in specific hypothalamic-pituitary-gonadal (HPG) axis elements to the surge is still largely unclear. Our lab has already studied mice carrying a gonadotrope-specific disruption of the essential clock component Bmal1 (GBmal1KO) by means of Cre-LoxP technology and found evidence that the clock intrinsic to gonadotropes has no major role in the generation of the preovulatory LH surge. This suggests that circadian clocks elsewhere must be required for surge production. Therefore, my aim was to explore the specific contributions of the intrinsic clocks in the GnRH-expressing neurons in the preoptic area (POA) of the hypothalamus and of the SCN to the regulation of the LH surge. To this end, we created mice that lack clock function selectively in either GnRH-expressing neurons or in the SCN by disrupting Bmal1 employing, again, CreLoxP technology. We were able to achieve Bmal1 ablation efficacies of approximately 76% and 94% in the hypothalamic GnRH neurons of our GnBmal1KO line and the SCN cells of our SCNBmal1KO line, respectively. Examinations of puberty onset, body weight, estrus cyclicity as well as serum gonadotropin levels in GnBmal1KO females, however, indicate that these animals were 3 reproductively normal under laboratory conditions. SCN-Bmal1KO females, on the contrary, displayed prolonged estrus cycles likely due to extended time spent in estrus. Moreover, these females also displayed blunted LH surge levels as well as a dysregulation of LH surge timing. FSH levels were highly dysregulated as well. As Bmal1-/- females completely lacked a LH surge, this suggests that either the residual BMAL1+ cells in the SCN of the SCN-Bmal1KOs were sufficient to produce the observed LH surge or that surge production depends on clocks at different levels of the HPG axis, possibly by a concerted action. Studying clocks in kisspeptin neurons, which are thought to serve a central role in reproduction, as well as clocks in the ovarian cells may, therefore, represent the crucial next step in the quest for the mechanism underlying the timed production of the LH surge. 4 L’abstrait L'horloge circadienne représente une auto-entretenue, oscillateur entraînable rythmes de ~ 24 heures de conduite. Le système circadien chez les mammifères est organisé de manière hiérarchique dans lequel le noyau suprachiasmatique (NSC) agit comme le maître stimulateur cardiaque, la coordination des oscillations d'horloges périphériques. Désynchronisation de ces oscillateurs dues au travail posté et voyage intercontinental peuvent contribuer à un dysfonctionnement de la reproduction, ce qui justifie l'étude du rôle que jouent horloges circadiennes dans la reproduction. Ovariectomie (OVX) complétées par les rongeurs flambée permissive estradiol présentent une hormone lutéinisante quotidienne (LH) même dans l'obscurité constante, arguant que la flambée est circadien réglementé. Alors que notre laboratoire a déjà montré que les souris déficientes de la fonction d'horloge dans tous les tissus (Bmal1 -/-) complètement manqué d'un pic de LH, la contribution des horloges individuelles dans hypothalamo-hypophyso-gonadique (HPG) éléments des axes spécifiques à la hausse est encore largement pas clair. Notre laboratoire a déjà étudié des souris porteuses d'une perturbation spécifique gonadotrope de la composante d'horloge essentielle Bmal1 (GBmal1KO) au moyen de la technologie Cre-LoxP et trouvé des preuves que l'horloge intrinsèque à gonadotropes n'a pas de rôle majeur dans la génération de la montée de LH préovulatoire. Ceci suggère que les horloges circadiennes doivent être ailleurs nécessaires pour la production de surtension. Par conséquent, mon objectif était d'explorer les contributions spécifiques des horloges intrinsèques dans les neurones à GnRH exprimant dans la région préoptique (POA) de l'hypothalamus et de la SCN pour la régulation de la montée de LH. À cette fin, nous avons créé des souris qui manquent de la fonction d'horloge de manière sélective soit dans des neurones exprimant la GnRH ou dans le SCN par perturber Bmal1 employant la technologie de nouveau, Cre-LoxP. Nous avons réussi 5 à obtenir Bmal1 ablation efficacités d'environ 76,10% et 93,7% dans les neurones à GnRH hypothalamiques de notre ligne GnBmal1KO et les cellules de notre ligne SCN SCN-Bmal1KO, respectivement. Examens apparition de la puberté, le poids corporel, cycle oestral ainsi que les niveaux de gonadotrophine sérique chez les femelles GnBmal1KO, cependant, indiquent que ces animaux étaient la reproduction normale dans des conditions de laboratoire. Femelles SCNBmal1KO, au contraire, affichent des cycles d'oestrus prolongés probables en raison de temps prolongé passé en oestrus. En outre, ces femmes affichent également les niveaux de surtension LH émoussées, ainsi qu'une dysrégulation de LH timing de surtension. Les taux de FSH ont été très dérégulée ainsi. Comme Bmal1 - / - femelles complètement manquait un pic de LH, ce qui suggère que soit les BMAL1 + cellules résiduelles dans le SCN du SCN-Bmal1KOs étaient suffisantes pour produire le pic de LH observé ou que la production de surtension dépend horloges à différents niveaux de l'axe HPG , éventuellement par une action concertée. Horloges étudient dans les neurones kisspeptin, qui sont censés jouer un rôle central dans la reproduction, ainsi que des horloges dans les cellules de l'ovaire peuvent, par conséquent, représentent la prochaine étape cruciale dans la quête pour le mécanisme sous-jacent de la production chronométrée de la montée de LH. 6 Co-Authorship Statement The GnBmal1KO mouse line was previously generated by Dr. Florian Storch and our former PhD student, Adrienne Chu, at the Douglas Mental Health University Institute. The SCNBmal1KO mouse line was generated by Dr. Henrik Oster at the University of Luebeck, Germany. Our GnRH antibody was generated by and provided to us by Dr. Henryk Urbanski of the Oregon Health & Science University. Lastly, mice were bred at the Douglas Hospital Animal Facility with the assistance of Aude Villemain and the Animal Facility staff. 7 Acknowledgements I sincerely thank Dr. Florian Storch for the opportunity to conduct these experiments under his direction in his laboratory. More importantly, I thank Dr. Storch for his mentorship, guidance, trust, support, understanding, encouragement, patience, and especially for believing in my abilities. Words cannot sufficiently express my gratitude. Thank you to members of my Master’s Advisory Committee, Dr. Nicolas Cermakian and Dr. Raj Duggavathi, for their input, suggestions, guidance, and support throughout my Master’s education. I also thank my Advisory Committee Chair, Dr. Joseph Rochford, for his guidance, encouragement, and for always promptly answering my inquiries. A sincere thank you is also in order for my lab mates, Lei Zhu, for taking a lot of time to train and help me in the lab; Ian Blum, for his wise advice and words of encouragement and help in the lab; and Adrienne Chu, for setting up these excellent experiments and for her patience in guiding me through them. Thank you, also, to our research technician, Aude Villemain, for her help in maintaining our mouse colonies and genotyping. I next wish to thank Dr. Maia Kokoeva for her generosity in letting me use her lab equipment as well as members of her lab, notably Tina Djogo and Sarah Robins, for training and helping me with immunohistochemistry and imaging. I also wish to thank my best friend, Paulina Achita, for understanding me so well and always being there for me. Thank you also to my friends, Sydney McQueen, Natalie Chow and Minghao Ji for their encouragement and support. Lastly, I would like to thank my mother for being my rock and loving me regardless of success or failure. Also thank you to my father for his love and support. I could not have done this without both of you. 8 Background The mammalian circadian timer represents a self-sustaining, entrainable oscillator driving rhythms of approximately 24 hours in behavior and physiology (1). Although clocks have been found in almost all cells and tissues in mammals, the suprachiasmatic nucleus (SCN), a paired structure located in the antero-ventral hypothalamus, is considered to be the central site for circadian rhythm generation (1) as electrolytic lesioning of the SCN permanently eliminated circadian rhythms in drinking and locomotor activity in rodents (2). However, the capacity of generating self-sustained circadian rhythms of gene expression has also been found in peripheral organs. Moreover, the ability to entrain peripheral clocks in tissues such as the liver or lung independently of the SCN with feeding cycles (3) prompted a quest for the physiological relevance of peripheral clocks and investigations in the architecture of the mammalian circadian oscillator hierarchy. Today we know that within the mammalian circadian timing system, SCN acts as the chief pacemaker that sets the period and phase coherence between and within the oscillators present elsewhere in the brain and peripheral organs. The circadian clock consists of molecular building blocks that generate rhythms cellautonomously, which relies to a great extent on transcriptional-translational feedback loops. At its core, a transcription factor heterodimer of BMAL1 partnering with CLOCK or NPAS2 rhythmically drives transcription of the Period (Per) and Cryptochrome (Cry) genes via binding to E-box promoter elements. Upon translation, PER and CRY proteins first accumulate in the cytoplasm and then translocate back to the nucleus to repress their own transcription. As PER and CRY levels decrease, the cycle begins anew, thereby generating daily rhythms in protein abundance (4). In addition to this core feedback loop, further feedback loops including one involving the nuclear receptors Rev-erbα and β have been shown to provide a link between the 9 positive and negative limbs of the molecular oscillator. Rev-erbα competes as transcriptional repressor with the transcriptional activating RORs at the Bmal1 promoter while Rev-erbβ is activated by BMAL1 and CLOCK and repressed by PER1/2 and CRY 1/2 (5, 6). It has been suggested that desynchrony between the SCN and peripheral tissue oscillators due to shift work or transmeridian/intercontinental travel may contribute to reproductive dysfunction (7). Shift-working women report menstrual irregularities including shortened or lengthened menstrual cycle length, increased menstrual pain, and changes in the amount of menstrual bleeding (7-9). These female shift workers also display higher rates of miscarriages and are at greater risk of preterm parturition and subfertility (7, 10). In addition, genetic perturbations of clock function in rodent models result in various reproductive deficits. For example, female mice deficient in the Bmal1 gene (Bmal1-/-) exhibit prolonged estrus cycle length and are infertile due to embryo implantation failure (11, 12). Our lab also previously found that these Bmal1-/- females lack the proestrus luteinizing hormone (LH) surge but are still able to spontaneously ovulate at the expected time, challenging the widely held view that LH surge is indispensible for ovulation (13). Similarly, Clock mutant females display a highly blunted LH surge as well as have irregular and extended estrus cycles and a high rate of pregnancy failure (14). Lastly, Per1-/-;Per2-/-;Cry2-/- and Per1-/-;Cry1-/-;Cry2-/- triple mutant mice, which are arrhythmic immediately upon release into constant darkness (DD) or light (LL) conditions, show a reduction in litter size and elevated pup mortality rates (15). The presence of reproductive deficits in various clock mutant animals indicates that a functional circadian clock is critical for normal and successful reproduction. However, as these mouse models carry clock gene manipulations throughout all cells, the individual contributions of the central (SCN) and peripheral oscillators to the observed reproductive phenotypes are still largely unclear. 10 Hypothalamic-pituitary-gonadal (HPG) axis The hypothalamic-pituitary-gonadal (HPG) axis is vital in the development and regulation of the body’s reproductive system. Gonadotropin-releasing hormone (GnRH), considered to be the top regulator of the axis, is primarily produced in neurons that reside in the preoptic area (POA) of the hypothalamus (16). These neurons send axons into the median eminence where GnRH is secreted into the portal capillary system, and from there, it is transported to the pituitary. In the pituitary, GnRH binds to its cognate receptor, GnRHR, at the membrane surface of the gonadotropes, which are located in the anterior portion of the hypophysis. GnRH binding raises intracellular calcium levels, stimulating the release of LH and FSH from pituitary gonadotropes into the circulation (17, 18). Although LH surge occurs only once per menstrual or estrus cycle, it is invariably staged at a specific time of day implicating an involvement of the circadian timing system. The first line of evidence was provided by Everett and Sawyer using barbiturate, a central nervous system (CNS) depressant, in rats (19). Application of barbiturate in a 1.5-hr window on the afternoon of proestrus did not simply delay the surge until depressant action subsided but led to a shift of the LH surge by exactly one day (19). This suggested that ovulation might be triggered by a daily neuronal signal generated by the CNS. Another key evidence linking circadian rhythms and the reproductive axis was provided using ovariectomized (OVX) rats administered with surgepermissive estradiol (E2). In these animals, LH showed a daily LH surge at the end of the light phase over multiple consecutive days in a light-dark cycle (LD) but also under constant dark (DD) conditions, clearly demonstrating circadian control of the LH surge (20). Importantly, SCN lesions resulting in the abolishment of circadian locomotor activity also prevented LH surge generation in E2-primed animals (21, 22). In addition, transplantation of SCN grafts that were 11 kept within semipermeable polymeric capsules to prevent neural outgrowth into SCN-lesioned females did not rescue the LH surge, suggesting that appropriate synaptic connections are required by SCN neurons to conduct the timing signal for LH surge generation (23). The importance of the SCN within the reproductive axis was further corroborated with behaviorally “split” OVX + E2 hamsters housed under constant lights (LL) conditions. These hamsters experience two bouts of locomotor activity that are 12 hours apart as consequence of the antiphasic oscillation of the left and right halves of the bilateral SCN (24). Both activity bouts are preceded by a LH surge, further supporting the notion that surge timing is controlled by the SCN (25). Collectively, these experiments strongly argue that clocks within the master SCN pacemaker structure are essential for the precise timing of the LH surge but also surge generation per se. Potential role of SCN neurons in HPG axis control The SCN is composed of two main neuronal groups based on peptide expression: approximately 37% of SCN neurons express arginine vasopressin (AVP) in a shell-like pattern within the outer portions of the SCN and approximately ~25% of SCN neurons express vasoactive intestinal peptide (VIP) in the ventromedial core portion of the SCN (1). Both groups of SCN neurons have been found to make direct and/or indirect connections to GnRH neurons residing in the POA (1). There is no evidence that AVP neurons form direct connections to GnRH-expressing neurons in the POA, but AVP-containing SCN efferents contact neurons in the anteroventral periventricular nuclei (AVPV) (26, 27) that release kisspeptin (Kiss1) protein (27). These Kiss1expressing neurons, in turn, have been shown to project onto GnRH neurons and exhibit c-fos 12 expression, a marker of neuronal activation, coinciding with the timing of the LH surge (28, 29). Abundant evidence, therefore, indicates that AVP neurons form an indirect connection to GnRH neurons via Kiss1-releasing neurons in the AVPV. Interestingly, release and synthesis of AVP from the SCN peaks near the time of the preovulatory GnRH surge, which stimulates a similar surge in LH and subsequent ovulation (30, 31). Moreover, co-cultures of hypothalamic POA and SCN explants from female rats release GnRH in response to AVP administration but not to VIP administration (31), suggesting that involvement of SCN in the neuroendocrine regulation of reproduction may be attributable only to the AVP subpopulation. Further evidence for a role of SCN-derived AVP in LH surge production is demonstrated in OVX + E2 rats with SCN lesions in which LH surging is rescued by intracranially-administered AVP (32). This stimulatory effect, however, varies depending on the time of AVP injection; no effects was observed when AVP was administered during the first half of the light period while a LH surge was induced in 30% of the animals when AVP was administered during the second half of the light period (33). Even in Clock mutant female mice, which seem to lack a defined LH surge, a precisely timed intracerebroventricular (i.c.v.) injection of AVP can stimulate LH release with approximately 50% of the animals reaching surge levels (34). The differential effects of AVP when administered at different times suggest that local clocks along the HPG axis, perhaps at the level of GnRH neurons and/or gonadotropes, are also involved in the preparation of the LH surge. In contrast to AVP, VIP-containing SCN efferents were observed to form direct contacts to GnRH neurons using both tract-tracing and immunocytochemical staining (35). In addition, approximately ~40% of all of the GnRH neurons analyzed by Smith and colleagues were found to contain VIP2 receptors, one of the two G protein-coupled receptors to which VIP binds (36). However, the overall effects of VIP on LH surge and reproduction remain ambiguous. Using 13 OVX + E2 rodents, it was found that VIP treatment induced c-fos expression in VIP-innervated GnRH neurons (37) as well as increased their firing rate, but this response is gated by E2 and the time of day (38). On the contrary, Weick and Stobie found that intracranial injections of VIP in OVX + E2 rats did not simulate LH surge but rather attenuated surge release (39). More recently, however, Sun and colleagues found, using OVX + E2 rats of both young and middle ages, that GnRH neurons can exert differential effects on LH release and c-fos expression upon VIP treatment, which depends on the age of the animal (40). I.c.v. infusion of VIP in young females delayed and blunted the peak LH release and decreased c-fos expression while VIP infusion in middle-aged females advanced and increased the LH surge (40). This age-dependent effect of VIP on LH surge induction may serve as a potential explanation for the ambiguous data discussed previously. Interestingly, fertility of Vip2-/- null mutant male mice was only mildly reduced and neither pregnancy rates nor neonatal litter sizes were affected in Vip2-/- females (41). This undermines the role of VIP in the regulation of LH surge and reproduction and further raises the likelihood that either AVP-producing SCN cells or intrinsic oscillators within the elements of the HPG axes such as the GnRH neurons and/or gonadotropes are required for proper LH surge generation. Role of gonadotrope clocks in reproduction As elaborated above, while it is evident that the circadian timing system plays a role in the regulation of the LH surge, it is unclear if this is exerted solely by SCN neurons or if intrinsic oscillators along the HPG axis are involved as well. Our lab has already examined the potential contribution of pituitary gonadotropes to reproductive physiology, including LH surge generation and ovulation (13). We generated mice that lacked the essential clock component 14 BMAL selectively in gonadotropes (GBmal1KO) using a Cre-loxP approach involving a “floxed” Bmal1 allele in conjunction with a GnRHR-Cre recombinase line. By comparing vaginal cytology in GBmal1KO and WT females, we found that although the proportion of time spent in each estrus stage did not differ significantly between the two groups, there was a significant increase in cycle length variance in GBmal1KO females compared with their WT littermates. Measurement of serum gonadotropin levels by taking tail vein blood consecutively for 5 to 7 days at the expected time of the LH surge (Zeitgeber time, ZT 11) revealed that although LH surge activation was not impaired in GBmal1KO animals, serum LH levels were significantly higher at all cycle stages in GBmal1KO mice than that in the WT. Specifically, the average overall serum LH baseline concentration was calculated to be 207 ± 41.1 pg/mL in the GBmal1KO mice while it was only 58.3 ± 16.6 pg/mL in the WT. Since the average LH surge level at ZT 11 on proestrus was also higher in GBmal1KO mice, the resulting proestrus LH surge-to-baseline ratio showed similar fold change in the two genotypes. Estrus FSH levels were also significantly elevated in the GBmal1KO females as compared with the WT. Moreover, genotype did not affect puberty onset, time to impregnation, gestation time, or litter size. In addition, like the WT animals, GBmal1KO females were fertile and were able to produce viable offspring, further suggesting that GBmal1KO females were reproductively normal. Since Bmal1/- mice completely lacked a LH surge, this suggests that circadian clocks in the SCN or in other organs along the HPG axis are responsible for surge induction (13). Potential role of GnRH neuronal clocks in HPG axis function According to the work reviewed above, GnRH neurons residing in the POA of the hypothalamus may be a good next candidate to investigate their contribution to the regulation of 15 the LH surge and reproduction in general. Indeed, endogenous clocks in these neurons have been observed both in vivo (42) and in immortalized, GnRH-secreting GT1-7 mouse hypothalamic cells (43, 44). Analysis of immunostaining on coronal sections of GnRH-eGFP transgenic mouse brains at 4-hour intervals over a 24-hour period revealed clear circadian rhythmicity for core clock proteins BMAL1 and PER2 antiphasic to one another, as expected for bona fide clock cells (42). The mouse GnRH-secreting cell line, GT1-7, were found to express core circadian clock genes including Bmal1, Clock, Per1, Per2, Per3, Cry1, and Cry2 (43, 44). Furthermore, mRNA levels of Bmal1, Per1, and Per2 were shown to fluctuate in a circadian manner when monitored for 54 hours (43, 44). Interestingly, transient expression of the dominant negative Clock-Δ19 protein in these GT1-7 cells significantly altered the pulse patterns of GnRH secretion, suggesting that an SCN-independent clock that is intrinsic to GnRH neurons is required for normal GnRH pulsatile secretion (44). Moreover, as discussed above, AVP administration induces LH surges by acting on Kiss1 neurons of the AVPV, which in turn activate GnRH neurons via Kiss1 secretion. However, LH surges could only be initiated by AVP stimulation during a specific daily time window, indicating potential gating, possibly by intrinsic clocks, at the level of Kiss1 and/or GnRH neurons. By examining c-fos expression in Kiss1 and GnRH neurons in response to AVP injections in Syrian hamsters, Williams and colleagues determined that the AVP-mediated excitation was gated by GnRH but not Kiss1 neurons (28). Therefore, intrinsic clocks within GnRH neurons may be critical for the temporal control of the LH surge. Accordingly, AVP neurons of the SCN and GnRH neurons represent excellent candidates for clock cells that have a critical role in the timing of reproductive functions. 16 Hypothesis Based on the available information in the literature and previous research already conducted in our laboratory, we hypothesized that the SCN and/or intrinsic clocks in the GnRHreleasing neurons in the hypothalamic POA play a critical role in the generation of luteinizing hormone surge and subsequent ovulation. To test this hypothesis, we examined two lines of mice that lack clock function specifically in the SCN or in the GnRH neurons. Intrinsic rhythms in these neurons can be abolished via disruption of the essential core clock component Bmal1, and will be discussed in detail in the methods section below. The resulting tissue-specific Bmal1KO lines, SCN-Bmal1KO and GnBmal1KO, were first assessed for BMAL1 ablation efficacy and then tested for reproductive phenotypes. These reproductive testing included assessment of average body weight, weight and age at puberty onset, estrus cycle structure and length, as well as serum levels of the gonadotropins LH and FSH. Thus, the overall aim of my thesis was to assess the role(s) of clocks intrinsic to (1) the SCN structure and (2) GnRH neurons in reproductive physiology. We expect that these studies will provide deeper insight into the mechanisms of reproductive timing but may also help to elucidate why shift work- and transmeridian air-travel is linked to reproductive dysfunctions. 17 Methodology Animal models We used two lines of mice that lacked clock function selectively in the SCN neurons and the GnRH-expressing neurons in the POA. This was accomplished by employing the Cre-loxP technology, which allowed for tissue-specific disruption of the essential clock component Bmal1. Specifically, we mated mice expressing a floxed allele of Bmal1 (45), where exon 8 is flanked by two loxP sites, with mice carrying Cre recombinase driven by a cell-type specific promoter. This resulted in the excision of Exon 8 of the Bmal1 gene only in cells that expressed Cre. Deletion of this exon led to BMAL1 deficiency and disrupted the transcriptional-translational feedback loop, thereby eliminating the intrinsic rhythm generation capacity from the affected cell. To elucidate the role that clocks specifically in SCN neurons play in reproduction, we employed the SCN-Bmal1 knockout (SCN-Bmal1KO) line, in which the Bmal1 gene has been disrupted preferentially in SCN neurons. This line of mice was generated by crossing Synaptotagmin10- (Syt10-) Cre knock-in mice (46) with our floxed Bmal1 mice. Syt10 transcripts are highly enriched throughout the adult SCN and are present only in a few other CNS sites (46). Moreover, there are multiple Synaptotagmin paralogs expressed in the SCN, which allows for compensation for the loss of Syt10 due to insertion of the Cre recombinase gene (46). To test for the role of GnRH neuronal clocks, we crossed floxed Bmal1 mice with mice carrying an internal ribosomal entry site (IRES)-Cre knock-in the GPR54 gene (23), resulting in GnBmal1KO mice. GPR54 is a G-protein coupled receptor that binds Kiss1 and is expressed in nearly all GnRH neurons (47). Therefore, knocking in IRES-Cre downstream of the GPR54 stop codon at the GPR54 locus results in a GnRH neuron-specific Cre driver. 18 Note that the mice were housed under a 12:12 light-dark cycle (LD) in the Douglas Animal Facility. Experiments were carried out in accordance with the protocol approved by the McGill University Animal Care Committee. 2 to 4-month-old female mice were used for experiments with littermates serving as experimental controls, unless otherwise stated. Validation of BMAL1 ablation Bmal1 gene ablation must first be validated in both SCN-Bmal1KO and GnBmal1KO lines. In SCN-Bmal1KO mice, we performed immunohistochemical staining of BMAL1 proteins in brain sections containing SCN neurons. Note that the BMAL1 antibody has already been validated and has good efficacy. For the staining, we used a total of 6 SCN-Bmal1KO females along with 5 corresponding wild type littermates in order to provide a robust data set to assess recombination efficacy and the identity/location of any remaining Bmal1-expressing SCN neurons. Specifically for each animal, we examined a total of 5, 8-µm-thick coronal sections encompassing the entire rostral-caudal extend of the SCN. In GnBmal1KO mice, we performed double immunohistochemical labeling using antibodies against BMAL1 and GnRH proteins on brain sections containing the hypothalamic POA. The GnRH antibody used in this experiment has previously been generated and validated by Dr. Henryk Urbanski (Oregon Health & Science University). For the staining, we employed 5 wild type and 7 GnBmal1KO female mice. For each animal, we again analyzed 5 coronal sections encompassing the POA that were each 40 µm in thickness. This allowed us to robustly determine the degree of Bmal1 disruption within the GnRH neuronal population. 19 Immunohistochemistry and histology For the SCN-Bmal1KO line, mice were anesthetized with ketamine/xylazine and the tissues were removed and drop-fixed with 4% paraformaldehyde in 0.1M PBS for 12-18 h at 4°C. Fixed tissues were first dehydrated with increasing percentages of ethanol in water and then the ethanol was removed from the tissue using Clearene solution (Leica, Concord, Ontario, Canada). The tissue was subsequently embedded in paraffin at 60°C, cooled to room temperature, and cut coronally into 8-µm sections using a microtome. The sections were mounted on SuperFrost Plus glass slides and stored at room temperature until use. For the GnBmal1KO line, mice were anesthetized with ketamine/xylazine and perfused transcardially with 4% paraformaldehyde in 0.1M PBS. Tissues were removed and postfixed in 4% paraformaldehyde in 0.1M PBS for 24 h at 4°C, incubated overnight with 30% sucrose in 0.1M PBS at 4°C, embedded in optimal cutting temperature compound (OCT, Tissue-Tek; Cedarline, Burlington, Ontario, Canada), and frozen in the cryostat at -18°C. Frozen tissues were cut into 40-µm sections using a cryostat, mounted on SuperFrost Plus glass slides, and store at -80°C until use. For immunohistochemical staining of the SCN-Bmal1KO line, the paraffin-embedded tissues were first deparaffinized with Clearene solution (Leica, Concord, Ontario, Canada) and then rehydrated with decreasing percentages of ethanol in water. Tissues were incubated in blocking solution (5% donkey serum, 0.01% Triton X-100 in PBS) for 1 h at room temperature and then incubated with rabbit anti-BMAL1 antibody (1:1000; Novus Biologicals, Oakville, Ontario, Canada) at 4°C overnight. The secondary antibody used was donkey anti-rabbit Alexa 488 (1:500; Invitrogen, Burlington, Ontario, Canada). 20 For double-immunohistochemical staining of the GnBmal1KO line, frozen tissues were defrosted in PBS at room temperature, incubated in blocking solution (3% donkey serum, 3% goat serum, 0.01% Triton X-100 in PBS) for 1 h at room temperature, and incubated with mouse anti-GnRH (1:1000; Dr. Henryk Urbanski, Oregon Health & Science University) and rabbit antiBMAL1 antibodies (1:1000; Novus Biologicals, Oakville, Ontario, Canada) at 4°C overnight. The secondary antibodies used were donkey anti-rabbit Alexa 488 and goat anti-mouse Alexa 568 (1:500; Invitrogen, Burlington, Ontario, Canada). Cells were counted using the Cell Counter Plugin of the ImageJ software. For the SCNBmal1KO line, SCN regions were first encircled and then DAPI+ cells as well as BMAL+/DAPI+ cells were counted. The percentages of BMAL+ cells in the SCN were calculated using the following formula: (number of BMAL+/DAPI+ cells/number of DAPI+ cells)*100%. For the GnBmal1KO line, GnRH+/DAPI+ and BMAL1+/GnRH+/DAPI+ cells encompassing the entire POA of each brain section were counted. The percentages of BMAL+ cells in the GnRH+ cells in the POA were calculated using the following formula: (number of BMAL1+/GnRH+/DAPI+ cells/number of GnRH+/DAPI+ cells)*100%. Assessment of reproductive phenotypes We have assessed an array of reproductive parameters in a minimum of 10 GnBmal1KO and 10 control females including body weight, weight and age at puberty onset, and estrus cycle structure and length. In addition, we have also examined serum levels of gonadotropin release including luteinizing hormone and follicle-stimulating hormone. An N of ~10 enables us to uncover even relatively slight alterations in, for instance, estrus cyclicity or LH surge behavior. 21 Female puberty onset was assessed by daily vaginal membrane breech inspection, as it is considered to be the first sign of puberty initiation. Estrus cytology was assessed by vaginal flushing using 0.1M PBS consecutively for 7 days and subsequent microscopical inspection of the shed cells.. For serum gonadotropin measurements, blood from female mice was collected from the tail vein at ZT11, the expected time of the LH surge, for 7 consecutive days. Blood samples were centrifuged and the resulting serum aliquots were analyzed for LH and FSH content on a Luminex 200 platform (Luminex, Toronto, Ontario, Canada) using a Milliplex rat pituitary magnetic beads panel as described by the manufacturer (catalog item RPT86K/RPTMAG-86K; Millipore, Mississauga, Ontario, Canada) (13). 22 Results GnBmalKO – Validation of BMAL1 Ablation In order to validate Bmal1 gene ablation in the GnBmal1KO line, we have performed double immunohistochemical labeling using antibodies against BMAL1 and GnRH proteins on coronal brain sections containing the hypothalamic POA from both GnBmal1KO and control littermates (Fig. 1A). We found that 88±1.% of GnRH-positive cells in the control animals were BMAL1-positive while only 24±4.6% of GnRH-positive cells in GnBmal1KO females immunoreacted with the BMAL1 antibody (Fig. 1B). T-test analysis confirmed a significant difference between the percentages of GnRH cells that were BMAL1-positive in the two genotypes: t(10) = 11.27, p < 0.0001 (two-tailed). 23 A B Figure 1. BMAL1 ablation validation in GnBmal1KO females. A. Double immunolabeling for BMAL1 and GnRH of coronal sections containing the hypothalamic preoptic area from WT and GnBmal1KO females. Boxed areas are shown enlarged in the other images. B. Quantification of the fraction of GnRH-positive cells that immunoreacted with BMAL1antibodies; n = 5 for WT and n = 7 for GnBmal1KO. By student’s t test: *, p < 0.0001. Bars indicate mean ± SEM. 24 GnBmal1KO - Puberty Onset and Body Weight Body weight was recorded daily for GnBmal1KO and control littermates from 24 to 57 days of age. Repeated measures ANOVA revealed no significant age-adjusted differences in body weight (p = 1.000, Fig. 2C). GnBmal1KO and control females were also inspected daily for vaginal membrane breech in order to determine the age of puberty onset. GnBmal1KO animals had an average puberty onset (±SEM) of 31.5±0.5 days of age while their wild-type littermates had an average puberty onset of 31.5±0.5 days (Fig. 2A). An unpaired t-test on the data was conducted and it revealed that the average ages of puberty onset were not significantly different between GnBmal1KO and control females, t(25) = 0.049, p = 0.96 (two-tailed). Lastly, the weight at the time of puberty onset between GnBmal1KO (14.1±0.2 g) and control (13.8±0.3 g) animals were compared and an unpaired t-test revealed that the two means were not significantly different, t(25) = 0.86, p = 0.34(two-tailed), arguing that there was no genotype effect on gross reproductive development. 25 Figure 2. Puberty onset and body weight. A. The average age of puberty onset in WT-control (n = 12) and GnBmal1KO (n = 15) females (n.s., not significant, p = 0.96). B. The average weight at the time of puberty onset in WT-control (n = 12) and GnBmal1KO (n = 15) females (n.s., not significant, p = 0.40). C. Mean daily body weight of WT (n = 5) and GnBmal1KO (n = 9) females from postnatal days 24 to 57 (n.s., not significant, p = 1.000). Bars/time points indicate mean ± SEM. 26 GnBmal1KO - Estrus Cyclicity In order to determine whether Bmal1 disruption in GnRH neurons resulted in alteration of estrus cycle and structure, we assessed estrus cytology in GnBmal1KO and control females (Fig. 3). The proportion of time spent in diestrus I/diestrus II (DI/II), proestrus (PE), and estrus (E) did not differ significantly between the two genotypes [(F(1,23) = 1.4, p = 0.25, Fig. 3A]. Moreover, GnBmal1KO and WT-control animals spent averages of (±SEM) 1.8±0..2 and 1.5±0.1 consecutive days in estrus per cycle, respectively (Fig. 3B), which were not statistically different [unpaired t-test, two tailed, t(54) = 1.32, p = 0.19]. The average estrus cycle lengths also did not differ significantly between GnBmal1KO (5.4±0.4 days) and WT animals [6.2±0.6 days; unpaired t-test, two tailed, t(50) = 1.13, p = 0.26; Fig. 3C]. Figure 3. Estrus cyclicity. A. Proportion of time spent in diestrus I/diestrus II (DI/DII), proestrus (PE), and estrus (E) (t-test, p = 0.24). B. Average consecutive days spent in estrus each cycle for GnBmal1KO (n = 36) and control (n = 20) animals (t-test, p = 0.1910). C. Average cycle length in GnBmal1KO (n = 36) and control (n = 16) animals (t-test, p = 0.26). Bars indicate mean±SEM. 27 GnBmal1KO – Gonadotropin assessments In order to determine whether serum gonadotropin levels, i.e., LH and FSH levels, differ between GnBmal1KO and WT-control animals, blood was collected from the tail vein of each individual animal at ZT11, the time when LH normally surges on proestrus day, for 7 consecutive days. The serum from the samples was then analyzed for LH and FSH content on a Luminex 200 platform, as described above. ZT11 LH serum levels across all estrus cycle stages were similar between WT and knockout animals. As expected, serum LH surged on the day of proestrus and returned to baseline levels on the following estrus day in both groups of animals (Fig. 4A). LH surge concentrations did not differ between GnBmal1KO (24.8±3.5 ng/ml) and the WTs [27.4±8.1 ng/ml; T-test, two-tailed, t(24) = 0.31, p = 0.76; Fig. 4B]. Similarly, GnBmal1KO and WT-control females showed mean baseline levels of LH of 0.28±0.04 and 0.23±0.027 ng/ml (Fig. 4C) that were statistically indifferent [t-test, two-tailed, t(118) = 0.90, p = 0.37]. 28 Figure 4. GnBmal1KO LH profile. A. Serum LH levels in representative individual GnBmal1KO (top) and control (bottom) animals measured daily at ZT 11 over 7 consecutive days. Red traces indicate LH levels, whereas black traces show estrus stage. B. Proestrus LH levels at ZT11, the expected time of surge, for control (n = 12) and GnBmal1KO (n = 14) animals (t-test, p = 0.76). C. Baseline LH levels at ZT11 for control (n = 57) and GnBmal1KO (n = 63) animals (t-test, p = 0.37); data points represent LH levels on non-proestrus days. Bars indicate mean ± SEM. FSH serum levels across all estrus cycle stages were also similar between WT-control and knockout animals. As expected, FSH trough levels were found on diestrus I/diestrus II (DI/DII) days and maximum values were found on the day of estrus, which likely reflect the secondary FSH surge (Fig. 5A). Detailed analyses of murine serum FSH fluctuations conducted previously in our lab indicated that the primary FSH surge occurs after the onset of proestrus, as it is triggered by the proestrus GnRH rise, while the secondary FSH surge occurs around estrus day and relies on activin and inhibin signaling (13, 48). Direct comparisons of the average FSH 29 levels on DI/DII, proestrus (PE), and estrus (E) days did not reveal any significant differences between the genotypes [Fig. 5B; ANOVA, F(1,15) = 0.32, p = 0.58]. Figure 5. GnBmal1KO FSH profile. A. Serum FSH levels in individual, representative GnBmal1KO (top) and WT-control (bottom) animals measured daily at ZT 11 over 7 consecutive days. Red traces indicated FSH levels and black traces indicate estrus stage. B. Serum FSH levels at diestrus I/diestrus II (DI/II), proestrus (PE), and estrus (E) for control (n = 9) and GnBmal1KO (n = 6) animals (t-test, p = 0.58). Bars indicate mean ± SEM. SCN-Bmal1KO – Validation of BMAL1 Ablation In order to validate Bmal1 gene ablation in the SCN-Bmal1KO line, we have performed immunohistochemical staining against BMAL1 in brain sections encompassing the SCN from both SCN-Bmal1KO and their WT littermates (Fig. 6A). Our results indicate that approximately 58.6±3.0% of the cells the SCN contained BMAL1 in WT animals, while only 7.3±0.7% of SCN 30 cells contained BMAL1 in SCN-Bmal1KO animals (Fig. 6B), a difference that was highly significant [t-test, two-tailed, t(20) = 17.81, p < 0.0001]. Figure 6. BMAL1 expression in the SCN of SCN-Bmal1KO and WT-control animals. A. Immunohistochemical staining of brain sections encompassing the SCN in SCN-Bmal1KO and control female animals using BMAL1 antiserum. Boxed areas are shown enlarged in the third and forth rows. Dashed lines outline the SCN regions based on the high density of cell nuclei that is characteristic for the SCN. B. Quantification of fraction of SCN cells that immunoreacted with BMAL1 antibodies; n = 10 for WT and n = 12 for SCN-Bmal1KO animals. Student’s t test: *, p < 0.0001. Bars indicate mean ± SEM. 31 SCN-Bmal1KO – Estrus Cyclicity We also assessed estrus cytology in SCN-Bmal1KO and control females (Fig. 7). We found that SCN-Bmal1KO females spent significantly more time in estrus (2.5±0.2 days, n = 92) compared to their WT littermates [1.8±0.1 days, n = 114; t(204) = 4.47, p < 0.0001; Fig. 7A, 7E], very reminiscent of what was previously observed in Bmal1-/- females (13). Upon closer examination, more than 99% of the control animals spent either 1, 2 or 3 days in estrus while only approximately 61% of the knockout animals spent 3 days or less in estrus (Fig. 7D). SCNBmal1KO females also showed an increase in overall cycle length compared to the controls (Fig. 7C), which has also previously been reported for Bmal1-/- animals (12, 13). On average, we found that SCN-Bmal1KO females cycled approximately once every 6.4±0.2 days (n = 86) while WT animals cycled only once every 5.4±0.1 days (n= 107), a difference that was also highly significant [t test, two-tailed, t(191) = 3.779, p = 0.0002]. Examination of the cycle length distribution within the two groups revealed a peak at 5 days for the WTs, while the knockout animals showed a much wider spread with a shallower peak at around 6 days (Fig. 7B). 32 Figure 7. SCN-Bmal1KO estrus cycle characteristics. A. Estrus cycles in representative SCNBmal1KO and control female animals. B. Cycle length distribution within groups. C. Mean cycle lengths in SCN-Bmal1KO (n = 86) and control animals (n = 107). By Student’s t test: *, p = 0.0002. D. Distribution of consecutive days spent in estrus in SCN-Bmal1KO and control animals. By ANOVA: *, p < 0.001. E. Mean consecutive days spent in estrus in SCN-Bmal1KO and control animals. By Student’s t test: *, p < 0.0001; n = 114 for WT and n = 92 for SCNBmal1KO. Bars indicate mean ± SEM. 33 SCN-Bmal1KO – Gonadotropin assessments Serum gonadotropin levels were also assessed in SCN-Bmal1KO and the corresponding control animals. Again, blood was collected from the tail vein of each individual animal at ZT11 for 7 consecutive days and the corresponding serum samples were analyzed for LH and FSH content. Estrus cyclicity was assessed via vaginal lavage concurrently and plotted against serum LH (Fig. 8) and FSH levels (Fig. 9) for three representative SCN-Bmal1KO and three representation control animals, respectively. In general, serum LH levels in SCN-Bmal1KO females appear to be lower at all cycle stages than the LH levels in the controls (Figure 8A). Moreover, LH surge timing in SCN-Bmal1KO animals seems to be dysregulated, occuring more frequently and at inconsistent times (Fig. 8A). Although SCN-Bmal1KO females were able to mount a LH surge, a Mann-Whitney test revealed that the average LH surge level in SCNBmal1KO females was significantly lower than that in the controls (p < 0.02; Fig. 8B). 34 Figure 8. SCN-BmalKO LH profile. A. Serum LH levels in individual, representive SCNBmal1KO (top) and WT-control (bottom) animals measured daily at ZT 11 over 7 consecutive days. Blue traces indicated LH levels and black traces indicate estrus stage. B. Average serum LH amplitude at ZT11, the expected time of surge, for control (n = 4) and SCN-Bmal1KO (n = 5) animals. *, p < 0.02, Mann-Whitney test. Bars indicate mean ± SEM. FSH serum levels also seemed to be dysregulated in SCN-Bmal1KO animals. While FSH levels appeared to fall and rise with estrus stage in WT-controls, no general trend could be observed for the knockout animals (Fig. 9A). A repeated-measure two-way ANOVA conducted on the data revealed that the average serum FSH concentration at any given estrus cycle stage did not differ significantly between the two genotypes [p = 0.28; Fig. 9B]. 35 Figure 9. SCN-Bmal1KO FSH profile across the estrus cycle. A. Serum FSH levels in individual, representative SCN-Bmal1KO (top) and control (bottom) animals measured daily at ZT 11 over 7 consecutive days. Blue traces indicated FSH levels and black traces indicate estrus stage. B. Serum FSH levels at diestrus I/diestrus II (DI/II), proestrus (PE), and estrus (E) for control (n = 5) and GnBmal1KO (n = 5) animals. Non-significant, p = 0.28, ANOVA. Bars indicate mean ± SEM. In order to determine whether the proestrus LH surge was indeed blunted in SCNBmal1KO females or rather shifted to a time other than ZT11, blood samples were taken at 4 consecutive time points between ZT 7 and 13 on the days of proestrus and estrus (Fig. 10). Serum analysis revealed the expected proestrus surge at ZT 11 in all of the examined control 36 animals (Fig. 10B), whereas SCN-Bmal1KO females did not show concordant peaks at ZT 11 (Fig. 10A). Moreover, knockout females displayed lower LH levels at all time points as compared to the controls; two-way ANOVA and subsequent Bonferroni’s Multiple Comparison Test revealed that the mean LH levels were significantly different between the two groups at ZT 11 [t(40) = 4.007, p < 0.001; Fig. 10C] (49). As expected, LH remained at baseline levels throughout all estrus time points in the control animals, while not all knockout animals displayed baseline LH levels (Fig. 10D-F), indicating that the LH surge might have been delayed or shifted in the knockouts from proestrus to estrus day. 37 Figure 10. LH profiles on proestrus and estrus days. A and B. Proestrus levels of LH from ZT 7 to 13 for individual SCN-Bmal1KO (A, n = 6) and control (B, n = 5) females, respectively. C. Averaged data of A and B. By ANOVA: *, p < 0.001. D and E. Estrus levels of LH from ZT 7 to 13 for individual SCN-Bmal1KO (D, n = 4) and control (E, n = 4) females, respectively. F. Averaged data of D and E. Non-significant, p = 0.37, ANOVA. Bars indicate mean ± SEM. 38 Discussion The lack of reproductive phenotype displayed by GnBmal1KO mice may be due to the potent ability of kisspeptin in resetting the hypothalamic GnRH pulse generator to a new zero time point (50). Given that Kiss1 secretion itself is pulsatile (51), this suggests that the also pulsatile secretion of GnRH could be purely influenced by Kiss1 stimulation and not set by endogenous clocks within GnRH-expressing neurons. Although it has been mentioned previously that GnRH neurons require an intrinsic circadian clock for normal GnRH secretion as transient expression of the dominant negative Clock-Δ19 protein in GT1-7 cells altered GnRH pulse patterns (44), it is plausible that Kiss1 proteins can reverse altered GnRH secretory patterns by resetting the GnRH pulse generator each time upon stimulation. As a result, elimination of clock function within GnRH-expressing neurons would not eliminate normal pulsatile secretion of GnRH due to undisrupted Kiss1 stimulation, therefore producing no deficits in LH/FSH production and in other reproductive phenotypes. Indeed, both indirect and direct evidence suggests that Kiss1 neurons in the arcuate nucleus act as the central pacemaker driving pulsatile GnRH secretion (49, 51, 52). Moreover, the regulation of GnRH secretion by kisspeptin-secreting neurons are mediated via kisspeptinGPR54 signaling as female mice with deletions in the GPR54 gene (GPR54 knockouts) showed disruptions in tonic GnRH/LH secretion (53). Since Kiss1 neurons are essential in generating the full preovulatory LH surge as Kiss1 antagonist treatment significantly blunted LH surge production (54), Kiss1 appears to be a key locus of integration for circadian signaling necessary for the LH surge production and ovulation. It has been established that circadian pattern of Kiss1 expression persisted even under constant darkness (DD) conditions, suggesting that rhythms in these neurons are regulated endogenously (55). As peak Kiss1 expression in DD also coincided 39 with the LH surge (55), elimination of the endogenous clock function in Kiss1 neurons may result in faulty timing of Kiss1 secretion and thus reproductive deficits. However, the circadian rhythm of Kiss1 secretion could also be solely controlled through upstream projections from the SCN. Therefore, evaluating the involvement of the endogenous circadian clock within Kiss1 neurons or the SCN in LH surge generation and other aspects of reproduction could be seen as the next logical step in unraveling the role of the circadian timing system in reproductive physiology. Interestingly, both the WT control animals in the GnBmal1KO and the SCN-Bmal1KO experiments displayed slightly longer estrous cycle lengths—5.4±0.1 days and 6.2±0.6 days respectively—than normally reported in laboratory mice [4-5 days (56)]. It has been reported that prolonged and irregular cycles are usually observed among female mice that are grouped together in the absence of male mice (57). As the animals used in this experiment were also housed together (4 per cage) in the absence of males, this presents a possible explanation for the apparent tendency towards longer estrous cycles in the WT controls.. It is important to note that GnRH-expressing neurons are enveloped in specialized glial cells known as astrocytes (58) and that the number of astrocytes apposed to GnRH neurons undergoes circadian fluctuation on proestrus, but not estrus, in young rats (59), suggesting that astrocytic plasticity may also play a role in the timing and generation of LH surge. It has been demonstrated that astrocytes are competent circadian oscillators – astroglia cultures derived from Per1::luc rats or PER2-LUC mice express circadian rhythms that can be phase entrained and shifted by external cues such as temperature cycles (60). Although it is unknown whether these in vivo rhythms in glial plasticity are produced by clocks intrinsic to astrocytes, it is conceivable that they affect GnRH neuron-astrocyte interactions, thereby regulating GnRH secretion from 40 GnRH neurons. Of note, there is now evidence suggesting that astrocytes apposing GnRH neurons in the median eminence regulate GnRH secretion by two mechanisms. One mechanism involves the synthesis and release of growth factors including transforming growth factor (TGF) β1, insulin-like growth factor-1 (IGF-1), basic fibroblast growth factor (bFGF) and various members of the epidermal growth factor (EGF) family by hypothalamic astroglial cells which are able to regulate GnRH neuronal function (61). The other mechanism involves the highly dynamic structural relationship between astrocytes and GnRH neurons in the median eminence. As GnRH neurons and their processes are profusely juxtaposed with astrocytes, rearrangements in connectivity between the two groups of cells can affect GnRH terminal contacts, thereby controlling GnRH release into the median eminence (61). Therefore, astrocytes in close apposition to GnRH neuronal terminals in the ME may play an important role in LH surge generation and ovulation. Finally, our results demonstrated that 23.9±4.7% of GnRH-positive neurons still expressed BMAL1, thereby retaining clock function. Moreover, dual-labeling experiments showed that approximately 30% of GnRH neurons did not express GPR54 (62), effectively precluding them from transcribing Cre from the knock-in locus and thus from Bmal1 Exon 8 excision. Therefore, GPR54-negative GnRH neurons could very likely account for the incomplete Bmal1 ablation observed in GnRH-expressing neurons. As these cells still contained a functional circadian clock, they may be sufficient for the generation of the preovulatory LH surge. Notably, GPR54 is also expressed in cells in the dentate gyrus of the hippocampus, septum, anteroventral nucleus of the thalamus, posterior hypothalamus, periaqueductal grey, supramammillary and ponctine nculei, and dorsal cochlear nucleus (62). As we did not examine BMAL1 expression in the GPR54 neurons of these regions in our GnBmal1KO mice nor in their 41 WT controls, it remains to be determined if intrinsic clock function in those GPR54+ cells plays a role in reproduction. Although no reproductive phenotype was observed in GnBmal1KO animals, tissuespecific abolishment of clock function in the SCN resulted in various reproductive deficits in our SCN-Bmal1KO females. SCN-Bmal1KO mice were capable of cycling albeit with longer time spent in estrus and prolonged overall estrus cycle lengths. This is similar to what was observed in global Bmal1 knockout (Bmal1-/-) mice (13). Although a functional endogenous circadian clock is not required for estrus cyclicity, it was postulated to provide daily cues to entrain the estrus oscillator periods to multiples of 24 hours and thus phase-locking it to the solar day environment (13). Given that estrus cycle length and structure in SCN-Bmal1KO females mirror that observed previously in Bmal1-/- females (13), it is likely that the daily cues entraining the estrus oscillator period originated from the endogenous clock within the SCN. Bmal1-/- animals were shown to lack a LH surge and showed severely blunted primary FSH surge, while the secondary FSH surge remained unaffected (13). Although SCN-Bmal1KO females displayed LH surges, their levels were significantly dampened in comparison with the control animals. Moreover, SCN-Bmal1KO animals lacked tight control of surge timing with regards to estrus day but also time-of-day. Since abolishment of clock function in SCN neurons did not eliminate LH surge production while it is completely absent in Bmal1-/- animals, this suggests that other tissue clocks along the HPG axis may also contribute to or can compensate for the lack of surge induction signal that emanates from the SCN. Mice lacking clock function specifically in gonadotropes (GBmal1KO) showed a mild, if any, phenotype in terms of LH surge production, indicating that clocks in these tissues may be capable of functional compensation when the SCN clock has been compromised and vice versa. Alternatively, clocks 42 in either the Kiss1 neurons or the ovary may possess a more prominent role than gonadotrope clocks with regards to regulating LH levels. While Syt10-Cre activity has been detected throughout the seminiferous tubules of the testis (46), fluorescent reporter expression was only very sporadically detected in the ovary (K.F. Storch, unpublished), arguing that the phenotypes we observed in SCN-Bmal1KO are not due to the loss of ovarian BMAL1. In order to ensure that the attenuated LH surge observed in SCN-Bmal1KO females was not caused by a shift in surge timing, we sampled blood taken at consecutive Zeitgeber times (ZT) 7, 9, 11, and 13 on proestrus day. Whereas the control animals exhibited consistent LH surges at precisely ZT 11, SCN-Bmal1KO animals displayed substantial dysregulation in surge timing. In addition, LH surge levels in the knockout animals were reduced in comparison to the controls even when time points other than ZT11 were taken into account. These observations demonstrate that selective elimination of clock function throughout the SCN results in dysregulation of LH surge timing as well as reduction in surge amplitude on the day of proestrus. Blood was also sampled at ZT 7, 9, 11, and 13 on estrus day. As expected, control females exhibited baseline LH levels throughout estrus. SCN-Bmal1KO females, on the other hand, again displayed highly variable serum LH concentrations across the collection time points that frequently reached surge levels. Consistently, knockout animals exhibited increased LH levels in comparison to the controls at all estrus time points sampled. Overall, SCN-Bmal1KO females retained the ability to generate LH surge but lacked control in surge timing. It is important to note that approximately 7.29% of the cells in the SCN still expressed BMAL1, thereby conceivably retaining the ability to generate endogenous circadian rhythms. It is thus possible, that these remaining cells are sufficient to generate the observed LH surge, albeit in a highly dysregulated manner, likely due to their inability to create a coherent rhythmic 43 output. Interestingly, it has been shown that the dorsomedial (dm) SCN clock can act independently of the ventrolateral (vl) region of the SCN as well as the light-dark cycle to time Kiss1 expression in the AVPV (64). In addition, these dmSCN neurons synthesize and release AVP into the cerebrospinal fluid (CSF) and its rhythm is always in phase with the LH surge (64). Since the expression of the Avp gene in the SCN relies on the BMAL1/CLOCK complex binding to an E box element in the Avp promoter (65), neurons that do not express BMAL1 would be expected to lack AVP expression. Therefore, disruption of the Bmal1 gene throughout the SCN would be expected to result in a complete abolition of AVP expression in this brain structure. However, a preliminary examination of the location of the few BMAL1-positive neurons that were retained in the SCN-Bmal1KOs shows most of them to be residing in the dorsomedial region of the SCN, raising the speculation that these neurons are also AVP-positive. It might be, therefore, possible that the release of AVP from (some of) the few remaining BMAL1-positive SCN neurons in the SCN-Bmal1KO animals is sufficient to induce the observed LH surge. A future work should be aimed at determining the level of AVP expression in the SCN of the SCNBmal1KO animals. If AVP is indeed present in the SCN of these mice, it could serve as an explanation for the observed ability to still mount an LH surge despite their inability to produce sustained circadian rhythms in locomotor activity (46); if, on the other hand, AVP turns out to be completely absent from the SCN, then we could conclude that AVP is not required for LH surge generation. 44 Conclusion and Future Directions We determined a BMAL1 ablation efficacy of approximately 76.10% in the hypothalamic GnRH neurons in our GnBmal1KO line. Examinations of age and weight at puberty onset, body weight, and estrus cycle structure and length indicate no overt reproductive deficits in these animals. Moreover, assessments of LH and FSH levels also support the notion that GnBmal1KO females are reproductively normal under laboratory conditions. Therefore, endogenous clocks in GnRH neurons appear to play a minor, if any, role in LH surge production and reproduction in general. In our SCN-Bmal1KO line, we detected a BMAL1 ablation efficacy of 93.7% in the SCN proper. An examination of estrus cyclicity in these KO females indicated prolonged time spent in estrus as well as an increase in the overall estrus cycle length. Moreover, these SCN-Bmal1KO females displayed blunted LH surge levels as well as dysregulation in LH surge timing. FSH serum levels also seemed to be dysregulated in SCN-Bmal1KOs. A closer examination of LH levels on the days of proestrus and estrus revealed a lack of LH surge timing control amongst the KO females on proestrus day. Furthermore, these mice frequently showed surge levels of serum LH concentrations on estrus days, which are normally not observed in any intact rodents. Given that elimination of endogenous rhythm generation in GnRH neurons did not result in any reproductive deficits and considering the importance of Kiss1 neurons in LH surge production and reproduction, a next step towards elucidating the role of the circadian clock in reproduction physiology would be to examine the specific contribution of the clock intrinsic to Kiss1 neurons to gonadotropin control and reproduction. It would also be informative to examine the role that astrocytes in apposition to GnRH neurons and their intrinsic clocks play in LH surge and ovulation, as they have been shown to intimately regulate GnRH secretion. Moreover, as 45 global Bmal1 knockout females completely lack the preovulatory rise in LH (13) while SCNBmal1KO females retained the ability to produce LH surges albeit in a blunted manner, we hypothesize that endogenous clocks in either Kiss1 neurons or in the ovary may also be involved in regulating gonadotropin levels, representing an exciting topic for future studies. Lastly, as the position of the remaining BMAL1-positive neurons in the SCN-Bmal1KO animals seemed to be located at the so-called dorsomedial shell-region of the SCN, which harbors the AVP-producing clock cells, future experiments should also be aimed at determining if AVP is still produced in the SCN (-shell) of these animals. If AVP is absent from the SCN, then we can conclude that this peptide isn’t a requirement for LH surge initiation, which is contrary to what has been suggested in the literature. We hope that these studies will allow us to gain insights into the mechanisms that underlie shift work- and trans-meridian air-travel-related reproductive dysfunctions. Our work may, however, also provide a fresh look at the causes of common reproductive dysfunctions observed in humans such as polycystic ovarian syndrome or premature ovarian failure. 46 References 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. K. J. Tonsfeldt, P. E. Chappell, Clocks on top: the role of the circadian clock in the hypothalamic and pituitary regulation of endocrine physiology. Mol Cell Endocrinol 349, 3-‐12 (2012). F. K. Stephan, I. Zucker, Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions. Proceedings of the National Academy of Sciences of the United States of America 69, 1583-‐1586 (1972). K. A. Stokkan, S. Yamazaki, H. Tei, Y. Sakaki, M. Menaker, Entrainment of the circadian clock in the liver by feeding. Science 291, 490-‐493 (2001). S. M. Reppert, D. R. Weaver, Coordination of circadian timing in mammals. Nature 418, 935-‐941 (2002). N. Preitner et al., Orphan nuclear receptors, molecular clockwork, and the entrainment of peripheral oscillators. Novartis Found Symp 253, 89-‐99; discussion 99-‐109 (2003). T. K. Sato et al., A functional genomics strategy reveals Rora as a component of the mammalian circadian clock. Neuron 43, 527-‐537 (2004). M. M. Mahoney, Shift work, jet lag, and female reproduction. International journal of endocrinology 2010, 813764 (2010). C. C. Lawson et al., Rotating shift work and menstrual cycle characteristics. Epidemiology 22, 305-‐312 (2011). S. Labyak, S. Lava, F. Turek, P. Zee, Effects of shiftwork on sleep and menstrual function in nurses. Health care for women international 23, 703-‐714 (2002). L. Bisanti, J. Olsen, O. Basso, P. Thonneau, W. Karmaus, Shift work and subfecundity: a European multicenter study. European Study Group on Infertility and Subfecundity. Journal of occupational and environmental medicine / American College of Occupational and Environmental Medicine 38, 352-‐358 (1996). M. J. Boden, T. J. Varcoe, A. Voultsios, D. J. Kennaway, Reproductive biology of female Bmal1 null mice. Reproduction 139, 1077-‐1090 (2010). C. K. Ratajczak, K. L. Boehle, L. J. Muglia, Impaired steroidogenesis and implantation failure in Bmal1-‐/-‐ mice. Endocrinology 150, 1879-‐1885 (2009). A. Chu et al., Global but not gonadotrope-‐specific disruption of Bmal1 abolishes the luteinizing hormone surge without affecting ovulation. Endocrinology 154, 2924-‐ 2935 (2013). B. H. Miller et al., Circadian clock mutation disrupts estrous cyclicity and maintenance of pregnancy. Curr Biol 14, 1367-‐1373 (2004). H. Oster, G. T. van der Horst, U. Albrecht, Daily variation of clock output gene activation in behaviorally arrhythmic mPer/mCry triple mutant mice. Chronobiology international 20, 683-‐695 (2003). M. J. Skynner, R. Slater, J. A. Sim, N. D. Allen, A. E. Herbison, Promoter transgenics reveal multiple gonadotropin-‐releasing hormone-‐I-‐expressing cell populations of different embryological origin in mouse brain. The Journal of neuroscience : the official journal of the Society for Neuroscience 19, 5955-‐5966 (1999). Z. Naor, Signaling by G-‐protein-‐coupled receptor (GPCR): studies on the GnRH receptor. Frontiers in neuroendocrinology 30, 10-‐29 (2009). 47 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. 32. A. J. Pawson, A. S. McNeilly, The pituitary effects of GnRH. Animal reproduction science 88, 75-‐94 (2005). J. W. Everett, C. H. Sawyer, A 24-‐hour periodicity in the "LH-‐release apparatus" of female rats, disclosed by barbiturate sedation. Endocrinology 47, 198-‐218 (1950). S. J. C. Legan, G.A.; Karsch, F.J., Role of Estrogen as Initiator of Daily LH Surges in the Ovariectomized Rat. Endocrinology 96, 50-‐56 (1975). G. D. S. Gray, P.; Tallentire, D.; Davidson, J.M., Effects of lesions in various structures of the suprachiasmatic-‐preoptic region on LH regulation and sexual behavior in female rats. Neuroendocrinology 25, 174-‐191 (1978). W. J. Schwartz, P. Zimmerman, Lesions of the suprachiasmatic nucleus disrupt circadian locomotor rhythms in the mouse. Physiol Behav 49, 1283-‐1287 (1991). S. Wen et al., Functional characterization of genetically labeled gonadotropes. Endocrinology 149, 2701-‐2711 (2008). H. O. de la, Antiphase Oscillation of the Left and Right Suprachiasmatic Nuclei. Science 290, 799-‐801 (2000). H. O. de la Iglesia, J. Meyer, W. J. Schwartz, Lateralization of circadian pacemaker output: Activation of left-‐ and right-‐sided luteinizing hormone-‐releasing hormone neurons involves a neural rather than a humoral pathway. The Journal of neuroscience : the official journal of the Society for Neuroscience 23, 7412-‐7414 (2003). R. E. Watson, Jr., M. C. Langub, Jr., M. G. Engle, B. E. Maley, Estrogen-‐receptive neurons in the anteroventral periventricular nucleus are synaptic targets of the suprachiasmatic nucleus and peri-‐suprachiasmatic region. Brain research 689, 254-‐ 264 (1995). B. Vida et al., Evidence for suprachiasmatic vasopressin neurones innervating kisspeptin neurones in the rostral periventricular area of the mouse brain: regulation by oestrogen. Journal of neuroendocrinology 22, 1032-‐1039 (2010). W. P. Williams, 3rd, S. G. Jarjisian, J. D. Mikkelsen, L. J. Kriegsfeld, Circadian control of kisspeptin and a gated GnRH response mediate the preovulatory luteinizing hormone surge. Endocrinology 152, 595-‐606 (2011). W. W. Le, K. A. Berghorn, S. Rassnick, G. E. Hoffman, Periventricular preoptic area neurons coactivated with luteinizing hormone (LH)-‐releasing hormone (LHRH) neurons at the time of the LH surge are LHRH afferents. Endocrinology 140, 510-‐519 (1999). A. Kalsbeek, R. M. Buijs, M. Engelmann, C. T. Wotjak, R. Landgraf, In vivo measurement of a diurnal variation in vasopressin release in the rat suprachiasmatic nucleus. Brain research 682, 75-‐82 (1995). T. Funabashi, K. Shinohara, D. Mitsushima, F. Kimura, Gonadotropin-‐releasing hormone exhibits circadian rhythm in phase with arginine-‐vasopressin in co-‐ cultures of the female rat preoptic area and suprachiasmatic nucleus. Journal of neuroendocrinology 12, 521-‐528 (2000). I. F. Palm, E. M. Van Der Beek, V. M. Wiegant, R. M. Buijs, A. Kalsbeek, Vasopressin induces a luteinizing hormone surge in ovariectomized, estradiol-‐treated rats with lesions of the suprachiasmatic nucleus. Neuroscience 93, 659-‐666 (1999). 48 33. 34. 35. 36. 37. 38. 39. 40. 41. 42. 43. 44. 45. 46. 47. 48. I. F. Palm, E. M. van der Beek, V. M. Wiegant, R. M. Buijs, A. Kalsbeek, The stimulatory effect of vasopressin on the luteinizing hormone surge in ovariectomized, estradiol-‐ treated rats is time-‐dependent. Brain research 901, 109-‐116 (2001). B. H. Miller et al., Vasopressin regulation of the proestrous luteinizing hormone surge in wild-‐type and Clock mutant mice. Biology of reproduction 75, 778-‐784 (2006). E. M. Van der Beek, T. L. Horvath, V. M. Wiegant, R. Van den Hurk, R. M. Buijs, Evidence for a direct neuronal pathway from the suprachiasmatic nucleus to the gonadotropin-‐releasing hormone system: combined tracing and light and electron microscopic immunocytochemical studies. The Journal of comparative neurology 384, 569-‐579 (1997). M. J. Smith, L. Jiennes, P. M. Wise, Localization of the VIP2 receptor protein on GnRH neurons in the female rat. Endocrinology 141, 4317-‐4320 (2000). E. M. van der Beek et al., Preferential induction of c-‐fos immunoreactivity in vasoactive intestinal polypeptide-‐innervated gonadotropin-‐releasing hormone neurons during a steroid-‐induced luteinizing hormone surge in the female rat. Endocrinology 134, 2636-‐2644 (1994). C. A. Christian, S. M. Moenter, Vasoactive intestinal polypeptide can excite gonadotropin-‐releasing hormone neurons in a manner dependent on estradiol and gated by time of day. Endocrinology 149, 3130-‐3136 (2008). R. F. Weick, K. M. Stobie, Vasoactive intestinal peptide inhibits the steroid-‐induced LH surge in the ovariectomized rat. The Journal of endocrinology 133, 433-‐437 (1992). Y. Sun, J. Shu, K. Kyei, G. S. Neal-‐Perry, Intracerebroventricular infusion of vasoactive intestinal Peptide rescues the luteinizing hormone surge in middle-‐aged female rats. Frontiers in endocrinology 3, 24 (2012). H. Dolatshad et al., Developmental and reproductive performance in circadian mutant mice. Human reproduction 21, 68-‐79 (2006). J. R. Hickok, S. A. Tischkau, In vivo circadian rhythms in gonadotropin-‐releasing hormone neurons. Neuroendocrinology 91, 110-‐120 (2010). J. M. Gillespie, B. P. Chan, D. Roy, F. Cai, D. D. Belsham, Expression of circadian rhythm genes in gonadotropin-‐releasing hormone-‐secreting GT1-‐7 neurons. Endocrinology 144, 5285-‐5292 (2003). P. E. Chappell, R. S. White, P. L. Mellon, Circadian gene expression regulates pulsatile gonadotropin-‐releasing hormone (GnRH) secretory patterns in the hypothalamic GnRH-‐secreting GT1-‐7 cell line. The Journal of neuroscience : the official journal of the Society for Neuroscience 23, 11202-‐11213 (2003). K. F. Storch et al., Intrinsic circadian clock of the mammalian retina: importance for retinal processing of visual information. Cell 130, 730-‐741 (2007). J. Husse, X. Zhou, A. Shostak, H. Oster, G. Eichele, Synaptotagmin10-‐Cre, a driver to disrupt clock genes in the SCN. Journal of biological rhythms 26, 379-‐389 (2011). C. Mayer, U. Boehm, Female reproductive maturation in the absence of kisspeptin/GPR54 signaling. Nature neuroscience 14, 704-‐710 (2011). T. K. Woodruff et al., Inhibin A and inhibin B are inversely correlated to follicle-‐ stimulating hormone, yet are discordant during the follicular phase of the rat 49 49. 50. 51. 52. 53. 54. 55. 56. 57. 58. 59. 60. 61. 62. estrous cycle, and inhibin A is expressed in a sexually dimorphic manner. Endocrinology 137, 5463-‐5467 (1996). A. K. Roseweir et al., Discovery of potent kisspeptin antagonists delineate physiological mechanisms of gonadotropin regulation. The Journal of neuroscience : the official journal of the Society for Neuroscience 29, 3920-‐3929 (2009). Y. M. Chan et al., Kisspeptin resets the hypothalamic GnRH clock in men. J Clin Endocrinol Metab 96, E908-‐915 (2011). K. L. Keen, F. H. Wegner, S. R. Bloom, M. A. Ghatei, E. Terasawa, An increase in kisspeptin-‐54 release occurs with the pubertal increase in luteinizing hormone-‐ releasing hormone-‐1 release in the stalk-‐median eminence of female rhesus monkeys in vivo. Endocrinology 149, 4151-‐4157 (2008). Y. Wakabayashi et al., Neurokinin B and dynorphin A in kisspeptin neurons of the arcuate nucleus participate in generation of periodic oscillation of neural activity driving pulsatile gonadotropin-‐releasing hormone secretion in the goat. The Journal of neuroscience : the official journal of the Society for Neuroscience 30, 3124-‐3132 (2010). H. M. Dungan et al., The role of kisspeptin-‐GPR54 signaling in the tonic regulation and surge release of gonadotropin-‐releasing hormone/luteinizing hormone. The Journal of neuroscience : the official journal of the Society for Neuroscience 27, 12088-‐ 12095 (2007). J. T. Smith et al., Kisspeptin is essential for the full preovulatory LH surge and stimulates GnRH release from the isolated ovine median eminence. Endocrinology 152, 1001-‐1012 (2011). L. J. Kriegsfeld, Circadian regulation of kisspeptin in female reproductive functioning. Adv Exp Med Biol 784, 385-‐410 (2013). C. S. Caligioni, Assessing reproductive status/stages in mice. Curr Protoc Neurosci Appendix 4, Appendix 4I (2009). F. S. vom Saal, F. H. Bronson, Variation in length of the estrous cycle in mice due to former intrauterine proximity to male fetuses. Biology of reproduction 22, 777-‐780 (1980). K. M. Glanowska, S. M. Moenter, Endocannabinoids and prostaglandins both contribute to GnRH neuron-‐GABAergic afferent local feedback circuits. J Neurophysiol 106, 3073-‐3081 (2011). A. B. Cashion, M. J. Smith, P. M. Wise, The morphometry of astrocytes in the rostral preoptic area exhibits a diurnal rhythm on proestrus: relationship to the luteinizing hormone surge and effects of age. Endocrinology 144, 274-‐280 (2003). L. M. Prolo, J. S. Takahashi, E. D. Herzog, Circadian rhythm generation and entrainment in astrocytes. The Journal of neuroscience : the official journal of the Society for Neuroscience 25, 404-‐408 (2005). S. R. Ojeda, A. Lomniczi, U. S. Sandau, Glial-‐gonadotrophin hormone (GnRH) neurone interactions in the median eminence and the control of GnRH secretion. Journal of neuroendocrinology 20, 732-‐742 (2008). A. E. Herbison, X. de Tassigny, J. Doran, W. H. Colledge, Distribution and postnatal development of Gpr54 gene expression in mouse brain and gonadotropin-‐releasing hormone neurons. Endocrinology 151, 312-‐321 (2010). 50 63. 64. 65. M. D. Muzumdar, B. Tasic, K. Miyamichi, L. Li, L. Luo, A global double-‐fluorescent Cre reporter mouse. Genesis 45, 593-‐605 (2007). B. L. Smarr, E. Morris, H. O. de la Iglesia, The dorsomedial suprachiasmatic nucleus times circadian expression of Kiss1 and the luteinizing hormone surge. Endocrinology 153, 2839-‐2850 (2012). X. Jin et al., A molecular mechanism regulating rhythmic output from the suprachiasmatic circadian clock. Cell 96, 57-‐68 (1999). 51