Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
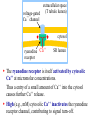
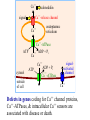
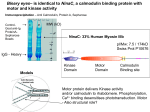
Advanced Biochemistry Chapter 02 Transport and Signaling Biochemistry of Metabolism Lecture 06 Calcium Signals Ca++ Ca++-release channel signal Ca Modulation of cytosolic Ca++: Cytosolic [Ca++] is usually <1 μM, except during a Ca++ signal event. ATP calmodulin Ca endoplasmic reticulum ++ Ca++-ATPase ++ ADP + Pi Ca++ cytosol outside of cell ATP ADP + Pi Ca++-ATPase signalactivated channel Ca++ Ca++-ATPase pumps in plasma membranes & ER membranes maintain this low concentration by transporting Ca++ away from the cytosol, out of the cell or into the ER. Ca++ Extracellular [Ca++] in mammals is in the mM range. Opening of plasma membrane Ca++ channels may initiate or sustain a Ca++ signal. Ca++-release channel signal Ca ATP calmodulin Ca endoplasmic reticulum ++ Ca++-ATPase ++ ADP + Pi Ca++ cytosol outside of cell ATP ADP + Pi Ca++-ATPase signalactivated channel Ca++ [Ca++] is also relatively high in the lumen of the ER, which serves as the major internal reservoir from which Ca++ is released to the cytosol during Ca++ signaling. Mitochondria and lysosomes also serve as reservoirs for Ca++ subject to release under certain conditions. Ca++-binding proteins in the ER lumen "buffer" free [Ca++], and increase the capacity for Ca++ storage. ER Ca++-binding proteins have 20-50 low-affinity Ca++binding sites per molecule, consisting of acidic residues. Examples: Calsequestrin is in the lumen of the sarcoplasmic reticulum (SR), a specialized ER of muscle. Calreticulin is in the lumen of the ER of non-muscle cells. It also has a role in protein folding. Ca++ concentration, within the cytosol or other cell compartments, may be monitored using indicator dyes or proteins that are either luminescent or change their fluorescence when they bind Ca++. Fluorescent indicators used with confocal fluorescence microscopy can provide high-resolution imaging and quantitation of Ca++ fluctuations within cells. Ca++ regulates many cellular reactions and processes. A transient increase in cytosolic Ca++ may be localized to the vicinity of one or a few Ca++release or Ca++entry channels. Ca++ Ca++-release channel signal Ca ATP calmodulin Ca endoplasmic reticulum ++ Ca++-ATPase ++ ADP + Pi Ca++ signalactivated channel ADP + Pi Such a localized ATP cytosol ++ ++ Ca -ATPase Ca “puff” or outside “spark” may Ca++ of cell activate effectors that induce additional Ca++ release, leading to a more widespread increase in cytosolic Ca++. A “wave” of higher Ca++ may spread to neighboring cells. For example, see a website maintained by E. Niggli showing recordings of Ca++ sparks and waves, using fluorescent Ca++ indicators. Ryanodine Receptor: A Ca++ Release Channel A large Ca++ release channel in the membrane of muscle sarcoplasmic reticulum (SR) is called the ryanodine receptor, because of sensitivity to inhibition by a plant alkaloid ryanodine. Skeletal and cardiac muscle contraction is activated when Ca++ is released from the SR lumen to the cytosol via the ryanodine receptor. extracellular space T tubules: invaginations of (T tubule lumen) voltage-gated muscle plasma membrane. Ca++ channel Voltage-gated Ca++ channels in the T tubule membrane interact with ryanodine receptors in the closely apposed SR membrane. cytosol ++ Ca ryanodine SR lumen receptor Activation of voltage-gated Ca++ channels, by an action potential in the T tubule, leads to opening of ryanodinesensitive Ca++-release channels. Ca++ moves from the SR lumen to the cytosol, passing through the transmembrane part of the ryanodine receptor, & then through the receptor's cytoplasmic assembly. extracellular space (T tubule lumen) voltage-gated Ca++ channel cytosol ++ Ca ryanodine SR lumen receptor The ryanodine receptor is itself activated by cytosolic Ca++ at micromolar concentrations. Thus a entry of a small amount of Ca++ into the cytosol causes further Ca++ release. High (e.g., mM) cytosolic Ca++ inactivates the ryanodine receptor channel, contributing to signal turn-off. Three views of a 3D reconstruction of the structure of the ryanodine-sensitive calcium channel at 30 Å resolution, based on micrographs obtained by EM at varied tilt angles. These images were provided by Terrence Wagenknecht of the Wadsworth Center, NY State Dept. of Health. Animation of conformational changes during channel opening & closing. A somewhat higher resolution structure of the ryanodine receptor channel now available indicates the presence of bent a-helices adjacent to the lumen in the transmembrane pore domain. But an atomic resolution structure of the whole channel has not yet been achieved. For diagrams see article by Ludtke et al. (journal subscription required). IP3 receptor Ca++ OPO32 H release channel In many mammalian cells, IP3 (inositol-1,4,5-trisphosphate) triggers Ca++ release from the ER. The 2nd messenger IP3 is produced, e.g., in response to hormonal signals, from the membrane lipid phosphatidylinositol. The IP3 receptor is a ligand-gated Ca++-release channel embedded in ER membranes. OPO32 OH H OH OH H H H H IP3 OPO32 inositol-1,3,4-trisphosphate Ca++ Ca++-release channel IP3 ++ Ca ATP calmodulin endoplasmic reticulum Ca++-ATPase ++ ADP + Pi Ca Ca++ Ca++-release channel IP3 ++ Ca ATP calmodulin endoplasmic reticulum Ca++-ATPase ADP + Pi Ca++ The IP3 receptor (IP3-activated Ca++-release channel) is distinct from but partly homologous to the ryanodine receptor channel. IP3 binds to a cytosolic domain of the receptor promoting channel opening. IP3 may displace a regulatory phospho-protein IRBIT, which binds at the same site. Diagram (RIKEN Inst) Ca++ also binds to the ligand-binding domain of the IP3 receptor, & promotes channel opening. Ca++ Ca++-release channel IP3 ++ Ca However, high cytosolic Ca++, which develops after channel opening, promotes channel closure. ATP calmodulin endoplasmic reticulum Ca++-ATPase ++ ADP + Pi Ca Thus both IP3-activated & ryanodine-sensitive channels are activated by low cytosolic Ca++ & inhibited by high Ca++. The feedback inhibition of Ca++ release by high cytosolic Ca++, along with activity of Ca++-ATPase pumps, contributes to signal turnoff & makes possible observed oscillations in Ca++ concentration. Ca++ Ca++-release channel IP3 ++ Ca View an animation of the overall process of Ca++ cycling. ATP calmodulin endoplasmic reticulum Ca++-ATPase ADP + Pi Ca++ Structures of cytosolic domains of the IP3 receptor, including the IP3 binding site, have been solved, but the pore structure of the receptor has not yet been determined at atomic resolution. See website for a low-resolution structure of IP3 receptor. (AIST, Japan, findings of C. Sato et al.) PDB 1CDM glutamate H H3N+ COO C CH2 Calmodulin, a Ca++activated switch protein, mediates many of the signal functions of Ca++. Calmodulin cooperatively binds 4 Ca++. CH2 C O ++ Ca O aspartate (Asp) H H3N+ helix-loop-helix motif in calmodulin C COO CH2 COO At each binding site, Ca++ interacts with O atoms, mainly of Glu & Asp side-chain carboxyl groups, & of the protein backbone, in a loop between 2 a-helices at right angles. This helix-loop-helix motif is called an EF hand. There are 4 helix-loophelix motifs, 2 at each end of calmodulin, which is dumbbell shaped. Calmodulin Ca++ ( ) Ca++ binding promotes a Target conformational change peptide that exposes hydrophobic residues along a concave patch on each of the 2 lobes. These are involved in protein-protein interactions. Ca++-calmodulin then changes conformation again as it wraps around the target domain of a protein. A typical calmodulin-binding target domain is a (+) charged, amphipathic a-helix, with polar & non-polar surfaces. Terminal methyl groups of Met side-chains of calmodulin participate in binding to hydrophobic residues in target domains of some enzymes regulated by calmodulin. methionine (Met) H H3N+ C COO CH2 CH2 S CH3 However the interaction of Ca++-calmodulin with some target proteins is different from what is described here. Some proteins have bound calmodulin as part of their quaternary structure, even in the absence of Ca++. In either case, Ca++ binding to calmodulin may induce a conformational change that alters target protein activity. Many enzymes are regulated by Ca++-calmodulin. E.g.: Some protein kinases that transfer phosphate from ATP to hydroxyl residues on other enzymes to be regulated, are activated by Ca++-calmodulin. These are referred to as CaM Kinases. The plasma membrane Ca++-ATPase that pumps Ca++ out of the cell is one of the target proteins activated by Ca++-calmodulin. Thus cytosolic Ca++ itself contributes further to turning off Ca++ signals. View an animation of Ca++-activated binding of calmodulin to a target peptide. Ca++ Ca++-release channel signal Ca ATP calmodulin Ca endoplasmic reticulum ++ Ca++-ATPase ++ ADP + Pi Ca++ cytosol outside of cell ATP ADP + Pi Ca++-ATPase signalactivated channel Ca++ Defects in genes coding for Ca++ channel proteins, Ca++-ATPases, & intracellular Ca++ sensors are associated with disease or death.