Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
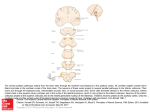
SUPPLEMENTARY NOTE Effects of scanner noise on MEMRI results MRI scanner noise, which presents a unique problem for fMRI studies of the auditory brainS1, should not contribute significantly to the MEMRI signals in mice for several reasons. First, mice are more sensitive to higher than lower frequency sound from the onset of hearingS2,S3, and the noise characteristics of our MRI protocol has no significant frequency components above 5-kHz while the sound stimuli used for these studies covered the major audible range of mice, up to 60-kHz (Supplementary Fig. 1). Furthermore, the MEMRI signal reflects the accumulative effect of sound stimulation over the 24-h exposure period, which should dominate any effects due to MRI noise over the 2-h imaging period. Finally, the mice in these studies were anesthetized during imaging which is expected to significantly decrease brain metabolism and activity during the acquisition of MRI dataS4. The excellent agreement of our results with those produced by other non-MRI mapping methods confirms that MEMRI provides an accurate measure of sound-evoked activity, independent of scanner noise. Effects of hearing loss during development Previous studies have shown that unilateral sound deprivation or cochlear ablation during early stages of development can induce cellular alterations in the auditory brainstem. Unilateral sound deprivation during early stages of ferret development was reported to induce an increase in the number of ipsilateral projections from the functional CN to ICS5, similar to the effects seen after cochlear ablation in neonatal gerbils and ferretsS6,S7. In the case of cochlear ablation during early postnatal development, the resulting sensorineural hearing loss is associated with more profound neuroplasticity changes than CHL, including an increased sensitivity and discharge level in response to stimulation of IC neurons ipsilateral to the functional earS8. At this point, there are few data on the neuroplasticity effects of CHL during early postnatal brain development. Future studies, combining MEMRI and histological analyses, should provide important insights into the cellular alterations and subsequent changes in auditory activity in a variety of mouse models of hearing loss. Transport of systemically administered Mn into neurons Relevant to our studies are the mechanisms of transport of the IP injected Mn into the brain. It is known that Mn can bind transferrin (Tf-Mn) and then be transported across the BBB via receptor-mediated endocytosisS9. Nevertheless, the same level of injected Mn is taken up in normal and hypotransferrinemic miceS10, demonstrating that other transport systems also mediate Mn uptake. Furthermore, free Mn is transported into the brain more rapidly than Tf-MnS11 , again suggesting that Tf-mediated transport is not the primary system for Mn uptake. At the level of MRI detection, previous studies have shown that injected Mn rapidly accumulates in the ventricular cerebral spinal fluid, and is taken up in the brain parenchyma more slowly over a period of hours unless the BBB is disruptedS12S15 , suggesting that Mn uptake is via absorption through the ependymal surfaces. Additionally, there may be an axonal tract-tracing component to the Mn uptake, which can cross multiple synapses as demonstrated in the olfactory systemS16. A number of studies indicate that a primary mechanism for transporting divalent Mn2+ into neurons is through calcium channelsS17,S18 , which provided the original motivation to develop MEMRI approachesS12. Indeed, Mn2+ entry through calcium channels is used to quench the fluorescence of fura-2 in a standard, widely applied assay to measure cellular Ca2+ influxS19. Taken together, these data indicate that systemically administered Mn 2 diffuses into the brain in free ionic form, entering cells either directly through calcium channels, or indirectly through tract tracing, providing an excellent opportunity for apping brain activity with MRI. Supplementary References S1. Amaro, E. Jr., Williams, S.C., Shergill, S.S., Fu, C.H., MacSweeney, M., Picchioni, M.M., Brammer, M.J., & McGuire, P.K. Acoustic noise and functional magnetic resonance imaging: current strategies and future prospects. J. Magn. Reson. Imaging 16, 497-510 (2002). S2. Romand, R., & Ehret, G. Development of tonotopy in the inferior colliculus. I. Electrophysiological mapping in house mice. Brain Res. Dev. Brain Res. 54, 221-234 (1990). S3. Saunders, J.C., Dolgin. K.G., & Lowry, L.D. The maturation of frequency selectivity in C57BL/6J mice studied with auditory evoked response tuning curves. Brain Res. 187, 69-79 (1980). S4. Wang, Z.X., Ryan, A.F., & Woolf, N.K. Pentobarbital and ketamine alter the pattern of 2-deoxyglucose uptake in the central auditory system of the gerbil. Hear. Res. 27, 145-155 (1987). S5. Moore, D. R., Hutchings, M.E., King, A.J., & Kowalchuk, N.E. Auditory brainstem of the ferret: some effects of rearing with a unilateral ear plug on the cochlea, cochlear nucleus, and projections to the inferior colliculus. J. Neurosci. 9, 1213-1222 (1989). S6. Nordeen, K. W., Killackey, H.P., & Kitzes, L.M. Ascending projections to the inferior colliculus following unilateral cochlear ablation in the neonatal gerbil, Meriones unguiculatus. J. Comp. Neurol. 214, 144-153 (1983). 3 S7. Moore, D. R., & Kowalchuk, N.E. Auditory brainstem of the ferret: effects of unilateral cochlear lesions on cochlear nucleus volume and projections to the inferior colliculus. J. Comp. Neurol. 272, 503-515 (1988). S8. Kitzes, L. M., & Semple, M.N. Single-unit responses in the inferior colliculus: effects of neonatal unilateral cochlear ablation. J. Neurophysiol. 53, 1499-1516 (1985). S9. Aschner, M., & Gannon, M. Manganese transport across the rat blood-brain barrier: saturable and transferrin-dependent transport mechanisms. Brain Res. Bull. 33, 345349 (1994). S10. Malecki, E. A., Devenyi, A.G., Beard, J.L., & Connor, J.R. Transferrin response in normal and iron-deficient mice heterozygotic for hypotransferrinemia; effects on iron and manganese accumulation. Biometals 11, 265-276 (1998). S11. Sotogaku, N., Oku, N., & Takeda, A. Manganese concentration in mouse brain after intravenous injection. J. Neurosci. Res. 61, 350-356 (2000). S12. Lin, Y. & Koretsky, A.P. Manganese ion enhanced T1-weighted MRI during brain activation: an approach to direct imaging of brain function. Magn. Reson. Med. 38, 378-388 (1997). S13. Watanabe, T., Natt, O., Boretius, S., Frahm, J., & Michaelis, T. In vivo 3D MRI staining of mouse brain after subcutaneous application of MnCl2. Magn. Reson. Med. 48, 852-859 (2002). S14. Aoki, I., Wu, Y.J., Silva, A.C., Lynch, R.M., & Koretsky, A.P. In vivo detection of neuroarchitecture in the rodent brain using manganese-enhanced MRI. Neuroimage 22, 1046-59 (2004). 4 S15. Zaim Wadghiri, Y., Blind, J.A., Duan, X., Moreno, C., Yu, X., Joyner, A.L., & Turnbull, D.H. Manganese-enhanced magnetic resonance imaging (MEMRI) of mouse brain development. NMR Biomed. 17, 613-619 (2004). S16. Pautler, R. G., & Koretsky, A.P. Tracing odor-induced activation in the olfactory bulbs of mice using manganese-enhanced magnetic resonance imaging. Neuroimage 16, 441-448 (2002). S17. Ochi, R. Manganese action potentials in mammalian cardiac muscle. Experimentia 31, 1048-1049 (1975). S18. Drapeau, P., & Nachsen, D.A. Manganese fluxes and manganese-dependent neurotransmitter release in presynaptic nerve endings isolated from rat brain. J. Physiol. 348, 493-510 (1984). S19. Merritt, J. E., Jacob, R., & Hallam, T.J. Use of manganese to discriminate between calcium influx and mobilization from internal stores in stimulated human neutrophils. J. Biol. Chem. 264, 1522-1527 (1989). 5 Peak sound pressure level (dB) Supplementary Figure 1 Frequency spectra measured for the MRI scanner and the sound-stimuli used in these MEMRI experiments. 90 a 90 b 90 60 60 60 30 30 30 0 0 0 90 20 40 60 80 100 120 d 0 0 90 20 40 60 80 100 120 e 0 90 60 60 60 30 30 30 0 0 0 20 40 60 80 100 120 c 20 40 60 80 100 120 20 40 60 80 100 120 f 0 0 20 40 60 80 100 120 0 Frequency (kHz) (a) Background noise in the MRI room, with the scanner turned off. (b) Noise measured 2.5-m in front of the magnet bore with the gradient-echo sequence running. (c-f) Noise measured at the center of the cage, inside the acoustic isolation chamber during the presentation of sound stimuli: (c) no stimulation; (d) broadband, 1-59 kHz; (e) High frequency broadband, 20-50 kHz; (f) Pure tone, 40 kHz. In each case, the maximum amplitude by frequency is plotted for a 10-s acquisition period. Supplementary Figure 2 Sound-stimulation produced significant MEMRI enhancement in auditory brainstem nuclei. MR images were acquired before (Pre) and 24-h after (Post 24h) injection of MnCl2. For the 24-h period following MnCl2 injection, mice were maintained in the acoustic isolation chamber with either no sound stimulation (a; n = 7) or with sound stimulation (b; n = 7). Compared to no stimulation, the mice exposed to 24-h of sound stimulation showed significant enhancement (two-tail t-test) in both inferior colliculus (IC, arrows) and cochlear nucleus (CN, arrowheads), (c; *P < 0.05, n = 7), while there was no difference in the caudate putamen (CPu). Data were analyzed by first defining the MEMRI signal change in each brain region: CN' = CNPost - CNPre; IC' = ICPost - ICPre; CPu' = CPuPost - CPuPre. As in other data presented, enhancement was normalized to the CPu in each mouse: Enhancement = [(IC', CN') – CPu'] / CPu'. (a) Images are shown pre- and 24-h post- MnCl2 injection at P21 and then at P6w in the same mouse, which experienced uni-CHL at P21 (top time line). MEMRI enhancement can be seen in the ipsi-CN (arrowhead) at both P21 and P6w, and in the contra-IC (arrow), referenced to the functional ear at P21 only. The pre-scan at P6w shows that the Mn has cleared from the previous injection at P21. Images from a second mouse, acquired pre- and 24-h post- MnCl2 injection once only at P6w show similar ipsi-CN enhancement (arrowhead; bottom timeline). (b) Quantitative MEMRI data acquired at P6w from mice injected twice with MnCl2 at P21 and P6w (n = 7; right) were the same as those injected only once at P6w (n = 7; left). In both cases, enhancement was significant (two-tail t-test) only in ipsi-CN (*P < 0.001) but not in contra-IC.