Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
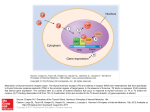
Comparative Biochemistry and Physiology Part B 132 (2002) 299–308 Review Internalization of the chicken growth hormone receptor complex and its effect on biological functions夞 ¨ a,*, Lieve Vleuricka, Marc Ederyb, Eddy Decuyperec, Veerle M. Darrasa Eduard R. Kuhn a Laboratory of Comparative Endocrinology, Zoological Institute, K.U. Leuven, B-3000 Leuven, Belgium INSERM U344, Faculte´ de Medicine Necker, 156, rue de Vaugirard, F-75730 Paris Cedex 15, France c Laboratory of Physiology and Immunology of Domestic Animals, Faculty of Agriculture and Applied Biological Sciences, K.U. Leuven, B-3001 Leuven, Belgium b Received 27 January 2001; received in revised form 17 April 2001; accepted 2 May 2001 Abstract In the chicken, as in mammals, GH is a pleiotropic cytokine that plays a central role in growth differentiation and metabolism by altering gene expression in target cells. In the growing and adult chicken it stimulates gene expression of IGF-I and inhibits gene transcription of the type III deiodinating enzyme (D3) and by doing so also increases T3 concentrations. GH binding to its receptor leads to internalization of the GH–GHR complex to the Golgi apparatus. This process is linked to the episodic release pattern of GH during growth. At the same time, a sharp decline of the expression of cGHR occurs at hatching. An in vitro study using a COS-7 cell line transfected with the cDNA of the chicken GHR, revealed that GHR immunofluorescence was found in the perinuclear region and on the plasma membrane. Following GH-induced internalization, GH and GHR were colocalized in endocytic and later in large lysosomal vesicles. Neither receptor nor ligand was transferred to the nucleus as confirmed by confocal laser microscopy. The JAKySTAT pathway however, as reported for mammalian GH receptors, mediated GH-induced gene transcription in chickens. 䊚 2002 Elsevier Science Inc. All rights reserved. Keywords: Colocalization; Endocytosis of GH–GHR; Gene transcription; Golgi apparatus; Growth hormone; Ontogeny of receptors; Thyroid; Growth 1. Introduction Several hormones are involved in the control of intermediary metabolism and growth of which the 夞 This paper was submitted as part of the proceedings of the 20th Conference of European Comparative Endocrinologists, organized under the auspices of the European Society of Comparative Endocrinology, held in Faro, Portugal, September 5–9, 2000. *Corresponding author. Tel.: q32-16-32-3990; fax: q3216-32-4262. E-mail address: ¨ [email protected] (E.R. Kuhn). main ones can be divided into at least three groups: a. GH and the insulin-like growth factors (IGF-I and II): these hormones are hydrophylic and have plasma membrane bound receptors. b. The thyroid hormones and other growth factors. c. Steroids. During the life cycle of vertebrates, different endocrine mechanisms may control growth and the intermediary metabolism. So, during embryonic and fetal growth body mass increases by both cell multiplication and cell enlargement. GH seems not 1096-4959/02/$ - see front matter 䊚 2002 Elsevier Science Inc. All rights reserved. PII: S 1 0 9 6 - 4 9 5 9 Ž 0 2 . 0 0 0 3 7 - 4 300 ¨ et al. / Comparative Biochemistry and Physiology Part B 132 (2002) 299–308 E.R. Kuhn Fig. 1. Relative importance of hormones in vertebrate growth: B, birth or hatching; and P, puberty. to be implicated, but insulin, and possibly the IGFs together with thyroid hormones may play a dominant role. Recently, GH receptor (GHR) binding was demonstrated in the fetal rat liver together with downstream signaling via the Janus kinase-2 tyrosine kinase and signal transducer and activator of transcription (STAT1 and STAT5) (Phornphutkul et al., 2000), whereas in the chicken embryo the GH gene is expressed in numerous extrapituitary sites (Harvey et al., 2000). This may implicate the presence of other, still unknown activities for GH during this period. Only after parturition or hatching, GH will become important for growth by stimulating the production of IGF and interacting profoundly with thyroid hormones. Gonadal steroids will induce a growth spur by fusing the epiphyseal cartilage left and by doing so, will terminate at least the process of longitudinal growth. All hormones again are involved in maintaining body weight in the postpuberal period and in preparing the animal for a successful reproductive phase (Fig. 1). Thyroid hormones and steroids are hydrophobic and directly stimulate the rate of gene transcription and protein synthesis by binding on nuclear receptors. However, GH will only be biologically active after association with its receptor located in the plasma membrane of a target cell. The existence of dwarf lines in chicken mutated in their GHR gene illustrates this principle (Decuypere et al., 1991; Burnside et al., 1991). The consequent physiological alterations are very similar to the situation known in the human Laron type dwarf- ism, which is also the result of a GHR dysfunction (Brown et al., 1993). In both dwarf lines, embryonic and fetal growth is normal, stressing the fact that GHR functionality is not required for normal growth prenatally. 2. Link between GH productionybiological activity and thyroid hormone The activity of the somatotropic and thyrotropic axes are tightly linked together in mammalian and avian species. In mammals, GH does not always control growth rate and does not always stimulate somatomedin production, particularly during food restriction and fetal life or hypothyroidism. In all these situations this phenomenon is associated with reduced T3 levels, suggesting a significant influence of thyroid function on GH action and body growth. Thyroid hormones also stimulate the production of IGF and consequently fetal growth and even body growth rate in dwarf animals is stimulated by T3 (Cabello and Wrutniak, 1989) (Figs. 1 and 2). In the growing chicken, thyroid hormones do play an important role in growth as indicated by the administration of these hormones, by thyroidectomy or by treatment with anti-thyroid drugs (King and May, 1984). There is also evidence for the growth promoting effect of GH, of which plasma levels are high in the young domestic fowl. An injection of an antiserum to avian GH depresses growth, which is restored by GH substitution. ¨ et al. / Comparative Biochemistry and Physiology Part B 132 (2002) 299–308 E.R. Kuhn 301 Fig. 2. Growth hormone and gene expression related to growth. Contrary to these observations, administrations of chicken GH (cGH) failed to increase growth rate in poultry (Vasilatos-Younken et al., 1988; Cogburn et al., 1997). This has been attributed to a failure of GH supplementation to raise further the already high plasma concentrations of T3, contrary to the late embryonic situation. It has been shown that after hatching low levels of the T3 inactivating enzyme D3 are responsible for the high levels of T3. Gene Expression of D3 is inhibited by GH, mainly at the level of D3 gene transcription. This effect of growth hormone on D3 expression per¨ et al., 1993; Darras et sists throughout life (Kuhn al., 1993, 1994). GH therefore controls both induction of IGF-I transcription (Rosselot et al., 1995) and inhibition of D3 transcription in the chicken. We would like to consider the down-regulation of D3 by GH as part of its biological function in the chicken (Fig. 2) There is evidence that thyroid hormones may be critical regulators for IGF-I in mammals and chicken (Cabello and Wrutniak, 1989). Also the thyroid status profoundly influences circulating IGF-I bioactivity and local IGF-I production in a tissuespecific manner in the rat (Thomas et al., 1993), and in the chicken, circulating IGF-I concentra- tions were reported to decrease in chickens administered supplemental T3 (Vasilatos-Younken et al., 1997; Tixier-Boichard et al., 1990), whereas hypothyroid chickens have a lower growth rate and a lower IGF-I production (Decuypere et al., 1987). 3. Ontogeny of growth promoting hormones and the GH-receptor As in mammals during fetal intra-uterine life, the chicken IGF-I secretory pattern seems not to depend on GH secretion during embryogenesis. Plasma concentrations of GH are first detectable on day 17 of incubation, then rise gradually with larger increases occurring during the week following hatching (Harvey et al., 1979). However, no effect of GH releasing hormones on IGF-I concentration was observed during incubation (Huybrechts et al., 1985), whereas plasma T3 ¨ concentrations are increased (Kuhn et al., 1990). The hepatic cGH receptor (cGHR) gene expression is detectable at early ages in the embryonic chicken. Expression of the cGHR is transient, with a sharp decline at hatching (Burnside and Cogburn, 1992). The variations of mRNA in chicken embryos are consistent with reported data on GH binding Table 1 Correlation analysis of plasma GH concentrations and GH binding to liver microsomal fractions of saline- and GH-injected embryos (18-day-old) over a period of 4 h following injection Total receptors Free receptors Spearman correlation coefficient Probability ))R) Spearman correlation coefficient Probability ))R) Salineinjected GHinjected 0.16 Ps0.50 0.24 Ps0.31 y0.85 P-0.0001 y0.81 P-0.0001 302 ¨ et al. / Comparative Biochemistry and Physiology Part B 132 (2002) 299–308 E.R. Kuhn Fig. 3. Ontogeny of GH, IGF-I, T3, GH binding and GHR expression (mRNA) in the chicken; H, hatching. (Vanderpooten et al., 1991a,b). The sharp rise of plasma GH, starting immediately before hatching, seems to down-regulate GHR expression in late embryonic and early post-hatch life (see Fig. 3). The post-hatch cGHR expression gradually increases. From week 2 post-hatch onwards, hormone secretion and receptor expression are mature and GH-dependent growth can occur. Later, GH secretion declines, whereas hepatic binding to the cGHR and cGHR mRNA levels progressively increase with age (Scanes and Harvey, 1981; Vanderpooten et al., 1991a,b; Burnside and Cogburn, 1992; Mao et al., 1997). Throughout development and growth, circulating GH concentrations are inversely related to hepatic GHR expression, both at the protein and mRNA level (Burnside and Cogburn, 1992). Although the exact developmental factor regulating GHR synthesis is not known, it is tempting to speculate that GH down-regulates GHR expression, either directly or by IGF-I induction. 4. Receptor mediated endocytosis in the chicken 4.1. GH and GH-binding In mammals, the processes following GH binding to the GHR have been studied in detail. Subsequent to GH binding to the plasma membrane-situated GHR, the GH–GHR complex is internalized to an acid cell compartment where degradation occurs (Barrazone et al., 1980; Murphy and Lazarus, 1984; Roupas and Herington, 1988). In chicken, several observations point to the same mechanism. a. Liver of growing chicks contains very little GH receptors, though fast growth at this age is paralleled by high plasma GH concentrations (Vanderpooten et al., 1991a,b). b. Adult laying hens which fasted for three days exhibit higher circulating GH levels but lower GH binding to liver membranes (Vanderpooten et al., 1989). c. Sex-linked dwarf chickens show low or no hepatic GH binding but circulating GH is increased (Scanes et al., 1983; Leung et al., ¨ 1987; Huybrechts et al., 1987; Kuhn et al., 1989; Vanderpooten et al., 1991b). d. Following hypophysectomy plasma GH drops below the detection limit but hepatic GHR are increased. Moreover, the GHR increase by hypophysectomy is normalized by GH supplementation (Vanderpooten et al., 1991a). e. And finally, the episodic release of GH secretion in young chickens, resulting in pulsatile GH concentrations, does not cause extensive GHR down-regulation (Johnson, 1988, 1989). Moreover, administration of GH following this pattern, increases growth, whereas an injection or infusion of GH does not (Vasilatos-Younken et al., 1988; Cogburn et al., 1997). Therefore, it was suggested that continuously high circulating concentrations of GH down-regulate liver GHR number (Vanderpooten et al., ¨ et al. / Comparative Biochemistry and Physiology Part B 132 (2002) 299–308 E.R. Kuhn 1991a), and that the cell replaces internalized GH receptors, either by de novo synthesis andyor by recycling of GH receptors to the plasma membrane (Fig. 3). This conclusion is supported by a recent experiment, where 18-day-old embryos and 15-day-old chicks were injected with 10 and 12.5 mg cGH, respectively, and both GH plasma concentrations and binding to liver microsomal fractions were measured. A negative GH–GHR correlation coefficient is the main result, especially in embryos where the GH-dependent decrease over a period of 4 h in GH binding is striking (Vleurick, 1999) (Table 1). As this study was performed using microsomal fractions that contain membrane vesicles derived from miscellaneous cell compartments (Amar-Costesec et al., 1974), intracellular shifts of GHR remained unnoticed. Therefore, a study was planned to isolate subcellular fractions from livers of saline- and GH-injected birds. 4.2. Intracellular distribution of GHRs In mammals, the intracellular distribution of the GHR was examined and it was found that, depending on their location in the cell, variations in GHRs are differently related to changes in plasma GH (Hochberg et al., 1993). Binding of human GH to lactogenic receptors was demonstrated by electron microscope autoradiography in Golgi apparatus fractions prepared from rat liver (Bergeron et al., 1978). Later, also somatogenic binding sites were detected in Golgi fractions (Husman et al., 1985). Immediately following a spontaneous plasma GH pulse, somatogenic and lactogenic GH binding to rat hepatic cell membrane fractions declines, whereas in Golgi apparatus fractions binding increases (Bick et al., 1989a,b). Previous binding studies on chicken liver made use of microsomal fractions or heterogeneous membrane fractions prepared by differential centrifugation of a homogenate. Microsomes are membrane vesicles of diverse subcellular origin. Endoplasmic reticulum fragments are well represented, next to lysosomes, Golgi apparatus and plasma membrane derived vesicles (Amar-Costesec et al., 1974). Since all these membranes may contain GHRs, procedures to obtain highly purified plasma membrane and Golgi apparatus fractions from one liver are desirable in order to get a better 303 insight in internalization, degradation and recycling of GHRs in chicken liver. Chicken liver plasma membranes minimally contaminated with Golgi apparatus-derived vesicles, were prepared and identified using two marker enzymes, alkaline phosphatase (AP) and alkaline phosphodiesterase (APD) (adapted from Beaufay et al., 1974). Golgi apparatus fractions were identified by galactosyltransferase (GT) as a marker (Van Veldhoven and Mannaerts, 1991). From the analysis of the fractions obtained by differential centrifugation and from the discontinuous gradients, it is clear that in general GH binding co-migrated with GT activity and not with AP and APD activities. Therefore, the majority of GH binding seems to reside in the Golgi apparatus (Vleurick et al., 1999a; Fig. 4). Also in rat, GHR number in plasma membrane constitutes only a fraction of those in the Golgi apparatus (Posner et al., 1979). As removal of endogenous GH was difficult, our results do not totally exclude the presence of occupied receptors in the plasma membrane. However, it is known that GHR occupancy in adult chicken liver is very low due to their low plasma GH levels (Vanderpooten et al., 1989). Although GHR concentration in the plasma membrane is surely low, the absence of the GHR from the plasma membrane is unlikely. It is generally accepted that GH receptors in the plasma membrane are indispensable for transduction of GH signals (Sotiropoulos et al., 1994). Moreover, in earlier studies conducted in our laboratory it was found that cGH is active in adult laying hens. Within 2 h after injections of cGH plasma T4 levels are decreased and T3 levels increased through a reduced hepatic breakdown of T3 (Darras et al., 1992). To provoke these GH-dependent changes, only a minimal GHR number is required if after binding the signal is multiplied in the secondary messenger cascade. Signal transduction by the cGHR involved the same molecules that were identified for the mammalian GHR (Vleurick et al., 1999c, see below) The GH-binding study indicated that the high GHR content of microsomal fractions prepared from adult hen liver is largely due to its Golgi localization. Since for GH sensitivity only receptors at the cell surface are relevant, the question rises whether the intracellular GHR has a physiological role per se or is merely a consequence of excess synthesis. 304 ¨ et al. / Comparative Biochemistry and Physiology Part B 132 (2002) 299–308 E.R. Kuhn It has been reported that transcription of the GHR gene increases with age (Burnside and Cogburn, 1992) (Fig. 3). A large intracellular stock is built up if constitutive GHR synthesis goes along with low GHR degradation. When circulating GH levels are low, as is the case in adult chickens, binding and degradation of internalized receptors are reduced. In general, the presence of intracellular receptors ensures quick reversal of transient GH refractoriness as result of receptor internalization, since at least in mammals, GH receptors can be transferred from the intracellular Golgi stock to the plasma membrane (Hochberg et al., 1993). 4.3. Endocytosis of GH–GHR complex following GH injections Next to the receptor synthesis pathway, the presence of intracellular receptors has also been connected to endocytosis of hormone–receptor complexes. When radioactive bovine GH is injected in rats, label appears subsequently in hepatic plasma membrane, endocytic compartment and lysosomes (Husman et al., 1988). The endosomal fractions in the latter study were 40–50-fold enriched in the Golgi marker GT. Other authors also reported on the presence of GH binding in the Golgi apparatus due to internalization. Immediately following a spontaneous plasma GH pulse, somatogenic and lactogenic GH binding to rat hepatic cell membrane fractions declines, whereas in Golgi fractions binding increases (Bick et al., 1989b). Following binding to GH receptors located in the plasma membrane, GH–GHR complexes enter the cell by endocytosis, ligand and receptor dissociate and internalized receptors are either degraded or recycled to the plasma membrane, presumably through the Golgi apparatus (Hochberg et al., 1993). In the following figure, the internalization model proposed by these researchers is represented (Fig. 5). In the adult chicken, 25 mgykg of cGH was injected intravenously and total receptor binding was measured 10 min and 1 h afterwards. A positive correlation was found between plasma GH and total GHR binding of the Golgi fraction in the GH-injected but not in the saline group. No changes in free (unoccupied) receptors were observed. These experiments therefore demonstrate that in the chicken, as in its mammalian counterpart, the GHR is internalized following GH binding (Vleurick et al., 1998). 4.4. In vitro study of GH–GHR interaction Association of GH to its receptor leads to the subsequent association of a second GHR molecule (dimerization) (Cunningham et al., 1991). The conserved regions in the intracellular domain intervene in receptor-mediated internalization (Allevato et al., 1995; Vleurick et al., 1999c) and in signal transduction by Janus kinase-2 (JAK-2) and signal transducers and activators of transcription (STAT proteins) (Finidori and Kelly, 1995; Vleurick et al., 1999c). However, in addition to the JAKySTAT secondary messenger pathway, a direct role in gene transcription was also postulated for GH, GHR and binding protein for GH (GHBP) in view of their association with chromatin, subsequently to nuclear translocation through the endosomal pathway (Lobie et al., 1991, 1992, 1994a,b). In the chicken intracellular trafficking of GH and its receptor was examined in COS-7 cells transfected with the chicken GHR and detected using immunofluorescence studies. In the absence of GH, GHR immunofluorescence is found in the perinuclear region and on the plasma membrane. Internalization of both hormone and receptor is induced by addition of GH after 20 min. Partial colocalization of GH and GHR is found in endocytic and later, in large lysosomal vesicles. Neither receptor nor ligand is transferred to the nucleus, as confirmed by confocal laser microscopy (Vleu- Fig. 4. Comparison of enrichment in marker enzyme and GH receptors (GHR) in purified plasma membranes and Golgi fractions. Values below one indicate enrichment of GHR content. AP, alkaline phosphatase; APD, alkaline phosphodiesterase; and GT, galactosyltransferase (Vleurick et al., 1999c). ¨ et al. / Comparative Biochemistry and Physiology Part B 132 (2002) 299–308 E.R. Kuhn Fig. 5. Schematic model of the spontaneous changes of the hGH receptors and their internalization to Golgi membranes (modified after Hochberg et al., 1993). rick et al., 1998). Moreover, large amounts of cGHBP were detected in the culture medium and cell lysate of these cGHR-transfected cells, indicating that the internalization process may also serve as the production site for GHBP, following proteolytic cleavage of the GHR (Vleurick et al., 1999b,c). In summary, a model based on literature data and the present results, resumes all the processes involved in GH–cGHR interaction (Figs. 4–6). GH-induced receptor dimerization promotes association and activation of JAK2 with the receptor. The activated JAK2 is autophosphorylated and phosphorylates the intracellular domain of the GHR at multiple sites. The STAT proteins are then recruited to the activated JAKyGHR complex and 305 become substrates for phosphorylation by JAK2. Phosphorylated STAT proteins dimerize and dissociate from the receptor, and are translocated to the nucleus where they bind consensus response elements and affect expression of target genes. In addition, adapter protein 2 (AP-2) is recruited from the cytosol to the membrane by Tyr-based internalization signals in the receptor’s intracellular domain presumably with the help of a membranebound high affinity AP-2 docking protein (not shown). Membrane-bound AP-2 complexes then initiate clathrin coat assembly, a highly coordinated process resulting in the entrapment and concentration of GH–GHR complexes. Membrane invagination presumably arises from rearrangements within the clathrin lattice. Finally, dynamin is required to pinch off the clathrin-coated vesicle. An uncharacterized factor (X) interacts with the identified b-turn, enhancing internalization of GH by mammalian GHR as compared to the cGHR. Clathrin is recycled from the coated vesicles. In endocytic vesicles, the slightly acid pH is sufficient to dissociate hormone and receptor, enabling separate processing of GH and GHR. Endocytic vesicles are either directed to a degrading or a recycling pathway. However, the extensive colocalization of GH and GHR suggests that hormone and receptor are processed in the same cell compartments. Endocytic vesicles may fuse with lysosomes leading to the degrada- Fig. 6. Schematic representation of the GH–GHR interaction and all subsequent events (Vleurick, 1999). 306 ¨ et al. / Comparative Biochemistry and Physiology Part B 132 (2002) 299–308 E.R. Kuhn tion of their content. GHBP, the extracellular domain of the GHR, may be generated in the lysosome by proteolysis of the full-length GHR. Recycling vesicles return directly to the plasma membrane or are transferred transiently to the perinuclear region, where the Golgi apparatus is located. In addition, to the recycled receptors, the Golgi apparatus contains large amounts of newly synthesized GH receptors, due to its role in protein processing (Vleurick, 1999; Vleurick et al., 1999b,c). References Allevato, G., Billestrup, N., Goujon, L., et al., 1995. Identification of phenylalanine 346 in the rat growth hormone receptor as being critical for ligand-mediated internalization and down-regulation. J. Biol. Chem. 270, 17210–17214. Amar-Costesec, A., Beaufay, H., Wibo, M., et al., 1974. Analytical study of microsomes and isolated subcellular membranes from rat liver. II Preparation and composition of the microsomal fraction. J. Cell. Biol. 61, 201–212. Barrazone, P., Lesniak, M.A., Gorden, P., Van Obberghen, E., Carpentier, J.-L., Orci, L., 1980. Binding, internalization, and lysosomal association of 125 human growth hormone in cultured human lymphocytes: a quantitative morphological and biochemical study. J. Cell. Biol. 87, 360–369. Beaufay, H., Amar-Costesec, A., Feytmans, F., et al., 1974. Analytical study of microsomes and isolated subcellular membranes from rat liver. Biochemical methods. J. Cell. Biol. 61, 188–200. Bergeron, J.J.M., Posner, B.I., Josefsberg, Z., Sikstrom, R., 1978. Intracellular polypeptide hormone receptors. The demonstration of specific binding sites for insulin and human growth hormone in Golgi fractions isolated from the liver of female rats. J. Biol. Chem. 253, 4058–4066. Bick, T., Youdim, M.B.H., Hochberg, Z., 1989a. Adaptation of liver membrane somatogenic and lactogenic growth hormone (GH) binding to the spontaneous pulsation of GH secretion in the male rat. Endocrinology 125, 1711–1717. Bick, T., Youdim, M.B.H., Hochberg, Z., 1989b. The dynamics of somatogenic and lactogenic growth hormone binding: internalization to Golgi fractions in the male rat. Endocrinology 125, 1718–1722. ¨ Brown, M.R., Meacham, L.R., Pfaffle, R.W., et al., 1993. The molecular defect in Laron syndrome: exon deletions. In: Laron, Z., Parks, J.S. (Eds.), Lessons from Laron Syndrome (LS) 1966–1992. A model of GH and IGF–I action and ¨ interaction. Karger, Basel, pp. 127–131. Burnside, J., Liou, S.S., Cogburn, L.A., 1991. Molecular cloning of the chicken growth hormone receptor complementary deoxyribonucleic acid: mutation of the gene in sexlinked dwarf chickens. Endocrinol 128, 3183–3192. Burnside, J., Cogburn, L.A., 1992. Developmental expression of hepatic growth hormone receptor and insulin-like growth factor-I mRNA in the chicken. Mol. Cell. Endcrinol. 89, 91–96. Cabello, G., Wrutniak, C., 1989. Thyroid hormone and growth: relationships with growth hormone effects and regulation. Reprod. Nutr. Dev. 29, 387–402. Cogburn, L.A., Mao, J.N.C., Burnside, J., 1997. The growth hormone receptor in growth and development. In: Harvey, S., Etches, S. (Eds.), Perspectives in Avian Endocrinology. J. Endocrinology Ltd, Bristol, UK, pp. 101–117. Cunningham, B.C., Ultsch, M., de Vos, A.M., Mulkerrin, M.G., Clauser, K.R., Wells, J.A., 1991. Dimerization of the extracellular domain of the human growth hormone receptor by a single hormone molecule. Science 254, 821–825. ¨ Darras, V.M., Berghman, L.R., Vanderpooten, A., Kuhn, E.R., 1992. Growth hormone acutely decreases type III iodothyronine deiodinase in chicken liver. FEBS. Lett. 310, 5–8. Darras, V.M., Rudas, P., Visser, T.J., et al., 1993. Endogenous growth hormone controls high plasma levels of 3,39,5triiodothyronine (T3) in growing chickens by decreasing the T3-degrading type III deiodinase activity. Dom. Anim. Endocrinol. 10, 55–65. ¨ Darras, V.M., Kuhn, E.R., Decuypere, E., 1994. Comparative aspects of maturation and control of thyroid hormone deiodination during development. Isr. J. Zool. 40, 383–400. Decuypere, E., Buyse, J., Scanes, C.G., Huybrechts, L.M., ¨ Kuhn, E.R., 1987. Effects of hyper- or hypothyroid status on growth, adiposity and levels of growth hormone, somatomedin C and thyroid metabolism in broiler chickens. Reprod. Nutr. Develop. 27, 555–565. ¨ Decuypere, E., Huybrechts, L.M., Kuhn, E.R., Tixier-Boichard, ´ M., Merat, P., 1991. Physiological alterations associated with the chicken sex-linked dwarfing gene. Crit. Rev. Poultry. Biol. 3, 191–221. Finidori, J., Kelly, P.A., 1995. Cytokine receptor signaling through two novel families of transducer molecules: Janus kinases, and signal transducers and activators of transcription. J. Endocrinol. 147, 11–23. Harvey, S., Davison, T.F., Chadwick, A., 1979. Ontogeny of growth hormone and prolactin secretion in the domestic fowl (Gallus domesticus). Gen. Comp. Endocrinol. 39, 270–273. Harvey S., Lavelin I., Sanders ET., Pines M. Growth hormone and embryonic growth: which came first: the chicken or the egg. Abstract 6.01, VII International Symposium on Avian Endocrinology. Banares Hindu University, Varanasi, India, 2000. Hochberg, Z., Phillip, M., Youdim, M.B.H., Amit, T., 1993. Regulation of the growth hormone (GH) receptor and GHbinding protein by GH pulsatility. Metabolism 142, 1617–1623. Husman, B., Andersson, G., Norstedt, G., Gustafsson, J.-A., 1985. Characterization and subcellular distribution of the somatogenic receptor in rat liver. Endocrinology 116, 2605–2611. Husman, B., Gustafsson, J.-A., Andersson, G., 1988. Receptormediated endoytosis and degradation of bovine growth hormone in rat liver. Mol. Cell. Endocrinol. 59, 13–25. Huybrechts, L.M., Decuypere, E., Scanes, C.G., Callewaert, ¨ P., Peys, E., Kuhn, E.R., 1985. Human pancreatic growth hormone releasing factor stimulates growth hormone secretion in perinatal dwarf and control chickens. Hormone Metab. Res. 17, 690–691. ´ ¨ Huybrechts, L.M., Kuhn, E.R., Decuypere, E., Merat, P., Scanes, C.G., 1987. Plasma concentrations of growth hor- ¨ et al. / Comparative Biochemistry and Physiology Part B 132 (2002) 299–308 E.R. Kuhn mone and somatomedin C in dwarf and normal chickens. Reprod. Nutr. Dev. 27, 547–553. Johnson, R.J., 1988. Diminution of pulsatile growth hormone secretion in the domestic fowl (Gallus domesticus): evidence for sexual dimorphism. J. Endocrinol. 119, 101–109. Johnson, R.J., 1989. Growth physiology and biotechnology: potential to improve broiler production. World Poultry Sci. J. 46, 228–240. King, D.B., May, J.D., 1984. Thyroidal influence on body growth. J. Exp. Zool. 232, 453–460. ¨ Kuhn, E.R., Huybrechts, L.M., Vanderpooten, A., Berghman, L., 1989. A decreased capacity of hepatic growth hormone (GH) receptors and failure of thyrotrophin-releasing hormone to stimulate the peripheral conversion of thyroxine into tri-iodothyronine in sex-linked dwarf broiler hens. Reprod. Nutr. Dev. 29, 461–467. ¨ Kuhn, E.R., Huybrechts, L.M., Darras, V.M., Meeuwis, R., Decuypere, E., 1990. Impaired peripheral T3 production but normal induced thyroid hormone secretion in the sex-linked dwarf chick embryo. Reprod. Nutr. Dev. 30, 193–201. ¨ Kuhn, E.R., Bergman, L.R., Moons, L., Vandesande, F., Decuypere, E., Darras, V.M., 1993. Hypothalamic and peripheral control of thyroid function during the life cycle of the chicken. In: Sharp, P.J. (Ed.), Avian Endocrinology. Bristol; Journal of Endocrinology Ltd, pp. 29–46. Leung, F.C., Styles, W.J., Rosenblum, C.I., Lilburn, M.S., Marsh, J.A., 1987. Diminished hepatic growth hormone receptor binding in sex-linked dwarf broiler and Leghorn chickens. P. Soc. Exp. Biol. Med. 184, 234–238. Lobie, P.E., Barnard, R., Waters, M.J., 1991. The nuclear growth hormone receptor-binding protein. J. Biol. Chem. 266, 19947–19954. ´ J., Wang, B.S., Baumbach, W.R., Lobie, P.E., Garcia-Aragon, Waters, M.J., 1992. Cellular localization of the growth hormone binding protein in the rat. Endocrinology 130, 3057–3065. Lobie, P.E., Mertani, H., Morel, G., Morales-Bustos, O., Norstedt, G., Waters, M.J., 1994a. Receptor-mediated nuclear translocation of growth hormone. J. Biol. Chem. 269, 21330–21339. Lobie, P.E., Wood, T.J.J., Min Chen, C., Waters, M.J., Norstedt, G., 1994b. Nuclear translocation and anchorage of the growth hormone receptor. J. Biol. Chem. 269, 31735–31746. Mao, J.N.C., Cogburn, L.A., Burnside, J., 1997. Growth hormone down-regulates growth hormone receptor mRNA in chickens but developmental increases in growth hormone receptor occur independently of growth hormone action. Mol. Cell. Endocrinol. 129, 135–143. Murphy, L.J., Lazarus, L., 1984. The mouse fibroblast growth hormone receptor: ligand processing and receptor modulation and turnover. Endocrinology 115, 1625–1632. Phornphutkul, C., Frick, C.P., Goodman, H.H., Berry, S.A., Grappuso, P.A., 2000. Hepatic growth hormone signaling in the late gestation fetal rat. Endocrinology 141, 3527–3533. Posner, B.I., Josefsberg, Z., Bergeron, J.J.M., 1979. Intracellular polypeptide hormone receptors: characterization and induction of lactogen receptors in the Golgi apparatus of the rat. J. Biol. Chem. 254, 12494–12499. Rosselot, G., McMurtry, J.P., Vasilatos-Younken, R., Czerwinski, S., 1995. Effect of exogenous chicken growth hormone (CGH) administration on insulin-like growth factor-I (IGFI) gene expression in domestic fowl. Mol. Cell. Endocrinol. 114, 157–166. 307 Roupas, P., Herington, A., 1988. Intracellular processing of growth hormone receptors by adipocytes in primary culture. Mol. Cell. Endocrinol. 57, 93–99. Scanes, C.G., Harvey, S., 1981. Growth hormone and prolactin in avian species. Life Sci. 28, 2895–2902. Scanes, C.G., Marsh, J., Decuypere, E., Rudas, P., 1983. Abnormalities in the plasma concentrations of thyroxine, tri-iodothyronine and growth hormone in sex-linked dwarf and autosomal White Leghorn domestic fowl (Gallus domesticus). J. Endocrinol. 97, 127–135. Sotiropoulos, A., Perrot-Applanat, M., Dinerstein, H., et al., 1994. Distinct cytoplasmatic regions of the growth hormone receptor are required for activation of JAK2, mitogenactivated protein kinase, and transcription. Endocrinology 135, 1292–1298. Thomas, H.R., Miell, J.P., Taylor, A.M., 1993. Endocrine and cardiac paracrine actions of insulin-like growth factor-I (IGF-I) during thyroid dysfunction in the rat: is IGF-I implicated in the mechanism of heart weightybody weight change during abnormal thyroid function? J. Mol. Endocrinol. 10, 313–323. ¨ Tixier-Boichard, M., Decuypere, E., Huybrechts, L., Kuhn, E., ´ Merat, M., 1990. Effects of dietary T3 on growth parameters and hormone levels in normal and sex-linked dwarf chickens’. Domest. Anim. Endocrinol. 7, 753–786. ¨ Vanderpooten, A., Dewil, E., Decuypere, E., Kuhn, E.R., 1989. The effect of fasting on growth hormone receptor in liver membranes of adult chicken. Med. Sci. Res. 17, 1031–1032. Vanderpooten, A., Darras, V.M., Huybrechts, L.M., Rudas, P., ¨ Decuypere, E., Kuhn, E.R., 1991a. Effect of hypophysectomy and acute administration of growth hormone (GH) on GH-receptor binding in chick liver membranes. J. Endocrinol. 129, 275–281. ¨ Vanderpooten, A., Huybrechts, L.M., Decuypere, E., Kuhn, E.R., 1991. Differences in hepatic growth hormone receptor binding during development of normal and dwarf chickens. Reprod. Nutr. Dev. 31, 47–55. Van Veldhoven, P.P., Mannaerts, G.P., 1991. Subcellular localization and membrane topology of sphingosine y,y phosphate lyase in rat liver. J. Biol. Chem. 266, 12502–12507. Vasilatos-Younken, R., Cravener, T.L., Cogburn, L.A., Mast, M.G., Wellenreiter, R.H., 1988. Effect of pattern of administration on the response of exogenous, pituitary-derived chicken growth hormone by broiler-strain pullets. Gen. Comp. Endocrinol. 71, 268–283. Vasilatos-Younken, R., Dunnington, E.A., Siegel, P.B., McMurtry, J.P., 1997. Tissue-specific alterations in insulin-like growth factor-I concentrations in response to 3,39,59-triiodoL-thyronine supplementation in the growth hormone-receptor deficient sex-linked dwarf chicken. Gen. Comp. Endocrinol. 105, 31–39. ¨ Vleurick, L., Van Veldhoven, P., Decuypere, E., Kuhn, E.R., 1998. Intracellular growth hormone receptors in chicken liver. In: Vaudry, H., Tonon, M.-C., Roubos, E.W., De Loof, A. (Eds.), Annals of the New York Academy of Sciences, vol. 839. Trends in Comparative Endocrinology and Neurobiology. Eighteenth Conference of European Comparative Endocrinologists: From Molecular to Integrative Biology. pp. 538–540. 308 ¨ et al. / Comparative Biochemistry and Physiology Part B 132 (2002) 299–308 E.R. Kuhn Vleurick, L., 1999. Interaction of growth hormone and growth hormone receptor in chicken (Gallus domesticus) liver. Dissertationes de Agricultura 387, 1–118. Vleurick, L., Kuhn, E.R., Decuypere, E., Van Veldhoven, P.P., ¨ 1999a. Isolation of plasma membranes and Golgi apparatus from a single chicken liver homogenate. J. Cell. Biochem. 72, 349–355. ¨ Vleurick, L., Kuhn, E.R., Decuypere, E., Burnside, J., Pezet, A., Edery, M., 1999b. Generation of chicken growth hormone-binding proteins by proteolysis. Gen. Comp. Endocrinol. 113, 283–289. ¨ Vleurick, L., Pezet, A., Kuhn, E.R., Decuypere, E., Edery, M., 1999c. A b-turn endocytic code is required for optimal internalization of the growth hormone receptor but not for a-adaptin association. Mol. Endocrinol. 13, 1823–1831.