Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
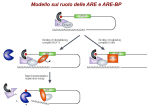
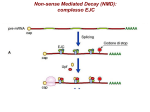
Messenger RNA maturation Dr.ssa Mariangela Morlando Features of RNA molecule Synthetised from a DNA template (transcription) Single strand molecule Contains ribose in place of deoxyribose Contains Uracil in place of Thymine Types of RNA molecules Coding RNA which are translted into proteins: messenger RNA(mRNA) Non coding RNA which are never translated into protein but work as RNA with structural and/or regulatory function. Messenger RNA mRNA structures AUG UAA cap AAAAAAAAAAAAA 3’ UTR 5’ UTR ORF Eucaryotic gene expression transcription START site exons gene upstream elements promoter elements TRANSCRIPTION CAPPING introns pre-mRNA m7 G SPLICING POLYADENYLATION m7 G m7 G AAAAAAAAAn AAAAAAAAAn TRANSPORT TRANSLATION polyadenylated pre-mRNA mature mRNA Eucaryotic gene expression transcription START site exons gene upstream elements promoter elements TRANSCRIPTION CAPPING introns pre-mRNA m7 G SPLICING POLYADENYLATION m7 G m7 G AAAAAAAAAn AAAAAAAAAn TRANSPORT TRANSLATION polyadenylated pre-mRNA mature mRNA Why the Cap structure is important? 1) RNA stability 2) Favours the mRNA transport to the cytoplasm 3) Increases translation (it binds to eIF4E that belongs to translation initiation complex) 7-metilguanosina This involves the addition of a modified guanine base to the 50 end of the RNA. Specifically, it is a methylated guanine, and it is joined to the RNA transcript by an unusual 5‘–5’ linkage involving three phosphates •CAP is added at very early stage of transcription initiation estremità 5’ •The 5’-5’ phosphodiester bond makes della RNA catena the molecule resistant to the exonuclease activity. •I vitro synthetized RNA without CAP are rapidly degraded pre-mRNA capping The 5’ cap is created in three enzymatic steps: 1. 1. 1. a phosphate group is removed from the 5’ end of the transcript. GMP moiety is added. GMP nucleotide is modified by the addition of a methyl group. RNA triphosphatase RNA guanylyltransferase RNA(G-7-) methyltransferase 5’ CAP favours the mRNA transport to the cytoplasm CBC: CAP binding complex Phosphorylation state of the CTD of RNA polymerase II along transcription cycle DSIF/NELF-mediated checkpoint to ensure pre-mRNA capping. TFIIH phosphorylate s the RNAP II CTD on Ser 5. DSIF interacts with RNAP II shortly after initiation. NELF recognizes the RNAP II–DSIF complex and halts elongation. This pause allows the recruitment of the capping enzyme (CE) which adds a 5′-cap to the nascent transcript NELF is released by the concerted action of P-TEFb phosphorylation of DSIF and the CTD on Ser 2, PRMT1/5 methylation of Spt5, and the capping enzyme itself. FCP1 may also participate, as FCP1 is required to release the capping enzyme. The precise mechanism causing NELF release is unknown. Robert J. Sims III et al. Genes Dev. 2004;18:2437-2468 Eucaryotic gene expression transcription START site exons gene upstream elements promoter elements TRANSCRIPTION CAPPING introns pre-mRNA m7 G SPLICING POLYADENYLATION m7 G m7 G AAAAAAAAAn AAAAAAAAAn TRANSPORT TRANSLATION polyadenylated pre-mRNA mature mRNA What is the function of the polyA tail? 1) RNA stability 2) Favours the mRNA transport to the cytoplasm 3) Increases translation efficiency by favouting the loading of ribosomal 40S subunit 4) mRNA 3’ end formation allows efficient transcription termination. Looking for consenus sequences SEQUENCE ALLINEAMENT OF cDNAs STARTING FROM THE POLYA TAIL 15-20 nt 5’UTR globin Ig Ovoalbumin Polymerase Myc 3’UTR AUG AUG UAA GGUUAUCCAUCAAUAAA…….GCUAUACGCAAAA n UAA CCACUGGGCCAAUAAA…….GCUAUACGCAAAA n AUG UAA GCAACCUCGAAUAAA…….GCUAUACGCAAAA n AUG UAA AUCUGGAGGAAUAAA…….GCUAUACGCAAAA n AUG UAA UAUAGAUCCAAUAAA…….GCUAUACGCAAAA n AAUAAA consensus consensus Polyadenylation efficiency Influence of consensus sequence AAUAAA Efficiency Site-specific mutation Recombinant DNA preparation mutazione * transfection mut1 mut2 mut3 WT polyA HeLa cells Northern blot RNA extraction After 24 h 3’ end formation in mammalian cells Cleavage and polyadenylation site 5’ AAUAAA GU rich 3’ 15-20 nt 5’ AAUAAA OH P GU rich degradation 5’ AAUAAA A250 3’ SPECIFIC COMPLEXES ARE INVOLVED IN mRNA 3’END PROCESSING Cleavage and Polyadenylation Specific Factor sito di taglio CPSF 5’ AAUAAA CA CFII Cleavage Stimulation Factor CstF 3 ’ PAP GU-rich CFI Poly-A polymerase Curr Opin Cell Biol. 2004 Jun;16(3):272-8 band shift/EMSA assay: to study RNAprotein interaction in vitro Labelled RNA In vitro transcription 1 Labelled RNA 2 Purified protein band shift/EMSA assay: to study RNAprotein interaction in vitro 2 1 Labelled RNA Purified protein RNA-protein complex Free RNA SPECIFIC COMPLEXES ARE INVOLVED IN mRNA 3’END PROCESSING Cleavage and Polyadenylation Specific Factor sito di taglio CPSF 5’ AAUAAA CFII Cleavage Stimulation Factor CstF 3 ’ PAP EMSA assay for testing the interaction between CPSF 160 and consensus sequence AAUAAA CA GU-rich CFI AAUAAA Poly-A polymerase AAGAAA HeLa NE Curr Opin Cell Biol. 2004 Jun;16(3):272-8 complex sito di taglio free RNA 3’-End Formation: RNA Processing Cleavage and Polyadenylation Specific Factor Cleavage Factors Cleavage Stimulation Factor 25 26 degradation 27 28 29 CAP and polyA tail influence efficient translation 3’ end formation occurs during transcritpion Polyadenylation is linked to termination Torpedo Model mRNA AAAAAAAAA pre-mRNA CPSF pA Xrn2 CstF RNAPII RNAPII Polyadenylation is linked to termination Torpedo model Allosteric model Variations at the transcriptome 3′ end—when processing gets alternative Approximately up to 70 % of the transcriptome is affected by a mechanism widely referred to as “alternative 3′ end cleavage and polyadenylation” (APA) APA can be regulated on the level of mRNA 3′ end processing by various cis- and trans-acting determinants: (1)the intrinsic strength of sequence elements, (2) the concentration or activity of polyadenylation factors, and/or (3) tissue- or stage-specific regulatory factors are the most important key players APA is highly regulated during development Proliferating cells tend to use upstream (“proximal”) PASs and produce mRNAs with shorter 3′UTRs, while quiescent/differentiated cells favor downstream (“distal”) PASs and produce mRNAs with longer 3′UTRs In contrast during somatic reprogramming or tumorigenesis, proximal PASs are favored leading to shorter 3′UTRs APA transcript isoforms of the same gene can encode different proteins and/or change the 3′UTR properties, leading to: - the inclusion or exclusion of mRNA stabilizing or destabilizing elements, - miRNA target sites, - result in different translation efficiencies - result in different subcellular localization Alternative polyadenylation of the immunoglobulin μ heavy chain gene sub obtimal pA Secreted terminus Plasma cells obtimal pA Membrane bound terminus B cells Secreted terminus Membrane bound terminus CSTF-64 levels control the alternative processing of mRNA. In not activated B cells the limiting concentration of CSTF allows the recognition of the stronger polyadenylation signal. The immuglobulin produced will then contain a portion for binding to the membrane. After the activation of B cells CSTF levels are increased and this allows the use of the weaker polyadenylation site that will be preferentially used because it will be the first to be transcribed. CstF-64 The polyadenylation factor CstF-64 regulates alternative processing of IgM heavy chain premRNA during B cell differentiation. Takagaki Y, Seipelt RL, Peterson ML, Manley JL. Cell 1996 Nov 29;87(5):941-52 U1A: an example of on-off regulation of the cleavage and polyadenylation step in RNA processing U1A is a protein that binds to U1snRNA Obtimal U1A binding site U1A is able to bind to its mRNA The mRNA encoding for U1A protein has a suboptimal polyA site (AUUAAA) and 2 sequences similar to the U1A binding site on U1snRNA U1A EMSA assay with increasing amount of U1A protein revealed that U1A binds its own mRNA and that this latter possessed two U1A binding sites RNA free - U1A autoregulation When the level of U1A exceeds that of U1 snRNA, the excess U1A protein binds to its two binding sites in U1A pre-mRNAs. Since U1A binding does not prevent cleavage of the pre-mRNAs, free 3′ ends are generated in the normal fashion In the absence of polyadenylation, however, both cleavage products are rapidly degraded by an exonuclease, so no functional U1A mRNA is produced. As a result, synthesis of U1A protein is decreased until all the excess is used in formation of new U1 snRNPs. Sub-obtimal U1A binding site sub-obtimal site influenza A by its connection to the cellular 3′ end apparatus has devised an efficient way to specifically shut off cellular gene expression The influenza A NS1 protein is one of the most abundant proteins synthesized in infected cells It interacts with the cellular 30 kDa subunit of CPSF. By sequestering CPSF30 it prevents the binding of the CPSF complex to the RNA substrate thus inhibits 3′ end cleavage and polyadenylation of the host pre-mRNAs by preventing NS1 also targets PABPN1, which inhibits the processive synthesis of long poly(A) tails catalyzed by PAP As mRNA processing represents a prerequisite for cytoplasmic export, the uncleaved host premRNAs are retained in the nucleus, while viral RNAs are still exported. APA in human disease - Loss‐of‐function mutations of globin mRNA 3′ end processing are a well‐recognized cause of thalassemias. Thalassemia patients have different mutations that result in an alteration of the AAUAAA hexanucleotide of the α‐globin and β‐globin genes, and invariably inactivate or severely inhibit gene expression. - Gain‐of‐function mutation stimulating 3′ end processing in the prothrombin (coagulation factor II; F2) gene. The G → A mutation converts the physiologically inefficient GC F2 cleavage site into the mechanistically most efficient CA dinucleotide, which increases cleavage site recognition and results in an enhancement of prothrombin mRNA and protein expression. The result in an increased risk to develop thrombosis (referred to as thrombophilia). - defective PAP causes cell‐cycle arrest in the G0–G1 phases, the activity of PAP thus likely reflects the proliferative activity of cells PAP mRNA is overexpressed in human carcinomas of the breast, colon, ovary and pancreas and polyadenylation activity is significantly enhanced in aggressive acute leukemias and Burkitt lymphoma compared with less aggressive chronic leukemias and normal lymphocytes - mRNAs with shorter 3′UTRs tend to be more stable or globally elevated eventually leading to higher protein output switching to shorter 3′UTRs may allow proto-oncogenes to escape from inhibition by miRNAs, thereby resulting in oncogene activation also in the absence of genetic alterations. Histone mRNA processing is polyadenylation independent Common factors in RNA processing pathways CPSF-73 endonucleolytic activity Poly(A)+ mRNAs Histone mRNAs CPSF-73 also displays 5′ → 3′ exoribonucleolytic activity that is responsible for the decay of the downstream cleavage product