Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
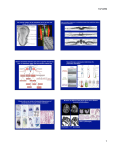
Building a bristle from scratch Flies have many types of sensory organs, which are the parts of the nervous system that take in information (such as light, touch, smell, sound) from the environment. After the eyes and antennae, the most visible sensory organs on the fly are the external sensory bristles that cover the fly. We will concentrate on the ones on the back of the fly, which are relatively easy to see. Anatomy of Drosophila bristles The back (or notum) has two classes of bristles – macrochaetes (the large ones) and microchaetes (the rows of small ones). The developmental processes that give rise to them are very similar, but the timing for when they appear varies. Figure 1. Fly bristles. On the left is an SEM image of a wild-type fly. In the upper right is a closeup of a single bristle (the smaller hairs are not part of the nervous system). In the lower right is a schematic of bristle structure. It is made up of four cells. All the large cell bodies are beneath the surface of the fly. The shaft (sh) and socket (so) cells make the visible bristle shaft and socket, respectively. The neuron (ne) and glial/sheath (gl) cells make no surface structures. The start of bristle development – the proneural cluster and the specification of the sensory organ precursor cell. Macrochaetes start developing during larval stages while microchaetes develop during pupal stages. Before this, all the tissue is simply epithelial tissue. Certain genes (which we will not discuss further) are expressed in a “prepattern” which defines regions that are capable of becoming part of the nervous system. These regions are clusters of many cells and are called proneural clusters (PNCs). Each cell in a PNC is capable of producing a bristle at this time point but ONLY ONE cell in each cluster eventually produces a bristle. How does this process occur? Figure 2: Establishment of the proneural cluster and selection of the SOP via Notch signaling. Each of the four squares in Figure 2 represents a region of cells that will give rise to a single bristle. Once the prepattern in laid down (yellow bands in first square), certain cells gain the potential to become part of the nervous system. These cells are colored yellow in the second square and constitute the PNC. What defines the PNC genetically is that ALL of the cells begin to express genes we call the proneural genes. One proneural gene that some of you will be using in your crosses is called scute (sc). Scute is expressed in all the cells of the proneural cluster early on but as the cells in the cluster talk to one another (via Notch signaling), the expression is eventually turned off in all but one cell (the red cell in the third and fourth squares). That cell becomes the sensory organ precursor (SOP) cell. Another gene that acts in concert with the proneural genes is senseless (sens). Both sc and sens mutants have similar phenotypes. Figure 3. A mutant fly that lacks the scute gene. The surface of the fly is devoid of all bristles. The bristle lineage – the divisions of the SOP and the specification of each cell. Once the SOP has been specified, it divides creating an “A” cell and a “B” cell. Each of these cells also divides, making the four cells of the bristle. The two daughters of the “A” cell become the socket and shaft cells. The two daughters of the “B” cell become the sheath/glial cell and neuron. (Point of information – there is one extra division that I’m not showing. The “B” cell divides twice. For the first division, one of its progeny goes on and dies by programmed cell death – it is a vestigial component of the sense organ) How do these cells decide who will adopt which fate? After each division, the two daughter cells communicate with each other via Notch signaling. The cell that receives the signal (the “A” cell, socket cell, and glial cell) are inhibited from adopting the other potential fate (“B” cell, shaft cell, neuron, respectively). For example, on the bottom left of Figure 3, the socket cell receives an inhibitory Notch signal from the shaft cell. In weak Notch mutants, you can get no Figure 4: The bristle lineage. The SOP is shown at the top and the cells that arise from it by division are below it. The T bars indicate an inhibitory signal through the Notch receptor from one cell to the other. sockets and two shafts. In stronger Notch mutants, each of the cell fate decisions is affected, so that after the first division, both cells become “B” cells and then after the second division, all four cells become neurons (Figure 5). Figure 5: Effects of loss of Notch function. On the left is a single developing bristle from a wild type fly stained for the neuron (red) and the sheath cell (green). The shaft and socket cells are also present but not visible with this preparation. On the right is a single developing bristle from a Notch mutant. All four cells are neurons (red). The green sheath cell has been transformed and the socket and shaft cells as well (if you looked at the surface of the fly, it would look bald). Members of the Notch pathway The genes involved in Notch signaling have been well defined and the signaling pathway is fairly well understood (Figure 6). The key elements include 1. Notch. This gene encodes a cell surface receptor on the cell that receives the signal (although it is also found on the sending cell). 2. Delta. This encodes the ligand (i.e. the cell surface molecule on the cell that sends the signal). When Delta on one cell binds to Notch on the other cell, the intracellular part of the Notch protein is enzymatically cleaved and moves to the nucleus. 3. Supressor of Hairless (Su(H)). Su(H) encodes a transcription factor. It binds to a DNA sequence and can turn genes on or off. When there is no Notch signal, it is bound by a corepressor and turns genes off. When there is a Notch signal, the intracellular part of Notch binds to Su(H) and changes it into an activator. Therefore, there are a certain class of genes that are responsive to a Notch signal. Without the signal, they are off (Su(H) is repressing them) but with the signal, they are on (the intracellular part of Notch with Su(H) is activating them). 4. Hairless. Hairless protein can bind to Su(H) and prevent it from interacting with the intracellular part of Notch. Therefore, Hairless acts against the Notch signal. Paradoxically, Hairless is found in all cells so how does one ever get Notch signaling? If there is a high level of Notch signaling, it overcomes the effects of Hairless. If there is a low level, then there is enough Hairless around to prevent the signal from working. This suggests that Hairless is acting to fine-tune the response to the signal and make sure cells don’t just respond immediately to a weak signal. Figure 6: How the Notch signaling pathway works. The Delta protein on the sending cell binds to the Notch protein on the receiving cell. The intracellular part of Notch is cleaved and travels to the nucleus. Su(H) is normally repressing genes (left side) via a co-repressor but the fragment of Notch that is cleaved is able to dislodge the corepressor and help the Su(H) become an activator (right side). Hairless (not shown) is normally helping to keep Su(H) from interacting with small amounts of Notch when the signal is not strong. Cells that do not receive (or are resistant to) the Notch signal adopt one fate (e.g. shaft cell) while cells that receive (or are sensitive to) the Notch signal adopt another fate (e.g. socket cell). Differentiation Once the cells decide who they are supposed to be, they must differentiate and become those cells. One gene that has been shown to be important in the differentiation of the shaft and sheath cell is Pax2. When Pax2 is mutated, all four cells of the sense organ are still present but the shaft cell does not produce a shaft and the sheath cell also does not differentiate properly (Figure 7). Figure 7: A Pax2 mutant. Note that although almost all the shafts are gone, the sockets are still present.