Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Speech perception wikipedia , lookup
Sound localization wikipedia , lookup
Hearing loss wikipedia , lookup
Lip reading wikipedia , lookup
Auditory processing disorder wikipedia , lookup
Audiology and hearing health professionals in developed and developing countries wikipedia , lookup
Noise-induced hearing loss wikipedia , lookup
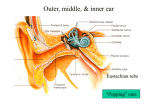
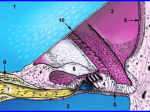
The physiology of hearing impairment 1 Young ED (2011) Neural coding of sound with cochlear damage. In C.G. Le Prell, D. Henderson, R.R. Fay, and A.N. Popper (Eds.) Noise Induced Hearing Loss: Scientific Advances. New York, Springer. (in press 2011). Moore BCJ (2007) Cochlear Hearing Loss. John Wiley and Sons. Liberman MC (1984) Single-neuron labeling and chronic cochlear pathology. I. Threshold shift and characteristic-frequency shift. Hearing Res 16:33-41. Liberman MC, Dodds LW (1984) Single-neuron labeling and chronic cochlear pathology. III. Stereocilia damage and alterations of threshold tuning curves. Hearing Res 16:55-74. Liberman MC, Kiang NY (1984) Single-neuron labeling and chronic cochlear pathology. IV. Stereocilia damage and alterations in rate- and phase-level functions. Hearing Res 16:75-90. Liberman MC, Dodds LW (1984) Single-neuron labeling and chronic cochlear pathology. II. Stereocilia damage and alterations of spontaneous discharge rates. Hearing Res 16:43-53. Moore BCJ (2004) Testing the concept of softness imperception: Loudness near threshold for hearingimpaired ears. J Acoust Soc Am 115:3103-3111. Moore BC et al. A test for the diagnosis of dead regions in the cochlea. Br J Audiol. 34:205 (2000). Hearing impairment can affect each of the steps in auditory transduction Conductive - damage (eardrum puncture), pathological processes (bone growth), or blockage of external and middle ear (not shown above). Metabolic - damage or degeneration of the stria vascularis (SV) or other components of cochlear homeostasis, indirectly affecting hair-cell transduction Sensorineural - damage to, degeneration of, or decreased gain of hair cells (IHC, OHC), degeneration of auditory nerve fibers (SGN, AN), or central auditory neurons and circuits. These lecture focuses on sensorineural hearing impairment (SNHL) and the functional consequences of damage to hair cells or auditory nerve fibers. SNHL has four well-studied causes: Acoustic trauma - exposure to very loud sounds for a long enough time to damage hair cells or auditory-nerve fibers. Ototoxic substances - some antibiotics (gentamycin), cancer drugs (cisplatin), and various other toxins specifically damage hair cells or other components of the cochlea. Genetic defects - any genetic defect that affects an essential functional element of cochlear physiology can cause deafness (e.g. Usher’s syndrome, caused by a defect in a myosin or in one of four other genes). Aging - which appears to operate indirectly via degeneration of the stria vascularis, leading to a reduction in the endolymphatic potential (EP) and a decrease in gain of hair cells. The perceptual deficits of SNHL: Loss of audibility Loudness recruitment Degraded frequency tuning Auditory neuropathy, degraded temporal precision Tinnitus To illustrate two of the primary behavioral effects of hearing impairment, look at equalloudness contours, along which tones of different frequencies sound equally loud. Equal-loudness contours Braida et al. Two problems: Threshold shift and recruitment. 2. Loudness growth is faster than normal (recruitment) 1. Soft sounds can't be heard Braida et al. The largest problem in hearing impairment is loss of audibility, sounds cannot be heard. The extent to which audibility alone can account for the degraded performance of hearing impaired listeners is evaluated with the articulation index. f1 AI = P ∫ I( f ) W ( f ) df f0 P - proficiency (fudge factor) I(f) - relative importance of different frequencies for understanding speech, empirically determined in normal listeners W(f) - fraction of the speech that is audible above noise masking or elevated threshold (roughly the red region at right) AI analysis shows that, with parameters P and I(f) set for normal listeners, the AI accurately predicts loss of performance in mild to moderately impaired listeners (<50 dB), but not in moderate-to-severe group (>55 dB). Apparently there are additional problems in such subjects. The difference is usually worse in older persons. normal thr. shift < 50 dB Note The task was recognition of spoken word lists with 9 different degrees of LP, HP, and BP filtering. thr. shift > 55 dB Pavlovic, 1984 The second problem, recruitment: loudness grows more rapidly than normal over some range, so that sounds of ≈80-100 dB are equally loud in a normal and an impaired listener. Note the narrowing of the dynamic range from the green to the red arrow. The behavior of impaired loudness near threshold is controversial. Some (Buus and Florentine, 2001) claim that loudness is elevated right at threshold, but the preponderance of evidence indicates that this is not so and that loudness growth is more like the blue curve (Moore, 2004). Recruitment is important in hearing-aid design, because the reduced dynamic range requires compressor circuits in aids. These circuits are problematic in that they are nonlinear and distort the sound. Buus and Florentine, 2001 The third problem: psychophysical auditory filters are wider in hearing-impaired ears. This means the ability to analyze sound by its frequency content is impaired. Note broadening (same arrow) Glasberg and Moore, 1986 The implications of broadened auditory filters: A vowel spectrum The same spectrum on the cochlea’s frequency scale (in units of auditory filter bandwidths) . . . the same with moderate hearing impairment (auditory filters 2x and 4x wider) The theoretical excitation pattern on the basilar membrane for this stimulus in a normal ear (the spectrum smoothed by auditory filters) Note the loss of spectral detail, e.g. the locations of the formant peaks, with decreased cochlear filtering. This is one likely explanation of below-AI performance in speech perception and suggests problems with background noise. Number of ERBs, E! Moore, 1995 The fourth problem: auditory neuropathy in which the thresholds are normal or show mild impairment, but subjects have considerable difficulty with complex perception, like speech. Thresholds of a person at two ages (19 and 32) show mild impairment (<60 dB) in both ears. However, the speech perception performance (% of words on an open set test) is much worse than expected from the hearing loss (green and orange symbols). The other data are from a population of similar subjects. Starr et al. 1996 Auditory neuropathy subjects show little or no ABR, meaning a lack of synchronous activity in the CNS. However, their DPOAEs (distortion product otoacoustic emissions) are normal, showing no deficit in outer hair cell function. Note inversion of waveform, this is CM OAE noise Starr et al. 1996 The physiological correlates of perceptual problems in hearing impairment. Inner and outer hair cells: outer hair cells amplify BM displacement; inner hair cells are the transducers for ANFs C. Smith Hair cell damage has been studied experimentally with acoustic trauma. These data show auditory nerve tuning and thresholds after such trauma (temporary threshold shift has been allowed to resolve). Note the 1. threshold shifts and 2. broadening of tuning when compared to normal tuning curves (gray). Substantial trauma was observed here in the absence of much hair cell loss. (The exposure was a 50 Hz noise band centered at 2 kHz, 115 dB for 2 hr, 54 day recovery) Liberman and Dodds, 1984 Another example of hair cell loss from acoustic trauma. In this case there is a dead region marked by the red band. At these frequencies, no auditory nerve fibers were found and there was substantial hair cell damage. normal threshold Dead regions have been described in psychophysical tests. Sophisticated masking methods can detect such regions (see Moore, 2007). Importantly, amplifying sounds at frequencies in a dead region does not help speech perception, even though it should increase audibility (from the AI). The reason is that sounds in a dead region are being heard through the spread of tuning curves from adjacent regions with existing IHCs and ANFs. ! No auditory nerve fibers were recorded in the region where IHCs were mostly missing. Liberman and Mulroy, 1982 What about the regions where hair cells remain, but substantial threshold shifts are still observed (the blue rectangle)? Liberman and Mulroy, 1982 A view of normal hair cells from above, showing the orderly rows of cilia. Kandel et al., 2000 Pickles, 1988 Cilia in the normal guinea pig cochlea, showing tip lengths (arrows) between the tips of shorter cilia and the sides of their longer neighbors. Pickles 1988 But intact hair cells may have damaged cilia after acoustic trauma Inner hair cell after acoustic trauma; damage is most severe to tallest cilia. Arrows show intact tip links, arrowheads show broken ones Outer hair cell after acoustic trauma; cilia are fused and missing tip links. Pickles, 1988 Normal View used to evaluate cilia in Liberman’s experiments Acoustic trauma Liberman and Dodds, 1984 Generally there was a good correlation between threshold shift and ciliary damage, in ears with surviving hair cells but substantial threshold shifts. Liberman and Mulroy, 1982 Acoustic trauma produces a complex and variable mixture of IHC and OHC damage. Ototoxic antibiotics (kanamycin) produce a simpler lesion in which regions with only OHC damage can be found. Unit 50 was localized using HRP injection to the middle of such a region (dashed line). This example shows the effects of a severe outer hair cell lesion, with little inner hair cell damage, including minimal cilia damage. Liberman and Dodds, 1984 Summary of the effects of different types of hair-cell damage IHC damage produces threshold shift with no loss of tuning. Liberman and Dodds, 1984 OHC damage produces threshold shift and broadened tuning. Auditory nerve fibers are apparently driven by two opposite-phase processes, called C1 and C2; examples are shown below for two fibers from normal animals. C1 C2 C1 C2 C2 tuning C1 tuning C1 C2 Apparently no C2 here Liberman and Kiang, 1984 Acoustic trauma seems to specifically affect C1 without modifying C2, shown below in two fibers from traumatized ears. reduced normal C1 C2 reduced normal C1 C2 C2 only Liberman and Kiang, 1984 Hypersensitive tails in W-shaped tuning curves occur when there is no sign of a C1C2 transition (no phase shift), presumably in the absence of C1. Liberman and Kiang correlated hypersensitive tails with cochlear regions where the IHCs are missing the tall row of stereocilia. Maybe C1 is the IHC driven through its tall stereocilia by sharply-tuned (due to the OHC) BM motion and C2 is the IHC driven through its short stereocilia. hypersensitive tail Normal fiber, same BF Liberman MC, Kiang, NY-S. Single-neuron labeling and chronic cochlear pathology. IV. Stereocilia damage and alterations in rate and phase-level functions. Hearing Res. 16:75-90 (1984). Fiber in traumatized ear No C1-C2 transition Liberman and Kiang, 1984