Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
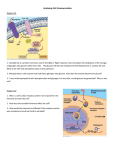
Article in press - uncorrected proof Biol. Chem., Vol. 390, pp. 125–135, February 2009 • Copyright by Walter de Gruyter • Berlin • New York. DOI 10.1515/BC.2009.013 Exploring the pathogenesis of renal cell carcinoma: pathway and bioinformatics analysis of dysregulated genes and proteins* Alexander D. Romaschin1,2, Youssef Youssef1, Tsz-fung F. Chow1, K.W. Michael Siu3, Leroi V. DeSouza3, R. John Honey4, Robert Stewart4, Kenneth T. Pace4 and George M. Yousef1,2,** Department of Laboratory Medicine and the Keenan Research Center in the Li Ka Shing Knowledge Institute, St. Michael’s Hospital Toronto, Toronto, M5B 1W8, Canada 2 Department of Laboratory Medicine and Pathobiology, University of Toronto, Toronto, M5B 1W8, Canada 3 Department of Chemistry and Center for Research in Mass Spectrometry, York University, Toronto, M3J 1P3, Canada 4 Division of Urology, Department of Surgery, St. Michael’s Hospital Toronto, University of Toronto, Toronto, M5B 1W8, Canada Keywords: bioinformatics; gene ontology (GO) consortium; kidney cancer; mass spectrometry; mammalian target of rapamycin (mTOR); pathway analysis; proteomics; renal cell carcinoma; tumor markers. 1 ** Corresponding author e-mail: [email protected] Abstract We recently identified a group of proteins which are dysregulated in renal cell carcinoma (RCC). In this study, we performed bioinformatics and pathway analysis of these proteins. Proteins were mapped to gene ontology biological processes. The upregulated proteins tend to cluster in processes, such as cancer initiation and progression. In addition, we identified a number of pathways that are significantly enriched in RCC. Some of these are ‘common’ pathways which are dysregulated in many cancers, but we also identified a number of pathways which were not previously linked to RCC. In addition to their potential prognostic values, many of these pathways have a potential as therapeutic targets for RCC. To verify our findings, we compared our proteins to a pool of datasets from published reports. Although there were only a minimal number of common proteins, there was a significant overlap between the identified pathways in the two groups. Moreover, out of 16 individually discovered genes identified by a literature search, 10 were found to be related to our dysregulated pathways. We also verified the upregulation of the mammalian target of rapamycin signaling pathway in RCC by immunohistochemistry. Finally, we highlight the potential clinical applications of pathway analysis in kidney cancer. *Supplementary material to this article can be accessed from the journal’s online edition at http://www.reference-global.com/ toc/bchm/390/2. Introduction A popular approach in understanding the pathogenesis of initiation and/or progression of cancer has been through the employment of genome-wide expression analysis at the mRNA level. More recently, proteomic analysis is becoming an increasingly important analytical tool which overcomes many of the limitations of RNA analysis (Faca et al., 2007). Unlike genomic studies where individual changes may have no functional significance, protein expression is closely aligned to cellular activity. Classically, individual analyses of tumor markers, especially the highly dysregulated ones, and their correlation with clinicopathological parameters helps in assessment of their potential clinical utility (DeSouza et al., 2005). More recently, however, focus of attention has begun to switch into a more ‘global’ analysis of dysregulated genes and proteins in order to obtain a better understanding of the potential ‘cross-talks’ between them (Setlur et al., 2007). Global analyses usually yield an enormous amount of information about molecules which are involved in cancer pathogenesis and can be used as tumor markers. Several databases and analytical tools are now available to the research community including the Gene Ontology (GO) consortium (for classification of genes according to their subcellular compartmentalization, biological processes and molecular functions), pathway analysis, and protein-protein interaction databases and analytical algorithms. Renal cell carcinoma (RCC) is one of the top 10 most frequent malignancies in Western societies. It is known to be one of the most therapy-resistant cancers. So far, little is known about the molecular changes in RCC, and there are no tumor markers which are of clinical use for diagnostic, prognostic, and treatment purposes. In our efforts to elucidate the pathogenesis of RCC initiation and progression, we recently identified a group of proteins which are dysregulated in kidney cancer tissues compared to their normal counterparts (Siu et al., 2008). In this paper, we further perform bioinformatics analysis including pathway and functional analysis. We validated one of the dysregulated pathways, the mammalian target of rapamycin (mTOR) signaling pathway, by immunohistochemistry. We also verified our bioinformatics results in silico through comparative analy- 2009/202 Article in press - uncorrected proof 126 A.D. Romaschin et al. sis with previously published dysregulated mRNAs and proteins in kidney cancer. Finally, we highlight the potential clinical applications of pathway analysis in RCC. Results Gene ontology analysis Dysregulated proteins in RCC were mapped to GO biological processes using multiple search engines, as described in the materials section. Figure 1 shows a partial list of the more significant biological processes which are dysregulated in kidney cancer compared to their normal counterparts. As expected, the upregulated proteins tend to cluster in biological processes which are related to cancer initiation and progression, e.g., cell cycle, cell motility, anti-apoptosis, and proteolysis. Other interesting processes include glycolysis (discussed below), RNA splicing, protein folding, signal transduction, and protein transport (Figure 1A). The downregulated proteins show a significantly different biological process clustering (Figure 1B). Some of these processes can be directly related to the malignant process, e.g., apoptosis, regulation of processing through cell cycle, signal transduction, and electron transport, while others, e.g., lipid metabolic processes, tricarboxylic acid (TCA) cycle, need to be investigated in relation to kidney cancer (discussed further below). Molecular function clustering was also carried out through the GO consortium. Enriched molecular functions among upregulated proteins include RNA binding, magnesium and metal ion binding, ATP, calcium and zinc ion binding, and oxidoreductase activity, while downregulated proteins show a significant clustering in actin binding, calcium, and zinc ion binding (data not shown). Pathway analysis Table 1 shows a partial list of the significantly enriched pathways in the clear cell type of RCC. Some of these pathways are ‘common’ pathways which are expected to be dysregulated in many cancers, such as the cell cycle, apoptosis, MAP kinase pathway, and cell adhesion, etc. We also identified a number of interesting pathways which were not previously linked to RCC, including insulin signaling, peroxisome proliferator-activated receptor (PPAR) signaling, hemostasis and blood coagulation, pyruvate metabolism and TCA cycle, formation of eEF1B complex, and regulation of actin cytoskeleton pathways. Literature searches showed that many of these pathways were previously linked to other malignancies. As discussed below, some of these pathways have a potential as therapeutic targets for RCC. It is important to mention that only few genes match to the kidney cancer pathway. This pathway, however, encompasses pathogeneses of all histological types of kidney cancers and is not restricted to the clear cell type of RCC which was analyzed in our study. It is also important to emphasize that statistical significance (either the impact factor or the p-value) is not equivalent to the biological significance in all cases, as the total number of molecules in a given pathway varies according to the database used, and some pathways are loosely defined and incorporate many subdivisions. Figure 1 Pie chart showing a partial list of the significantly altered biological processes in renal cell carcinoma, based on GO analysis. The biological processes which are enriched with upregulated proteins are significantly different from those with downregulated proteins. See text for details. Some irrelevant pathways were identified because of the presence of overlap in some segments with other pathways, e.g., Escherichia coli infection is overlapping with the cytoskeleton rearrangements and the apoptotic pathways. Activated and suppressed pathways in RCC To elucidate the functional relevance of dysregulated pathways, we analyzed them according to the number of upregulated and downregulated components. Although protein upregulation might not necessarily lead to the activation of a pathway, we can still assume activation of the pathway when the majority of its components are upregulated and vice versa. Figure 2 shows a representative graph of the up- and downregulated pathways. The upregulated pathways were those related to apoptosis, cell cycle, regulation of cytoskeleton, and cell adhesion. Many signaling pathways were also found to be upregulated, including PPAR signaling, insulin signaling, Wnt signaling, TGF-b signaling, and MAP kinase pathways. Article in press - uncorrected proof Pathway analysis in kidney cancer 127 Table 1 Partial list of significantly enriched pathways among dysregulated proteins in RCC. Pathway name Cell cycle Apoptosis TCA cycle Hemostasis and coagulation Regulation of actin cytoskeleton Cell-cell adhesion Leukocyte transendothelial migration PPAR signaling pathway Insulin signaling pathway Wnt signaling pathway TGF-b signaling pathway MAPK signaling pathway VEGF signaling pathway mTOR signaling pathway Calcium signaling pathway Rho GTPases signaling pathway Regulation of gene expression UTR-mediated translational regulation DNA repair mRNA processing Translation Metabolism of small molecules Metabolism of amino acids Metabolism of ketone body Metabolism of lipoprotein Metabolism of nucleotide Metabolism of xenobiotics Metabolism of lipid and lipoprotein Renal cell carcinoma Pathogenic Escherichia coli infection Antigen processing and presentation Axon guidance ECM receptor interaction Natural killer cell-mediated cytotoxicity Electron transport chain Formation of eEF1B complex Total molecules Positive matches Search enginea Significance factorb Upregulatedc Downregulatedc 114 84 49 21 127 69 208 195 116 70 135 149 84 256 70 47 175 132 310 109 85 136 124 70 72 5 13 81 68 118 69 49 82 128 87 131 76 3 9 6 6 10 14 13 31 23 14 13 13 8 5 11 4 2 4 8 41 17 5 18 23 29 20 4 6 16 11 16 3 15 10 9 6 6 6 2 1 1 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 2 2 2 2 2 2 2 2 2 2 2 2 2 1 1 1 1 1 1 2 2 5.261 3.439 3.80E-01 5.70E-05 8.60E-01 16.607 25.089 18.560 12.138 16.427 8.982 2.777 2.405 2.176 1.987 0.951 0.309 9.50E-01 7.30E-02 2.50E-02 9.20E-01 2.10E-01 1.10E-02 2.30E-11 2.10E-05 3.70E-04 6.70E-04 7.70E-03 5.20E-02 8.80E-02 1.168 25.929 9.144 4.528 3.292 1.780 7.50E-01 2.50E-02 4 4 2 0 4 4 9 12 6 7 3 6 4 5 2 1 2 3 21 9 1 8 13 18 4 1 5 7 7 9 1 11 3 5 3 3 0 1 3 2 3 9 8 5 7 6 6 5 2 1 1 3 2 1 0 1 11 3 2 6 5 7 13 3 0 8 0 6 2 3 3 3 1 3 5 1 Search engines used are: (1) Onto-Tools and (2) Reactome. Significance factor is the ‘impact factor’ for the Onto-Tools search engine and the ‘p’-value for Reactome. c Only proteins with G1.5-fold change were defined as up- or downregulated. a b Also, pathways related to regulation of gene expression, translational regulation, metabolism of lipoproteins, and xenobiotics were upregulated. Downregulated pathways included the TCA cycle, electron transport chain, coagulation and hemostasis, and the immune system signaling pathways. Also, DNA repair, amino acid metabolism, and ketone body metabolism showed underexpression of many members. Certain pathways, such as the actin cytoskeleton, cell adhesion, and antigen presentations pathways, showed a mixed pattern of up- and downregulation of its components. Our findings are quite comparable to previously published functional analyses in RCC (Boer et al., 2001; Gieseg et al., 2002). Differences can be attributed to the fact that previous studies were based on mRNA microarray analysis rather than proteins. Also, we analyzed ‘pathway’ dysregulation which is more accurate than functional taxonomy analysis which does not take into account the dynamics or dependencies that would be required to fully describe a pathway. Specimen heterogeneity may be another important factor. While many of the previous studies were based on analysis of a mixture of histological types, we focused only on the clear cell subtype of RCC. Validation of pathway analysis by independent datasets To verify our findings, we compared our results to a pool of datasets from 12 previously published lists of dysregulated genes and proteins in kidney cancer (Rae et al., 2000; Boer et al., 2001; Takahashi et al., 2001; Higgins et al., 2003; Lenburg et al., 2003; Seliger et al., 2003a,b; Liou et al., 2004; Shi et al., 2004; Perego et al., 2005; Craven et al., 2006; Perroud et al., 2006). This pool included both mRNAs and proteins, analyzed using a variety of techniques including differential display, microarray, and proteomic profiling on a spectrum of samples ranging from cell lines to tissues, urine, and blood. There was a minimal overlap (10–20%) between our signature protein list and previously published results (data not shown), which is consistent with the average Article in press - uncorrected proof 128 A.D. Romaschin et al. Pathway analysis of individually isolated protein markers To further validate our findings, we performed pathway analyses on previously identified ‘individual’ diagnostic markers for RCC to examine if they belong to our significant pathways. Out of 16 genes and proteins identified by literature searches, 10 were found to be related to our list of dysregulated pathways in kidney cancer. These markers are listed in Table 3, along with their clinical significance. Clinical applications of pathway analysis Figure 2 The frequency of up- and downregulated proteins in significantly dysregulated pathways in RCC. Dysregulated proteins were mapped to pathways and were classified as over- or underexpressed according to quantitative fold changes from normal kidney counterparts. overlap observed when analyzing protein datasets from different experiments (Yu et al., 2007). Focusing only on proteomic profiling studies, we compared our results with four previously published reports of differential protein expression in kidney cancer (Sarto et al., 1997; Shi et al., 2004; Perego et al., 2005; Craven et al., 2006). Only 24 proteins from our list of dysregulated proteins were identified in the other reports; 15 of them by one report, five by two reports and four by three studies (data not shown). We then hypothesized that these genes, despite the minimal overlap, are representatives of common pathways and that technical variations lead to identification of different members of the same pathway in each study. We performed a comparative pathway analysis between our proteins and the pool of datasets. A total of 3969 non-redundant genes and proteins were compiled from published literature and compared to our list of 869 dysregulated proteins against a database of 3030 pathway events. A total of 541 genes from the literature pool matched to 708 events (pathways), compared to a total of 253 of our proteins which matched 380 pathway events. As shown in Table 2, there was a significant overlap between the identified pathways in the two groups. These pathways include cell cycle, apoptosis, TCA cycle, insulin signaling pathways, PPAR receptor signaling, regulation of actin cytoskeleton, and hemostasis, among others (compare Tables 1 and 2). An interesting observation is that despite this significant overlap, the actual number of common proteins identified in these pathways was 10–20% on average, further proving our hypothesis that these pathways are involved in RCC pathogenesis, regardless of the genes identified. Technical differences between studies favor the identification of certain proteins in each study. In addition to their value in delineating cancer pathogenesis, we hypothesized that dysregulated pathways have direct clinical applications. We performed a literature search to examine the potential clinical applications of dysregulated pathways in both kidney and other cancer types. Table 4 shows selected clinical applications of the significantly altered pathways in different types of cancers. These applications range from being markers of aggressiveness and metastasis to therapeutic applications. The latter include immunotherapy, targeted molecular therapy in addition to chemotherapy. Promising preliminary results were observed in a variety of cancers, including breast, colon, and lung cancers, in addition to kidney cancer. Immunohistochemical validation of pathway analysis The mTOR pathway was chosen for immunohistochemical (IHC) validation because of its potential clinical value. Inhibitors of the mTOR have shown promising efficacy in early stage trials in patients with advanced RCC (Radulovic and Bjelogrlic, 2007). Both phosphorylated mTOR and S6 protein were found to be overexpressed in RCC compared to the normal kidney (p-0.05), in agreement with previously published studies (Lin et al., 2006; see Figure 3). Semi-quantitative RT-PCR validation of pathway analysis Three components in the coagulation pathway: F2, KNG1, and SERPING1, which were found to be dysregulated, were validated with semi-quantitative RT-PCR. The PCR result supported the original findings that F2 was underexpressed in RCC compared to normal kidney, while KNG1 and SERPING1 were overexpressed in RCC compared to normal kidney (p-0.05) (Figure 4). Discussion To our knowledge, this is the first study, at the protein level, which analyzes pathway changes in RCC based on quantitative protein expression from kidney cancer tissues. Global analysis has several advantages over the traditional, single molecule approach. Proteins are known to function in regulated networks or pathways, and bioinformatic approaches are thus useful in delineating the dynamic interactions between them. Moreover, while in individual protein analysis we focus on ‘highly’ dysregu- Article in press - uncorrected proof Pathway analysis in kidney cancer 129 Table 2 Comparative pathway analysis between our dysregulated proteins in RCC and a pool of 12 studies. Pathway Total pathway molecules Our study Comparison poola Common proteins Common protein symbols Search engineb Cell cycle 114 9 16 5 PCNA, SFN, YWHAB, YWHAG, YWHAH 1 Apoptosis 84 6 9 2 AIFM1, CYCS 1 Regulation of actin cytoskeleton 208 40 44 12 ACTB, ARPC3, F2, GSN, MSN, MYH10, MYH14, MYLK, RAC1, RAC2, RDX, ROCK2 1 Regulation of gene expression 310 41 48 7 EEF1A1, EIF4A1, RPS8, RPLP2, RPS28, RPS25, EEF2 2 Cell-cell adhesion 195 23 53 5 ACTB, COL1A1, RAC1, RAC2, ROCK2 1 21 10 10 5 OGDH, ACO2, SUCLG1, SUCLG2, PDHB 2 150 14 56 7 2 69 13 34 8 RAC1, ALB, F2, SERPINA1, HRG, FGA, FGB C3, CFB, F2, FGA, FGB, KNG1, SERPINA1, SERPING1 Leukocyte transendothelial migration 116 14 31 5 ACTB, MSN, RAC1, RAC2, ROCK2 1 Insulin signaling pathway 135 13 32 9 CALM2, FBP1, PCK1, PCK2, PFKP, PKLR, PKM2, PYGB, PYGL 1 PPAR signaling pathway 70 13 30 8 ACSL1, APOA1, FABP1, PABP5, FABP7, PCK1, PCK2, UBC 1 Immune system signaling pathway 325 7 78 3 IGHV4-31, F2, C3 2 Wnt signaling pathway 149 8 24 3 RAC1, RAC2, ROCK2 1 MAPK signaling pathway 256 11 38 3 MAPT, RAC1, RAC2 1 TGF-b signaling pathway 84 5 12 2 DCN, ROCK2 1 VEGF signaling pathway 70 4 7 2 RAC1, RAC2 1 Metabolism of small molecules 70 29 29 17 FBP1, ENO2, CALM1, ALDOA, KHK, TPI1, ALDOB, PYGL, PFKP, PCK1, PGK1, PKLR, HK1, PGMI, PGAM1, ENO1, PKM2 2 Metabolism of amino acids 72 20 39 13 OGDH, HIBADH, GATM, SHMT1, GOT1, CKB, ACAT1, GLUD1, QDPR, ALDH9A1, ALDH6A1, IVD, HIBCH 2 120 16 36 8 OXCT1, HMGCL, APOE, ALB, ACAT1, HADHA, SDCI, P4HB 2 Metabolism of xenobiotics 68 11 23 8 GSTP1, AHCY, ALDH1A1, NNMT, GSS, ACSM2B, ALDH2, GSTA2 2 Renal cell carcinoma 69 3 12 1 RAC1 1 TCA cycle Hemostasis and coagulation Metabolism of lipids and lipoprotein 1 Article in press - uncorrected proof 130 A.D. Romaschin et al. Table 2 (Continued) Pathway Total pathway molecules Our study Comparison poola Common proteins Common protein symbols Search engineb Antigen processing and presentation 82 10 29 8 CALR, CANX, CTSB, HLA- DRB1, HSP90AA1, HSPA5, LGMN, PDIA3 1 Pathogenic Escherichia coli rinfection 49 15 15 5 ACTB, CDH1, ROCK2, TUBB, TUBB2C 1 128 9 31 4 DPYSL2, RAC1, RAC2, ROCK2 1 Axon guidance A pool of 12 published studies on RCC was used for analysis. See text for full references. b Search engines used are: (1) Onto-Tools and (2) Reactome. a lated proteins (significantly up- or downregulated) which can be verified experimentally, pathway analysis allows the understanding of the role of minimally altered proteins which might be significant in the context of a biological pathway (Setlur et al., 2007). Many of the overrepresented pathways in our analysis were also shown to be dysregulated in other malignancies. Cell cycle pathway, MAP kinase, and cell adhesion pathways were implicated to play a role in the development of prostate cancer and to be signatures of metastatic potential (Setlur et al., 2007). Cross-talks between many of these pathways have been previously documented in various malignancies, e.g., the interaction between both the EGFR and PPAR signaling axis with the PI3K/Akt pathway in bladder cancer (Kassouf et al., 2006). Pathway alterations can have significant clinical applications. In addition to therapeutic intervention, pathwayderived signatures can also be used to predict prognosis in many cancers, e.g., breast cancer (Yu et al., 2007). An important pathway that was identified is the pyruvate metabolism and the TCA cycle, where 10 out of a Table 3 Individually discovered RCC biomarkers which belong to pathways identified in our study. Gene symbol Clinical application NNMT Potential diagnostic and prognostic biomarker Independent prognostic factor Overexpressed in RCC Overexpressed in the majority of renal cell carcinomas Identified in our study Reference Related pathway Yes Yao et al., 2005 Yes Yes Yes Takayama et al., 2006 Buchner et al., 2007 Hanada et al., 2001 Plexin B1 FGF5 Downregulated in RCC Overexpressed in RCC No No Gomez Roman et al., 2008 Hanada et al., 2001 MDM2 Overexpressed in RCC No Seliger et al., 2003a ABP1 Antigen for potential immunotherapy in RCC Promotes renal tumorigenesis Prognostic factor in RCC No Seliger et al., 2003a Metabolism of xenobiotics Nucleotide metabolism PPAR signaling pathway Glycolysis Gluconeogenesis Insulin signaling Axon guidance Regulation of actin cytoskeleton Melanoma, glioma, chronic myeloid leukemia and prostate cancer Amino acid metabolism Yes Yes Yan et al., 2007 George and Bukowski, 2007 Gluconeogenesis Rho GTPase signaling pathway ECGF1 FABP7 PKM2 PGK1 LDHB Table 4 Potential clinical utility of significantly altered pathways in RCC. Pathway Clinical application Cancer type Reference Glycolysis mTOR VEGF signaling pathway MAPK Immunotherapy targets Molecular targeted therapy Targeted therapy Therapeutic Cell cycle Focal adhesion Focal adhesion Wnt signaling Prognosis and therapeutic decision Hanada et al., 2001 Hanna et al., 2008 Sosman et al., 2007 Chen et al., 2001 van Spronsen et al., 2005 Vincenzi et al., 2006 Draghici et al., 2007 van Nimwegen and van de Water, 2007 He et al., 2005; Mikami et al., 2005 TGF-b Marker of invasiveness and metastatic capacity Kidney cancer Kidney cancer Kidney cancer Breast cancer Kidney cancer Lung cancer Lung cancer Breast cancer Colorectal cancer and sarcoma Breast cancer Anticancer therapy Therapeutic Todorovic-Rakovic, 2005 Article in press - uncorrected proof Pathway analysis in kidney cancer 131 Figure 3 mTOR pathway and overexpressions of its components in RCC. (A) The mTOR pathway. (B,C) Immunohistochemical staining of the phosphorylated S6 protein in normal kidney (B) and renal cell carcinoma (C). (D,E) Immunohistochemical staining of the phosphorylated mTOR protein in normal kidney (D) and renal cell carcinoma (E). Both proteins show overexpression in RCC compared to the normal kidney. Article in press - uncorrected proof 132 A.D. Romaschin et al. Figure 4 Semi-quantitative RT-PCR analysis of three dysregulated components (F2, KNG1, SERPING1) in the coagulation pathway. total of 21 events in the pathway were detected as dysregulated proteins in our list. In contrast to normal proliferating cells, tumor cells have to survive in environments with varying oxygen and nutrient supplies (Mazurek et al., 2005). The increase in lactate dehydrogenase and the activation of the pyruvate kinase pathway indicate active anaerobic glycolysis which is a reflection of the hypoxic conditions known to be an integral component of the pathogenesis of RCC (Turner et al., 2002; Kaelin, 2003). Our results are consistent with a recent report (Perroud et al., 2006) whereby pathway changes in RCC were analyzed. In another recent study, Hwa et al. (2006) pointed out dysregulation of hexokinase in RCC patients. Another interesting pathway is the hemostasis and blood coagulation pathway. RCC is known to be accompanied with dysregulation of the coagulation mechanism and a propensity for vascular thrombosis in the renal vein and inferior vena cava in up to 46% of cases (Hoehn and Hermanek, 1983). This has been confirmed by a number of coagulation studies (Zacharski et al., 1986). Moreover, recent studies have also shown that the essential factors of hemostasis support cancer growth and proliferation through providing a supportive scaffold for tumor angiogenesis (Rak et al., 2006). They also directly play a role in cancer cell ability for proliferation, migration, and induction of proteolysis (Buller et al., 2007). These findings inspired therapeutic applications, and a number of studies has evaluated the effects of anticoagulants on tumor growth and recurrence in various types of cancer (Buller et al., 2007). In addition, some components of the coagulation factors, e.g., tissue factor, is found in urine and might serve as cancer biomarkers for urologic cancers (Lwaleed et al., 2007). As shown in Table 1, the insulin signaling pathway is significantly presented in RCC (impact factor 8.982), with 13 pathway molecules identified in our list. The link between insulin signaling and kidney cancer is not surprising. Epidemiological studies have shown that insulinresistance states, characterized by hyperinsulinemia, are associated with an increased risk of a number of malignancies, including kidney cancer (Belfiore, 2007), and that insulin receptor is overexpressed in cancer. It has also been recently shown that insulin can control the mTOR signaling pathway (which is known to be dysregulated in RCC) by phosphorylation, mediated through PI3kinase signaling (Wang et al., 2005). Interestingly, our upregulated proteins in this pathway included ribosomal protein S6 kinase, a downstream target in the mTOR pathway which mediates rapid phosphorylation of ribosomal protein S6 on multiple serine residues in response to insulin or several classes of mitogens. This might have important implications for both the prevention and treatment of kidney cancer. They underline the concept that hyperinsulinemia, associated with insulin resistance and obesity, should be treated to avoid an increased risk of cancer. They also represent potential molecular targets for cancer therapy. Recent studies have shown that the level of insulin signaling is key in the regulation of cancer stem cells which function through at least two tumor suppressor genes (Narbonne and Roy, 2008). Our IHC results showing upregulation of members of the mTOR pathway in kidney cancer is not unprecedented. In fact, this pathway has recently attracted much attention for targeted therapy in kidney cancer, and the first successful phase III clinical trial involving mTOR inhibitors has been recently published (Hudes et al., 2007). The pathway involved in eEF1B complex formation (2 of 3 elements found) is presumed to have an essential role in the control of gene expression (Le et al., 2006). Regulation of actin cytoskeleton was also identified with potential involvement in RCC pathogenesis. This is not surprising, as loss of von Hippel-Lindau (VHL) tumor suppressor gene function occurs in familial and most sporadic clear cell RCC. VHL loss has been shown to allow robust RCC cell motility, invasiveness, and morphogenesis (Koochekpour et al., 1999). A recent study showed that VHL gene product represses oncogenic b-catenin signaling in renal carcinoma cells (Peruzzi et al., 2006). The PPAR signaling pathway also has a potential role in the course of RCC. PPARs are transcription factors which strongly influence molecular events in normal and cancer cells (Michalik et al., 2004). PPARs are nuclear receptors for linoleic and arachidonic acid metabolites. It has been shown that PPAR-g promotes colonic tumorigenesis, and that non-steroidal anti-inflammatory drugs (NSAIDs) suppress PPAR-d activity in colon cancer cells. Moreover, breast cancer cell lines that express PPAR-g can be prompted to undergo growth arrest and differentiation when treated with synthetic PPAR-g ligands. ANGPTL4, a member of this pathway, was present in very large amounts in RCC tumor cells, but not in benign kidney. Both as a protein encoded by a target gene of PPAR-a and PPAR-g, which have been shown to be associated with the regulation of lipid metabolism and/or glucose homeostasis and as a hypoxia-inducible secreted protein, ANGPTL4 has potential for use as a new diagnostic tool and a therapeutic target, modulating angiogenesis in tumors and ischemic tissues (Le et al., 2003). Changes in the lipid and lipoprotein metabolism can also be of clinical significance. The metabolic syndrome Article in press - uncorrected proof Pathway analysis in kidney cancer 133 is composed of cardiovascular risk factors, including increased body mass index/waist circumference, blood pressure, plasma glucose, and triglycerides, as well as decreased high-density lipoprotein cholesterol. Interestingly, most of the components of the metabolic syndrome have individually been linked in some way to the development of cancer, including kidney cancer (Cowey and Hardy, 2006). Our study provides a better resolution of the apparent differences and lack of consistency among studies, which might be related to many factors including the material used for analysis (tissue vs. serum vs. cell lines, etc.), the molecules investigated (genes or proteins) or the methodology used (microarray vs. proteomics, etc.). Regardless of these differences, we showed that these results can be very informative, being linked to specific pathways which are worth investigating. The recent success of molecular-targeted therapeutics in treating cancers where oncogenic pathways are well defined offers promise for overcoming the resistance to conventional chemotherapy displayed by RCC. Our results provide a rationale for the assessment of agents that target these pathways for efficacy against RCC. Materials and methods Quantitative protein expression in RCC We analyzed differentially expressed proteins in RCC compared to normal counterparts from the same patient using quantitative mass spectrometry analysis and simultaneous labeling. Overand underexpression were calculated as fold changes in protein concentrations. A summary of the procedure and complete list of the dysregulated proteins can be found in our previously published results (Siu et al., 2008), and in the online supplementary material. Bioinformatic analysis Official gene symbols, GenBank accession numbers and SwissProt IDs were compiled and converted through web tools of the Cancer Genome Anatomy Project (Krizman et al., 1999), The Database for Annotation, Visualization and Integrated Discovery (DAVID) (Dennis et al., 2003), and the ExPASy Proteomics Server. Dysregulated proteins were mapped to the Gene Ontology (GO) (Harris et al., 2004). We used a cut-off value of 1.5fold change for up- or downregulation. GO analysis was carried out using the Onto-Tools and DAVID algorithms. It should be noted that a GO biological process is not equivalent to a pathway, as GO does not try to represent the dynamics or dependencies which would be required to fully describe a pathway. For in silico validation, we utilized previously published genes and proteins which were shown to be differentially expressed in kidney cancer. Lists of studies included are indicated in the results section for each analysis. Literature searches were carried out through PubMed, BioMedCentral, and Google search engines. Pathway analysis Pathway analysis was performed through the KEGG pathway database (Kanehisa et al., 2008), DAVID Bioinformatics Resources (Dennis et al., 2003), the Reactome Knowledgebase of Human Biological Pathways and Processes (Vastrik et al., 2007), the Onto-Tools integrated databases (Draghici et al., 2003), and the BioCarta Pathways (http://www.biocarta.com/ index.asp). Official gene symbols and SwissProt accession numbers were used to map proteins to various pathways. Significantly presented pathways in the ‘Reactome’ databases were calculated by unadjusted probability (hypergeometric distribution) of seeing a given number or more genes in an event by chance, and significance was expressed by a p-value. Onto-Tool analysis calculates a perturbation factor for each input gene, which reflects the relative importance of each differentially regulated gene. Significance is presented as an impact factor of the entire pathway and takes into consideration the proportion of differentially regulated genes in the pathway and gene perturbation factors of all genes in the pathway. Immunohistochemical validation Paraffin blocks were sectioned 4 mm thick, mounted on slides, and dried overnight. Sections were deparaffinized in xylene and rehydrated through decreasing graded alcohols. Slides were immunostained using the Benchmark XT (Ventana, Tucson, AZ, USA) with monoclonal antibodies for phospho-mTOR (Ser2448) (Cell Signaling Tech, Danvers, MA, USA), and phosphor-S6 ribosomal protein (Ser235/236) (Cell Signaling Tech). Immune complexes were visualized by incubation with diaminobenzidine, and sections were counterstained with hematoxylin. All slides were reviewed and scored independently by two pathologists. Staining was scored for both percent positivity (in a scale from 1 to 4: 5–25%, 26–50%, 51–75%, and 76–100%, respectively) and intensity of staining (in a scale from 1 to 3: weak, moderate, and strong, respectively). An expression factor was assigned to each slide which is a multiplication of the positivity and intensity factors. Tissue collection and RNA extraction Tissues from 10 ccRCC tumors and their adjacent normal kidney tissues were extracted and flash frozen in liquid nitrogen immediately afterwards. In total, 20 mg of tissue from each sample were used in total RNA extraction using the miRNeasy Mini Kit and its recommended protocol (Qiagen Canada, Mississauga, ON, Canada). Semi-quantitative RT-PCR Total RNA RT reaction was performed on the RNA extractions using the OmniScript RT kit and its recommended protocol (Qiagen Canada). Primers designed to amplify the cDNA sequences of the targeted genes were obtained from Operon Biotechnologies (Huntsville, AL, USA); F2 forward primer: 59-ACT ACC GAG GGC ATG TGA AC-39, F2 reverse primer: 59-GCT GCA CAG CTG AGT TGA AG-39, KNG1 forward primer: 59-GGA ATC ACA GTC CGA GGA AA-39, KNG1 reverse primer: 59-AAG TTC AAT CCA GCC ACC AC-39, SERPING1 forward primer: 59-ATT CTC CTA CCC AGC CCA CT-39, SERPING1 reverse primer: 59-GGC GTC ACT GTT GTT GCT TA-39. The PCR reaction is performed with melting temperature at 948C, annealing temperature at 608C, and extension temperature at 728C for 30 cycles. The identity of the products was verified by an automated DNA sequencer (Applied Biosystems, Foster City, CA, USA). Equal amounts of PCR products and a house-keeping gene ran on 1.5% agarose gel and was visualized by ethidium bromide. Semi-quantitative expression data were calculated by densitometry measurements of band intensities. Article in press - uncorrected proof 134 A.D. Romaschin et al. References Belfiore, A. (2007). The role of insulin receptor isoforms and hybrid insulin/IGF-I receptors in human cancer. Curr. Pharm. Des. 13, 671–686. Boer, J.M., Huber, W.K., Sultmann, H., Wilmer, F., von Heydebreck, A., Haas, S., Korn, B., Gunawan, B., Vente, A., Fuzesi, L., et al. (2001). Identification and classification of differentially expressed genes in renal cell carcinoma by expression profiling on a global human 31,500-element cDNA array. Genome Res. 11, 1861–1870. Buchner, A., Castro, M., Hennig, A., Popp, T., Assmann, G., Hofstetter, A., Stief, C., and Zimmermann, W. (2007). wTranscriptome analyses in renal cell carcinoma. Combination of laser microdissection and microarraysx (in German). Urologe A 46, 1170–1175. Buller, H.R., van Doormaal, F.F., van Sluis, G.L., and Kamphuisen, P.W. (2007). Cancer and thrombosis: from molecular mechanisms to clinical presentations. J. Thromb. Haemost. 5 (Suppl. 1), 246–254. Chen, Z., Gibson, T.B., Robinson, F., Silvestro, L., Pearson, G., Xu, B., Wright, A., Vanderbilt, C., and Cobb, M.H. (2001). MAP kinases. Chem. Rev. 101, 2449–2476. Cowey, S. and Hardy, R.W. (2006). The metabolic syndrome: a high-risk state for cancer? Am. J. Pathol. 169, 1505–1522. Craven, R.A., Stanley, A.J., Hanrahan, S., Dods, J., Unwin, R., Totty, N., Harnden, P., Eardley, I., Selby, P.J., and Banks, R.E. (2006). Proteomic analysis of primary cell lines identifies protein changes present in renal cell carcinoma. Proteomics 6, 2853–2864. Dennis, G. Jr., Sherman, B.T., Hosack, D.A., Yang, J., Gao, W., Lane, H.C., and Lempicki, R.A. (2003). DAVID: database for annotation, visualization, and integrated discovery. Genome Biol. 4, 3. DeSouza, L., Diehl, G., Rodrigues, M.J., Guo, J., Romaschin, A.D., Colgan, T.J., and Siu, K.W. (2005). Search for cancer markers from endometrial tissues using differentially labeled tags iTRAQ and cICAT with multidimensional liquid chromatography and tandem mass spectrometry. J. Proteome Res. 4, 377–386. Draghici, S., Khatri, P., Bhavsar, P., Shah, A., Krawetz, S.A., and Tainsky, M.A. (2003). Onto-Tools, the toolkit of the modern biologist: Onto-Express, Onto-Compare, Onto-Design and Onto-Translate. Nucleic Acids Res. 31, 3775–3781. Draghici, S., Khatri, P., Tarca, A.L., Amin, K., Done, A., Voichita, C., Georgescu, C., and Romero, R. (2007). A systems biology approach for pathway level analysis. Genome Res. 17, 1537–1545. Faca, V., Krasnoselsky, A., and Hanash, S. (2007). Innovative proteomic approaches for cancer biomarker discovery. Biotechniques 43, 279, 281–3, 285. George, S. and Bukowski, R.M. (2007). Biomarkers in clear cell renal cell carcinoma. Expert. Rev. Anticancer Ther. 7, 1737–1747. Gieseg, M.A., Cody, T., Man, M.Z., Madore, S.J., Rubin, M.A., and Kaldjian, E.P. (2002). Expression profiling of human renal carcinomas with functional taxonomic analysis. BMC Bioinform. 3, 26. Gomez Roman, J.J., Garay, G.O., Saenz, P., Escuredo, K., Sanz, I.C., Gutkind, S., Junquera, C., Simon, L., Martinez, A., Fernandez Luna, J.L., and Val-Bernal, J.F. (2008). Plexin B1 is downregulated in renal cell carcinomas and modulates cell growth. Transl. Res. 151, 134–140. Hanada, K., Perry-Lalley, D.M., Ohnmacht, G.A., Bettinotti, M.P., and Yang, J.C. (2001). Identification of fibroblast growth factor-5 as an overexpressed antigen in multiple human adenocarcinomas. Cancer Res. 61, 5511–5516. Hanna, S.C., Heathcote, S.A., and Kim, W.Y. (2008). mTOR pathway in renal cell carcinoma. Expert. Rev. Anticancer Ther. 8, 283–292. Harris, M.A., Clark, J., Ireland, A., Lomax, J., Ashburner, M., Foulger, R., Eilbeck, K., Lewis, S., Marshall, B., Mungall, C., et al. (2004). The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 32, D258–D261. He, B., Reguart, N., You, L., Mazieres, J., Xu, Z., Lee, A.Y., Mikami, I., McCormick, F., and Jablons, D.M. (2005). Blockade of Wnt-1 signaling induces apoptosis in human colorectal cancer cells containing downstream mutations. Oncogene 24, 3054–3058. Higgins, J.P., Shinghal, R., Gill, H., Reese, J.H., Terris, M., Cohen, R.J., Fero, M., Pollack, J.R., van de Rijn, M., and Brooks, J.D. (2003). Gene expression patterns in renal cell carcinoma assessed by complementary DNA microarray. Am. J. Pathol. 162, 925–932. Hoehn, W. and Hermanek, P. (1983). Invasion of veins in renal cell carcinoma – frequency, correlation and prognosis. Eur. Urol. 9, 276–280. Hudes, G., Carducci, M., Tomczak, P., Dutcher, J., Figlin, R., Kapoor, A., Staroslawska, E., Sosman, J., McDermott, D., Bodrogi, I., et al. (2007). Temsirolimus, interferon a, or both for advanced renal-cell carcinoma. N. Engl. J. Med. 356, 2271–2281. Hwa, J.S., Kim, H.J., Goo, B.M., Park, H.J., Kim, C.W., Chung, K.H., Park, H.C., Chang, S.H., Kim, Y.W., Kim, D.R., et al. (2006). The expression of ketohexokinase is diminished in human clear cell type of renal cell carcinoma. Proteomics 6, 1077–1084. Kaelin, W.G. Jr. (2003). The von Hippel-Lindau gene, kidney cancer, and oxygen sensing. J. Am. Soc. Nephrol. 14, 2703– 2711. Kanehisa, M., Araki, M., Goto, S., Hattori, M., Hirakawa, M., Itoh, M., Katayama, T., Kawashima, S., Okuda, S., Tokimatsu, T., et al. (2008). KEGG for linking genomes to life and the environment. Nucleic Acids Res. 36, D480–D484. Kassouf, W., Chintharlapalli, S., Abdelrahim, M., Nelkin, G., Safe, S., and Kamat, A.M. (2006). Inhibition of bladder tumor growth by 1, 1-bis(39-indolyl)-1-(p-substitutedphenyl)methanes: a new class of peroxisome proliferator-activated receptor gamma agonists. Cancer Res. 66, 412–418. Koochekpour, S., Jeffers, M., Wang, P.H., Gong, C., Taylor, G.A., Roessler, L.M., Stearman, R., Vasselli, J.R., Stetler-Stevenson, W.G., Kaelin, W.G. Jr., et al. (1999). The von HippelLindau tumor suppressor gene inhibits hepatocyte growth factor/scatter factor-induced invasion and branching morphogenesis in renal carcinoma cells. Mol. Cell. Biol. 19, 5902–5912. Krizman, D.B., Wagner, L., Lash, A., Strausberg, R.L., and Emmert-Buck, M.R. (1999). The Cancer Genome Anatomy Project: EST sequencing and the genetics of cancer progression. Neoplasia 1, 101–106. Le, J.S., Amy, C., Cazes, A., Monnot, C., Lamande, N., Favier, J., Philippe, J., Sibony, M., Gasc, J.M., Corvol, P., et al. (2003). Angiopoietin-like 4 is a proangiogenic factor produced during ischemia and in conventional renal cell carcinoma. Am. J. Pathol. 162, 1521–1528. Le, S.F., Boulben, S., Le, B.R., Cormier, P., Morales, J., Belle, R., and Mulner-Lorillon, O. (2006). eEF1B: at the dawn of the 21st century. Biochim. Biophys. Acta 1759, 13–31. Lenburg, M.E., Liou, L.S., Gerry, N.P., Frampton, G.M., Cohen, H.T., and Christman, M.F. (2003). Previously unidentified changes in renal cell carcinoma gene expression identified by parametric analysis of microarray data. BMC Cancer 3, 31. Lin, F., Zhang, P.L., Yang, X.J., Prichard, J.W., Lun, M., and Brown, R.E. (2006). Morphoproteomic and molecular concomitants of an overexpressed and activated mTOR pathway in renal cell carcinomas. Ann. Clin. Lab. Sci. 36, 283–293. Liou, L.S., Shi, T., Duan, Z.H., Sadhukhan, P., Der, S.D., Novick, A.A., Hissong, J., Skacel, M., Almasan, A., and DiDonato, J.A. (2004). Microarray gene expression profiling and analysis in renal cell carcinoma. BMC Urol. 4, 9. Article in press - uncorrected proof Pathway analysis in kidney cancer 135 Lwaleed, B.A., Cooper, A.J., Voegeli, D., and Getliffe, K. (2007). Tissue factor: a critical role in inflammation and cancer. Biol. Res. Nurs. 9, 97–107. Mazurek, S., Boschek, C.B., Hugo, F., and Eigenbrodt, E. (2005). Pyruvate kinase type M2 and its role in tumor growth and spreading. Semin. Cancer Biol. 15, 300–308. Michalik, L., Desvergne, B., and Wahli, W. (2004). Peroxisomeproliferator-activated receptors and cancers: complex stories. Nat. Rev. Cancer 4, 61–70. Mikami, I., You, L., He, B., Xu, Z., Batra, S., Lee, A.Y., Mazieres, J., Reguart, N., Uematsu, K., Koizumi, K., and Jablons, D.M. (2005). Efficacy of Wnt-1 monoclonal antibody in sarcoma cells. BMC Cancer 5, 53. Narbonne, P. and Roy, R. (2008). Genes that affect both cell growth and polarity mediate stem cell quiescence. Front. Biosci. 13, 995–1002. Perego, R.A., Bianchi, C., Corizzato, M., Eroini, B., Torsello, B., Valsecchi, C., Di Fonzo, A., Cordani, N., Favini, P., Ferrero, S., et al. (2005). Primary cell cultures arising from normal kidney and renal cell carcinoma retain the proteomic profile of corresponding tissues. J. Proteome Res. 4, 1503–1510. Perroud, B., Lee, J., Valkova, N., Dhirapong, A., Lin, P.Y., Fiehn, O., Kultz, D., and Weiss, R.H. (2006). Pathway analysis of kidney cancer using proteomics and metabolic profiling. Mol. Cancer 5, 64. Peruzzi, B., Athauda, G., and Bottaro, D.P. (2006). The von Hippel-Lindau tumor suppressor gene product represses oncogenic b-catenin signaling in renal carcinoma cells. Proc. Natl. Acad. Sci. USA 103, 14531–14536. Radulovic, S. and Bjelogrlic, S.K. (2007). Sunitinib, sorafenib and mTOR inhibitors in renal cancer. J. BUON 12 (Suppl. 1), S151–S162. Rae, F.K., Stephenson, S.A., Nicol, D.L., and Clements, J.A. (2000). Novel association of a diverse range of genes with renal cell carcinoma as identified by differential display. Int. J. Cancer 88, 726–732. Rak, J., Yu, J.L., Luyendyk, J., and Mackman, N. (2006). Oncogenes, Trousseau syndrome, and cancer-related changes in the coagulom of mice and humans. Cancer Res. 66, 10643–10646. Sarto, C., Marocchi, A., Sanchez, J.C., Giannone, D., Frutiger, S., Golaz, O., Wilkins, M.R., Doro, G., Cappellano, F., Hughes, G., et al. (1997). Renal cell carcinoma and normal kidney protein expression. Electrophoresis 18, 599–604. Seliger, B., Lichtenfels, R., and Kellner, R. (2003a). Detection of renal cell carcinoma-associated markers via proteome- and other ‘ome’-based analyses. Brief. Funct. Genomics Proteomics 2, 194–212. Seliger, B., Menig, M., Lichtenfels, R., Atkins, D., Bukur, J., Halder, T.M., Kersten, M., Harder, A., Ackermann, A., Beck, J., et al. (2003b). Identification of markers for the selection of patients undergoing renal cell carcinoma-specific immunotherapy. Proteomics 3, 979–990. Setlur, S.R., Royce, T.E., Sboner, A., Mosquera, J.M., Demichelis, F., Hofer, M.D., Mertz, K.D., Gerstein, M., and Rubin, M.A. (2007). Integrative microarray analysis of pathways dysregulated in metastatic prostate cancer. Cancer Res. 67, 10296–10303. Shi, T., Dong, F., Liou, L.S., Duan, Z.H., Novick, A.C., and DiDonato, J.A. (2004). Differential protein profiling in renal-cell carcinoma. Mol. Carcinog. 40, 47–61. Siu, M.K., DeSouza, L., Scorilas, A., Romaschin, A., Honey, J., Stewart, R., Pace, K., Youssef, Y., Chow, F., and Yousef, G. (2008). Differential protein expressions in renal cell carcino- ma: new biomarker discovery by mass spectrometry. J. Proteome Res., in press. Sosman, J.A., Puzanov, I., and Atkins, M.B. (2007). Opportunities and obstacles to combination targeted therapy in renal cell cancer. Clin. Cancer Res. 13, 764s–769s. Takahashi, M., Rhodes, D.R., Furge, K.A., Kanayama, H., Kagawa, S., Haab, B.B., and Teh, B.T. (2001). Gene expression profiling of clear cell renal cell carcinoma: gene identification and prognostic classification. Proc. Natl. Acad. Sci. USA 98, 9754–9759. Takayama, T., Mugiya, S., Sugiyama, T., Aoki, T., Furuse, H., Liu, H., Hirano, Y., Kai, F., Ushiyama, T., and Ozono, S. (2006). High levels of thymidine phosphorylase as an independent prognostic factor in renal cell carcinoma. Jpn. J. Clin. Oncol. 36, 564–569. Todorovic-Rakovic, N. (2005). TGF-b 1 could be a missing link in the interplay between ER and HER-2 in breast cancer. Med. Hypoth. 65, 546–551. Turner, K.J., Moore, J.W., Jones, A., Taylor, C.F., Cuthbert-Heavens, D., Han, C., Leek, R.D., Gatter, K.C., Maxwell, P.H., Ratcliffe, P.J., et al. (2002). Expression of hypoxia-inducible factors in human renal cancer: relationship to angiogenesis and to the von Hippel-Lindau gene mutation. Cancer Res. 62, 2957–2961. van Nimwegen, M.J. and van de Water, B. (2007). Focal adhesion kinase: a potential target in cancer therapy. Biochem. Pharmacol. 73, 597–609. van Spronsen, D.J., Mulders, P.F., and de Mulder, P.H. (2005). Novel treatments for metastatic renal cell carcinoma. Crit. Rev. Oncol. Hematol. 55, 177–191. Vastrik, I., D’Eustachio, P., Schmidt, E., Joshi-Tope, G., Gopinath, G., Croft, D., de Bono, B., Gillespie, M., Jassal, B., Lewis, S., et al. (2007). Reactome: a knowledge base of biologic pathways and processes. Genome Biol. 8, R39. Vincenzi, B., Schiavon, G., Silletta, M., Santini, D., Perrone, G., Di, M.M., Angeletti, S., Baldi, A., and Tonini, G. (2006). Cell cycle alterations and lung cancer. Histol. Histopathol. 21, 423–435. Wang, X., Beugnet, A., Murakami, M., Yamanaka, S., and Proud, C.G. (2005). Distinct signaling events downstream of mTOR cooperate to mediate the effects of amino acids and insulin on initiation factor 4E-binding proteins. Mol. Cell. Biol. 25, 2558–2572. Yan, Q., Bartz, S., Mao, M., Li, L., and Kaelin, W.G. Jr. (2007). The hypoxia-inducible factor 2a N-terminal and C-terminal transactivation domains cooperate to promote renal tumorigenesis in vivo. Mol. Cell. Biol. 27, 2092–2102. Yao, M., Tabuchi, H., Nagashima, Y., Baba, M., Nakaigawa, N., Ishiguro, H., Hamada, K., Inayama, Y., Kishida, T., Hattori, K., et al. (2005). Gene expression analysis of renal carcinoma: adipose differentiation-related protein as a potential diagnostic and prognostic biomarker for clear-cell renal carcinoma. J. Pathol. 205, 377–387. Yu, J.X., Sieuwerts, A.M., Zhang, Y., Martens, J.W., Smid, M., Klijn, J.G., Wang, Y., and Foekens, J.A. (2007). Pathway analysis of gene signatures predicting metastasis of node-negative primary breast cancer. BMC Cancer 7, 182. Zacharski, L.R., Memoli, V.A., and Rousseau, S.M. (1986). Coagulation-cancer interaction in situ in renal cell carcinoma. Blood 68, 394–399. Received July 8, 2008; accepted October 1, 2008