Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
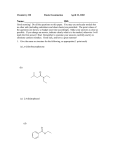
547th MEETING, LONDON 657 Iron-Binding Compounds Formed by Micrococcus denitri9cans G. H. TAIT Department of Chemical Pathology, St. Mary's Hospital Medical School, London W2 l P C , U.K. 2,3-Dihydroxybenzoic acid and a number of compounds containing it are excreted by some micro-organisms grown in media deficient in iron (Neilands, 1972). In this report it is shown that Micrococcus denitrificans grown in media deficient in iron excretes 2,3dihydroxybenzoic acid and two previously undescribed compounds containing 2,3dihydroxybenzoic acid and spermidine. When M. denitri$caris is grown anaerobically in a defined nitrate medium deficient in iron or containing high concentrations of calcium and copper, or aerobically in a defined ammonia medium deficient in iron (Tait, 1973), compounds which chelate iron accumulate in the medium. Three compounds have been isolated which contain catechol residues as shown by their reaction with Arnow's nitrite-molybdate reagent (cf. Corbin & Bulen, 1969). Two of the compounds were extracted into ethyl acetate from medium acidified to pH2. On extracting the ethyl acetate with 1 . 2 ~ - N a H C o one ~ , of them (compound I) was extracted into the aqueous phase; the other (compound 111) remained in the organic phase. These compounds were purified by chromatography on silicic acid. Compound I was crystallized from hot water and compound 111was obtained as a white powder by adding a concentrated solution in ethanol to water and freeze-drying the suspension. The acidified medium from an iron-deficient aerobic culture, after extraction with ethyl acetate, was freeze-dried. The solid was extracted with ethanol, the ethanolic solution was dried, and the residue was dissolved in water. Compound I1 was purified by chromatography on Zeokarb 225 [H+]and then on CM-cellulose. Compound I was found to be 2,3-dihydroxybenzoic acid. The melting point, infra-red spectrum, ultra violet spectra in aqueous solutions of different pH values and in organic solvents, and the chromatographic behaviour on paper in a number of solvents were identical with those of an authentic sample. Compound I1 contained 2,3-dihydroxybenzoic acid and spermidine in a molar ratio of 2/1. It did not react with ninhydrin, showing that both primary amine groups of spermidine were blocked. It reacted with fluorodinitrobenzene, and from the spectrum of the product (Dubin, 1960) it was shown that the secondary amine group of spermidine was free. It is concluded that compound I1 is N'N8-bis-(2,3-dihydroxybenzoyl)spermidine. Compound I11 on acid hydrolysis yielded 2,3-dihydroxybenzoic acid, 2-hydroxybenzoicacid, L-threonineand spermidine in a molar ratio of 2 :1 : 1 : 1. Partial acid hydrolysis yielded a dipeptide containing threonine and spermidine. The spectrum of the product after reaction with fluorodinitrobenzene showed that the dipeptide did not have a free secondary amine group, and thus it is probable that threonine is linked by its carboxyl group to the secondary amine group of spermidine. On the basis of the enzymic experiments described below, and from the structure assigned to compound 11, it is probable that compound I11 is 2-hydroxybenzoyl-N-~-threonyl-N~[N'N~-bis-(2,3-dihydroxybenzoy1)lspermidine. This compound forms a red equimolar complex with Fe3+. A concentrated suspension of iron-deficient M . denitrgcans was disrupted by ultrasonication and centrifuged at 25000g for 30min. The clear supernatant, after dialysis, was used as enzyme. The assay mixture contained Tris-HC1 buffer, pH8.8, MgCI2, dithiothreitol, ATP, enzyme and, where added, L-threonine, spermidine, 2,3-dihydroxybenzoic acid and 2-hydroxybenzoic acid. [14C]Threonine or ['4C]spermidine was also added. After incubation HCI and ethyl acetate were added. After thorough mixing the ethyl acetate layer was removed. Portions of the ethyl acetate were taken for measurement of radioactivity and for paper chromatography. On incubation of [14C]spermidine and 2,3-dihydroxybenzoic acid, with or without threonine, a radioactive compound with the chromatographic properties of compound Vol. 2 658 BIOCHEMICAL SOCIETY TRANSACTIONS I1 was formed. No radioactivity was extracted into ethyl acetate in the absence of 2,3dihydroxybenzoic acid, or when 2-hydroxybenzoic acid was used in its place. A radioactive compound with the chromatographic properties of compound 111 was formed on incubation of [14C]spermidine, threonine, 2,3-dihydroxybenzoic acid and 2-hydroxybenzoic acid. On incubation of [‘4C]threonine, spermidine, 2,3-dihydroxybenzoic acid and 2hydroxybenzoic acid radioactive compound I11 was formed. In the presence of only one of the aromatic acids only small amounts of compounds containing [14C]threoninewere extracted into ethyl acetate. When compound I1 was incubated with [14C]threonine and 2-hydroxybenzoic acid it was converted into radioactive compound 111. Incubation of compound I1 with [‘4C]threonine and 2,3-dihydroxybenzoic acid resulted in the formation of only a small amount of a radioactive compound with the chromatographic properties of compound 111. I thank Mrs. C. Eggleton for her excellent technical assistance and Dr. R. C. Davies for helpful advice and discussions. Corbin, J. L. & Bulen, W. A. (1969) Biochemistry 8,757-762 Dubin, D. T. (1960) J. Biol. Chem. 235 783-786 Neilands, J. B. (1972) Struct. Bonding 11, 145-170 Tait, G. H. (1973) Biochem. J. 131, 389-403 Effect of Growth Conditions and Dibntyryl Adenosine 3’:5’-Cyclic Monophosphate Treatment on L-Glutamate and L-Alanine Transport in BMK28-CB3 Cells DOUGLAS M. SCOTT and JOHN A. PATEMAN Institute of Genetics, University of Glasgow, Glasgow GI 1 5JS, U.K. Intracellular concentrations of naturally occurring amino acids and their analogues have been shown to influence the inward transport of those molecules in bacterial, fungal and certain mammalian cells either by transinhibition (Ring & Heinz, 1966; Belkhode & Scholefield, 1969; Pall, 1971) or by control mechanisms effective on protein synthesis (Pall, 1971 ;Phang et al., 1971). Treatment of tissue slices or cells in culture with various hormones alters amino acid transport rates (Krawitt et al., 1970; Baril et al., 1969). Receptors of hormones of the ‘primary messenger class’ (Robinson et al., 1971) are closely related to and may be part of an adenyl cyclase system, and act to alter intracellular concentrations of the ‘secondary messenger’ cyclic AMP. Reports have claimed that cyclic AMP or its dibutyryl derivative can alter the morphology of certain cell lines (Hsie &Puck, 1971;Hsie et al., 1971; Sheppard, 1971), affect growth rates of certain cell lines (Heidrick & Ryan, 1970; Blat et al., 1973), and alter the transport activities of various metabolites e.g. nucleotides (Hauschka et al., 1972; Kram et al., 1973), sugars (Kram & Tomkins, 1973) and phosphate (Blat et al., 1973). The present paper describes the effect on L-alanine and L-glutamate uptake after the following treatments (a), growth in the presence of certain amino acids (b), treatments that are reported to alter intracellular cyclic AMP concentrations or (c), treatment of the cells with dibutyryl cyclic AMP. L-Glutamate and L-alanine transport were studied by determining the incorporation of the appropriate labelled amino acid into cells grown in monolayers on coverslips or in petri dishes. Before transport studies the cells were grown in Glasgow’s modified Eagles medium plus 10% (v/v) or 0.5% (v/v) calf serum, with the addition in certain experiments of dibutyryl cyclic AMP (0.2-2m~)with or without theophylline (1 mM), or by theophylline (1 mM) alone. In other experiments the medium was supplemented with 1974