Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
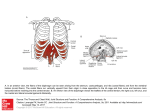
Electrophysiological Characteristics of Canine Atrial Plateau Fibers By Perry M . Hogan and Larry D. Davis Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 ABSTRACT Right atria from dog hearts were immersed in Tyrode's solution with the endocardial surface of the anterior wall exposed. Glass microelectrodes were used to impale fibers in diSerent anatomical areas of this preparation. Electrical characteristics of atrial plateau fibers located along the caval border of the crista terminalis were compared with those of regular atrial fibers from other sites. The former conducted at a rate (0.803 ±0.110 M/sec) two to three times more rapidly than pathways composed of regular atrial fibers. A period of supernormal excitability was demonstrated for plateau fibers but not for regular fibers. Graded premature responses could be elicited in plateau fibers by both electrical stimulation and propagated action potentials. Duration of the effective refractory period of plateau fibers (196.1 ± 2.5 msec) was longer than that of regular fibers (147.8 ± 5.4 msec). Plateau fibers required a lower level of membrane potential (—62.1 ± 0.8 mv) to generate a propagated action potential than did regular fibers (—65.5 ± 1.3 mv). Regular atrial fibers could respond at faster rates to high frequency stimulation than plateau fibers. KEY WORDS conduction velocity supernormal excitability premature excitation • Anatomical evidence for the existence of extranodal specialized fibers in mammalian atria was provided by the early work of Wenckebach (1), Thorel (2, 3), and Bachmann (4, 5). More recently, James (6-9) and others (10-14) have confirmed the presence of specialized pathways in the atria of several mammalian species. In each of these studies fibers were found in the atria which had certain histological characteristics similar to those of ventricular Purkinje fibers. Electrophysiological evidence for specialized atrial pathways was provided in the studies of Eyster and Meek (15-19). They From the Department of Physiology, Medical School, University of Wisconsin, Madison, Wisconsin 53706. This investigation was supported in part by U. S. Public Health Service Grants 1-R01-HE-12780-07 and 1-R01-HE-13375-01 and grants from the Wisconsin Heart Association and the Western New York Heart Association. Dr. Hogan's present address is Department of Physiology, State University of New York at Buffalo, Buffalo, New York 14214. Received August 31, 1970. Accepted for publication November 10, 1970. 62 effective refractory period rapid frequency stimulation showed that sinoventricular transmission could be impaired to different degrees by placing cuts along the borders of the sinoatrial node. From this they postulated that specialized pathways exist which normally convey the sinus impulse to the atrioventricular node. Lewis and his co-workers (20, 21) argued against this idea and in favor of the concept of radial spread of the sinus impulse with uniform velocity through homogeneous atrial muscle. More recent findings provided by studies in which cellular transmembrane potentials were recorded have shown the presence of extranodal specialized atrial fibers in the hearts of several species (22-34). Recent work from this laboratory has presented evidence for a type of specialized fiber in the right atrium of the dog (31, 34). The contour of the action potential of these fibers is remarkably similar to that of ventricular Purkinje fibers. Because of a prominent plateau during repolarization (phase 2), the fibers were called atrial plateau fibers. Such fibers were found consistently along the venous border of the crista terminalis. This is the Circulation Research, Vol. XXVIII, January 1971 63 ATRIAL PLATEAU FIBERS Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 approximate location of the posterior internodal tract described by James (6-9), and it was suggested (34) that atrial plateau fibers might compose this tract. In the present study, certain electrophysiological characteristics of these atrial plateau fibers were determined. Conduction velocity was measured along the pathway composed of platetau fibers and was compared with similar measurements along several pathways of the right and left atria composed predominantly of regular muscle fibers. Childers et al. (33), in their study of the specialized fibers of Bachmann's bundle, concluded that the presence of a phase of supernormal excitability during repolarization was the most "reliable and distinctive" feature of these specialized fibers. In the present study, atrial plateau fibers were shown to have a period of supernormal excitability. Finally, we studied the response of plateau fibers to premature excitation and measured the duration of the effective refractory period of both plateau and regular atrial muscle fibers. Methods Hearts were excised from dogs anesthetized with sodium pentobarbital (30 mg/kg, i.v.). A right atrial tissue preparation as described in an earlier report (34) was dissected from the heart and pinned under slight tension to a paraffin block in the tissue bath. Fibers located along the caval border of the crista terminalis were studied in the experiments described below. Based on the contour of the action potential they have been designated as atrial plateau fibers (34). In many preparations, a free-running strand of tissue similar to the false tendons of the ventricle connected the caval border of the crista terminalis with the upper pectinate muscles of the anterior free wall of the right atrium. The fibers comprising the free-running strand were found to be plateau fibers and were included in this study. The electrical characteristics of plateau fibers were compared with those of regular atrial muscle fibers located in the crista terminalis, pectinate muscles, or epicardial surface of the left atrial appendage. Tyrode's solution equilibrated in a reservoir with 95% oxygen and 5% carbon dioxide flowed continuously through the tissue bath at a rate of 25 ml/min. The composition of the solution in millimoles per liter was: NaCl 137, dextrose 5.5, KC1 2.7, Cad 2 2.7, MgCl2 0.5, NaH2PO4 1.8, Circulation Research, Vol. XXVIII, January 1971 NaHCOg 25. Temperature in the tissue bath was maintained at 35° to 37°C but remained constant during each experiment. The pH of the solution was 7.3 to 7.4. Transmembrane potentials of atrial fibers on the endocardial surface of the preparation were recorded using glass microelectrodes filled with 3M KC1. Details of the recording system used have been described previously (34). Conduction velocities along linear pathways in five anatomically distinct areas of the atria were determined by the method described by Paes de Carvalho et al. (22). Briefly, drive stimuli, 5 msec in duration and at a rate of 95/min, were applied to one end of the pathway under study through a unipolar stimulating electrode. One microelectrode was inserted into a cell close to the stimulating electrode. The onset of the action potential at this site served as a fixed reference point from which to time the arrival of the impulse at distal sites. In an alternative procedure, this reference electrode remained extracellular and the stimulus artifact was used as the reference point. A second microelectrode was used as an exploring electrode to impalefibersof the same type at increasing distances away from the reference electrode. The type of fibers, composing each pathway studied was based on inspection of the action potential recorded at each impalement site. The distance which separated the two microelectrodes was read to 0.1 mm from a vernier scale attached to the micromanipulator used to position the exploring microelectrode. The upstrokes of the action potentials from both proximal and distal sites were displayed at a fast oscilloscope sweep speed and photographed. For analysis these records were projected on the screen of a Benson-Lehner X-Y film reader. The distance between the upstrokes of the two action potentials was determined and converted to the time in milliseconds required for excitation to pass between the two recording sites. These data were plotted against the distance which had separated the two microelectrodes, and the slope of the resulting curve, computed using the least-square equation for linear regression, was taken as the conduction velocity along the pathway. Analysis of variance was used to test for the presence of significant differences between the groups of conduction velocities which were based on anatomical location. Duncan's multiple range test (35) was used to compare the mean of each group with the mean of every other group and thereby locate the site of significant difference between groups. To test for supernormal excitability in atrial plateau fibers, the method of Childers et al. (33) was used. A special stimulating and recording 64 Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 circuit was employed1 which made it possible to stimulate and record through the same intracellular microelectrode. Microelectrodes with relatively low tip impedance (5 megohms) were used to facilitate passage of stimulating current. For these experiments, the tissue was driven at a constant rate (95/min) by administering drive stimuli (SI) through a unipolar surface electrode. A test stimulus (S2) could be applied through the intracellular microelectrode at any preselected time after a given drive stimulus. Initially, S2 was delivered midway in the period of electrical diastole, well after the maximum diastolic potential was attained. The amplitude of S2 was increased until it succeeded in producing a premature response with each application. Then its amplitude was lowered just enough so that it repeatedly failed to cause a response. The interval between SI and S2 then was reduced by increments until S2 successfully produced a response. With further reduction of the interval between SI and S2, a point was reached when the latter stimulus again failed to produce a premature action potential. Subsequently, S2 was abruptly repositioned to mid-diastole, where it again failed to elicit responses. Responses to premature excitation and duration of the effective refractorv period of both plateau and regular atrial muscle fibers were measured by administering cathodal drive and test stimuli directly to the tissue under study. The fiber studied was always within 1 mm of the unipolar stimulating electrode. A test stimulus, 5 msec in duration and two times the diastolic threshold, could be applied at any time during a driven action potential. Initially, the test stimulus was given during the plateau or early phase 3 of the action potential and elicited no premature response. On subsequent trials the test pulse was moved by small increments progressively later in the cycle. Six to eight drive stimuli were allowed in succession before application of a single test pulse. Two methods were used to determine if the test response was propagated. When the earliest test response was of relatively large magnitude and depolarized to or beyond the line of zero potential difference, it was assumed to be propagated (36). The second method required the use of a second microelectrode inserted into a fiber located several millimeters from the stimulus site. The end of the effective refractory period was taken as the point at which an action potential elicited by the test stimulus propagated to the distal recording site (37). This method was HOGAN, DAVIS used in studies in which early test stimuli elicited graded responses. The ability of the fibers to respond to rapid rates of stimulation was determined by applying stimuli at progressively shorter cycle lengths. Cycle lengths of 630, 500, 430, 320, 250, 200, 160, and 100 msec were used. Starting with the longest cycle length, stimuli were applied for 5second intervals at progressively shorter cycle lengths. Records were taken continuously during the entire determination by moving the film past a vertically deflecting oscilloscope beam. Results CONDUCTION VELOCITY Conduction velocity was measured along linear pathways in five areas of the right and left atria. Four pathways located on the endocardial surface of the right atrium were (1) along the free-running strand, (2) along the caval border of the crista terminalis, (3) along the middle or auricular border of the crista terminalis, and (4) along the long axis of a pectinate muscle. The former two pathways were composed predominantly of atrial plateau fibers while the latter two pathways were composed of regular atrial fibers. A fifth conduction pathway along the epicardial surface of the left atrial appendage was composed of regular atrial fibers. Conduction velocities, as indicated from the calculated slope of sequential activation time versus linear distance plots, were determined for 36 such pathways in 12 atrial preparations. It was assumed that if the exploring electrode was moved along a linear path in the same direction as the wave of excitation that the TABLE 1 Conduction Velocity Analysis (Duncan's MultipleRange Test) M/sec 1. Epicardial left auricle 2. Pectinate muscle 3. Middle and auricular crista terminalis 4. Free-running strand 5. Caval crista terminalis N .344 .431 .604 (5) (5) (10) .786 .803 (4) (6) 1 Designed and constructed by Mr. Joseph French, Department of Rehabilitation Medicine, University of Wisconsin, Medical School, Madison, Wisconsin 53706. There is no significant difference at the 5% level between the mean values for 1 and 2, or 2 and 3, or 4 and 5. Circulation Research, Vol. KXVlll, January 1971 ATRIAL PLATEAU FIBERS 65 Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 ACTIVATION TIME (IT Graphs of linear distance versus activation time measured for five anatomically distinct areas of the canine atria: open stars = caval crista terminalis; triangles = free-running strand; circles = middle and auricular crista terminalis; solid stars = pectinate muscle; half circles = epicardial left appendage. The slope of each line indicates the conduction velocity for that pathway. The data points for pathways composed predominantly of atrial plateau fibers are fitted by solid straight lines. Those composed of regular atrial fibers are fitted by broken lines. Demonstration of supernormal excitability in an atrial plateau fiber. A: Test stimulus (S2) of subthreshold strength placed midway in electrical diastole (the stimulus artifact [arrow] seen during phase 4 indicates the time of stimulation). B: S2 stimulus elicited a response during supernormal period. C: S2 stimulus was ineffective when delivered earlier during repolarization. In this and subsequent figures, time and voltage calibrations are indicated by the horizontal parallel black lines. The vertical distance separating the lines equals 100 mv in all figures. In this figure the spikes on the top line are at 200-msec intervals. Circulation Research, Vol. XXVlll, January 1971 coordinate points would show little scatter and be fitted by a straight line (22). Six plots were rejected for their excessive scatter and nonlinearity. Table 1 summarizes the results of the remaining 30 experiments. Analysis of variance for the five conduction velocity groups indicated a significant difference between groups. To locate the site of this difference, Duncan's multiple-range test (35) was employed at the 5% level. Table 1 shows the ranked order of mean velocities and further indicates where there was a significant difference between groups. Conduction velocities along the free-running strand and caval border of the crista terminalis composed of plateau fibers were not significantly different and were the fastest of all pathways measured. Regular muscle fibers composed the other three pathways and conducted the impulse at a significantly slower rate (P < 0.005) when compared with the specialized cells. Figure 1 shows typical conduction velocity plots for the five anatomical areas investigated. 66 HOGAN, DAVIS Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 Premature responses observed during measurement of the effective refractory period in atrial plateau fibers. The upper action potential of each record is from a plateau fiber close to the stimulating electrode. The lower action potential of each record is from a plateau fiber several millimeters distal to the stimulating electrode. Application of the test stimulus is indicated by the appearance of the stimulus artifact on the repolarization phase of the action potential. A-E show graded responses in the proximal fiber as the test stimulus was given progressively later during repolarization. The premature response at the stimulus site was eventually propagated to the distal fiber as shown in F. Discussion in text. Time spikes at 100-msec intervals. SUPERNORMAL EXCITABILITY It has been suggested that a period of supernormal excitability is a characteristic of specialized cardiac fibers (33). In the present study, atrial fibers in the free-running strand and along the crista terminalis were tested for a period of supernormal excitability. The presence of such a period during the terminal phase of repolarization was demonstrated, as shown in Figure 2. An intracellular stimulus which failed to excite in mid-diastole (A) always was successful when applied shortly before complete repolarization (B), then again failed when applied earlier during phase 3 (C). Similar results were obtained in 17 fibers from five hearts studied. It was not possible to test for supernormal excitability in regular atrial fibers because the amount of current which could be passed through the microelectrode was inadequate to produce excitation of these fibers at any point in the cycle. RESPONSES TO PREMATURE STIMULATION In these experiments, cathodal stimuli were applied at various intervals during phase 3 of the action potential. Figure 3 shows the typical response of plateau fibers to this procedure. The earliest detectable response usually occurred midway during phase 3 repolarization and consisted of a depolariza- tion of small magnitude with slow rising velocity and short duration (A and B). When the test stimuli were given progressively later in the cycle, at higher levels of membrane potential, the test responses had proportionately larger magnitudes, rising velocities and durations (C-E). Such responses are called graded responses and conduct with decrement (38). Figure 3 (A-E) shows that the graded responses failed to propagate throughout the preparation, as indicated by the lack of a test response in the distal fiber. Eventually, a point was reached during the cycle at which a premature response elicited in the proximal fiber propagated to the distal recording site (F). This signaled the end of the effective refractory period in the proximal fiber as described below. Action potentials with normal contour were obtained only after full recovery of the membrane potential to the maximum diastolic level. In some experiments graded responses were not observed and the earliest test response obtained was of relatively large magnitude (Fig. 4). Experiments of this type were utilized in studies of the effective refractory period presented below. In other experiments, as shown in Figure 5, early test stimulation resulted in a response which occurred only after a long latent period (Fig. 5A). This Circulation Research, Vol. XXVlll, January 1971 ATRIAL PLATEAU FIBERS 67 D Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 FIGURE 4 Determination of the effective refractory period of atrial plateau fibers. A: Trial of test stimulation given while the fiber was still refractory. B: When the test stimulus was positioned a few milliseconds later in the cycle a large magnitude response was obtained, signaling the end of the effective refractory period. C and D: Same phenomenon in a plateau fiber in another preparation. Graded responses were not observed in the fibers shown in this figure. Time spikes at 100-msec intervals. FIGURE 5 Premature responses elicited in an atrial plateau fiber. A: Effect of a test stimulus given during early phase 3. After a long latent period a nearly normal response occurred. As the test stimulus was applied progressively later during repolarization (B-E), the test responses occurred after shorter latent periods and initially were graded in nature. Time spikes at 100-msec intervals. See text for discussion. response occurred at a high level of membrane potential and had an almost normal contour. When test stimuli were applied later in the cycle (B-E), graded responses as described above were obtained after short latent periods. This result appears identical to the "paradoxical event" described by Kao and Hoffman (38) in their studies of graded activity in Purkinje fibers. Graded responses also could be obtained in plateau fibers being stimulated by propagated action potentials. In the experiment shown in Circulation Research, Vol. XXVlll, January 1971 Figure 6, both drive and test stimuli were administered to regular atrial tissue. One microelectrode was located in a regular fiber close to the stimulating electrode (lower trace). A second microelectrode was located in a plateau fiber at a distance (upper trace). Early test stimulation produced a largemagnitude action potential in the regular fiber, but no response occurred in the plateau fiber. During subsequent trials, the test stimulus was given later in the cycle and elicited small-amplitude depolarizations and 68 HOGAN, DAVIS Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 FIGURE « dHTanlL7T% eZh record X T T ye 1 t™, aPPhed \° rf m fibBT CmSed thB TegUhT PWduCed by «** a ^opagated action potential The Shown in th ° Attorn trace of fiber 2 1 7u u ^ ' "^ ^-magnitude action potential in the regular fiber but no response in the plateau fiber (top trace). As the test stimulus u>al alied progre^vely later in the cycle (B-E), the propagated potential of the regular Zr eTcHed p Ziz:zorwith progressivdy Urm mamitude in the ature test **•» ^- T>™SPt:t vuhe elicited the series of rapid re- * " finally action potentials with more normal features in the plateau fiber. It is evident that graded responses in plateau fibers can be elicited by propagated action potentials and thus are not a phenomenon produced only by electrical stimulation. One final observation made during these studies is of interest. In several preparations, a single premature stimulus resulted in more than one extra response. Examples of this effect are shown in Figure 7. Similar observations have been made in studies of ventricular muscle and Purkinje fibers (39). DURATION OF THE EFFECTIVE REFRACTORY PERIOD The effective refractory period (ERP) is defined as the shortest interval between two propagated action potentials (40). The ERP Circulation Research, Vol. XXVlll, January 1971 ATRIAL PLATEAU FIBERS 69 ±5 Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 Determination of the effective refractory period of regular atrial fibers. The upper action potential is from a regular atrial fiber close to the stimulating electrode. The lower action potential is from a regular fiber at a distance. The application of the test stimulus is indicated by the stimulus artifact seen during repoUtrization. A-E show graded premature responses in the proximal fiber as the test stimulus was moved progressively later in the cycle. The end of the effective refractory period for the proximal fiber was signaled by the appearance of the propagated response in the distal fiber. (F). Time marks at 100-msec intervals. 160 I 200 100 I 160 FIGURE 9 Determination of the ability of regular atrial fibers (A) and plateau fibers (B) to respond at rapid frequency. The numbers indicate the stimulus cycle length in milliseconds. The vertical black bars indicate a change in cycle length. Time spikes at 100-msec intervals. of both plateau and regular atrial fibers was measured in the present experiments. Forty experiments were performed on plateau fibers. Records from typical experiments are shown in Figures 3 and 4. The ERP of plateau fibers persisted until the membrane repolarized to — 62.1 ±0.8 (mean±SE) mv. The duration of the ERP in these experiments was 196.1 ± 2.5 msec. The ERP of regular atrial muscle fibers was determined in 14 experiments. The level of membrane potential needed in these fibers to obtain propagated action potentials was — 65.5 ±1.3 mv. Duration of the ERP for regular fibers was 146.8 ±5.4 msec. Records from an experiment with regular atrial fibers are shown in Figure 8. Statistical comparison of duration of effective refractory periods for the two fiber types showed a significant difference (P<0.005). Circulation Research, Vol. XXVlll, January 1971 There also was a small but significant difference (P<0.025) in the level of membrane potential required to give a propagated response in each type of fiber. Other observations indicate that the refractory period of atrial plateau fibers is longer than that of regular muscle fibers. In most hearts it was possible to obtain full-sized action potentials in regular atrial fibers. (Figs. 6A, 7D, and 7E) before any response was observed in plateau fibers. Also, when the ability of both types of fibers to respond to rapid frequencies of stimulation was determined, it was found that regular atrial fibers could respond at faster rates than plateau fibers. In 7 of 11 preparations, regular atrial muscle fibers responded to every stimulus given at 600/min. In six of eight similar experiments on plateau fibers, the fiber ceased to respond to every stimulus when HOGAN, DAVIS 70 the rate was less than 600/min. Records from representative experiments are shown in Figure 9. Discussion Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 Atrial plateau fibers possess several characteristics exhibited by specialized conducting and impulse-generating fibers. For example, in an earlier study (34) it was demonstrated that atrial plateau fibers have electrical features similar to ventricular Purkinje fibers. In particular, they exhibit a long plateau phase during repolarization and inherent slow diastolic depolarization. The maximum rate of depolarization during the upstroke of the action potential of plateau fibers is greater than in other fibers. Further, plateau fibers are relatively resistant to inactivation by elevated extracellular potassium. Additional evidence of the specialized nature of atrial plateau fibers is indicated by the results of the present study. Measurements of conduction velocity, supernormal excitability and refractoriness of plateau fibers favor this concept. It was suggested earlier that atrial plateau fibers, by virtue of their rapid upstroke velocity, might be capable of rapid impulse conduction between S-A and A-V nodes. Actual measurements of conduction velocity, as presently done, support this suggestion. The most rapid linear conducting pathway measured was along the caval border of the crista terminalis and was composed of plateau fibers. This finding further demonstrates that in addition to regular atrial muscle fibers the crista terminalis contains specialized conducting fibers. Their anatomical location corresponds to the bundle first described by Thorel (2, 3) and later by James (6-9) as the posterior intemodal tract. Rapid conduction along atrial plateau fibers correlates with the findings of several investivators (22, 24, 27, 29, 41, 42) that excitation passes more rapidly along the crista terminalis than through adjacent atrial muscle. Regarding the absolute values for conduction velocities reported in this study, it was shown that atrial plateau fibers conduct the impulse two to three times more rapidly than regular atrial fibers. This agrees with the recent findings of Holsinger et al. (43) who showed, using cardiac electrograms, the same relative difference between areas containing the intemodal tracts and areas composed predominantly of regular atrial fibers. It should be noted that although plateau fibers conduct at relatively fast rates they do not compare with the very fast rates reported for ventricular Purkinje fibers (44). The intemodal pathways in the atrium are shorter than the Purkinje network of the ventricles and may not require the rapid conduction velocity of the latter fibers to synchronize atrial contraction and to provide preferential activation of the atrioventricular node. Another electrical feature of atrial plateau fibers is a period of supernormal excitability. In the present study, it was shown that supernormal excitability exists in the plateau fibers of the free-running strand and crista terminalis. This phenomenon has also been observed in the specialized fibers of Bachmann's bundle (33) and in ventricular Purkinje fibers (45) and thus provides additional evidence for the presence of specialized Purkinje-like fibers in the canine right atrium. The effective refractory period of atrial plateau fibers was consistently longer than that of other fibers. This difference is due primarily to delayed repolarization caused by the plateau phase. It is not due to the difference in level of membrane potential required to obtain propagated responses because plateau fibers regained excitability at a lower level of membrane potential than regular fibers. This difference tends to reduce the disparity in length of refractory periods between the two fiber types. The measured limits of the refractory period are dependent upon the characteristics of the stimulus used to produce premature excitation (46). In this study, to ensure valid comparisons, stimuli with identical characteristics of duration and intensity were used for determinations on both types of fibers. Two additional observations substantiate the directly measured difference in refractoriness between the two fiber types. First, in several experiments, non-plateau Circulation Research, Vol. XXVIII, January 1971 71 ATRIAL PLATEAU FIBERS Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 fibers showed full-sized action potentials before any response was observed in plateau fibers. Second, in experiments in which the ability to respond to rapid frequencies of stimulation was determined, plateau fibers usually ceased to respond at lower stimulation rates than did other fibers. The latter type of experiment is very similar to that introduced by Dawes (47) to measure duration of the refractory period. We have shown by direct measurement that the canine right atrium contains two types of fibers with markedly different electrophysiological characteristics—in particular, with respect to duration of refractoriness. This raises the question of which type of fiber is being evaluated. If the stimulus was applied only to plateau fibers, the observed maximal rate of response would be slower and the duration of the refractory period longer than if it was applied to other fibers. For this reason, it seems reasonable to propose that the site of atrial stimulation should be the same in control and experimental determinations of refractoriness when this technique is used. The difference in refractoriness between plateau and regular fibers is similar qualitatively to that known to exist between Purkinje fibers and ventricular muscle fibers (37, 38). In tissue preparations containing both types of fibers, it usually is possible to obtain largemagnitude premature action potentials in ventricular fibers before similar responses can be obtained in Purkinje fibers (37-39). In addition, when excitation of a Purkinje fiber is produced by action potentials propagated from ventricular muscle, local responses are frequently produced in the Purkinje fiber. In the present study we observed similar behavior with plateau and regular atrial fibers. Such responses may be of significance in the genesis of atrial arrhythmias. Several observations made in this and previous studies may aid in understanding the origin of atrial arrhythmias. As noted above, atrial plateau fibers have several electrophysiological characteristics in common with Purkinje fibers. Some investigators believe that the origin of ventricular arrhythmias can be Circulation Research, Vol. XXVIII, January 1971 explained best in terms of changes in excitability and conductivity of the latter fibers. Detailed discussion of the evidence for this idea can be found in the reviews of others (48-50). We have shown previously (34) that plateau fibers have inherent slow diastolic depolarization which can be enhanced by catecholamines to the point of spontaneous discharge of action potentials. Therefore it is possible that in intact animals atrial plateau fibers are responsible for development of atrial ectopic foci. In the present study, it was shown that premature excitation of plateau fibers, whether caused by electrical stimuli or propagated action potentials, produced graded responses. Such responses propagate slowly and decrementally (37, 38), and it is believed that the resulting delay or block of conduction is favorable for circus movement and reentry of excitation (48-50). A final point regarding the possible role of plateau fibers in atrial arrhythmias is based on the assumption that these fibers compose the posterior internodal tract. Sherf and James (51) considered the function of the three internodal tracts in transmission of excitation to the A-V node. They postulated that excitation would reach the A-V node first over the anterior or middle tracts or both rather than over the posterior tract. This is logical in view of the relative length of the different tracts and the likelihood that conduction velocity along each tract is similar (43). They postulated that in the normal situation excitation might pass retrograde for a distance along the posterior tract. One could speculate that in normal hearts the long refractory period of atrial plateau fibers would insure that excitation does not ascend any great distance over the posterior tract. If the situation should arise that conduction velocity over the posterior tract increased concomitantly with a reduction of duration of refractoriness, the entire pathway might become responsive to retrograde excitation. James (6) has called attention to the fact that the internodal tracts form a circuitous pathway around the orifices of the venae cavae. Movement of the impulse around such a circuit long has been proposed to 72 HOGAN, DAVIS Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 explain the atrial arrhythmias of flutter and fibrillation (46). One condition which may produce the changes in conduction velocity and refractoriness stated above is vagal stimulation or application of acetylcholine. This agent hyperpolarizes the membrane, increases the overshoot, and markedly shortens duration of the action potential of plateau fibers (34). The tendency of atrial arrhythmias to occur during parasympathetic activity or application of acetylcholine is well known (46). We recognize that these proposals are tentative but believe them to be consistent with the current state of knowledge concerning structure and function of the internodal tracts. References 1. WENCKENBACH, K.F.: Beitriige zur Kenntnis der menschlichen Herztiitigkeit. Arch Physiol 1907, pp 1-24. 2. THOREL, C : Vorlaufige Mitteilung iiber eine besondere Muskelverbindung zwischen der Cava superior und dem Hisschen Biindel (abstr). Munchen Med Wschr 56:2159, 1909. 3. THOREL, C : Ueber den Aufbau des Sinusknotens und seine Verbindung mit der Cava superior und den Weckenbachschen Bundeln. Munchen Med Wschr 57:183-186, 1910. 4. BACHMANN, G.: Inter-auricular time interval. Amer J Physiol 41:309-320, 1916. 5. BACHMANN, G.: The distribution of the vagus nerves to the sinoauricular junction of the mammalian heart. Amer J Physiol 63:300-337, 1922. 6. JAMES, T.N.: The connecting pathways between the sinus node and the AV node and between the right and left atrium in the human heart. Amer Heart J 66:498-508, 1963. 7. JAMES, T.N.: Anatomy of the AV node of the dog. Anat Rec 148:15-27, 1964. 8. JAMES, T.N.: Anatomy of the sinus node, AV node and os cordis of the beef heart. Anat Rec 153:361-371, 1965. 9. JAMES, T.N.: Anatomy of the cardiac conduction system in the rabbit. Cir Res 20:638-648, 1967. 10. TAKAVASU, M., TAKASAKI, H., NISHH, N., TATEISHI, Y., OSAWA, M., TAMAGAKI, A., FUJIWABA, M., SHINAGAWA, H., AND IKUTA, S.: Studies on the atrial stimulus conduction. Jap Circ J 18:414-420, 1955. 11. TAKAYASU, M.: Conduction of excitation from the sinus node to the atrioventricular node. Jap Circ J 31:373-380, 1967. 12. ROBB, J.S., AND PETRI, R.: Expansions of the atrioventricular system in the atria. In The Specialized Tissues of the Heart, edited by A. Paes de Carvalho, W. C. de Mello, and B. F. Hoffman, Amsterdam, Elsevier Publishing Company, 1961, pp 1-16. 13. MEREDITH, J., AND TITUS, J.L.: Anatomic atrial connections between sinus and A-V node. Circulation 37:566-579, 1968. 14. EMBERSON, J.W., AND CHALLICE, C.E.: Studies on the impulse conducting pathways in the atrium of the mammalian heart. Amer Heart J 79:653-667, 1970. 15. EYSTER, J.A.E., AND MEEK, W.J.: Experiments on the origin and propagation of the impulse in the heart: The point of primary negativity in the mammalian heart and the spread of negativity to other regions. Heart 5:119-135, 1913-1914. 16. EYSTER, J. A. E., AND MEEK, W. J.: Experiments on the origin and conduction of the cardiac impulse: VI. Conduction of the excitation from the sinoauricular node to the right auricle and auriculoventricular node. Arch Intern Med 18: 775-799, 1916. 17. EYSTER, J.A.E., AND MEEK, W.J.: Origin and conduction of the heart beat. Physiol Rev 1: 1-43, 1921. 18. EYSTER, J.A.E., AND MEEK, W.J.: Studies on the origin and conduction of the cardiac impulse: VIII. The permanent rhythm following destruction of the sinoauricular node. Amer J Physiol 61:117-129, 1922. 19. EYSTER, J.A.E., AND MEEK, W.J.: Experiments on the origin and conduction of the heart beat: IX. Sinoventricular conduction. Amer J Physiol 61:130-137, 1922. 20. LEWIS, T., MEAKINS, J., AND WHITE, P.D.: Excitatory process in the dog's heart: I. The auricles. Phil Trans Roy Soc London, Ser B 205:375-420, 1914. 21. LEWIS, T.: Mechanism and Graphic Registration of the Heart Beat, 3d ed. London, Shaw, 1925, p87. 22. 23. 24. 25. 26. PAES D E CARVALHO, A., D E MELLO, W.C., AND HOFFMAN, B.F.: Electrophysiological evidence for specialized fiber types in rabbit atrium. Amer J Physiol 196:483-488, 1959. TRAUTWEIN, W.W.: Elektrophysiologie der Herzmuskelfaser. Ergebn Physiol 51:131-198, 1961. HORIBE, H.: Studies on the spread of the right atrial activation by means of intracellular microelectrodes. Jap Circ J 25:583-593, 1961. MIYAUCHI, A.: Electrical events in specialized muscle fibers of a mammalian right atrium. Jap Heart J 3:357-372, 1962. HORIBA, M.: Stimulus conduction in atria studied by means of intracellular microelectrodes: I. That in Bachmann's bundle. Jap Heart J 4: 333-345, 1965. 27. SANO, T., AND YAMACISHI, S.: Circulation Research, Vol. XXVlll, Spread of January 1971 73 ATRIAL PLATEAU FIBERS excitation from the sinus node. Circ Res 16: 423-430, 1965. 28. WAGNER, M.L., LAZZARA, R., WEISS, R.M, AND HOFFMAN', B.F.: Specialized conducting fibers in the interatrial band. Cir Res 18:502-518, 1965. 29. YAMADA, K., HORIBA, M., SAKAIDA, Y., OKAJIMA, M., HORIBE, H., KOBAYASHI, T., MlYAUCHI, A., OlSHI. H., NONCAWA, A., ISHIKAWA, K., Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 AND TOYAMA, J.: Origination and transmission of impulses in the right auricle. Jap Heart J 6: 71-97, 1965. 30. MOORE, E.N.: Atrioventricular transmission in newborn calves. In Comparative Cardiology, edited by H. H. Hecht and D. K. Detweiler. Ann NY Acad Sci 127:113-126, 1965. 31. DAVIS, L.D., TEMTE, J.V., HELMER, P.R., AND MURPHY, Q.R.: Effects of cyclopropane and hypoxia on transmembrane potentials of atrial, ventricular, and Purkinje fibers. Circ Res 18: 692-704, 1966. 32. MOORE, E.N.: Phylogenetic observations on specialized cardiac tissue. Bull NY Acad Med 43:1138-1159, 1967. 33. CHILDERS, R.W., MERIDETH, J., AND MOE, G.K.: Supernormality in Bachman's bundle. Circ Res 22:363-370, 1968. 34. HOGAN, P.M., AND DAVIS, L.D.: Evidence for specialized fibers in the canine right atrium. Circ Res 23:387-396, 1968. 35. STEEL, R.G.D., AND TORRIE, J.H.: Principles and Procedures of Statistics. New York, McGraw-Hill Book Co, 1960, p 107. 36. MOORE, E.N, PRESTON, J.B., AND MOE, G.K.: Durations of transmembrane action potentials and functional refractory periods of canine false tendon and ventricular myocardium: Comparisons in single fibers. Circ Res 17: 259-273, 1965. 37. HOFFMAN, B.F., KAO, C.Y, AND SUCKLINC, E. E.: Refractoriness in cardiac muscle. Amer J Physiol 190:473-482, 1957. 38. KAO, C.Y., AND HOFFMAN, B.F.: Graded and web, Vol. XXVIII, January 1971 decremental responses in heart muscle fibers. Amer J Physiol 194:187-196, 1958. 39. DAVIS, L.D., AND TEMTE, J.V.: Electrophysio- logical actions of lidocaine on canine ventricular muscle and Purkinje fibers. Circ Res 24: 639-655, 1969. 40. Danax, A.N., AND LOVE, W.S.: Supposed lengthening of the absolute refractory period of frog's ventricular muscle by veratrine. Heart 13:77-85, 1926. 41. PUECH, P., ESCLAVISSAT, M., SODI-PAIXARES, D., AND CISNEROS, F.: Normal auricular activation in the dog's heart. Amer Heart J 47:174-191, 1954. 42. Oism, H.: Manner of stimulus conduction in atria. Jap Heart J 8:276-290, 1967. 43. HOLSINCER, J.W., WALLACE, A.G., AND SEALY, W.C.: Identification and surgical significance of the atrial internodal conduction tracts. Ann Surg 167:447-453, 1968. 44. DRAPER, M.H., AND WEIDMANN, S.: Cardiac resting and action potentials recorded with an intracellular electrode. J Physiol (London) 115:74-94, 1951. 45. WEIDMANN, S.: Effects of calcium ions and local anesthetics on electrical properties of Purkinje fibers. J Physiol (London) 129:568-582, 1955. 46. BROOKS, C . M C . C , HOFFMAN, B.F., SUCKLING, E.E, AND ORIAS, O.: Excitability of the Heart. New York, Grune & Stratton, Inc., 1955, pp 72, 148, and 205. 47. DAWES, G.S.: Synthetic substitutes for quinidine. Brit J Pharmacol 1:90-112, 1946. 48. HOFFMAN, B.F.: Genesis of cardiac arrhythmias. Progr Cardiovasc Dis 8:319-329, 1966. 49. WATANABE, Y, AND DREIFUS, L.S.: Newer con- cepts in the genesis of cardiac arrhythmias. Amer Heart J 76:114-135, 1968. 50. HAN, J.: Mechanisms of ventricular arrhythmias associated with myocardial infarction. Amer J Cardiol 24:800-813, 1969. 51. SHERF, L., AND JAMES, T.N.: New electrocardio- graphic concept: Synchronized sinoventricular conduction. Dis Chest 55:127-140, 1969. Electrophysiological Characteristics of Canine Atrial Plateau Fibers PERRY M. HOGAN and LARRY D. DAVIS Downloaded from http://circres.ahajournals.org/ by guest on June 18, 2017 Circ Res. 1971;28:62-73 doi: 10.1161/01.RES.28.1.62 Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Copyright © 1971 American Heart Association, Inc. All rights reserved. Print ISSN: 0009-7330. Online ISSN: 1524-4571 The online version of this article, along with updated information and services, is located on the World Wide Web at: http://circres.ahajournals.org/content/28/1/62 Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in the Permissions and Rights Question and Answer document. Reprints: Information about reprints can be found online at: http://www.lww.com/reprints Subscriptions: Information about subscribing to Circulation Research is online at: http://circres.ahajournals.org//subscriptions/