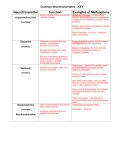
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
J Neurophysiol 93: 3029 –3031, 2005; doi:10.1152/classicessays.00026.2005. Editorial Focus ESSAYS ON APS CLASSIC PAPERS Synaptic chemistry in single neurons: GABA is identified as an inhibitory neurotransmitter Ronald Harris-Warrick Department of Neurobiology and Behavior, Cornell University, Ithaca, New York TODAY WE KNOW that there are many different neurotransmitters, each interacting with its spectrum of receptors to evoke a variety of different responses in neurons. However, in the 1950s, the idea that neurotransmission was primarily of a chemical rather than electrical nature was still a topic for controversy, at least in the United States. Only two neurotransmitters had been identified: acetylcholine as the transmitter of parasympathetic and preganglionic sympathetic neurons and spinal motoneurons, and norepinephrine as the transmitter of sympathetic neurons. Although it was understood that there are inhibitory as well as excitatory synapses, little was known of what compounds, or how many of them, might mediate inhibitory transmission; in addition to its inhibitory effect on the heart, acetylcholine appeared not to be involved. This problem was solved for the first time by Edward Kravitz (Fig. 1) and his colleagues in Stephen Kuffler’s group at Harvard Medical School: in an innovative series of biochemical and physiological experiments, they proved that ␥-aminobutyric acid, or GABA, is the inhibitory neurotransmitter at the lobster neuromuscular junction. Several of the crucial papers were published in the Journal of Neurophysiology, and two of them are discussed in this commentary. GABA was identified in the brain in 1950. Its function was unknown, but it was hypothesized to be involved in metabolism. Interest in GABA as an inhibitory neurotransmitter was initiated by Bazemore, Elliott, and Florey, who isolated inhibitory substances from brain that could block the spontaneous spike discharge of the crayfish stretch receptor preparation. Starting with 52 kg of beef brain, they were able to isolate an active extract that was called Factor I (1, 2). On further fractionation, Florey and colleagues found that the major active substance in Factor I was GABA, which had much higher Address for reprint requests and other correspondence: R. Harris-Warrick, Dept. of Neurobiology and Behavior, Cornell Univ., Ithaca, NY 14853 (E-mail: [email protected]). http://www.the-aps.org/publications/classics specific activity than any other compound. They cautiously concluded, “It thus seems probable that ␥-aminobutyric acid is concerned in some aspects of the regulation of physiological activity in the brain and other nervous structures. It is possibly a transmitter substance of inhibitory impulses. . .” (2). However, further work convinced Florey that he was wrong about GABA. Attempts to purify GABA or its synthetic enzyme from crustacean muscles or nerves were apparently unsuccessful. Furthermore, GABA failed to mimic the strychnine-sensitive inhibitory substance in the mammalian spinal cord (which we now know to be glycine). In an important review in 1961, Florey (5) concluded, “There is sufficient evidence that GABA is not the inhibitory transmitter, though it imitates the transmitter action in many crustacean preparations.” Mammalian neurophysiologists, including David Curtis and Hugh McClellan, had trouble mimicking the endogenous inhibitory transmitter effects with GABA, and at a major FIG. 1. Edward Kravitz. 0022-3077/05 $8.00 Copyright © 2005 The American Physiological Society 3029 Downloaded from http://jn.physiology.org/ by 10.220.33.4 on June 17, 2017 This essay looks at the historical significance of two APS classic papers that are freely available online: Kravitz EA, Kuffler SW, and Potter DD. Gamma-aminobutyric acid and other blocking compounds in Crustacea. III. Their relative concentrations in separated motor and inhibitory axons. J Neurophysiol 26: 739 –751, 1963 (http:// jn.physiology.org/cgi/reprint/26/5/739). Otsuka M, Kravitz EA, and Potter DD. Physiological and chemical architecture of a lobster ganglion with particular reference to gamma-aminobutyrate and glutamate. J Neurophysiol 30: 725–752, 1967 (http://jn.physiology.org/cgi/reprint/ 30/4/725). Editorial Focus 3030 2. David Potter. conference in 1959, most scientists concluded that GABA was probably not an inhibitory transmitter (16). A number of neurophysiologists studying the actions of GABA at the crustacean neuromuscular junction were convinced that this conclusion was premature. These workers took advantage of the extreme simplicity of muscle innervation in lobster and crayfish walking legs to further study the actions of GABA. The opener of the dactyl, for example, is only innervated by two axons, one excitatory and one inhibitory. These can be separately stimulated, allowing a pure excitatory or a pure inhibitory postsynaptic response to be detected and analyzed. Kuffler and Edwards (13) and Boistel and Fatt (3) showed that GABA mimics the action of the natural inhibitory transmitter at the crustacean stretch receptor and neuromuscular junction, causing little change in membrane potential but a significant decrease in input resistance. Both substances had the same reversal potential, which was due to a selective increase in chloride permeability. Stephen Kuffler, at Harvard, decided to organize a group to study the neurotransmitters used at the crustacean neuromuscular junction. Kuffler was lucky to recruit a talented electrophysiologist, David Potter (Fig. 2), and a talented biochemist, Edward Kravitz, who was then a postdoctoral fellow at the National Institutes of Health (NIH) studying the metabolism of opium alkaloids. This team was finally able to solve the GABA problem. Four criteria had to be met before the compound could be confirmed as a neurotransmitter. First, it had to mimic the physiological and pharmacological properties of the natural transmitter. This criterion had already been met for GABA at the crustacean neuromuscular junction (3). Second, the compound had to be present in the presynaptic neuron. This was a very tough assignment in the days before immunocytochemistry, and it was Kravitz’s job as the team biochemist to develop the microchemical techniques to meet this criterion. First, they showed that GABA is present in extracts of crayfish muscles and nerves (7). In a careful screen of compounds found in the central and the peripheral nervous system of lobsters, about ten compounds with blocking activity at the neuromuscular junction (reducing the amplitude of evoked excitatory junctional potentials) were isolated by water extraction, ion exchange columns, paper chromatography, and electrophoresis (4, 9). J Neurophysiol • VOL GABA had by far the highest specific activity (more than 50 times higher than the next most active compound, -alanine), but it only accounted for about 20% of the total blocking activity. Moreover, GABA was selectively concentrated in the small bundles that only had the excitatory and inhibitory axons. Kravitz decided that the only way to determine where GABA was located was to separate the single excitatory and inhibitory axons that innervate the dactyl of the walking leg and compare the concentrations of the blocking agents in each axon. This was a technical tour de force, described in the first paper featured in this commentary (8). Kravitz and colleagues carefully dissected the two axons apart from several joints of the walking legs of several hundred lobsters. In all, 5 meters of single axons were isolated for analysis. GABA was rapidly isolated using ion exchange columns and electrophoresis. To measure the tiny amounts of GABA in single axons, Kravitz developed two new and more sensitive assays for GABA. The first was based on the enzymatic conversion of GABA to succinate by the enzymes GABA-glutamic acid transaminase and succinic semialdehyde dehydrogenase, reducing NADP to NADPH in the process, which could be measured spectrophotometrically. The enzymes were isolated from a strain of Pseudomonas fluorescens that can grow on GABA alone as an energy source, which Kravitz had carried in his pocket to Harvard from the NIH. When there was insufficient GABA for this assay (1 nmol), a more sensitive fluorometric assay was developed where the newly synthesized NADPH was isolated and reconverted back to NADP for measurement; this method was sensitive down to 10 pmol. The very first experiments were startling: significant amounts of GABA were found in the inhibitory axons, whereas none was detectable from the excitatory axons. Further experiments showed that there is indeed a small amount of GABA (with still unknown function) in the excitatory motoneurons, but the ratio of GABA in the inhibitory to excitatory axons was about 100:1, with about 0.1 M GABA in the inhibitory axons (11). In another tour de force of single cell biochemistry, described in the second paper in this commentary (15), Otsuka, Kravitz, and Potter proceeded to compare levels of GABA in single identified neurons in lobster abdominal ganglia. Again, the lobster proved to be an ideal choice for these studies. The abdominal ganglion contains a number of very large neurons in stereotyped positions; these neurons do not have any synapses on their somata, so the isolated soma was not contaminated by transmitters in adherent presynaptic terminals. Neurophysiological studies were carried out to identify 21 pairs of neurons that innervate peripheral neuromusclar junctions: three pairs were inhibitory, innervating different muscles, whereas the rest were excitatory motoneurons. Kravitz and his colleagues developed a novel method to isolate these single neurons and assay their levels of GABA using the sensitive fluorometric assay. Almost none of the excitatory motoneurons had any detectable GABA by this assay, whereas all of the inhibitory neurons tested contained GABA. This was the first study of the chemical architecture of a region of the nervous system and stands as a model for studies that followed. Although these papers unambiguously showed that GABA is highly concentrated in inhibitory neurons compared with excitatory neurons, Kravitz and colleagues were not satisfied that they had proven that GABA is the inhibitory neurotransmitter of these neurons. Two more criteria for identification of a 93 • JUNE 2005 • www.jn.org Downloaded from http://jn.physiology.org/ by 10.220.33.4 on June 17, 2017 FIG. ESSAYS ON APS CLASSIC PAPERS Editorial Focus ESSAYS ON APS CLASSIC PAPERS J Neurophysiol • VOL elsewhere. Invertebrates have a long and successful record for major discoveries in neuroscience (for example, the ionic basis of the action potential, organization of small motor networks, mechanisms of neuromodulation, and the cellular basis of learning and memory), and they will continue to serve this function into the future. REFERENCES 1. Bazemore A, Elliott KAC, and Florey E. Factor I and ␥-aminobutyric acid. Nature 178: 1052–1053, 1956. 2. Bazemore AW, Elliott KAC, and Florey E. Isolation of Factor I. J Neurochem 1: 334 –339, 1957. 3. Boistel J and Fatt P. Membrane permeability change during inhibitory transmitter action in crustacean muscle. J Physiol 144: 176 –191, 1958. 4. Dudel J, Gryder R, Kaji A, Kuffler SW, and Potter DD. Gammaaminobutyric acid and other blocking compounds in Crustacea. I. Central nervous system. J Neurophysiol 26: 721–728, 1963. 5. Florey E. Comparative physiology: transmitter substances. Annu Rev Physiol 23: 501–528, 1961. 6. Hall ZW, Bownds MD, and Kravitz EA. The metabolism of gamma aminobutyric acid in the lobster nervous system. J Cell Biol 46: 290 –299, 1970. 7. Kravitz EA. Enzymic formation of gamma-aminobutyric acid in the peripheral and central nervous system of lobsters. J Neurochem 9: 363– 370, 1962. 8. Kravitz EA, Kuffler SW, and Potter DD. Gamma-aminobutyric acid and other blocking compounds in Crustacea. III. Their relative concentrations in separated motor and inhibitory axons. J Neurophysiol 26: 739 –751, 1963. 9. Kravitz EA, Kuffler SW, Potter DD, and Van Gelder NM. Gammaaminobutyric acid and other blocking compounds in Crustacea. II. Peripheral nervous system. J Neurophysiol 26: 729 –738, 1963. 10. Kravitz EA, Molinoff PB, and Hall ZW. A comparison of the enzymes and substrates of gamma-aminobutyric acid metabolism in lobster excitatory and inhibitory axons. Proc Natl Acad Sci USA 54: 778 –782, 1965. 11. Kravitz EA and Potter DD. A further study of the distribution of ␥-aminobutyric acid between excitatory and inhibitory axons of the lobster. J Neurochem 12: 323–328, 1965. 12. Kravitz EA, Potter DD, and Van Gelder NM. Gamma-aminobutyric acid and other blocking substances extracted from crab muscle. Nature 194: 382–383, 1962. 13. Kuffler S and Edwards C. Mechanism of gamma-aminobutyric acid (GABA) action and its relation to synaptic inhibition. J Neurophysiol 21: 589 – 610, 1958. 14. Otsuka M, Iversen LL, Hall ZW, and Kravitz EA. Release of gammaaminobutyric acid from inhibitory nerves of lobster. Proc Natl Acad Sci USA 56: 1110 –1115, 1966. 15. Otsuka M, Kravitz EA, and Potter DD. Physiological and chemical architecture of a lobster ganglion with particular reference to gammaaminobutyrate and glutamate. J Neurophysiol 30: 725–752, 1967. 16. Roberts E and Baxter DF. Inhibition in the Nervous System and GammaAminobutyric Acid. New York: Pergamon, 1960. 93 • JUNE 2005 • www.jn.org Downloaded from http://jn.physiology.org/ by 10.220.33.4 on June 17, 2017 transmitter remained to be met. The third criterion was to show that the transmitter is synthesized in the presynaptic neuron. This criterion was met by demonstrating the enzymatic synthesis of GABA from radioactive glutamate by glutamic acid decarboxylase (GAD), first in whole central or peripheral nervous tissue (7) and then in single excitatory and inhibitory axons (6, 7). These experiments demonstrated that the GAD concentration in inhibitory axons is at least 100 times higher than in the excitatory axons. The metabolic enzymes that break down GABA were found in similar amounts in the two nerves. The fourth and final criterion was to show that GABA is released from the inhibitory nerve terminal on stimulation. Again taking advantage of the simplicity of the lobster neuromuscular junction, Otsuka et al. (14) superfused opener muscles in the large crusher claw of lobsters while stimulating the single excitatory and inhibitory nerves innervating the muscle. GABA was isolated from the saline superfusing the muscle using ion exchange columns and was measured using the enzyme assay. On stimulation of the inhibitory axon, significant amounts of GABA were released, and the amount rose with the number of stimuli. Release was calcium dependent, as expected for a neurotransmitter. In contrast, no detectable GABA was released when the excitatory motoneuron was selectively stimulated. In further work, Kravitz and his students characterized all the enzymes in the GABA synthetic and metabolic pathway and finally showed that there is a specific reuptake mechanism for GABA that could serve to inactivate the released GABA. Throughout this work, Kuffler, Kravitz, and their colleagues were very careful not to overstate their case. It was only after all of these results were obtained that Kravitz finally wrote, without any qualifications, “␥-aminobutyric acid (GABA) is the inhibitory transmitter compound at the lobster neuromuscular junction.” (6). These experiments proved conclusively for the first time that GABA is an inhibitory neurotransmitter. As the results obtained by Kravitz and his very able colleagues (Otsuka, Iversen, Hall, Potter) supporting this accumulated, they were for the most part ignored by vertebrate neuroscientists. It was only a decade later that similar experiments were possible in the vertebrate central nervous system, verifying that GABA is indeed the most common inhibitory neurotransmitter in the brain, although certainly not the only one. Taking advantage of the numerical simplicity of the invertebrate nervous system, Kravitz and his collaborators were able to demonstrate a general principle of neuronal function that could not be proven 3031