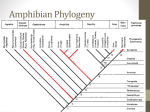
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
CHAPTER 13 Life Histories Richard C. Bruce 13.1 INTRODUCTION The life history of an animal is the sequence of morphogenetic stages from fertilization of the egg to senescence and death, which incorporates the probabilistic distributions of demographic parameters of an individual’s population as components of the life-history phenotype. At the population (or species) level, and in the context of ectothermic vertebrates, Dunham et al. (1989) have defined a life history in terms of a heritable set of rules that govern three categories of allocations: (1) allocation of time among such activities as feeding, mating, defense, and migration; (2) allocation of assimilated resources among growth, storage, maintenance, and reproduction; and (3) the mode of “packaging” of the reproductive allocation. An application of such an allocation-based definition in the study of salamander life histories was provided by Bernardo (1994). Implicit in the definition is the condition that life-history traits have a heritable basis; average plasticity in any trait reflects the average reaction norm, or the range in phenotypes expressed by a given genotype, averaged for all genotypes, over the range of environments experienced by all members of the population (Via 1993). Salamanders show greater diversity in life histories than any other vertebrate taxon of equivalent rank, which is all the more remarkable given the relatively small number (about 500) of known extant species. Beginning, somewhat arbitrarily, with the landmark studies of the plethodontids Desmognathus fuscus and Eurycea bislineata by Inez W. Wilder (1913, 1924) [who earlier published papers on salamanders under the name I. L. Whipple], this diversity has generated considerable interest by ecologists and herpetologists during the past 90 years. Many salamanders undergo a biphasic life cycle that includes a complex metamorphosis from an aquatic larva to a more terrestrial juvenile that ultimately matures to an adult. It is likely that this is the ancestral life cycle, given current knowledge of phylogenetic relationships within the Department of Biology, Western Carolina University, Cullowhee, NC 28723 USA "%& Reproductive Biology and Phylogeny of Urodela Urodela, as described by Larson et al. in chapter 2 of this volume, and among the three extant amphibian lineages (Zardoya and Meyer 2001). However, in numerous species in several independent lineages, the aquatic larva achieves reproductive maturity directly, without metamorphosis, a condition known as paedomorphosis. Within a species paedomorphosis may be facultative or obligatory. Moreover, paedomorphosis may involve the entire suite of somatic tissues and organs, or may be dissociative, i.e., partial. Conversely, many species of salamanders, particularly in the Plethodontidae, lack the aquatic larval form, and the eggs hatch as tiny versions of the adults, a condition known as direct development. Moreover, within a small clade of viviparous members of the Salamandridae, direct development is associated with the most unique life histories of any salamanders. Thus, assuming that extant salamanders represent a monophyletic lineage, the range of variation in living representatives is evidence of rampant evolution at the level of the life cycle in this taxon. Paedomorphosis, as a derivative of the biphasic life cycle that involves retention of the full suite of larval somatic characters, is usually considered an evolutionary blind alley in salamanders, in contrast to dissociative paedomorphosis in direct-developing forms (i.e., plethodontines and bolitoglossines among the plethodontids) that has provided evolutionary opportunities for adaptation to a wide range of terrestrial environments (Wake 1966, 1991; Wake and Hanken 1996). Carroll (1988) cited direct development in these plethodontids as models of the reproductive mode from which the amniote condition evolved (though the plethodontine/bolitoglossine lineages are themselves constrained by lunglessness). The model is supported by more recent studies on the relationship between egg size and several features of embryogenesis in these taxa (Collazo and Marks 1994; Collazo 1996). 13.2 HETEROCHRONY The variation in morphogenesis leading to life-cycle diversification in salamanders can be assessed under the general heading of heterochrony, a term that refers to evolutionary changes in the rate or timing of ontogenetic events. Following de Beer (1930), Gould (1977, 2000) defined heterochrony as “phyletic change in the onset or timing of development, so that the appearance or rate of development of a feature in a descendant ontogeny is either accelerated or retarded relative to the appearance or rate of development of the same feature in an ancestor’s ontogeny.” In this context, heterochrony is considered a macroevolutionary rather than a microevolutionary process. This distinction is often blurred in the literature, as Reilly et al. (1997) have pointed out. The first important exposition of the importance of heterochrony in macroevolution was provided by de Beer (1930, 1940), who utilized heterochronic arguments to refute the logical structure of Haeckel’s biogenetic law. Gould (1977) revised and simplified de Beer’s scheme, and reduced the processes involved to two basic categories – acceleration and retardation in Life Histories "%' the rate of development of either a whole organism or a particular subset of its organ systems. Gould argued that acceleration and retardation affect the reproductive and non-reproductive tissues and organs differently, such that he recognized four main categories of heterochrony, which he summarized conveniently in tabular form (Gould 1977: Table 3, p. 229). I have reorganized this table and substituted some names (Table 13.1). In this format it represents a convenient model (“the salamander model”) for considering the evolution of life-cycle variation in salamanders, under the assumption that the biphasic life cycle is ancestral. The conditions named in the body of the table are considered evolutionary results of heterochrony, achieved by the processes of acceleration and retardation. It is seen that paedomorphosis may be attained by two processes, acceleration in the development of reproductive organs, or progenesis, and retardation in the rate of development of the somatic organs, or neoteny. Acceleration in the rate of development of the somatic organs involves shortening of the larval period, and may lead to elimination of a freeliving larval stage from the life cycle, a condition known as direct development. The fourth category, hypermorphosis, is attained by retardation in the rate of development to sexual maturation, leading to a prolongation of the juvenile phase. The expected result of hypermorphosis in salamanders is attainment of larger body sizes, given that growth rates slow in these animals following maturation. Direct development and hypermorphosis are two aspects of peramorphosis, a term denoting derived ontogenies wherein morphogenesis extends the ancestral condition (Alberch et al. 1979). Table 13.1 The four categories of heterochrony that apply to organisms with biphasic life cycles. Special terms are available for the processes of acceleration in development of the reproductive tissues (progenesis) and retardation of development of the somatic tissues (neoteny). Based on Gould (1977: Table 3). See also Gould (2000) for clarification of the terminology. Tissues Æ Process Ø acceleration retardation somatic direct development neoteny Æ paedomorphosis reproductive progenesis Æ paedomorphosis hypermorphosis Later, Gould, with others, modified this theoretical construct by expanding the definition of heterochrony to include changes in timing as well as rate (Alberch et al. 1979). Changes in timing of growth or morphogenesis can affect either the initiation of the process (onset time) or its termination (offset time). Regarding the former, Alberch et al. recognized predisplacement as earlier onset and postdisplacement as later onset, the former leading to peramorphosis and the latter to paedomorphosis. In reference to the salamander model, preand postdisplacement in age at metamorphosis yield the same results as acceleration (Æ direct development) and neoteny (Æ paedomorphosis). "& Reproductive Biology and Phylogeny of Urodela Offset time is more complex, and involves the relationship between sexual maturation and growth. Although growth is indeterminate in salamanders, there is a tendency for growth to slow following maturation, such that the highest growth rates are attained in the larval and juvenile phases. Thus, acceleration in the rate of development to maturation, or progenesis, may be associated with an earlier offset time of growth; conversely, retardation of maturation, resulting in hypermorphosis, may be correlated with a delayed offset of growth. These relationships bring the salamander model into conformity with the formalistic structure of the heterochronic model of Alberch et al. (1979: Table 1) (see also McKinney and McNamara 1991: Fig. 2-3; Reilly et al. 1997: Fig. 1). Given that the salamander model concerns the entire life history, is dissociative only at the level of somatic versus reproductive organs, and is age-structured, it can be evaluated primarily in terms of the four rate phenomena provided in Table 1. The special case of differential metamorphosis in cryptobranchids, amphiumids, and a few other taxa is considered below. The life-history aspects of dissociative heterochrony and ontogenetic repatterning in direct-developing plethodontids are largely beyond the scope of this chapter (but see section 13.3.9 Direct Development). Reilly et al. (1997), in criticizing many aspects of the Alberch et al. (1979) approach, substituted several newer terms for the traditional categories and emphasized the need to differentiate between heterochrony at the interspecific (the original meaning) versus the intraspecific level. These authors introduced a new set of terms for intraspecific heterochrony. Although they noted that heterochrony applies to individual traits rather than the whole organism, it is useful in the context of salamander life histories to recognize dissociative heterochrony of the somatic and reproductive organs in their entirety; after all, metamorphosis involves a suite of traits across several organ systems, and promotes a niche shift from one environment to another, whereas sexual maturation affects not only the reproductive system but components of other organ systems that contribute the secondary sexual characters. The terms neoteny and progenesis, which have had such wide usage in amphibian biology, were targeted by Reilly et al. (1997) for special criticism. They recommended discarding neoteny, in favor of deceleration, mainly because of continuing inconsistency in usage. This seems a minor problem. They leveled similar criticism at the term progenesis, in particular the application of the term by various authors either to reproductive acceleration only, or to reproductive and somatic acceleration conjointly. They suggested substitution of the term hypomorphosis for the latter process. Reilly et al. noted that some authors, under the first definition, have assumed that reproductive acceleration is accompanied by continuing somatic development such that “normal terminal shape” is attained. Given that Gould (1977) provided clear definitions of both neoteny and progenesis in his Table 3, and that, in the case of the latter, some term is needed in amphibians for accelerated maturation independent of somatic acceleration, I retain both neoteny and progenesis (Table 1), as defined by Gould. The importance of retaining the terminology Life Histories "& lies in the observation that the landmark events of somatic metamorphosis and reproductive maturation may evolve independently in amphibians, as documented below. Thus, for the purposes of this analysis, the heterochronic categories of Gould, in a slightly modified format (Table 1), based on rate changes, and subsuming shifts in timing, provide a straightforward conceptual framework for evaluating phylogenetic diversification of urodelan life histories. A different category of evolutionary change in ontogeny is that of caenogenesis, defined as the interposition into the life cycle of specialized larval adaptations that have no effect on adult organization, and thus are technically non-heterochronic (Gould 1977). Caenogenesis has been influential in larval evolution in anurans (Orton 1953), but has played a far lesser role in salamanders (Wake and Hanken 1996). 13.3 VARIATION IN MAJOR FEATURES OF THE URODELAN LIFE HISTORY 13.3.1 Fertilization External fertilization in aquatic habitats is characteristic of the primitive families Cryptobranchidae, Hynobiidae, and Sirenidae. In Cryptobranchus alleganiensis the female deposits strings of eggs that are fertilized by the male as he moves beside or above her in the nest (Smith 1907). Communal spawning is known in the Japanese cryptobranchid, Andrias japonicus (Kawamichi and Ueda 1998). Pond-dwelling hynobiid females release their eggs in a pair of egg sacs, into which the male releases sperm; however, in one species of stream-dwelling hynobiid, Ranodon sibiricus, the male produces a large, stalkless spermatophore, to which a female or females attach their egg sacs (Nussbaum 1985). Although mating has not been observed in sirenids, histological properties of the oviduct of Siren intermedia suggest convincingly that fertilization is external (Sever et al. 1996a). All other families of salamanders have internal fertilization, involving deposition of a stalked spermatophore by the male partner, either in water or on land depending on the taxon. The spermatophore is produced by secretions of cloacal glands (Sever 1994), and consists of a stalk, surmounted by a cap in which the sperm are imbedded (Zalisko et al. 1984). The female partner clasps the spermatophore cap between her cloacal lips, sperm are extruded, and migrate to glandular spermathecae in the dorsal wall of her cloaca, where they may be stored for varying periods of time. At ovulation, in oviparous species, the sperm are expelled from the spermathecae, and fertilize the eggs as they pass into the cloaca. In viviparous species, fertilization occurs in the oviduct or at the oviducal-cloacal boundary (reviewed by Greven in chapter 12 of this volume). One benefit of internal fertilization utilizing spermatophores and spermathecal storage is that it allows separation of courtship/mating and oviposition/egg brooding in both space and time, and thus allows a species to more effectively utilize the spatial and temporal habitat resources available for reproduction. "& Reproductive Biology and Phylogeny of Urodela In some taxa, oviposition follows within a few days after sperm transfer, as in many pond-breeding species of the genus Ambystoma (summarized in Petranka 1998). In other pond-breeders, as well as stream-breeding and terrestrial species, sperm may be stored for several months, perhaps from one oviposition season to the next (Houck and Schwenk 1984; Sever et al. 1995; Sever et al. 1996b; Sever 2000) or maybe for multiple years, as suggested by Boisseau and Joly (1975) for the larviparous Salamandra salamandra. However, the evidence for multi-year sperm storage is not compelling, as discussed by Sever in chapter 9 of this volume. Sperm storage provides opportunities for multiple matings by females during a given reproductive season, and may engender sperm competition (Houck et al. 1985; Sever 2002). External fertilization is considered the ancestral state in salamanders, given that internal fertilization involving spermatophore production by cloacal glands in males and sperm storage in a spermatheca in females are characters otherwise unknown in vertebrates (Sever 1994). 13.3.2 Parity Oviparity. Nearly all salamanders are oviparous, and this is undoubtedly the ancestral reproductive state. Salthe (1969) recognized three principal modes of oviparity in salamanders: (1) abandonment of eggs in lentic waters, without parental care, e.g., most ambystomatids and salamandrids; (2) deposition of eggs in protected nests in streams, usually with parental care, e.g., dicamptodontids and many plethodontids; and (3) oviposition on land, under cover or in underground nests, usually with parental care, e.g., most plethodontids. He examined variation in egg size, clutch size, clutch volume, larval size, and size at metamorphosis, using regression and graphical methods. His results demonstrated a strong correlation between body volume of the female parent and clutch volume among species, across all reproductive modes, but pronounced variation among modes in the relationship between body volume and clutch size. Thus, Salthe found that species with parental care in modes 2 and 3 tend to have larger eggs and smaller clutches than mode 1 species which abandon their eggs in lentic habitats; he attributed the difference to greater mortality risks to eggs in pond habitats, where the provisioning of parental care in itself may be more difficult. Parental care. Parental care of the eggs in oviparous salamanders has been reviewed by Nussbaum (1985, 1987, and especially chapter 14 of this volume) and Crump (1995, 1996), who may be consulted for details beyond the scope of this chapter. In one of the three families with external fertilization, the Hynobiidae, the male parent may attend the clutch in some species of Hynobius, but this is poorly documented. In the Cryptobranchidae, however, the male parent attends the clutch in all three species. Within the latter family, in Andrias japonicus, communal spawning occurs, but the nests are dominated by a single large male (the “den-master”), who attends and defends the nest until the eggs hatch (Kawamichi and Ueda 1998). In the Sirenidae, parental care by Life Histories "&! the female parent is probable in Siren intermedia (Godley 1983), but care by either parent is unlikely in other members of the family, which scatter their eggs. Among taxa with internal fertilization, pond-breeding ambystomatids and salamandrids lack parental care. In stream-dwelling forms, parental care is lacking in rhyacotritonids and in the salamandrids Taricha rivularis and two of the three species of Euproctus. Apparently either parent may be involved in parental care in Proteus anguinus, at least in captive specimens (Vandel and Bouillon 1959; Vandel et al. 1964). In Necturus, although few precise data are available, one or the other parent may attend the clutch, depending on the species (Bishop 1941; Ashton and Braswell 1979). Otherwise, insofar as is known, care of the egg clutch is provided by the female parent in the Amphiumidae, Dicamptodontidae, most members of the Plethodontidae, and in the ambystomatid Ambystoma opacum (which oviposits in terrestrial habitats). However, parental care is variable in plethodontids that reproduce in ponds and scatter eggs, as well as in those that practice communal nesting, specifically Hemidactylium scutatum (Harris and Gill 1980; Breitenbach 1982) and some species of Batrachoseps (Jockusch and Mahoney 1997). In addition, neotropical plethodontids of the genera Nototriton and (probably) Oedipina lack parental care (Good and Wake 1993; Bruce 1998, 1999). At the other extreme, biparental attendance of the clutch has been observed in Bolitoglossa pesrubra (Ehmcke and Bolaños 1998). Clutch size. Within oviparous species of salamanders, clutch size or clutch volume tends to increase with female body size in many species (Tilley 1968; Bruce 1969; Salthe 1969; Salthe and Mecham 1974; Houck 1977a). Nevertheless, there is ordinarily a high level of residual variance in regressions of clutch size on body size, reflecting the influence of factors other than body size on egg number. Moreover, in some species clutch size is apparently uncorrelated with size of the female parent; in Desmognathus monticola, for example, this tendency is associated with high residual variances in clutch size in local populations having a very narrow range of adult female body size (Bruce and Hairston 1990; Bruce 1996). Hom (1987, 1988) has analyzed constraints on female growth following initial reproduction in desmognathines; her theoretical model suggests that individual females have little opportunity for growth once adulthood is achieved, and that any positive correlation between fecundity and body size may result mainly from variation among individuals in age and size at first reproduction. Overall, however, the general trend of the clutch size-body size relationship in salamanders suggests the operation of fecundity selection on females, defined as the selective advantage of increased fecundity that is gained from an increase in body size. Viviparity: larviparity and pueriparity. The confusing terminology associated with viviparity has been reviewed and revised recently by Greven (Greven 2002 and chapter 12 of this volume), which should be consulted for full treatments of the morphological, histological, and physiological aspects of "&" Reproductive Biology and Phylogeny of Urodela viviparity in salamanders. Here I restrict the discussion to those features of viviparity relevant to population ecology and life history. Greven introduced the terms larviparity for species that give birth to larvae and pueriparity for those that bear fully-metamorphosed juveniles. He clarified the terminology concerning variation in embryonic nutritional modes, as described in chapter 12. Although Greven recommended discarding the terms ovoviviparity and viviparity, I have retained the latter as a useful umbrella that covers both larviparity and pueriparity. Of the six known species of viviparous salamanders, all in the genus Salamandra, three (S. atra, S. lanzai, S. luschani) are pueriparous, whereas various populations of the S. salamandra complex are larviparous, pueriparous, or mixed. Dopazo and Korenblum (2000), in a study of S. salamandra, have hypothesized that pueriparity evolved from larviparity as an exaptation, as a result of intraoviducal competition for resources involving the provision of nutrient eggs by the female. Fecundity in viviparous salamanders differs between larviparous and pueriparous forms. In a larviparous population of Salamandra salamandra, Thiesmeier (1990) reported 8-58 (mean = 33) larvae per female per year, with the number positively correlated with female body weight. The correlation, though significant, was weak, with a high level of residual variance (r2 = 0.145). In contrast, in captive salamanders from a pueriparous population of this species, Thiesmeier et al. (1994) reported a range in brood size of 2-18 (mean = 9.2), with the number again positively correlated with body size of the female parent. Moreover, in both populations average offspring mass was positively correlated with mass of the female. The female reproductive cycle in (mainly) larviparous populations of S. salamandra is annual at low elevations and biennial at high elevations, and gestation periods vary from 3-9 mo to approximately 1 yr, respectively (Joly 1968; Joly et al. 1994). In contrast, in the obligate pueriparous species Salamandra atra, S. lanzai, and S. luschani the gestation periods are often lengthier, 2-4 yr in the former two species (Vilter 1986; Wake 1993; Guex and Greven 1994; Miaud et al. 2001) and 1 yr or longer in S. luschani (Özeti 1979; Polymeni 1994; Olgun et al. 2001). Fecundity in these species is low, ordinarily 2 offspring per brood, as reported in the preceding references and in chapter 12. Given these low numbers, selection favoring larger female body size in these three species may arise not from any advantage of greater fecundity, but possibly from the benefit of larger body sizes of offspring – a relationship which has been documented in both larviparous and pueriparous populations of S. salamandra (Thiesmeier 1990; Thiesmeier et al. 1994). However, I know of no comparable data for the strictly pueriparous species. The combination of long gestation period and low fecundity suggests high survivorship and lengthy life spans in the pueriparous species. Miaud et al. (2001) have reported maximum skeletochronological ages of 22 and 24 yr in two populations of Salamandra lanzai, in samples of 161 and 94 adults, respectively. In contrast, Olgun et al. (2001) determined a maximum age of Life Histories "&# only 10 yr in a sample of 67 adults from a single population of S. luschani. The latter authors compared age and size at maturity, adult survivorship, and life expectancy among this species, S. atra, S. salamandra, and two oviparous species of Triturus. Although adult survivorship appeared to be higher in the viviparous species, the heterogeneous nature of the data sets showed no convincing trends that could be ascribed to viviparity, and the authors concluded judiciously that additional studies are required to better determine relationships among these variables. 13.3.3 Larvae Morphology. Salamander larvae were differentiated as pond- and stream-type by Noble (1931). Later, Valentine and Dennis (1964) subdivided the latter into mountain-brook and stream categories on the basis of variation in branchial structures and tail fin. Duellman and Trueb (1986) largely followed the latter authors in their analysis of larval variation in urodeles. The following discussion is based mainly on these authors’ evaluations. Mountain-brook larvae represent one extreme in the adaptive diversification of salamander larvae, as illustrated by Rhyacotriton, and by Onychodactylus and some species of Batrachuperus among the hynobiids. Larvae of these forms show extreme reduction of the external gills and gill rakers; the gular fold has a fleshy, broadly concave margin. The body tends to be rounded or depressed, the tail is muscular, and the fin is reduced. These are characters associated with a bottom-dwelling life in swift mountain streams. The reduction of the lungs and ypsiloid cartilage in mountain-brook larvae in these genera are adaptations ostensibly serving to reduce buoyancy and facilitate purchase on the stream bottom. At the opposite extreme, pond-type larvae are found in sirenids, amphiumids, Necturus among the proteids, Hynobius among the hynobiids, most Ambystoma, and many pond-breeding salamandrids in several genera; e.g., Notophthalmus, Pleurodeles, Taricha, Triturus, and Tylototriton. They are characterized by bushy gills that have long, thin rami and numerous fimbriae, well-developed gill rakers, and a biconvex gular fold having a thin margin. The body tends to be short and laterally compressed; the weak tail has a high fin that originates on the trunk dorsally, and may extend forward of the cloacal aperture onto the trunk ventrally. The lungs are well-developed and, except in sirenids, amphiumids, and proteids, a ypsiloid cartilage is present in the ventral body wall just forward of the pelvis; the cartilage and associated muscles function to control the pitch of the larva suspended in the water column by regulating the distribution of air in the lungs (Whipple 1906). The lungs function as hydrostatic organs in pond-dwelling larvae; they allow the larva to vary its density in adjusting its position in the water column. In some species larvae undergo vertical movements between the pond bottom and the surface over the daily cycle (Anderson and Graham 1967; Hassinger et al. 1970; Anderson and Williamson 1974; Branch and Altig 1981). At hatching, pond-type larvae in many of the above taxa are provided with balancers, paired lateral "&$ Reproductive Biology and Phylogeny of Urodela projections of the head behind the eyes that prevent the larva from sinking into the soft substrate and aid it in maintaining an upright position (Fox 1984). Balancers are lost early in larval ontogeny. Larvae that are intermediate between mountain-brook and pond types are designated stream-type larvae; this category includes many plethodontids, a few ambystomatids, Ranodon and some Batrachuperus among hynobiids, Cryptobranchus, and the salamandrids Chioglossa, Mertensiella, Euproctus, and Salamandrina. Valentine and Dennis (1964) selected Gyrinophilus to illustrate the intermediate nature of the branchial structures and tail in stream-type larvae. Larvae in this category lack balancers (rudimentary in Salamandrina); a ypsiloid cartilage and lungs may be present, but are often reduced or absent, and are always lacking in plethodontids. The larva of the larviparous salamandrid, Salamandra salamandra, is apparently intermediate between stream- and pond-type; this species typically reproduces in streams, but may also utilize lentic habitats (Thiesmeier and Schuhmacher 1990; Thiesmeier 1994). Valentine and Dennis (1964) and Duellman and Trueb (1986) noted that among the plethodontids, pond-type larvae occur in Hemidactylium scutatum, Eurycea quadridigitata, and Stereochilus marginatus, all of which reproduce in ponds, bogs, swamps, and/or slow-moving streams. Hemidactylium scutatum has a very short larval period and metamorphoses at a small size. In both E. quadridigitata and S. marginatus the pond-type features are replaced by streamtype morphology early in larval ontogeny, and the latter species has streamtype morphology through most of the larval period (Bruce 1971; Birchfield and Bruce 2000). The absence of lungs in plethodontids restricts their larvae to microhabitats on the stream bottom or in detritus and vegetation mats, and may constrain evolution of the pond-type larval morphology in species that utilize lentic habitats. Variation in larval periods and facultative paedomorphosis. Inasmuch as larval development and paedomorphic trends have been studied most thoroughly in the Ambystomatidae, the latter is herein considered in some greater detail than the other families. Most species of Ambystoma in the United States and Canada have large egg clutches and relatively short larval periods, generally less than 1 yr and often in the range 2-4 mo (literature summarized in Petranka 1998). Larval survivorship tends to be low in such species (Anderson et al. 1971; Shoop 1974; Petranka 1984a). Those species distributed over a broad elevational range may prolong the larval period to a second or third year in cooler environments at higher elevations, e.g., A. gracile (Sprules 1974a; Eagleson 1976), A. tigrinum (Sexton and Bizer 1978), and A. macrodactylum (Kezer and Farner 1955; Anderson 1967; Howard and Wallace 1985). In the last species, Anderson (1967) suggested that reproduction is necessarily restricted to permanent ponds at higher elevations, where growth rates are lower and the metamorphic threshold is not attained until the second summer, 14 mo after hatching; whereas at lower elevations, where growth rates are higher, temporary ponds may be utilized and metamorphosis achieved Life Histories "&% in 3-4 mo in association with pond drying. Those species of Ambystoma that reproduce in both ponds and streams tend to have lengthier larval periods in the latter habitats. Thus, Petranka (1984b) demonstrated that in A. barbouri the larval period varies from about 40-70 days in pond populations to 60-100 days in stream populations; the higher growth rates, larger sizes, and earlier metamorphosis in the ponds are apparently due to higher temperatures and greater availability of food in these habitats. Facultative paedomorphosis is known in several species of Ambystoma in temperate North America, including A. gracile (Sprules 1974a), A. talpoideum (see below), and A. tigrinum. The last species shows extreme lability in larval growth rate and age at metamorphosis throughout its extensive range; paedomorphs occur in cold, high-elevation, permanent ponds in the Rocky Mountains of Colorado (Sexton and Bizer 1978) and in permanent bodies of water in arid environments of the Great Plains and deserts of the southwest (Rose and Armentrout 1976; Collins 1981). Sprules (1974b) hypothesized that facultative paedomorphosis in Ambystoma (and Notophthalmus viridescens) is an adaptation to environments where permanent, fish-free ponds and lakes are surrounded by harsh terrestrial habitats. Facultative paedomorphosis in Ambystoma has been most intensively studied in the mole salamander, A. talpoideum, in the southeastern United States. Populations of this species exhibit a broad range of life-history patterns (Petranka 1998). The most detailed studies of larval ecology have been conducted on South Carolina populations (Patterson 1978; Semlitsch 1985, 1987a, 1987b; Semlitsch and Gibbons 1985; Semlitsch et al. 1988). In these populations mating and oviposition occur in winter, with hatching following in February and March. In temporary ponds the larval period is typically 3-7 mo, with metamorphosis occurring in the summer as the ponds dry. In contrast, populations in relatively permanent ponds include both metamorphosing and paedomorphic individuals, and the latter often predominate. In such ponds, some larvae metamorphose in the autumn after a larval period of 6-8 mo; others metamorphose the next spring at 12-15 mo after attaining sexual maturity and reproducing in the winter. Paedomorphs may, however, remain in the ponds, postponing metamorphosis until later years. Populations of Ambystoma talpoideum occupying ephemeral and permanent ponds show some degree of genetic differentiation, as shown experimentally in the South Carolina populations by Semlitsch et al. (1990). These authors demonstrated variation among populations in the propensity to initiate metamorphosis or delay, in response to pond drying. Based on differential responses by larvae from the different pond categories, they suggested that variation among populations in the timing of metamorphosis was the result of either (1) selection for individual plasticity in expression of the paedomorphic or metamorphic phenotype, which requires a larva’s ability to assay the quality of the aquatic environment, or (2) genetic polymorphism in this trait, wherein an individual is either paedomorphic or metamorphic. Such a polymorphism could be maintained by spatial and temporal heterogeneity, under conditions "&& Reproductive Biology and Phylogeny of Urodela when a behavioral response by the larva is ineffective because of unreliability of the environmental cues. In relatively permanent ponds, where paedomorphosis is common, Harris et al. (1990) determined that the trait may have evolved by different genetic pathways In one of the more permanent South Carolina populations of Ambystoma talpoideum, Krenz and Sever (1995) demonstrated that paedomorphic individuals reproduce in autumn, prior to the migration of metamorphic males to the breeding ponds in early winter. They suggested that such a temporal difference confers partial reproductive isolation between paedomorphic and metamorphic components of the population. Ryan and Semlitsch (1998) studied experimentally the effects of density on the timing of metamorphosis and maturation in these same populations of Ambystoma talpoideum. Their most relevant finding concerned a low-density treatment, wherein paedomorphs attained sexual maturity before metamorphic individuals underwent metamorphosis; the latter, in fact, showed no evidence of maturation upon completion of metamorphosis. Such streamlining of the life cycle in paedomorphs provides for early reproduction (< 1 yr posthatching), and eliminates the costs of metamorphosis and subsequent migration to the breeding ponds. In contrast, metamorphs in these populations are at least 1 yr or more beyond hatching at first reproduction. Unlike some salamanders, where metamorphosis appears to be a prerequisite for maturation, facultative paedomorphosis in A. talpoideum and other ambystomatids demonstrates the possibility of decoupling metamorphosis and sexual maturation. Given that the source of larvae in their experiment was a naturally-metamorphic population in a temporary pond, Ryan and Semlitsch’s (1998) results illustrate a high degree of individual plasticity in metamorphosis in this species. The authors proposed that in the evolution of facultative paedomorphosis, the target of selection is age at maturation, with retention of larval morphology a secondary effect. Ideal conditions for such selection would include hydrologic reliability of the aquatic habitat, abundant resources, and low levels of predation and competition. Roff (1996) considered facultative paedomorphosis in salamanders as one of a large class of threshold traits in animals, wherein a discontinuous phenotypic trait, usually dimorphic, is an expression of genetic variation that controls an underlying trait (e.g., hormonal levels) that has a continuous distribution. He cited common garden experiments in Ambystoma gracile (Eagleson 1976), A. talpoideum (Semlitsch and Wilbur 1989; Harris et al. 1990), and Triturus carnifex (Kalezic et al. 1994) as evidence that the paedomorphic/ metamorphic dichotomy has a genetic basis. That the mode of inheritance is polygenic, at least in some cases, is suggested by the trait’s responsiveness to environmental variation, generating a norm of reaction, as seen in Notophthalmus viridescens (Harris 1987) and A. talpoideum (Semlitsch et al. 1990; Jackson and Semlitsch 1993). In hybridization experiments in A. mexicanum and A. tigrinum, Voss (1995) has shown that paedomorphosis has a polygenic basis in ambystomatids. Further evaluation of the threshold model in sala- Life Histories "&' mander life cycles will require detailed investigation of the genetic basis of the physiological and morphogenetic mechanisms that generate the dimorphism, in reference to the norm of reaction of the trait to the range of environments in which the trait is expressed. The highest incidence of paedomorphosis in ambystomatids is seen in the Mexican species, wherein a majority of the species are either obligatorily or facultatively paedomorphic. Most of these are lake-dwelling species of the Mexican plateau, such as the axolotl, Ambystoma mexicanum, and the achoque, A. dumerilii. Some stream-dwelling species of Mexican ambystomatids have variable, often lengthy larval periods, and their populations may also include paedomorphs; e.g., A. ordinarium (Anderson and Worthington 1971) and A. rosaceum (Anderson 1961). In the absence of ambystomatids, the pond-breeding niche in the Old World is occupied by hynobiids and salamandrids, which tend to share a common pattern of larval development. For example, in members of the Triturus cristatus complex, the larval component of the life history typically involves these elements: (1) oviposition and hatching in mid-late spring, followed by a larval period of several months; (2) overwintering by a small proportion of larvae in some populations, with metamorphosis occurring the next spring or summer after a larval period of about 1 yr; and (3) occasional paedomorphosis in some populations (Thiesmeier and Kupfer 2000). In the Japanese hynobiid Hynobius nebulosus, which breeds in small ponds and paddy fields, Kusano (1981) reported a larval period of 3-6 mo, extending from hatching in late April-early May to metamorphosis in July-October. However, he found that a small proportion of larvae overwinter in ponds, metamorphosing the next spring and summer from April to July, after a larval period of 12-15 mo. Other Japanese species of Hynobius may extend the larval period to a second or third year. In the pond-breeding H. retardatus, populations vary in the length of the larval period from < 1 yr to 2 and 3 yr, depending on temperature and elevation (Iwasaki and Wakahara 1999). The larval period of the stream-breeding H. kimurae varies according to habitat reliability: in a stream that typically dried in autumn all larvae metamorphosed in late summer, whereas in a permanent stream only 21% transformed in the first year, with the remainder overwintering and transforming in the second year (Misawa and Matsui 1997). Similar patterns occur in some European newts. Bell and Lawton (1975), in a study of Triturus vulgaris in England, found that oviposition occurs in three phases from March to June, and that early larvae metamorphose in the summer of the same year after a larval period of about 2 mo. However, some members of the late larval cohort overwinter, metamorphosing the next spring after a larval period of 9-11 mo. At more northern localities, in Scandanavia, larval overwintering has been reported in both T. vulgaris and T. cristatus (Hagström 1979). Yet Dolmen (1983) found that at extreme northern sites, near the limits of the range of both of these species in Norway, growth and development are rapid, with metamorphosis occurring after larval periods of "' Reproductive Biology and Phylogeny of Urodela 1-4 mo, and with no evidence of larval overwintering. He attributed the high growth/developmental rates to warm, sunny, summer days of long daylengths at these latitudes during the weeks following hatching in June. In stream-breeding hynobiids and salamandrids of Eurasia, the larval growth rates are often low and the larval periods may be prolonged, paralleling the situation found in stream-breeding North American ambystomatids, dicamptodontids, rhyacotritonids, and plethodontids (see below). For example, Bannikov (1949) reported a larval period of 2+ yr in the hynobiid Ranodon sibiricus, a high-elevation species that lives in and along small, fast-flowing mountain streams. In the mountain-stream salamandrid Mertensiella caucasica the larval period is estimated to vary from 2 to 4 yr (Tarkhnishvili 1994). Whereas high-elevation populations of the stream-dwelling salamandrid Euproctus asper have larval periods of about 2 yr, in lower-elevation populations the larval period is about 14 mo duration (Clergue-Gazeau and Beetschen 1966). Some other Old World streamside salamandrids may also have lengthy larval periods (summarized in Grossenbacher and Thiesmeier 1999). However, that of Salamandra salamandra is usually completed in several months, although overwintering sometimes occurs (Thiesmeier 1990, 1994). North American salamandrids overlap broadly with ambystomatids, and members of the two families often share breeding ponds. Yet salamandrids appear to exhibit less variation in the duration of the larval phase. In most populations of the several species of Notophthalmus and Taricha the larval phase is usually < 0.5 yr (Petranka 1998). However, the larval period may be longer in populations of N. viridescens having branchiate adults. In T. granulosa Farner and Kezer (1953) reported a larval period of about 1 yr in a highelevation population breeding in a cold-water lake. The two small families of stream-dwelling and streamside salamanders restricted to the Pacific Northwest of North America tend to have relatively lengthy larval periods. In Dicamptodon, with four species, facultative paedomorphosis is prevalent and paedomorphosis is the dominant life-history mode in D. copei; in other species, metamorphosis may occur after larval periods >2 yr in D. aterrimus and 1.5-3.0 yr (or even greater) in D. tenebrosus (Nussbaum and Clothier 1973). Although Dicamptodon and Rhyacotriton overlap broadly, paedomorphosis is unknown in the four species of Rhyacotriton; larval periods vary from 2.0-2.5 yr in R. variegatus to 3-4 yr in R. cascadae (Nussbaum and Tait 1977). In the two families of salamanders that undergo partial metamorphosis, the amphiumids and cryptobranchids, larval forms with external gills metamorphose very soon after hatching in the former; e.g., 1-3 wk in Amphiuma tridactylum (Ultsch and Arceneaux 1988). In the North American cryptobranchid, Cryptobranchus alleganiensis, gills are retained for 1.5-2.0 yr after hatching (Bishop 1941; Nickerson and Mays 1973). Larval periods of plethodontids with biphasic life cycles (Desmognathinae, Hemidactyliini) vary considerably. In desmognathines the stream-edge species tend to have larval periods £ 1 yr, whereas in the more aquatic and larger Life Histories "' species the larval phase is longer, usually between 2 and 4 yr (Tilley and Bernardo 1993: Table 1). Considerable variation is found in Desmognathus quadramaculatus, wherein the larval period ranges from 1-2 yr in some populations (Organ 1961; Camp et al. 2000; Beachy and Bruce 2003) to 3-4 yr in others (Bruce 1988a; Austin and Camp 1992; Camp et al. 2000; Bruce et al. 2002). Similar levels of variation in larval periods are found in hemidactyliines. The briefest larval phase, estimated at 1.5 mo (Blanchard 1923), occurs in Hemidactylium scutatum. However, the taxonomic allocation of this species to the Hemidactyliini is uncertain (Rose 1995; see also Larson et al. in chapter 2). In metamorphosing members of Eurycea the larval periods vary from 3 to 36 mo, with the shorter periods in pond-breeding and the longer in streambreeding populations/species (Ryan and Bruce 2000: Table 2). There is often considerable variation within species or populations. Voss (1993) demonstrated that such variation in E. wilderae is correlated with stream order; larvae in populations in first-order streams metamorphose after 12-15 mo, while those in higher-order streams prolong the larval phase an additional year. He proposed that the difference reflected variation in oviposition and hatching periods associated with variation in the thermal regimes of lower- and higherorder streams. The lengthiest larval periods in biphasic hemidactyliines, 3-5 yr, occur in Gyrinophilus porphyriticus (Bruce 1980) and perhaps in G. subterraneus (Besharse and Holsinger 1977). The related genera Pseudotriton and Stereochilus also have lengthy larval periods, which may exceed 2 yr (Ryan and Bruce 2000: Table 1). In both P. montanus (Bruce 1974, 1978) and P. ruber (Bruce 1972, 1974; Semlitsch 1983) there is a tendency for slower larval development and lengthier larval periods in higher-elevation populations. 13.3.4 Obligatory Paedomorphosis In reference to the salamander model, paedomorphosis ordinarily involves complete developmental truncation of the somatic tissues, such that larviform morphology is retained throughout life. Paedomorphosis, either facultative or obligatory, is known in nearly all salamander families, with the small Rhyacotritonidae being the single exception. Facultative paedomorphosis, as noted above, is often associated with relatively permanent and productive aquatic habitats that are surrounded by harsh terrestrial environments, with harshness deriving from abiotic and/or biotic factors. Obligatory paedomorphosis, where the trait is fixed genetically, often at the family level, is associated frequently with adaptation to large, generally permanent bodies of water, including large streams, rivers, lakes, and swamps, as seen in Amphiumidae, Cryptobranchidae, Sirenidae, and (in part) Proteidae. Members of the Amphiumidae and Cryptobranchidae have incomplete metamorphosis, and thus represent a special case of dissociative paedomorphosis. Given that members of these four families occur today in areas of eastern North America and eastern Asia where terrestrial salamanders are common and the abiotic "' Reproductive Biology and Phylogeny of Urodela terrestrial environment benign, the abandonment of the terrestrial phase of the life cycle in these lineages may represent a consequence of adaptation to an aquatic niche not fully accessible to fishes. Such a niche includes a bottomdwelling and burrowing mode of life under (or within) such cover objects as rocks, logs, leaf packs, and root masses, where paired limbs (rather than fins) are advantageous for bottom-crawling and purchase on the stream or lake bottom (though nearly dispensed with in the eel-like amphiumids and sirenids), and where retention of lung-breathing allows utilization of aquatic habitats that sometimes become hypoxic. Otherwise, obligatory paedomorphosis represents an adaptive route to exploitation of permanent aquatic habitats in harsh (dry, unproductive) terrestrial environments, including subterranean environments. Exemplars of this adaptive pathway are the proteid Proteus anguinus of southern Europe, several ambystomatid species of the Mexican Plateau, and numerous hemidactyliine plethodontids of eastern and central North America. Among the last, a particularly diverse adaptive radiation in Eurycea has occurred in the relatively arid Edwards Plateau, where an extensive aquifer supplies a vast network of surface and underground habitats. The assemblage includes epigean species that retain larval features typical of metamorphosing species of Eurycea, as well as cave-dwelling forms that have highly-derived morphologies. A question of interest is whether paedomorphosis in salamanders is an evolutionary product of progenesis or neoteny. Assuming that the ancestral life history was biphasic, the question can be addressed in facultative paedomorphs by comparing rates of development of paedomorphs and metamorphs. Thus, Ryan and Semlitsch (1998), in determining that in Ambystoma talpoideum paedomorphs mature at an earlier age than metamorphs but may in turn ultimately metamorphose themselves, demonstrated that paedomorphosis is a joint consequence of (1) accelerated maturation (= progenesis), and (2) retardation of somatic development or postdisplacement (later onset) of metamorphosis (= neoteny). Similar analyses could be conducted for other facultative paedomorphs. However, for obligate paedomorphosis such an evaluation becomes more difficult. This is especially true for the families of “giant” salamanders that have no close biphasic relatives. Although data are scanty, it is unlikely that early maturation has been a factor in these forms. In Necturus maculosus Bishop (1941) estimated sexually maturity at 5-6 yr, with males apparently maturing earlier than females. Cryptobranchids are a special case, since a partial metamorphosis does occur. In Cryptobranchus alleganiensis gill loss occurs at 1.5-2.0 yr, with maturation following at 4-5 yr in males and 7-8 yr in females (Taber et al. 1975; Peterson et al. 1983; Peterson et al. 1988). Thus, in this species partial paedomorphosis has apparently evolved from postdisplacement of part of the metamorphic process. Life Histories "'! As noted above, obligate paedomorphs are found in some families and genera also containing metamorphosing members, i.e., ambystomatids and plethodontids. Half (17/34) the species of hemidactyliine plethodontids are paedomorphic. Nearly all of these have congeneric relatives with biphasic life cycles. Thus, it may be possible to determine the evolutionary pathways to paedomorphosis using comparative methods once the systematics of these salamanders is stabilized. In Eurycea neotenes, I have suggested that paedomorphosis has evolved by progenesis, because age at maturation is lower than in several biphasic species of Eurycea (Bruce 1976). Sweet (1977) later discovered populations of E. neotenes in which metamorphosis does occur, varying in frequency among populations. The body-size distributions in his small samples tended to support the progenesis hypothesis; i.e., only the metamorphs were sexually mature, and they were larger than larviform adults in the populations where paedomorphosis is the rule, as in those I studied (Bruce 1976). However, this argument may be compromised, given the likelihood that the populations under comparison may belong to different species (Chippindale et al. 2000). 13.3.5 Metamorphosis Metamorphosis is the rapid change in the organization of the amphibian body plan that provides for a transition or niche shift from the wholly aquatic environment of the larva to the more terrestrial mode of life of the juvenile and adult. The most comprehensive study of the morphological reorganization that occurs at metamorphosis in salamanders is Wilder’s (1925) classic account of metamorphosis in Eurycea bislineata. Duellman and Trueb (1986) have provided a more recent summary of morphological and other changes a t metamorphosis in salamanders. The principal morphological changes include (1) resorption of the external gills and closure of the gill slits, (2) fusion of the gular fold to the throat, (3) changes in the branchial circulation and hyobranchial skeleton and musculature, (4) formation of a tongue, (5) widening of the mouth and development of the maxillary bones and teeth, (6) loss of labial folds, (7) resorption of the tail fin, and (8) complex changes in the epidermis and dermis of the skin, including reorganization of glandular elements and development of adult pigmentation. Wake and Hanken (1996) have suggested, citing Schoch (1992), that the abrupt metamorphosis in living amphibians, including salamanders, is a caenogenetic trait, given that the ancestral ontogeny in early tetrapods apparently involved a gradual transition from the larval to the adult condition. Metamorphosis in amphibians is regulated mainly through the hypothalamopituitary-thyroid and hypothalamo-pituitary-interrenal axes (Burggren and Just 1992; Shi 2000). Far more experimental studies have been conducted on anurans than on salamanders and the following brief summary is based largely on the former. The principal agents stimulating metamorphosis are thyroid hormones (TH), comprising the iodinated thyronine compounds T3 and T4, with the latter a precursor to the biologically more active T3. Secretion "'" Reproductive Biology and Phylogeny of Urodela of TH is regulated by thyroid stimulating hormone (TSH) from the pituitary, which in turn is controlled by corticotropin-releasing hormone (CRH) from the hypothalamus. The latter also regulates pituitary secretion of ACTH, which acts on the interrenal glands to stimulate secretion of such steroids as corticosterone and aldosterone, which in turn serve to modulate TH-induced metamorphosis, either antagonistically or synergistically, varying according to the target tissue and developmental stage (Hayes 1997). The recognition that CRH is the overall regulator of the metamorphic activities of both the thyroid and interrenal glands (Denver and Licht 1989) represents an important step toward development of an integrated model of the ecological and neuroendocrine regulation of metamorphosis (Denver 1997). Prolactin (PL) is a pituitary hormone which stimulates growth in amphibian larvae, and acts to inhibit TH-stimulated metamorphosis. It is regulated by thyrotropin-releasing hormone (TRH) produced in the hypothalamus. The precise role of prolactin remains unclear; however, inasmuch as PL levels increase at metamorphic climax, the hormone may act antagonistically on TH to regulate the sequence of morphogenetic changes in metamorphosis (Denver 1997). The several axes of the neuroendocrine system controlling larval growth and metamorphosis are modulated by positive and negative feedback loops that regulate the timing of hormone release (Shi 2000). Denver (1997) has argued that plasticity in the duration of the amphibian larval period is an adaptive response to habitat deterioration, requiring the larva’s ability to assess changes in habitat quality through various sensory modalities, and then to engender a response leading to metamorphosis, mediated through the higher brain centers and the hypothalamo-pituitary axis. Under this hypothesis, hypothalamic CRH is the essential hormone that activates both the pituitary thyrotropes (Æ TSH) and corticotropes (Æ ACTH) to stimulate secretion of the thyroid and interrenal agents of metamorphosis, i.e., TH and steroids, respectively. It was this author’s contention that the adaptive response to larval habitat uncertainty – i.e., plasticity in metamorphic timing – evolved by modifying an ancestral stress response mechanism to serve as the proximate mechanism controlling initiation of metamorphosis. 13.3.6 Maturation Endocrine system. Development to sexual maturation in amphibians is controlled by the hypothalamo-pituitary-gonadal axis. Again, much of our knowledge of the mechanism is based on studies of anurans, but a similar mechanism probably applies in urodeles (Jørgensen 1992; and especially chapter 8 herein). Gonadotropin releasing hormone (GnRH) of the hypothalamus stimulates the release of gonadotropins (GTH) from the pituitary, specifically luteinizing hormone (LH) and follicle stimulating hormone (FSH), which promote growth, maturation, and steroid secretion of the ovaries and testes. In females, circulating levels of GTH vary seasonally; relatively low, threshold levels are required for normal vitellogenesis, but a sharp increase is observed at the time of ovulation. In males, GTH is required for spermatogenesis; Life Histories "'# analogous to females, spermatogenesis proceeds in an environment of low circulating levels of GTH, but spermiation follows only after a pronounced rise in GTH levels. Jørgensen (1992) has suggested that a parallel in the annual cycle of GTH in males and females acts to synchronize ovulation and spermiation in a population’s annual breeding cycle. What is intriguing in amphibians is the interrelationship of the endocrine controls of metamorphosis and maturation, and the degree of independence of the two systems. Hayes (1997: Fig. 5) presented a schematic based on a broad range of research, some of it contradictory across species, which indicates a high level of integration of CRH-mediated metamorphosis and GnRHmediated maturation. However, the mechanisms of disengagement of the two systems in the diversification of amphibian life cycles in the direction of either paedomorphosis or direct development remain open questions. Variation in age at first reproduction.1 Duellman and Trueb (1986: Table 2-8) tabulated variation in age at first reproduction in selected species of salamanders based on a survey of the literature through 1978. Their compilation reflects the considerable interspecific variation in this important trait, but scarcely touches on variation within species. Although the Duellman and Trueb table is based on a heterogeneous set of studies of varying reliability, it does indicate that in salamanders generally, either (1) males and females attain maturity at the same age, or (2) females are older at first reproduction than males. This trend is correlated with patterns of sexual size dimorphism in salamanders (Shine 1979; Bruce 2000). Since the publication of Duellman and Trueb’s book, many studies of age structure in salamanders have utilized skeletochronology (Castanet and Smirina 1990), which provides a direct estimate of individual age based on a morphological trait, in contrast to earlier methods that often relied on group data (size-frequency histograms) to identify age classes. Both the older and newer methodologies have demonstrated pronounced geographic and altitudinal variation in age at first reproduction in many species. It is remarkable 1 Age at first reproduction of oviparous species of salamanders is indexed from the estimated month of an individual’s entry into the population as a zygote. In species with external fertilization, and in those species with internal fertilization in which oviposition follows shortly after mating, the age at first reproduction of both males and females will be a whole number of years. However, in many species with internal fertilization there is a lag between mating/sperm transfer and fertilization/oviposition. This means that in terms of demographic parameters, particularly survival to first reproduction, males “reproduce” some months before their offspring of the current year enter the population, and that during this period survival of a male is immaterial relative to the well-being of his current brood. Thus age at first reproduction in males in these species is more appropriately given in fractional units of years (or in months). Many authors have not distinguished between age at maturation and age at first reproduction. Given that spermatogenesis and vitellogenesis are initiated ordinarily many months prior to sperm transfer, fertilization, and oviposition, failure to differentiate between these parameters may lead to inconsistent and misleading comparisons. Moreover, for many species the lag time between mating and fertilization/ oviposition is not known. "'$ Reproductive Biology and Phylogeny of Urodela that salamanders, which are small vertebrates, often have relatively lengthy periods of development to maturation, such that growth rates are low and survival rates high, in comparison with other taxa of small ectothermic vertebrates. Thus, salamanders seemingly direct their available resources mainly to maintenance and storage, rather than to growth and reproduction, over considerable portions of their life spans. Pronounced variation in age at first reproduction is known in many species of salamanders. Some of the variation stems from geographic variation in this trait, but part originates within populations. In the following sections I consider the magnitude of this variable in several taxa whose life histories have been studied intensively using both traditional and skeletochronological methodologies. Notophthalmus viridescens. North American newts of the genera Taricha and Notophthalmus generally have short (< 0.5 yr) larval periods and lengthy juvenile stages of several years duration, such that sexual maturation is not reached until ages of five years or greater (Petranka 1998). In the eastern newt, Notophthalmus viridescens, earlier accounts of variation in age at first reproduction, based mainly on body size distributions (Bishop 1941; Chadwick 1944; Healy 1973, 1974), have been confirmed and extended by more recent skeletochronological studies (Forester and Lykens 1991; Caetano and Leclair 1996). The juvenile stage of this species is sufficiently distinct in morphology to merit the special name of red eft, which is terrestrial, in contrast to the generally aquatic adult. In some populations, defined by both geography (inland, upland) and habitat quality (permanency of ponds), the adults are wholly aquatic (Chadwick 1944; Hurlbert 1969; Healy 1975; Caetano and Leclair 1996); whereas in others the adults leave the breeding ponds for part of the year, with the phenology of the migration varying geographically (Murphy 1963; Hurlbert 1969; Gill 1978; Massey 1990). Among populations of this species having a life cycle that includes a juvenile red eft, this stage may extend from 2 to 7 yr; thus, age at first reproduction varies considerably. Healy (1974) estimated that the eft stage is 3-7 yr in inland Massachusetts populations, such that age at first reproduction varies from 4 to 8 yr. Using skeletochronology to age newts in a population in the Blue Ridge Mountains of Maryland, Forester and Lykens (1991) found that adults overlap efts in the age range 4-7 yr, which therefore yields an estimate of the age range at first reproduction of 4-8 yr. In contrast, for Quebec populations, Caetano and Leclair (1996), using skeletochronology, estimated age at first reproduction at 2-4 yr. Both Healy (1975) and Caetano and Leclair (1996), in noting that the minimum size in mature males is less than in females, concluded that males are smaller and younger than females at first reproduction. The latter authors’ skeletochronological results suggested that adult females average larger than males; however, the differences they reported were small and generally nonsignificant statistically. Life Histories "'% In contrast, in lowland and coastal populations of newts the red eft stage may be abbreviated or bypassed altogether. Larvae either metamorphose into an aquatic juvenile preceding development to maturity, or undergo partial metamorphosis whereby the aquatic juveniles and adults retain the external gills and some other larval features of the branchial region (Noble 1926, 1929; Bishop 1941; Brandon and Bremer 1966; Healy 1970, 1973, 1974). Although such adults are often referred to as paedomorphs, Brandon and Bremer (1966) called them “branchiate adults,” in consideration of the fact that they are fully metamorphosed otherwise. Adults in these populations may retain the ability to complete metamorphosis if the breeding ponds dry. In populations where the eft stage is skipped or foreshortened and/or facultative “paedomorphosis” in the form of gill retention is prevalent, sexual maturation may be attained in the first or second year, such that first reproduction occurs at an age of 1 or 2 yr (Healy 1974; Harris et al. 1988). Harris (1987) has shown that in the North Carolina Coastal Plain low larval density results in paedomorphosis or metamorphosis to aquatic adults and early reproduction at 1 yr, whereas high larval density promotes metamorphosis to an abbreviated eft stage and first reproduction at 2 yr. The prevalent life-history strategy in Notophthalmus viridescens, involving slow growth and development during the lengthy red eft stage, and consequent long generation time, is correlated with the presence of a highly toxic skin secretion (tetrodotoxin) that reduces the incidence of predation and the range of predators that will feed on newts, especially on the more toxic (and aposematic) red eft stage (Brodie 1968a). Similar and even more potent toxins are secreted by the skins of western newts of the genus Taricha (Wakely et al. 1966; Brodie 1968b), which have similar life histories. Thus, these North American newts have evolved an allocation strategy that emphasizes maintenance over rapid growth and early reproduction. However, the eastern newt, N. viridescens, retains considerable plasticity that allows it to switch to an alternative strategy of rapid growth and development to maturation under environmental conditions (permanent ponds, dry terrestrial environment) that select against the terrestrial eft stage. It is possible that the lower levels of toxicity of larvae, juveniles, and adults in the aquatic environment versus the much higher levels of toxicity of red efts in the terrestrial environment elicit greater selective pressures (i.e., from higher predation) for early reproduction in those populations that forgo the eft stage. Given that the alternative life-history strategies involve larval and adult stages that are aquatic, the main target of selection is expected to be the post-metamorphic juvenile stage (whether terrestrial or aquatic, and whether gilled or not). Old-World salamandrids. Detailed studies of several widespread species of European newts have demonstrated considerable geographic variation as well as within-population variation in age at first reproduction, mainly within the range 2-7 yr. Three such species are Triturus cristatus (Hagström 1977, 1979; Dolmen 1983; Francillon-Vieillot et al. 1990; Miaud et al. 1993), T. marmoratus (Caetano et al. 1985; Francillon-Vieillot et al. 1990; Arntzen and Teunis 1993; 498 Reproductive Biology and Phylogeny of Urodela Caetano and Castanet 1993; Díaz-Paniagua et al. 1996; Díaz-Paniagua 1998), and T. vulgaris (Bell 1977; Hagström 1977, 1979; Dolmen 1983). Variation in growth of T. cristatus and T. marmoratus has been analyzed recently by Arntzen (2000) using the von Bertalanffy growth model. He reported a trend in both species for females to attain larger sizes than males, which is correlated with a tendency for delayed maturity in females versus males in these species. The most extreme variation in age at first reproduction in European Triturus is found in the Alpine newt, Triturus alpestris, which occurs over a broad altitudinal range, extending from near sea level to elevations of 2,500 m. The pronounced variation in age at maturity is correlated, at least in part, with altitude. Miaud et al. (2000: Table 1) have summarized the results of eight recent skeletochronological studies of age structure in this species, which show that age at first reproduction varies from 1-2 or 3-4 yr in two populations at an elevation of 270 m, to 9-11 yr in a population at 2,200 m. Although growth rates (and the length of the growing season) are lower in the highelevation populations, the salamanders attain larger sizes at high versus low elevations, apparently a consequence of delayed maturity at the higher localities. In these populations, females tend to attain maturity at larger sizes than males, and mean adult body size is greater in females; however, there is no clear difference in age at maturation between the sexes. Marunouchi et al. (2000) reported somewhat similar variation in the Japanese newt, Cynops pyrrhogaster. Although this species is distributed over a much lesser elevational gradient than the Alpine newt, variation in age at maturity follows a pattern similar to that of the latter species, ranging from 3 yr at elevations less than 500 m to 4-7 yr at elevations of 500-1,140 m. Males were shown to mature at a younger age than females in nine of 12 populations of C. pyrrhogaster, and in all 12 adult females were larger than males in mean and maximum sizes. A recent skeletochronological study of two Alpine populations of Salamandra lanzai at slightly different altitudes has demonstrated considerable differences in age at maturity, apparently unrelated to variation in body size (Miaud et al. 2001). In the higher-elevation population minimal age of mature individuals was 8 yr in both sexes in a sample of 135, whereas in the lower-elevation population comparable ages in a sample of 203 were 3 yr in males and 6 yr in females. However, differences in body-size parameters were minimal. Desmognathinae. In the plethodontid subfamily Desmognathinae of eastern North America, interspecific variation in adult body size is tightly correlated with age at maturation: the larger species are older at first reproduction (Tilley and Bernardo 1993). The subfamily is unusual in that males mature earlier and at smaller sizes than females, as is common in salamanders, but then, in most species, males outgrow and/or outsurvive females, to attain greater maximum body sizes (Juterbock 1978; Bruce 1993). The larval/juvenile phases are relatively lengthy in desmognathines. Hom (1987, 1988) has argued that in members of the Desmognathus ochrophaeus complex, as well as Life Histories "'' in other desmognathines, special aspects of trophic ecology, energetics, behavior, and reproductive cycles are factors promoting selection for delayed maturation. The extreme desmognathine in adult body size and age at maturation, insofar as known, is Desmognathus quadramaculatus, a species of the southern Blue Ridge Mountains. Camp et al. (2000: Table 1), in tabulating data on variation in age/size at maturation in populations of this species over the entire latitudinal gradient of the geographic range, showed that age at maturation varies from approximately 3 to 6 yr, and is likely correlated in complex fashion with latitude, elevation, and rainfall. Other data, either not tabulated by Camp et al. (2000) or more recent, tend to confirm these relationships (Organ 1961; Castanet et al. 1996; Bruce et al. 2002; Beachy and Bruce 2003). Skeletochronological analysis by Castanet et al. (1996) and Bruce et al. (2002) demonstrated that age at first reproduction varies by approximately 1-2 yr between two nearby populations in the southern Blue Ridge (Wolf Creek: 6 yr, 7-8 yr; Coweeta: 7-8 yr, 9-10 yr), and that the differences are correlated with differences in adult body size. In contrast, Beachy and Bruce (2003), based on size distributions, estimated ages at first reproduction of 3-5 yr in a population of small-bodied D. quadramaculatus at a more northern locality in the southern Blue Ridge. Camp et al. (2000) concluded that variation in age and size at maturation in this species is determined largely by age and size at metamorphosis, which in turn is controlled by the effects of moisture on the length of the larval period and temperature on larval growth rate. Given that age at metamorphosis and age at maturation are known to vary independently in desmognathines, including D. quadramaculatus, a reconsideration of the Camp et al. (2000) hypothesis at a finer scale of resolution seems warranted. Geographic variation in age at first reproduction is also documented in other species of desmognathines. In the Desmognathus ochrophaeus complex, following earlier studies (Tilley 1973, 1974) of the life history of D. carolinensis, Tilley (1980) conducted an elegant mark-recapture investigation of D. ocoee, in which he was able to construct life tables for two populations at different elevations. His results showed that members of the higher-elevation population attain maturity a year later on average than those of the lower-elevation population. Bernardo (1994) later conducted reciprocal-transplant experiments using common gardens to examine the environmental and genetic contributions to variation in life-history traits in these same populations of D. ocoee. He reported no evidence of a genetic difference in growth rate or plasticity in this parameter between populations. However, the experiments did detect a genetic difference in development to sexual maturation, with the higher-elevation salamanders delaying maturation and having less plasticity in this trait than those at lower elevations. The results suggested that growth rate and maturation rate may evolve independently in this species. Bernardo speculated that the main target of selection in the D. ocoee metapopulation might be either age at maturation or body size, under alternative or complementary selective regimes. # Reproductive Biology and Phylogeny of Urodela In the first case, as suggested earlier by Tilley (1980), selection on age at maturation might be a response to lower versus higher extrinsic adult mortality in high- versus low-elevation environments, which has resulted in delayed maturation, larger body size, and larger clutch size and/or larger eggs in high-elevation populations. Or, under the second scenario, with body size as the target of selection, smaller body size may be favored at lower elevations as a response that reduces maintenance costs in relation to the higher metabolic rates at the higher temperatures of lower-elevation environments. Early maturity would follow as an outcome of correlational selection. Bernardo also suggested that larger body size and later maturity may be an outcome of selection for larger egg size, resulting in larger hatchlings and higher survival in overwintering larvae at colder, higher elevations. His analysis illustrates the complexities inherent in understanding variation in age and body size of maturation in salamanders (and other organisms), and the need to design research to encompass the full suite of allocation functions. 13.3.7 Reproductive Cycles Reproductive cycles in vertebrates may be categorized as (1) associated, dissociated, or continuous; (2) aseasonal or seasonal; or (3) annual, biennial, or irregular (Whittier and Crews 1987; Jørgensen 1992). Associated and dissociated cycles occur in temperate-zone salamanders that have seasonal reproduction. In the former case, spermatogenesis and oogenesis are synchronous, such that courtship and sperm release/transfer are followed immediately, in either order, by oviposition and fertilization. This kind of cycle is found in salamanders having external fertilization (hynobiids, cryptobranchids, sirenids) and in species with internal fertilization where males and females gather at breeding ponds for brief periods each year when weather conditions are favorable, e.g., many species of Ambystoma. In contrast, in dissociated cycles spermatogenesis and oogenesis are not necessarily synchronous. Courtship and sperm transfer occur weeks or months before oviposition; following transfer the sperm are retained and perhaps nourished (but see Sever, chapter 9 of this volume) in the spermatheca of the female until the oviposition season. Such cycles are characteristic of plethodontids. Continuous cycles, which are equivalent to aseasonal, involve year-round reproduction, and are probably uncommon in salamanders, except for some species of neotropical bolitoglossines in aseasonal environments. In reference to the latter, Houck determined that males in several Guatemalan species (Houck 1977a), including Bolitoglossa rostrata (Houck 1977b), had sperm in the testes and vasa deferentia throughout the year, leading her to conclude that courtship and mating may occur year round. However, females in several of these species, including B. rostrata, oviposited on a seasonal basis, depositing the clutch at the beginning of the dry season and brooding the clutch for as long as 5-6 mo, with hatching occurring at the start of the wet season. McDiarmid and Worthington (1970) had proposed a similar seasonal female cycle for some tropical bolitoglossines, but these authors and Houck Life Histories 501 (1977a) both noted that in other species oviposition is aseasonal. Thus, in the large clade of tropical plethodontids male and female reproductive patterns may differ, and interspecific variation in the female cycle is in part correlated with the magnitude of wet-dry seasonality. Nevertheless, there is at least one case of microsympatry of an aseasonal and a seasonal species (Houck 1977a). In her study of Bolitoglossa rostrata, Houck (1977b) concluded that females probably oviposit biennially rather than annually. Jørgensen (1992) has reviewed the evidence for annual, biennial, and irregular reproductive cycles in amphibians, particularly females; much of the evidence for salamanders is inconclusive or contradictory. Evidence for substitution of a biennial (or longer) cycle for an annual cycle is usually explained on the basis of the energetic costs to females of vitellogenesis and brooding of the clutch. In North American plethodontids, females of many species of Plethodon appear to reproduce biennially (Marvin 1996), whereas most desmognathines are annual (Bruce 1993). Some species in other families may reproduce over an even longer interval. For example, in Ambystoma maculatum, Husting (1965), in a mark-recapture study, estimated that only about one-third of the adults reproduce in a given year; how this finding relates to individual male and female reproductive cycles is unclear. The extreme periods of reproductive cycles in salamanders may be the 3 and 4 yr female cycles of low and high elevation populations of Salamandra atra (Guex and Greven 1994). 13.3.8 Longevity Early studies of life histories and population ecology of salamanders using the methodologies of mark-recapture and analysis of body-size distributions have shown that salamanders have high survivorship, lengthy mean generation times, and are long-lived, in comparison with other taxa of small ectothermic vertebrates (reviewed in Hairston 1987). Data on salamanders in captivity support this conclusion (e.g., Oliver 1955: Table 14; Duellman and Trueb 1986: Table 11-2; Hairston 1987: Table 2.2). Demographic studies of salamanders, the results encapsulated in survivorship curves and life tables, ordinarily have extended the analysis of survivorship and fecundity to ages of 10 yr and above (Organ 1961; Bell 1977; Tilley 1980; Hairston 1983; Bruce 1988b). Moreover, growth curves generated for salamanders, mainly using markrecapture methods, have suggested that some members of a population may have lengthy life spans. Thus, Marvin (2001) estimated that in Plethodon kentucki maximum ages are 13 yr in males and 16 yr in females. Other species may live much longer; growth studies of Cryptobranchus alleganiensis suggested that some individuals survive to > 25 yr (Taber et al. 1975; Peterson et al. 1983; Peterson et al. 1988). The development of skeletochronological techniques, in providing a means of accurately aging salamanders directly, has confirmed the conclusion that salamanders generally are long-lived. For example, numerous skeletochronological studies of European salamandrids in the genus Triturus have yielded maximum ages of 11 to 17 yr in various populations and species (Caetano 502 Reproductive Biology and Phylogeny of Urodela et al. 1985; Francillon-Vieillot et al. 1990; Caetano and Castanet 1993; Miaud et al. 1993). However, in samples of two high-elevation Alpine populations of T. alpestris the oldest individuals were 20 and 22 yr (Miaud et al. 2000). Similarly, in the pueriparous Salamandra lanzai maximum ages of 22 and 24 yr have been recorded for two Alpine populations (Miaud et al. 2001). In contrast, in S. luschani maximum ages were 8 yr in males and 10 yr in females (Olgun et al. 2001). An extreme age of 26 yr has been recorded for the stream-dwelling European salamandrid Euproctus asper (Montori 1990). Likewise, in the stream-breeding Mertensiella caucasica, lengthy life spans, to 26 yr, have been documented (Tarkhnishvili and Gokhelashvili 1994). However, in another streamside species, Chioglossa lusitanica (the putative sister species of M. caucasica) maximum skeletochronological age was only 10 yr (Lima et al. 2001). In the North American newt Notophthalmus viridescens skeletochronological ages of 9 yr and 13 yr have been documented in Maryland (Forester and Lykens 1991) and Canadian (Caetano and Leclair 1996) populations, respectively. Much greater ages, up to 32 yr, are attained by the ambystomatid Ambystoma maculatum (Flageole and Leclair 1992), which is broadly sympatric with Notophthalmus viridescens in North America. In studies of nearby populations of the same species, skeletochronological ages are generally concordant with age estimates obtained by other methods. For example, in Plethodon metcalfi Hairston (1983) generated a life table from size-frequency data. He projected the age column to 30 yr and estimated 0.001 survivorship to this age. In a skeletochronological evaluation of a sample of 109 individuals from a nearby population of this species (Ash et al. 2003), maximum ages were 9 yr (n = 3) and 10 yr (n = 1). For comparison, in Hairston’s life table survivals to 9 and 10 yr are estimated at 0.043 and 0.035. Given the errors inherent in the methodologies of both studies, the results appear to be relatively concordant. Another such comparison is possible for Desmognathus ocoee. In his longterm mark-recapture study, Tilley (1980) extended the life tables of two populations of this small species to 12 yr and 16 yr, and estimated survivorship to 10 yr to be 0.005 in the first and 0.013 in the second population. For two different but nearby populations of this species, Castanet et al. (1996) and Bruce et al. (2002) reported maximum skeletochronological ages of 10 yr and 8 yr in samples of 76 and 101 individuals. Had the samples been larger it is likely that maximum ages would have been increased to the limits set by Tilley. Castanet et al. (1996) and Bruce et al. (2002) also reported maximum skeletochronological ages of 11 yr in the medium-size species D. monticola and 15 yr in the larger D. quadramaculatus, in the same assemblages that included D. ocoee. Thus, longevity appears to be correlated with body size in this genus. On demographic criteria, it is expected that age at first reproduction and longevity (or average lifespan) are correlated. Such a trend was evident in the studies of Desmognathus cited above. Marvin (2001) examined the relationship in a small sample of plethodontid species; the correlation was positive, as predicted, but was non-significant statistically. At the intraspecific level, the Life Histories 503 data on geographic variation in Triturus alpestris summarized by Miaud et al. (2000) showed that longevity increases with age at first reproduction. It would be of interest to expand this type of analysis, using comparative methods, given the steady accumulation of age data from skeletochronological studies. 13.3.9 Direct Development The evolution of direct development in salamanders involves the elimination of a free-living larval stage from the life cycle, with metamorphosis completed in the embryonic stage, so that the hatchling that emerges from the egg capsule is a terrestrial juvenile. A majority of all salamander species undergo direct development. All but a handful of these are members of the plethodontid tribes Plethodontini and Bolitoglossini, in which the trait is fixed at the tribal level. Otherwise, direct development occurs in a very few species of desmognathine plethodontids and Old World salamandrids. In the latter taxon it is associated with viviparity. Direct development in salamanders is envisioned as a lifehistory strategy that has facilitated exploitation of the resources of terrestrial environments. It allows for reproduction in a broad range of terrestrial and arboreal habitats, including underground burrows, rotting logs, rock crevices, and epiphytic vegetation. Some direct-developing plethodontids, especially in the genera Aneides, Ensatina, and Hydromantes, have exceptionally large eggs, and this is reflected in lengthy periods of embryogenesis, meroblastic cleavage, and the formation of an embryonic disk on the uncleaved yolk (Collazo 1996; Wake and Hanken 1996). However, factors other than egg size per se, especially genome size, contribute to variation in these traits (Sessions and Larson 1987; Jockusch 1997). From a life-history perspective, it is of interest to consider whether direct development has evolved independently of changes in age at first reproduction in plethodontines and bolitoglossines. For comparison, the relative taxa are the biphasic plethodontids of the Desmognathinae and Hemidactyliini. Data on age at first reproduction in a few species of bolitoglossines (Houck 1977a, b; Salvidio 1993; Wake and Castanet 1995) and 13 species of Plethodon (Marvin 1996), in comparison with similar data for numerous biphasic species of desmognathines (Tilley and Bernardo 1993; Castanet et al. 1996; Bruce et al. 2002) and hemidactyliines (Ryan and Bruce 2000), show that the values for plethodontines and bolitoglossines are contained within the ranges of the latter two taxa. Thus, these data provide no evidence that evolutionary predisplacement of metamorphosis to the embryonic period was accompanied by precocity in sexual maturation. Although there are relatively few data on age at maturation in bolitoglossines, one special situation concerns an unusual report by Vial (1968) on salamanders of the Bolitoglossa subpalmata complex (Garcia-Paris et al. 2000). In estimating growth from mark-recapture data, Vial argued that males attain maturity at 4-9 yr and females at 9-13 yr. If correct, this would mean that evolutionary acceleration in somatic development was associated with delay in reproductive development (or hypermorphosis) in this lineage. However, 504 Reproductive Biology and Phylogeny of Urodela Vial’s conclusions were challenged by Houck (1982), on the basis of laboratory studies of growth in one population of this complex; it was her conclusion that maturity was attained at 1.5 yr in males and 3 yr in females, similar to values reported for plethodontines and other bolitoglossines. Wake and Hanken (1996) proposed that biphasic life cycles in extant amphibians incorporate caenogenetic features, particularly a complex and abrupt metamorphosis, in comparison (citing Schoch 1992) with the more gradual metamorphosis that was apparently a feature of their tetrapod ancestors. Given the validity of this hypothesis, these authors suggested that the evolution of direct development in plethodontids freed these salamanders from constraints evolved through caenogenesis in their biphasic predecessors, and promoted diversification along novel morphogenetic pathways, referred to as “ontogenetic repatterning,” that are associated with heterochronic shifts in developmental timing. Support for the “constraint” hypothesis has been provided by Deban and Marks’s (2002) analysis of the feeding mechanisms of biphasic and direct-developing plethodontids. The evolutionary opportunities available to direct-developing plethodontids are reflected in the extensive adaptive radiation and prolific speciation of the neotropical lineages of the family. 13.4 EVOLUTIONARY ECOLOGY OF URODELAN LIFE HISTORIES Ecological models of biphasic amphibian life histories have emphasized the timing of metamorphosis, considered as an ontogenetic niche shift between two habitats or modes of life, in reference to growth opportunities and survival probabilities in the two niches and the optimal distribution of reproduction over the organism’s life span. Age and size at metamorphosis are key parameters in the amphibian life cycle, as are age and size at first reproduction. However, metamorphosis is a morphogenetic landmark, whereas first reproduction is a demographic landmark that directly contributes to the fitness parameters r and R0. In the development of models of the urodelan life history, age at metamorphosis and age at first reproduction can be allowed to vary independently, given that direct development and paedomorphosis are common derived life-history modes. In contrast, in anurans, where paedomorphosis is unknown and unlikely to evolve (Wassersug 1974), attainment of maturation, of necessity, follows metamorphosis. The seminal paper of Wilbur and Collins (1973) focused on factors eliciting selection on metamorphic size in amphibians. In assuming that in any species there are lower and upper limits on body size at metamorphosis set by the evolutionary history of the taxon, the authors hypothesized that within those limits individual size at metamorphosis is determined by the larva’s body weight and recent growth history, which in turn reflect the quality of the physical and biotic habitat and the competitive milieu involving food and crowding. Their model was based on exacting experimental studies of both urodelan (Ambystoma) and anuran larvae. The model leads to the prediction Life Histories 505 that amphibian species in permanent aquatic habitats will show a narrow range of metamorphic sizes but broad variation in metamorphic age, whereas species adapted to temporary aquatic habitats will metamorphose over a wide range of size and may show a narrow or broad range of ages at metamorphosis. Although largely focused on the biphasic life cycle, Wilbur and Collins (1973) examined diversification of amphibian life cycles in the directions of both paedomorphosis and direct development within the framework of their model. Smith-Gill and Berven (1979) criticized Wilbur and Collins on the grounds that the timing of metamorphosis depends more on the differentiation rate than on growth rate, such that the latter, in itself, is not a good predictor of metamorphic size in some species. Yet it is apparent that Smith-Gill and Berven’s assertion that differentiation rate determines age at metamorphosis borders on the tautological, given that differentiation rate would seem to be equivalent to rate of development to metamorphic climax, especially since the authors used the Taylor-Kollros stages as their index of differentiation rate in anurans. However, their analysis did emphasize that factors other than body size and growth rate determine metamorphic timing. They concluded that temperature is a particularly important proximate factor in regulating growth and differentiation rates, and thus the duration of the larval period in North Temperate anurans and probably in urodeles as well; e.g., as reported for Ambystoma tigrinum by Sexton and Bizer (1978). Smith-Gill and Berven hypothesized that development rate is more sensitive to temperature than growth rate, so that over a temperature gradient involving either elevation or latitude later metamorphosis at larger sizes is expected at lower temperatures. Such a result has been reported by Beachy (1995) in an experimental study of the stream-breeding plethodontid Desmognathus ocoee. However, in another stream-breeding desmognathine, D. fuscus, Juterbock (1990) showed that populations at higher and cooler latitudes metamorphose later but at smaller sizes. The documented variation in both age and size at metamorphosis was minimal in the latter study, as was that of the environmental gradient. More recently, Bernardo and Reagan-Wallin (2002) have challenged the Smith-Gill and Berven hypothesis on the basis of data on variation in metamorphic size with elevation in six species of stream-dwelling plethodontid salamanders. They demonstrated that metamorphic size either decreases (four species) or remains constant (two species) with increasing elevation and cooler temperatures. Bernardo and Reagan-Wallin did not provide data on metamorphic timing for the six species in their study. Nevertheless, these data, like those of the earlier study of Beachy (1995), strengthen the concept that the relationship between environmental temperature regimes and larval growth and developmental rates in salamanders is complex, is confounded by other factors, and may differ between pond- and stream-developing larvae. In a review of complex life cycles, with emphasis on amphibians, Wilbur (1980) extended an argument of the Wilbur and Collins (1973) model, that the biphasic amphibian life cycle is an adaptation that allows the organism to exploit the resources of two environments, wherein the larval stage is 506 Reproductive Biology and Phylogeny of Urodela dedicated to growth and the adult stage to dispersal and reproduction. In order to understand the selective factors involved in maintenance of the biphasic life cycle, Wilbur emphasized the need to determine whether densitydependent regulation of amphibian populations occurs in the larval stage, postmetamorphic stage, or both. At the time his paper was written there had been several studies demonstrating density-dependent regulation in the larval stage, but there were virtually no data on regulation in the adult stage. Wilbur’s (1972) own research had demonstrated density dependence in larvae of several species of Ambystoma, as have later studies of this genus (e.g., Stenhouse et al. 1983; Stenhouse 1985). The effects of density may involve growth, survival, and/or length of the larval period. It was Wilbur’s (1980) contention that the timing of metamorphosis represents a tradeoff between the opportunities for rapid growth in the aquatic environment and the risks of mortality from predation and pond drying. The model is most applicable to pond-breeding salamanders; larvae in stream-breeding species are less constrained in terms of dispersal and habitat drying, growth opportunities are lower, and the niche shift at metamorphosis may be less drastic. For example, density effects on growth and mortality were not demonstrated in a series of experimental studies of plethodontid larvae of several species in mountain streams, wherein predation may be the main regulatory agent (Beachy 1993, 1994, 1997). The latter author (Beachy 1997) suggested that plasticity in metamorphic timing in response to either growth history or predation risks may be less in the more reliable but less productive environments of mountain streams than in ponds. Drawing largely on the arguments presented by Wilbur and Collins (1973), Whiteman (1994) formulated three hypotheses to account for facultative paedomorphosis, and examined the evidence for each. The paedomorph advantage hypothesis proposes that paedomorphs have higher fitness than metamorphs, such advantages stemming from either abiotic or biotic factors. Alternatively, the best-of-a-bad-lot hypothesis predicts that paedomorphosis results from the failure of slower-growing larvae to reach a critical size threshold for survival in the terrestrial environment, and hence maturation is favored in the larval morph rather than continued larval growth at a low rate. Whiteman also suggested, under what he called the dimorphic paedomorph hypothesis, that both mechanisms might operate in a population, based on the magnitude of variation in larval growth rates. He concluded that variation in larval growth and metamorphic patterns in a number of species and populations tend to support either the paedomorph advantage or best-of-abad-lot hypothesis. Werner (1986) expanded the Wilbur and Collins model by incorporating attributes of the terrestrial phase of the life history into a predictive model of optimal metamorphic size in amphibians. It was Werner’s claim that models focusing only on larval growth rates (and other attributes of the larval phase) are incomplete, because factors operating in later stages of the life history influence selection on size at metamorphosis. These factors include, especially, Life Histories 507 growth and mortality in the aquatic habitat, and growth, mortality, and fecundity in the terrestrial habitat. Werner’s model is based on the premises that (1) ecological effects are mediated through size rather than age, (2) sizespecific growth, mortality, and fecundity rates are the parameters needed to specify population growth, and (3) optimal size at metamorphosis is the size that maximizes the instantaneous population growth rate (r). Werner illustrated these relationships in a series of graphical models based on a simplified version of his mathematical formulation of the relationships — namely the ratio of size-specific mortality rate to growth rate, m/g, for the aquatic and terrestrial phases of the life cycle. Under this model, selection promoting a biphasic life cycle requires switching of the m/g curves for the aquatic and terrestrial phases, with optimal size at metamorphosis determined by the size at which the switch occurs. The ecological conditions under which metamorphosis is eliminated from the life history through the evolution of paedomorphosis or direct development involve a shift in either of the m/g curves in order to eliminate the switch. The generality of Werner’s model is illustrated by his evaluation of those species of newts (e.g., Notophthalmus viridescens), which in passing through three post-hatching stages — larva, eft, adult -- have the most complex life histories of any amphibian species. Although Werner recognized that other factors affect the course of the amphibian life history (such factors as temperature and seasonality, hydrologic variability, and the costs in survival and energetics of the metamorphic process itself), the effects of these factors were not explicitly built into the model. Ludwig and Rowe (1990) and especially Rowe and Ludwig (1991) built upon Werner’s model by introducing a time constraint for reproduction, either a fixed time (T) by which individuals must attain the minimal size required for reproduction, as in explosive breeders, or a time span (season within a year), as in prolonged breeders. Time is not equivalent to age in their model because the starting points (T0) are young, premetamorphic individuals whose ages may vary. Thus, the range in body sizes at T0 is due both to variation in date of birth and to random variation in growth rate early in the life cycle. The fitness gains in the aquatic and terrestrial habitats are conjoint functions of growth and survival in each; the goal in optimizing the timing of the switch from the first to the second habitat is that of maximizing the “payoff” (reproduction) at time T. The model does not consider reproduction beyond the first year. Under the conditions of the model, Rowe and Ludwig showed that initial variation in body size within a population leads to sizedependent variation in the timing of metamorphosis; the shape of the switching curve varies according to the differentials between the two habitats in both growth rate and mortality rate. Both the Werner (1986) and Rowe and Ludwig (1991) formulations emphasize that optimization of the timing of metamorphosis in amphibians with biphasic life cycles involves tradeoffs in growth/survivorship probabilities between the two niches, unlike the Wilbur and Collins model (1973) that deals with the dynamics of larval growth and survival only. Rowe and Ludwig (1991), in contrast to Werner (1986), 508 Reproductive Biology and Phylogeny of Urodela introduce a time constraint on reproduction that provides for variation in optimal size and time at metamorphosis according to individual growth trajectories. More recently, Day and Rowe (2002) have re-examined the Wilbur-Collins model, as applied to the timing of either metamorphosis (as in the original model) or maturation (i.e., any major life-history transition). These authors noted that many species of amphibians and other animals show a reduction in either age at metamorphosis or age at maturity, but an increase in body size, with increased conditions for growth. Such norms of reaction are not necessarily predicted by the Wilbur-Collins model. Day and Rowe incorporated developmental thresholds into an optimality model, and examined reaction norms for age and body size of either metamorphosis or maturation. They focused on “overhead” thresholds, defined as the minimum body size at which an individual can meet the baseline costs of either transition, such that benefits accrue only above the threshold size. Variation in threshold size generates reaction norms (Day and Rowe 2002: Fig. 3) that are in accord with much of the empirical and experimental data they cited, including several anuran studies among amphibians. In particular, one common pattern in anurans is that larvae that experience low growth rates due to food limitation metamorphose at older ages and smaller sizes. Such a pattern has also been demonstrated in the salamander Ambystoma barbouri (Petranka 1984b). However, other experimental studies of salamanders have generated contrary results in terms of the response to variation in food supply. Of particular interest are food-limitation experiments by Beachy (1995) on Desmognathus ocoee and O’Laughlin and Harris (2000) on Hemidactylium scutatum. In both experiments increases in the food ration resulted in increases in size at metamorphosis but not in a shortening of the larval period. In the context of the threshold model, such results would require very low threshold sizes at metamorphosis (Day and Rowe 2002: Fig. 3c). Both Beachy and O’Laughlin and Harris suggested that the phylogenetic history of plethodontids (putatively a stream-adapted taxon) may have involved loss of plasticity in metamorphic timing. Day and Rowe’s model may apply to either age at metamorphosis or age at maturation. Sexual maturation and ensuing first reproduction are key transitions in all organisms, regardless of the life-cycle mode, because they involve a major shift in allocation of resources to reproduction, at a cost to growth, maintenance, and storage (Bernardo 1993). Applicable theoretical models depend on the pattern of variation in population parameters and the environment (Stearns 1992; Roff 2002). Depending on these circumstances, either r, the rate of population increase, or R0, the net reproductive rate, may be selected as a fitness measure. The following account is based largely on Roff (2002). The characteristic equation is given as: z w 1= w l( x) m(x)e–rxdx, x =a or, alternatively, 1 = Â l( x) m (x)e–rx, x=a where a = age at first reproduction, w = age at last reproduction, x = age, l(x) = survival to age x, and m(x) = fecundity at age x. In a stationary population, Life Histories 509 where r = 0, the characteristic equation can be written: z w R0 = w l( x ) m(x)dx, or R0 = Â l( x) m(x). x=a x=a Depending on whether the population is increasing/decreasing or stationary, optimal age at first reproduction is the age that maximizes either r or R0. This is the peak of the graph of r or R0 plotted against age, and thus is the age where ∂r/∂a or ∂R0/∂a = 0. As these maxima do not necessarily coincide, the correct choice of a fitness measure is critically important (Roff 2002: Fig. 4.23). The principal application of this general approach to the study of a salamander life history is Kusano’s (1980, 1981, 1982) investigation of the life history and population dynamics of the hynobiid Hynobius nebulosus in Japan. This author combined mark-recapture methods in the field with experimental studies of growth in the laboratory to estimate key demographic parameters in a population of this species. Although Kusano’s study extended over a 7-yr period only, his data suggested a relatively stationary population, but with severe annual fluctuations in larval survival, ranging from 0.002 to 0.107 during a larval period of about 0.5 yr. His study also generated survivorship values for postmetamorphic individuals (mean = 0.7 annually), but the data for females especially were of uncertain reliability. Kusano (1982) converted size-fecundity data for females to age-fecundity values based on the growth data. Age at first reproduction was estimated as 4 yr in males and 5 yr in females. Kusano (1982) selected r as the fitness measure, and calculated values for this parameter using the equation: w 1= Â l( x) m(x) e-rx. x=a In solving the equation for the calculated schedules of survival and fecundity, Kusano determined that r was maximized at a = 5 yr in females, which is the value of a determined directly from empirical data (Kusano 1982: Fig. 8). The close match is somewhat surprising, given the uncertainties of the female survivorship data. By substituting alternative survival schedules, Kusano concluded that increasing premetamorphic survival generates selection for earlier maturation, whereas increasing survival after metamorphosis generates selection for later maturation. This is an expected result for some categories of populations wherein mortality rates vary with age and body size, as opposed to morphogenetic stage (Roff 2002). That is, a decrease in adult survival generates selection for an increase in reproductive effort in earlier age classes (lower age at first reproduction), whereas a decrease in survival of prereproductives generates selection for a decrease in reproductive effort (greater age at first reproduction) in later age classes (Murphy 1968; Michod 1979). It remains problematical whether salamanders with biphasic life cycles, like Hynobius nebulosus, where mortality rates and their variances vary between larval and postmetamorphic stages, and where the differences may be largely determined by stage rather than age or size, meet the requirements of the 510 Reproductive Biology and Phylogeny of Urodela above model. Under a stage-dependent model, a decrease in postmetamorphic survival should result in selection to prolong the larval (growth) stage and thus delay maturation, whereas decreased larval survival should elicit selection for early metamorphosis and thus early maturation. The latter scenario would require the dependence of maturation on metamorphosis, which may in fact be negligible in many salamander taxa. Nevertheless, it is conceivable that the effects of selection on age at first reproduction and reproductive effort in salamanders differ between species with monophasic and biphasic life cycles. Kusano’s method of analysis could be extended to other species where agespecific survival and fecundity data have been generated; e.g., life tables for various species (Bell 1977; Tilley 1980; Hairston 1983; Bruce 1988b). However, given the wide range in age at first reproduction in many salamander species, as documented earlier, it is necessary to plan demographic studies for a sufficient term to adequately determine the degree of stability in the population and its environment. Hairston (1987) has proposed that the evaluation of population stability requires a minimal study period of one generation, which is often several years in salamanders. It is also desirable to extend the analysis to two or more populations that represent known variation in the parameter in question, as, for example, the two populations of Desmognathus ocoee investigated by Tilley (1980) and Bernardo (1994). 13.5 CONCLUSIONS The occurrence of metamorphosis, paedomorphosis, and direct development in salamanders is correlated with a lack of morphogenetic and morphological specialization, in comparison with vertebrates generally, and with anurans and caecilians specifically, in a taxon in which the ancestral life history was biphasic. Within the Amphibia, whereas biphasic life cycles and direct development are found in anurans, caecilians, and urodelans, only the last have exploited the paedomorphic mode of life. One set of morphological features that undoubtedly played a significant role in amphibian life-history diversification is the variety of respiratory structures found in this taxon: gills, lungs, buccopharynx, and skin are variously used in gas exchange (Boutilier et al. 1992; Shoemaker et al. 1992). Evolution of a thin, glandular, vascularized skin provided a major respiratory organ that functions well in both aquatic and terrestrial environments. Amphibians, including salamanders, generally lack the specializations (i.e., bony scales of the dermis and keratinized derivatives of the epidermis) that preclude a major respiratory role for the skin in other vertebrates, with the noteworthy exceptions of many fishes, especially air breathers, and various aquatic reptiles (Feder and Burggren 1985; Graham 1997). Wassersug (1974) explained convincingly how ecological and morphological specializations constrain the evolution of paedomorphosis in anurans; thus, the considerable degree of life-history diversification in the latter taxon is limited to elaboration of the biphasic and direct-development modes. Life Histories 511 Whether similar constraints operate on caecilians is unknown; however, the developmental pattern of biphasic caecilians, involving hatching at a relatively advanced stage (Breckenridge et al. 1987), might predispose caecilians to selection for direct development. From a macroevolutionary perspective, i.e., the fixation of paedomorphosis and direct development in several higher taxa, it is obvious that age at metamorphosis and age at maturation have been relatively free to evolve independently in salamanders. This conclusion also follows from microevolutionary studies of variation in metamorphic and maturational schedules in some species that exhibit facultative paedomorphosis (e.g., Ryan and Semlitsch 1998). Facultative direct development is a much rarer occurrence, but has been documented in the salamandrid Salamandra salamandra (Joly 1986; Thiesmeier and Haker 1990; Thiesmeier 1994; Thiesmeier et al. 1994). Evolutionary independence of these key life-history transitions suggests independence of the underlying physiological and endocrinological mechanisms that regulate metamorphosis and maturation in salamanders. Direct development is a prevalent life-cycle mode in one family only, the Plethodontidae, where it is correlated with adaptive radiations yielding a majority of all salamander species (Wake and Hanken 1996). The most diverse radiation has occurred in one lineage, the bolitoglossines, and is associated with expansion deep into the Neotropics, where salamanders otherwise are absent. Conceptually, direct development may represent an outcome of acceleration in the rate of somatic development (a category of peramorphosis), involving predisplacement (earlier onset) of metamorphosis, which has eliminated caenogenetic features that trace back to the biphasic life cycles of ancestral urodelans. An important unresolved question is how metamorphosis during embryogenesis in direct-developing taxa is regulated by the endocrine system. A very different mode of direct development has evolved in the salamandrid genus Salamandra. It can be argued that the most derived life histories of salamanders are found in this taxon, wherein remarkable discontinuities have arisen, first, between other salamandrids and various forms of the S. salamandra complex, and, secondly, between the latter and the obligate pueriparous species S. atra, S. lanzai, and S. luschani. The extreme life-history traits shown by the members of the latter group include long gestation periods of 2-4 yr, tiny broods of two offspring, and lengthy life spans. It would be of great interest to relate the variation in these traits to the distribution of survivorship over the life spans of these species. In the species-rich families of salamanders with biphasic life cycles, the pond-breeding niche shared by hynobiids and salamandrids in the Old World is occupied primarily by ambystomatids in the New World, with only the widespread salamandrid Notophthalmus viridescens showing the adaptive flexibility to successfully coexist with ambystomatids. The stream-breeding niche occupied by other members of the same Old World families is shared by plethodontids, dicamptodontids, rhyacotritonids, and some members of other 512 Reproductive Biology and Phylogeny of Urodela families in the New World. In all of these families, among related taxa having biphasic life cycles, larval periods tend to be shorter in pond-breeding than in stream-breeding populations or species, and in low- versus high-elevation populations/species. These differences are reflected, generally, in lower ages at first reproduction and shorter mean generation times in pond/lowland versus stream/upland forms. This is surely related to higher temperatures and/or greater productivity in lowland and/or pond habitats in comparison to those of uplands and/or streams. An interesting and largely unresolved question is that of population regulation in pond versus stream species, in the context of maintenance and evolution of biphasic cycles, as postulated in the models of Wilbur and Collins (1973), Werner (1986), Rowe and Ludwig (1991), and Day and Rowe (2002). The resolution of this question will require much more comprehensive and long-term studies than have yet been attempted. Variation in age and body size at first reproduction, generation time, and life expectancy is extreme among species and within many species of salamanders, regardless of life-history mode. Some of the intraspecific variation is correlated with latitude and altitude, and represents either direct environmental effects on growth and development or an adaptive response to the environmental gradient. In most species exhibiting such variation, the genetic contribution to the response is unknown. The recent development of skeletochronological methodology is providing opportunities for examining more precisely the relationships among these and other life-history and demographic parameters. Preliminary assessments of such variation have been attempted at both the intra- and interspecific level (e. g., Castanet et al. 1996; Bruce 1996; Miaud et al. 2000; Olgun et al. 2001; Bruce et al. 2002). The integration of these types of studies with manipulative experimentation (e.g., common gardens) represents an approach for determining the genetic basis of the observed variation in these traits. Such a research program can provide a framework for phylogenetic analysis of life-history variation using comparative methods. 13.6 ACKNOWLEDGMENTS This chapter could not have been completed without the outstanding services provided by the staff of Hunter Library at Western Carolina University. Brenda Moore, in particular, was unrelenting in obtaining essential books and papers. The chapter has benefited from an incisive reading of an earlier version by Christopher K. Beachy. Burkhard Thiesmeier was unstinting in helping me obtain several books and papers on European salamanders. 13.7 LITERATURE CITED Alberch, P., Gould, S.J., Oster, G.F. and Wake, D.B. 1979. Size and shape in ontogeny and phylogeny. Paleobiology 5: 296-317. Anderson, J.D. 1961. The life history and systematics of Ambystoma rosaceum. Copeia 1961: 371-377. Life Histories 513 Anderson, J.D. 1967. A comparison of the life histories of coastal and montane populations of Ambystoma macrodactylum in California. American Midland Naturalist 77: 323-355. Anderson, J.D. and Graham, R.E. 1967. Vertical migration and stratification of larval Ambystoma. Copeia 1967: 371-374. Anderson, J.D., Hassinger, D.D. and Dalrymple, G.H. 1971. Natural mortality of eggs and larvae of Ambystoma t. tigrinum. Ecology 52: 1107-1112. Anderson, J.D. and Williamson, G.K. 1974. Nocturnal stratification in larvae of the mole salamander, Ambystoma talpoideum. Herpetologica 30: 28-29. Anderson, J.D. and Worthington, R.D. 1971. The life history of the Mexican salamander Ambystoma ordinarium Taylor. Herpetologica 27: 165-176. Arntzen, J.W. 2000. A growth curve for the newt Triturus cristatus. Journal of Herpetology 34: 227-232. Arntzen, J.W. and Teunis, S.F.M. 1993. A six year study on the population dynamics of the crested newt (Triturus cristatus) following the colonization of a newly created pond. Herpetological Journal 3: 99-110. Ash, A.N., Bruce, R.C., Castanet, J. and Francillon-Vieillot, H. 2003. Population parameters of Plethodon metcalfi on a ten year old clearcut and in nearby forest in the southern Blue Ridge Mountains. Journal of Herpetology (in press). Ashton, R.E., Jr. and Braswell, A.L. 1979. Nest and larvae of the Neuse River waterdog, Necturus lewisi (Brimley) (Amphibia: Proteidae). Brimleyana 1: 15-22. Austin, R.M., Jr. and Camp, C.D. 1992. Larval development of black-bellied salamanders, Desmognathus quadramaculatus, in northeastern Georgia. Herpetologica 48: 313-317. Bannikov, A.G. 1949. Notes on the biology of Ranodon sibiricus. Doklady Akademiia Nauk USSR 65: 237-240. [English language version translated by E. Levi, edited by F. B. Turner] Beachy, C.K. 1993. Guild structure in streamside salamander communities: a test for interactions among larval plethodontid salamanders. Journal of Herpetology 27: 465-468. Beachy, C.K. 1994. Community ecology in streams: effects of two species of predatory salamanders on a prey species of salamander. Herpetologica 50: 129-136. Beachy, C.K. 1995. Effects of larval growth history on metamorphosis in a streamdwelling salamander (Desmognathus ochrophaeus). Journal of Herpetology 29: 375-382. Beachy, C.K. 1997. Effects of predatory larval Desmognathus quadramaculatus on growth, survival and metamorphosis of larval Eurycea wilderae. Copeia 1997: 131-137. Beachy, C.K. and Bruce, R.C. 2003. Miniaturization is due to accelerated maturation in the large plethodontid Desmognathus quadramaculatus. Amphibia-Reptilia (in press). Bell, G. 1977. The life of the smooth newt (Triturus vulgaris) after metamorphosis. Ecological Monographs 47: 279-299. Bell, G. and Lawton, J.H. 1975. The ecology of the eggs and larvae of the smooth newt (Triturus vulgaris (Linn.)). Journal of Animal Ecology 44: 393-423. Bernardo, J. 1993. Determinants of maturation in animals. Trends in Ecology and Evolution 8: 166-173. Bernardo, J. 1994. Experimental analysis of allocation in two divergent, natural salamander populations. American Naturalist 143: 14-38. #" Reproductive Biology and Phylogeny of Urodela Bernardo, J. and Reagan-Wallin, N.L. 2002. Plethodontid salamanders do not conform to “general rules” for ectotherm life histories: insights from allocation models about why simple models do not make accurate predictions. Oikos 97: 398-414. Besharse, J.C. and Holsinger, J.R. 1977. Gyrinophilus subterraneus, a new troglobitic salamander from southern West Virginia. Copeia 1977: 624-634. Birchfield, G.L. and Bruce, R.C. 2000. Morphometric variation among larvae of four species of lungless salamanders (Caudata: Plethodontidae). Herpetologica 56: 332-342. Bishop, S.C. 1941. The salamanders of New York. New York State Museum Bulletin (324): 1-365. Blanchard, F.N. 1923. The life history of the four-toed salamander. American Naturalist 57: 262-268. Boisseau, C. and Joly, J. 1975. Transport and survival of spermatozoa in female Amphibia. Pp. 94-104. In E. S. E. Hafez and C. G. Thibauld (eds), The Biology of Spermatozoa: Transport, Survival, and Fertilizing Ability. S. Karger AG, Basel, Switzerland. Boutilier, R.G., Stiffler, D.F. and Toews, D.P. 1992. Exchange of respiratory gases, ions, and water in amphibious and aquatic amphibians. Pp. 81-124. In M. E. Feder and W. W. Burggren (eds), Environmental Physiology of the Amphibians. University of Chicago Press, Chicago, Illinois. Branch, L.C. and Altig, R. 1981. Nocturnal stratification of three species of Ambystoma larvae. Copeia 1981: 870-873. Brandon, R.A. and Bremer, D.J. 1966. Neotenic newts, Notophthalmus viridescens louisianensis, in southern Illinois. Herpetologica 22: 213-217. Breckenridge, W.R., Nathanael, S. and Pereira, L. 1987. Some aspects of the biology and development of Ichthyophis glutinosus (Amphibia: Gymnophiona). Journal of Zoology, London 211: 437-449. Breitenbach, G.L. 1982. The frequency of communal nesting and solitary brooding in the salamander, Hemidactylium scutatum. Journal of Herpetology 16: 341-346. Brodie, E.D., Jr. 1968a. Investigations on the skin toxin of the red-spotted newt, Notophthalmus viridescens viridescens. American Midland Naturalist 80: 276-280. Brodie, E.D., Jr. 1968b. Investigations on the skin toxin of the adult rough-skinned newt, Taricha granulosa. Copeia 1968: 307-313. Bruce, R.C. 1969. Fecundity in primitive plethodontid salamanders. Evolution 23: 50-54. Bruce, R.C. 1971. Life cycle and population structure of the salamander Stereochilus marginatus in North Carolina. Copeia 1971: 234-246. Bruce, R.C. 1972. The larval life of the red salamander, Pseudotriton ruber. Journal of Herpetology 6: 43-51. Bruce, R.C. 1974. Larval development of the salamanders Pseudotriton montanus and P. ruber. American Midland Naturalist 92: 173-190. Bruce, R.C. 1976. Population structure, life history and evolution of paedogenesis in the salamander Eurycea neotenes. Copeia 1976: 242-249. Bruce, R.C. 1978. A comparison of the larval periods of Blue Ridge and Piedmont mud salamanders (Pseudotriton montanus). Herpetologica 34: 325-332. Bruce, R.C. 1980. A model of the larval period of the spring salamander, Gyrinophilus porphyriticus, based on size-frequency distributions. Herpetologica 36: 78-86. Life Histories 515 Bruce, R.C. 1988a. Life history variation in the salamander Desmognathus quadramaculatus. Herpetologica 44: 218-227. Bruce, R.C. 1988b. An ecological life table for the salamander Eurycea wilderae. Copeia 1988: 15-26. Bruce, R.C. 1993. Sexual size dimorphism in desmognathine salamanders. Copeia 1993: 313-318. Bruce, R.C. 1996. Life-history perspective of adaptive radiation in desmognathine salamanders. Copeia 1996: 783-790. Bruce, R.C. 1998. Nesting habits, eggs, and hatchlings of the salamander Nototriton picadoi (Plethodontidae: Bolitoglossini). Herpetologica 54: 13-18. Bruce, R.C. 1999. Life history attributes of a rare neotropical salamander, Nototriton picadoi (Plethodontidae: Bolitoglossini). Herpetological Review 30: 76-78. Bruce, R.C. 2000. Sexual size dimorphism in the Plethodontidae. Pp. 243-260. In R. C. Bruce, R. G. Jaeger and L. D. Houck (eds), The Biology of Plethodontid Salamanders. Kluwer Academic/Plenum Publishers, New York, NY. Bruce, R.C., Castanet, J. and Francillon-Vieillot, H. 2002. Skeletochronological analysis of variation in age structure, body size, and life history in three species of desmognathine salamanders. Herpetologica 58: 181-193. Bruce, R.C. and Hairston, N.G., Sr. 1990. Life-history correlates of body-size differences between two populations of the salamander, Desmognathus monticola. Journal of Herpetology 24: 126-134. Burggren, W.W. and Just, J.L. 1992. Developmental changes in physiological systems. Pp. 467-530. In M. E. Feder and W. W. Burggren (eds), Environmental Physiology of the Amphibians. University of Chicago Press, Chicago, Illinois. Caetano, M.H. and Castanet, J. 1993. Variability and microevolutionary patterns in Triturus marmoratus from Portugal: age, size, longevity and individual growth. Amphibia-Reptilia 14: 117-129. Caetano, M.H., Castanet, J. and Francillon, H. 1985. Détermination de l’âge de Triturus marmoratus marmoratus (Latreille 1800) du Parc National de Peneda Gerês (Portugal) par squelettochronologie. Amphibia-Reptilia 6: 117-132. Caetano, M.H. and Leclair, R., Jr. 1996. Growth and population structure of redspotted newts (Notophthalmus viridescens) in permanent lakes of the Laurentian Shield, Quebec. Copeia 1996: 866-874. Camp, C.D., Marshall, J.L. and Austin, R.M., Jr. 2000. The evolution of adult body size in black-bellied salamanders (Desmognathus quadramaculatus complex). Canadian Journal of Zoology 78: 1712-1722. Carroll, R.L. 1988. Vertebrate Paleontology and Evolution. W. H. Freeman and Company, New York, NY. 698 pp. Castanet, J., Francillon-Vieillot, H. and Bruce, R.C. 1996. Age estimation in desmognathine salamanders assessed by skeletochronology. Herpetologica 52: 160-171. Castanet, J. and Smirina, E. 1990. Introduction to the skeletochronological method in amphibians and reptiles. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 11: 191-196. Chadwick, C.S. 1944. Observations on the life cycle of the common newt in western North Carolina. American Midland Naturalist 32: 491-494. Chippindale, P.T., Price, A.H., Wiens, J.J. and Hillis, D.M. 2000. Phylogenetic relationships and systematic revision of central Texas hemidactyliine plethodontid salamanders. Herpetological Monographs 14: 1-80. 516 Reproductive Biology and Phylogeny of Urodela Clergue-Gazeau, M. and Beetschen, J.C. 1966. Contribution à l’étude de la reproduction de l’urodèle Euproctus asper (Dugês) dans les Pyrénées centrales et ariégeoises. Annales de Limnologie 2: 217-226. Collazo, A. 1996. Evolutionary correlations between early development and life history in plethodontid salamanders and teleost fishes. American Zoologist 36: 116-131. Collazo, A. and Marks, S.B. 1994. Development of Gyrinophilus porphyriticus: identification of the ancestral developmental pattern in the salamander family Plethodontidae. Journal of Experimental Zoology 268: 239-258. Collins, J.P. 1981. Distribution, habitats and life history variation in the tiger salamander, Ambystoma tigrinum, in east-central and southeast Arizona. Copeia 1981: 666-675. Crump, M.L. 1995. Parental care. Pp. 518-567. In H. Heatwole and B. K. Sullivan (eds), Amphibian Biology, Volume 2, Social Behaviour. Surrey Beatty and Sons, Chipping Norton, Australia. Crump, M.L. 1996. Parental care among the Amphibia. Advances in the Study of Behavior 25: 109-144. Day, T. and Rowe, L. 2002. Developmental thresholds and the evolution of reaction norms for age and size at life-history transitions. American Naturalist 159: 338-350. Deban, S.M. and Marks, S.B. 2002. Metamorphosis and evolution of feeding behaviour in salamanders of the family Plethodontidae. Zoological Journal of the Linnean Society 134: 375-400. de Beer, G.R. 1930. Embryology and Evolution. Clarendon Press, Oxford, U.K. 116 pp. de Beer, G.R. 1940. Embryos and Ancestors. Clarendon Press, Oxford, U.K. 108 pp. Denver, R.J. 1997. Proximate mechanisms of phenotypic plasticity in amphibian metamorphosis. American Zoologist 37: 172-184. Denver, R.J. and Licht, P. 1989. Neuropeptide stimulation of thyrotropin secretion in the larval bullfrog: evidence for a common neuroregulator of thyroid and interrenal activity in metamorphosis. Journal of Experimental Zoology 252: 101-104. Díaz-Paniagua, C. 1998. Reproductive dynamics of a population of small marbled newts (Triturus marmoratus pygmaeus) in south-western Spain. Herpetological Journal 8: 93-98. Díaz-Paniagua, C., Mateo, J.A. and Andreu, A.C. 1996. Age and size structure of populations of small marbled newts (Triturus marmoratus pygmaeus) from Doñana National Park (SW Spain). A case of dwarfism among dwarfs. Journal of Zoology, London 239: 83-92. Dolmen, D. 1983. Growth and size of Triturus vulgaris and T. cristatus (Amphibia) in different parts of Norway. Holarctic Ecology 6: 356-371. Dopazo, H.J. and Korenblum, M. 2000. Viviparity in Salamandra salamandra (Amphibia: Salamandridae): adaptation or exaptation? Herpetologica 56: 144-152. Duellman, W. E. and Trueb, L. 1986. Biology of Amphibians. McGraw-Hill Book Company, New York, NY. 670 pp. Dunham, A.E., Grant, B.W. and Overall, K.L. 1989. Interfaces between biophysical and physiological ecology and the population ecology of terrestrial vertebrate ectotherms. Physiological Zoology 62: 335-355. Eagleson, G.W. 1976. A comparison of the life histories and growth patterns of populations of the salamander Ambystoma gracile (Baird) from permanent low-altitude and montane lakes. Canadian Journal of Zoology 54: 2098-2111. Life Histories 517 Ehmcke, J. and Bolaños, F. 1998. Brutpflege bei Bolitoglossa subpalmata. Salamandra 34: 223-226. Farner, D.S. and Kezer, J. 1953. Notes on the amphibians and reptiles of Crater Lake National Park. American Midland Naturalist 50: 448-462. Feder, M.E. and Burggren, W.W. 1985. Cutaneous gas exchange in vertebrates: design, patterns, control and implications. Biological Reviews 60: 1-45. Flageole, S. and Leclair, R., Jr. 1992. Étude démographique d’une population de salamandres (Ambystoma maculatum) à l’aide de la méthode squelettochronologique. Canadian Journal of Zoology 70: 740-749. Forester, D.C. and Lykens, D.V. 1991. Age structure in a population of red-spotted newts from the Allegheny Plateau of Maryland. Journal of Herpetology 25: 373376. Fox, H. 1984. Amphibian Morphogenesis. The Humana Press Inc., Clifton, New Jersey. 301 pp. Francillon-Vieillot, H., Arntzen, J.W. and Géraudie, J. 1990. Age, growth and longevity of sympatric Triturus cristatus, T. marmoratus and their hybrids (Amphibia, Urodela): a skeletochronological comparison. Journal of Herpetology 24: 13-22. Garcia-Paris, M., Good, D.A., Parra-Olea, G. and Wake, D.B. 2000. Biodiversity of Costa Rican salamanders: implications of high levels of genetic differentiation and phylogeographic structure for species formation. Proceedings of the National Academy of Sciences, USA 97: 1640-1647. Gill, D.E. 1978. The metapopulation ecology of the red-spotted newt, Notophthalmus viridescens (Rafinesque). Ecological Monographs 48: 145-166. Godley, J.S. 1983. Observations on the courtship, nests and young of Siren intermedia in southern Florida. American Midland Naturalist 110: 215-219. Good, D.A. and Wake, D.B. 1993. Systematic studies of the Costa Rican moss salamanders, genus Nototriton, with descriptions of three new species. Herpetological Monographs 7: 131-159. Gould, S.J. 1977. Ontogeny and Phylogeny. Harvard University Press, Cambridge, Massachusetts. 501 pp. Gould, S.J. 2000. Of coiled oysters and big brains: how to rescue the terminology of heterochrony, now gone astray. Evolution and Development 2: 241-248. Graham, J.B. 1997. Air-Breathing Fishes: Evolution, Diversity, and Adaptation. Academic Press, San Diego, California. 299 pp. Greven, H. 2002. The urodele oviduct and its secretions in and after G. von Wahlert’s doctoral thesis “Eileter, Laich und Kloake der Salamandriden.” Bonner Zoologische Monographien 50: 25-61. Grossenbacher, K. and Thiesmeier, B. (eds). 1999. Handbuch der Reptilien und Amphibien Europas, Band 4/I, Schwanzlurche (Urodela) I. AULA-Verlag, Wiesbaden, Germany. 405 pp. Guex, G-D. and Greven, H. 1994. Structural and physiological aspects of viviparity in Salamandra atra. Pp. 161-208. In H. Greven and B. Thiesmeier (eds), Biology of Salamandra and Mertensiella. Mertensiella Supplement 4, DGHT, Bonn, Germany. Hagström, T. 1977. Growth studies and ageing methods for adult Triturus vulgaris L. and T. cristatus Laurenti (Urodela, Salamandridae). Zoologica Scripta 6: 61-68. Hagström, T. 1979. Population ecology of Triturus cristatus and T. vulgaris (Urodela) in SW Sweden. Holarctic Ecology 2: 108-114. Hairston, N.G. 1983. Growth, survival and reproduction of Plethodon jordani: trade-offs between selective pressures. Copeia 1983: 1024-1035. 518 Reproductive Biology and Phylogeny of Urodela Hairston, N.G., Sr. 1987. Community Ecology and Salamander Guilds. Cambridge University Press, Cambridge, U.K. 230 pp. Harris, R.N. 1987. Density-dependent paedomorphosis in the salamander Notophthalmus viridescens dorsalis. Ecology 68: 705-712. Harris, R.N., Alford, R.A. and Wilbur, H.M. 1988. Density and phenology of Notophthalmus viridescens dorsalis in a natural pond. Herpetologica 44: 234-242. Harris, R.N. and Gill, D.E. 1980. Communal nesting, brooding behavior, and embryonic survival of the four-toed salamander Hemidactylium scutatum. Herpetologica 36: 141-144. Harris, R.N., Semlitsch, R.D., Wilbur, H.M. and Fauth, J.E. 1990. Local variation in the genetic basis of paedomorphosis in the salamander Ambystoma talpoideum. Evolution 44: 1588-1603. Hassinger, D.D., Anderson, J.D. and Dalrymple, G.H. 1970. The early life history and ecology of Ambystoma tigrinum and Ambystoma opacum in New Jersey. American Midland Naturalist 84: 474-495. Hayes, T.B. 1997. Steroids as potential modulators of thyroid hormone activity in anuran metamorphosis. American Zoologist 37: 185-194. Healy, W.R. 1970. Reduction of neoteny in Massachusetts populations of Notophthalmus viridescens. Copeia 1970: 578-581. Healy, W.R. 1973. Life history variation and the growth of juvenile Notophthalmus viridescens from Massachusetts. Copeia 1973: 641-647. Healy, W.R. 1974. Population consequences of alternative life histories in Notophthalmus v. viridescens. Copeia 1974: 221-229. Healy, W.R. 1975. Breeding and postlarval migrations of the red-spotted newt, Notophthalmus viridescens, in Massachusetts. Ecology 56:673-680. Hom, C.L. 1987. Control theory predictions of reproductive allocation in female dusky salamanders. Journal of Mathematical Biology 25:289-306. Hom, C.L. 1988. Optimal reproductive allocation in female dusky salamanders: a quantitative test. American Naturalist 131:71-90. Houck, L.D. 1977a. Life history patterns and reproductive biology of neotropical salamanders. Pp. 43-72. In D. H. Taylor and S. I. Guttman (eds), The Reproductive Biology of Amphibians. Plenum Press, New York, NY. Houck, L.D. 1977b. Reproductive biology of a neotropical salamander, Bolitoglossa rostrata. Copeia 1977:70-83. Houck, L.D. 1982. Growth rates and age at maturity for the plethodontid salamander Bolitoglossa subpalmata. Copeia 1982:474-478. Houck, L.D. and Schwenk, K. 1984. The potential for long-term sperm competition in a plethodontid salamander. Herpetologica 40:410-415. Houck, L.D., Tilley, S.G. and Arnold, S.J. 1985. Sperm competition in a plethodontid salamander: preliminary results. Journal of Herpetology 19:420-423. Howard, J.H. and Wallace, R.L. 1985. Life history characteristics of populations of the long-toed salamander (Ambystoma macrodactylum) from different altitudes. American Midland Naturalist 113: 361-373. Hurlbert, S.H. 1969. The breeding migrations and interhabitat wandering of the vermilion-spotted newt Notophthalmus viridescens (Rafinesque). Ecological Monographs 39: 465-488. Husting, E.L. 1965. Survival and breeding structure in a population of Ambystoma maculatum. Copeia 1965: 352-362. Life Histories 519 Iwasaki, F. and Wakahara, M. 1999. Adaptable larval life histories in different populations of the salamander, Hynobius retardatus, living in various habitats. Zoological Science (Tokyo) 16: 667-674. Jackson, M.E. and Semlitsch, R.D. 1993. Paedomorphosis in the salamander Ambystoma talpoideum: effects of a fish predator. Ecology 74: 342-350. Jockusch, E.L. 1997. An evolutionary correlate of genome size change in plethodontid salamanders. Proceedings of the Royal Society, London B 264: 597-604. Jockusch, E.L. and Mahoney, M.J. 1997. Communal oviposition and lack of parental care in Batrachoseps nigriventris (Caudata: Plethodontidae) with a discussion of the evolution of breeding behavior in plethodontid salamanders. Copeia 1997: 697-705. Joly, J. 1968. Données écologiques sur la salamandre tachetée Salamandra salamandra (L.). Annales des Sciences Naturelles, Zoologie, Paris, 12e Série 10: 301-366. Joly, J. 1986. La reproduction de la salamandre terrestre (Salamandra salamandra L.). Pp. 471–486. In P.P. Grasse and M. Delsol (eds), Traité de Zoologie, Amphibiens, Tome 14. Masson, Paris, France. Joly, J., Chesnel, F. and Boujard, D. 1994. Biological adaptations and reproductive strategies in the genus Salamandra. Pp. 255-269. In H. Greven and B. Thiesmeier (eds.), Biology of Salamandra and Mertensiella. Mertensiella Supplement 4, DGHT, Bonn, Germany. Jørgensen, C.B. 1992. Growth and reproduction. Pp. 439-466. In M. E. Feder and W. W. Burggren (eds), Environmental Physiology of the Amphibians. University of Chicago Press, Chicago, Illinois. Juterbock, J.E. 1978. Sexual dimorphism and maturity characteristics of three species of Desmognathus (Amphibia, Urodela, Plethodontidae). Journal of Herpetology 12: 217-230. Juterbock, J.E. 1990. Variation in larval growth and metamorphosis in the salamander Desmognathus fuscus. Herpetologica 46: 291-303. Kalezic, M.L., Cvetkovic, D., Djorovic, A. and Dzukic, G. 1994. Paedomorphosis and differences in life-history traits of two neighbouring crested newt (Triturus carnifex) populations. Herpetological Journal 4: 151-158. Kawamichi, T. and Ueda, H. 1998. Spawning at nests of extra-large males in the giant salamander Andrias japonicus. Journal of Herpetology 32: 133-136. Kezer, J. and Farner, D.S. 1955. Life history patterns of the salamander Ambystoma macrodactylum in the high Cascade Mountains of southern Oregon. Copeia 1955: 127-131. Krenz, J.D. and Sever, D.M. 1995. Mating and oviposition in paedomorphic Ambystoma talpoideum precedes the arrival of terrestrial males. Herpetologica 51: 387-393. Kusano, T. 1980. Breeding and egg survival of a population of a salamander, Hynobius nebulosus tokyoensis Tago. Researches on Population Ecology 21: 181-196. Kusano, T. 1981. Growth and survival rate of the larvae of Hynobius nebulosus tokyoensis Tago (Amphibia, Hynobiidae). Researches on Population Ecology 23: 360-378. Kusano, T. 1982. Postmetamorphic growth, survival, and age at first reproduction of the salamander, Hynobius nebulosus tokyoensis Tago in relation to a consideration on the optimal timing of first reproduction. Researches on Population Ecology 24: 329-344. Lima, V., Arntzen, J.W. and Ferrand, N.M. 2001. Age structure and growth pattern in two populations of the golden-striped salamander Chioglossa lusitanica (Caudata, Salamandridae). Amphibia-Reptilia 22: 55-68. 520 Reproductive Biology and Phylogeny of Urodela Ludwig, D. and Rowe, L. 1990. Life-history strategies for energy gain and predator avoidance under time constraints. American Naturalist 135: 686-707. Marunouchi, J., Ueda, H. and Ochi, O. 2000. Variation in age and size among breeding populations at different altitudes in the Japanese newts, Cynops pyrrhogaster. Amphibia-Reptilia 21: 381-396. Marvin, G.A. 1996. Life history and population characteristics of the salamander Plethodon kentucki with a review of Plethodon life histories. American Midland Naturalist 136: 385-400. Marvin, G.A. 2001. Age, growth, and long-term site fidelity in the terrestrial plethodontid salamander Plethodon kentucki. Copeia 2001: 108-117. Massey, A. 1990. Notes on the reproductive ecology of red-spotted newts (Notophthalmus viridescens). Journal of Herpetology 24: 106-107. McDiarmid, R.W. and Worthington, R.D. 1970. Concerning the reproductive habits of tropical plethodontid salamanders. Herpetologica 26: 57-70. McKinney, M.L. and McNamara, K.J. 1991. Heterochrony: the Evolution of Ontogeny. Plenum Press, New York, NY. 437 pp. Miaud, C., Andreone, F., Ribéron, A., De Michelis, S., Clima, V., Castanet, J., Francillon-Vieillot, H. and Guyétant, R. 2001. Variations in age, size at maturity and gestation duration among two neighbouring populations of the alpine salamander (Salamandra lanzai). Journal of Zoology, London 254: 251-260. Miaud, C., Guyétant, R. and Faber, H. 2000. Age, size, and growth of the Alpine newt, Triturus alpestris (Urodela: Salamandridae), at high altitude and a review of life-history trait variation throughout its range. Herpetologica 56: 135-144. Miaud, C., Joly, P. and Castanet, J. 1993. Variation in age structures in a subdivided population of Triturus cristatus. Canadian Journal of Zoology 71: 1874-1879. Michod, R.E. 1979. Evolution of life histories in response to age-specific mortality factors. American Naturalist 113: 531-550. Misawa, Y. and Matsui, M. 1997. Larval life history variation in two populations of the Japanese salamander Hynobius kimurae (Amphibia, Urodela). Zoological Science (Tokyo) 14: 257-262. Montori, A. 1990. Skeletochronological results in the pyrenean newt Euproctus asper (Dugès, 1852) from one prepyrenean population. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 11: 209-211. Murphy, G.I. 1968. Pattern in life history and the environment. American Naturalist 102: 391-403. Murphy, T.D. 1963. Amphibian populations and movements at a small semipermanent pond in Orange County, North Carolina. Ph.D. Dissertation, Duke University, Durham, North Carolina. Nickerson, M.A. and Mays, C.E. 1973. The Hellbenders: North American “Giant Salamanders.” Milwaukee Public Museum, Publications in Biology and Geology No.1. 106 pp. Noble, G.K. 1926. The Long Island newt: a contribution to the life history of Triturus viridescens. American Museum Novitates No. 228: 1-11. Noble, G.K. 1929. Further observations on the life-history of the newt, Triturus viridescens. American Museum Novitates No. 348: 1-22. Noble, G. K. 1931. The Biology of the Amphibia. McGraw-Hill Book Company, Inc., New York, NY. 577 pp. Nussbaum, R.A. 1985. The evolution of parental care in salamanders. Miscellaneous Publications, Museum of Zoology, University of Michigan No. 169: 1-50. Life Histories 521 Nussbaum, R.A. 1987. Parental care and egg size in salamanders: an examination of the safe harbor hypothesis. Researches on Population Ecology 29: 27-44. Nussbaum, R.A. and Clothier, G.W. 1973. Population structure, growth, and size of larval Dicamptodon ensatus (Eschscholtz). Northwest Science 47: 218-227. Nussbaum, R.A. and Tait, C.K. 1977. Aspects of the life history and ecology of the Olympic salamander, Rhyacotriton olympicus (Gaige). American Midland Naturalist 98: 176-199. O’Laughlin, B.E. and Harris, R.N. 2000. Models of metamorphic timing: an experimental evaluation with the pond-dwelling salamander Hemidactylium scutatum (Caudata: Plethodontidae). Oecologia 124: 343-350. Olgun, K., Miaud, C. and Gautier, P. 2001. Age, growth, and survivorship in the viviparous salamander Mertensiella luschani from southwestern Turkey. Canadian Journal of Zoology 79: 1559-1567. Oliver, J.A. 1955. The Natural History of North American Amphibians and Reptiles. D. Van Nostrand Company, Inc., Princeton, New Jersey. 359 pp. Organ, J.A. 1961. Studies of the local distribution, life history, and population dynamics of the salamander genus Desmognathus in Virginia. Ecological Monographs 31: 189-220. Orton, G.L. 1953. The systematics of vertebrate larvae. Systematic Zoology 2: 63-75. Özeti, N. 1979. Reproductive biology of the salamander Mertensiella luschani antalyana. Herpetologica 35: 193-197. Patterson, K.K. 1978. Life history aspects of paedogenic populations of the mole salamander, Ambystoma talpoideum. Copeia 1978: 649-655. Peterson, C.L., Metter, D.E., Miller, B.T., Wilkinson, R.F. and Topping, M.S. 1988. Demography of the hellbender Cryptobranchus alleganiensis in the Ozarks. American Midland Naturalist 119: 291-303. Peterson, C.L., Wilkinson, R.F., Jr., Topping, M.S. and Metter, D.E. 1983. Age and growth of the Ozark hellbender (Cryptobranchus alleganiensis bishopi). Copeia 1983: 225-231. Petranka, J.W. 1984a. Incubation, larval growth, and embryonic and larval survivorship of smallmouth salamanders (Ambystoma texanum) in streams. Copeia 1984: 862-868. Petranka, J.W. 1984b. Sources of interpopulational variation in growth responses of larval salamanders. Ecology 65: 1857-1865. Petranka, J.W. 1998. Salamanders of the United States and Canada. Smithsonian Institution Press, Washington, D.C. 587 pp. Polymeni, R.M. 1994. On the biology of Mertensiella luschani (Steindachner, 1891): a review. Pp. 301-314. In H. Greven and B. Thiesmeier (eds), Biology of Salamandra and Mertensiella. Mertensiella, Supplement 4, DGHT, Bonn, Germany. Reilly, S.M., Wiley, E.O. and Meinhardt, D.J. 1997. An integrative approach to heterochrony: the distinction between interspecific and intraspecific phenomena. Biological Journal of the Linnean Society 60: 119-143. Roff, D.A. 1996. The evolution of threshold traits in animals. Quarterly Review of Biology 71: 3-35. Roff, D.A. 2002. Life History Evolution. Sinauer Associates, Inc. Sunderland, Massachusetts. 527 pp. Rose, C.S. 1995. Intraspecific variation in ceratobranchial number in Hemidactylium scutatum (Amphibia: Plethodontidae): developmental and systematic implications. Copeia 1995: 228-232. 522 Reproductive Biology and Phylogeny of Urodela Rose, F.L. and Armentrout, D. 1976. Adaptive strategies of Ambystoma tigrinum Green inhabiting the Llano Estacado of west Texas. Journal of Animal Ecology 45: 713-729. Rowe, L. and Ludwig, D. 1991. Size and timing of metamorphosis in complex life cycles: time constraints and variation. Ecology 72: 413-427. Ryan, T.J. and Bruce, R.C. 2000. Life history evolution and adaptive radiation of hemidactyliine salamanders. Pp. 303-326. In R.C. Bruce, R.G. Jaeger and L.D. Houck (eds), The Biology of Plethodontid Salamanders. Kluwer Academic/Plenum Publishers, New York, NY. Ryan, T.J. and Semlitsch, R.D. 1998. Intraspecific heterochrony and life history evolution: decoupling somatic and sexual development in a facultatively paedomorphic salamander. Proceedings of the National Academy of Sciences, USA 95: 5643-5648. Salthe, S.N. 1969. Reproductive modes and the number and sizes of ova in the urodeles. American Midland Naturalist 81: 467-490. Salthe, S.N. and Mecham, J.S. 1974. Reproductive and courtship patterns. Pp. 309-521. In B. Lofts (ed), Physiology of the Amphibia, Volume II. Academic Press, New York, NY. Salvidio, S. 1993. Life history of the European plethodontid salamander Speleomantes ambrosii (Amphibia, Caudata). Herpetological Journal 3: 55-59. Schoch, R.R. 1992. Comparative ontogeny of early Permian branchiosaurid amphibians from southwestern Germany. Palaeontographica 222: 43-83. Semlitsch, R.D. 1983. Growth and metamorphosis of larval red salamanders (Pseudotriton ruber) on the Coastal Plain of South Carolina. Herpetologica 39: 48-52. Semlitsch, R.D. 1985. Reproductive strategy of a facultatively paedomorphic salamander Ambystoma talpoideum. Oecologia 65: 305-313. Semlitsch, R.D. 1987a. Relationship of pond drying to the reproductive success of the salamander Ambystoma talpoideum. Copeia 1987: 61-69. Semlitsch, R.D. 1987b. Paedomorphosis in Ambystoma talpoideum: effects of density, food, and pond drying. Ecology 68: 994-1002. Semlitsch, R.D. and Gibbons, J.W. 1985. Phenotypic variation in metamorphosis and paedomorphosis in the salamander Ambystoma talpoideum. Ecology 66: 1123-1130. Semlitsch, R.D., Harris, R.N. and Wilbur, H.M. 1990. Paedomorphosis in Ambystoma talpoideum: maintenance of population variation and alternative life-history pathways. Evolution 44: 1604-1613. Semlitsch, R.D., Scott, D.E. and Pechmann, J.H.K. 1988. Time and size at metamorphosis related to adult fitness in Ambystoma talpoideum. Ecology 69: 184-192. Semlitsch, R.D. and Wilbur, H.M. 1989. Artificial selection for paedomorphosis in the salamander Ambystoma talpoideum. Evolution 43: 105-112. Sessions, S.K. and Larson, A. 1987. Developmental correlates of genome size in plethodontid salamanders and their implications for genome evolution. Evolution 41: 1239-1251. Sever, D.M. 1994. Comparative anatomy and phylogeny of the cloacae of salamanders (Amphibia: Caudata). VII. Plethodontidae. Herpetological Monographs 8: 276-337. Sever, D.M. 2000. Sperm storage in female plethodontids with especial reference to the Desmognathinae. Pp. 345-369. In R. C. Bruce, R. G. Jaeger and L. D. Houck (eds), The Biology of Plethodontid Salamanders. Kluwer Academic/Plenum Publishers, New York, NY. Life Histories 523 Sever, D.M. 2002. Female sperm storage in amphibians. Journal of Experimental Zoology 292: 165-179. Sever, D.M., Krenz, J.D., Johnson, K.M. and Rania, L.C. 1995. Morphology and evolutionary implications of the annual cycle of secretion and sperm storage in spermathecae of the salamander Ambystoma opacum (Amphibia: Ambystomatidae). Journal of Morphology 223: 35-46. Sever, D.M., Rania, L.C. and Krenz, J.D. 1996a. Reproduction of the salamander Siren intermedia Le Conte with especial reference to oviducal anatomy and mode of fertilization. Journal of Morphology 227: 335-348. Sever, D.M., Rania, L.C. and Krenz, J.D. 1996b. Annual cycle of sperm storage in spermathecae of the red-spotted newt, Notophthalmus viridescens (Amphibia: Salamandridae). Journal of Morphology 227: 155-170. Sexton, O.J. and Bizer, J.R. 1978. Life history patterns of Ambystoma tigrinum in montane Colorado. American Midland Naturalist 99: 101-118. Shi, Y-B. 2000. Amphibian Metamorphosis: from Morphology to Molecular Biology. John Wiley & Sons, Inc., New York, NY. 288 pp. Shine, R. 1979. Sexual selection and sexual dimorphism in the Amphibia. Copeia 1979: 297-306. Shoemaker, V.H., Hillman, S.S., Hillyard, S.D., Jackson, D.C., McClanahan, L.L., Withers, P.C. and Wygoda, M.L. 1992. Exchange of water, ions, and respiratory gases in terrestrial amphibians. Pp. 125-150. In M. E. Feder and W.W. Burggren (eds), Environmental Physiology of the Amphibians. University of Chicago Press, Chicago, Illinois. Shoop, C.R. 1974. Yearly variation in larval survival of Ambystoma maculatum. Ecology 55: 440-444. Smith, B.G. 1907. The life history and habits of Cryptobranchus allegheniensis. Biological Bulletin 13: 5-39. Smith-Gill, S.J. and Berven, K.A. 1979. Predicting amphibian metamorphosis. American Naturalist 113: 563-585. Sprules, W.G. 1974a. Environmental factors and the incidence of neoteny in Ambystoma gracile (Baird) (Amphibia: Caudata). Canadian Journal of Zoology 52: 1545-1552. Sprules, W.G. 1974b. The adaptive significance of paedogenesis in North American species of Ambystoma (Amphibia: Caudata): an hypothesis. Canadian Journal of Zoology 52: 393-400. Stearns, S.C. 1992. The Evolution of Life Histories. Oxford University Press, Oxford, U.K. 249 pp. Stenhouse, S.L. 1985. Interdemic variation in predation on salamander larvae. Ecology 66: 1706-1717. Stenhouse, S.L., Hairston, N.G. and Cobey, A.E. 1983. Predation and competition in Ambystoma larvae: field and laboratory experiments. Journal of Herpetology 17: 210-220. Sweet, S.S. 1977. Natural metamorphosis in Eurycea neotenes, and the generic allocation of the Texas Eurycea (Amphibia: Plethodontidae). Herpetologica 33: 364-375. Taber, C.A., Wilkinson, R.F., Jr. and Topping, M.S. 1975. Age and growth of hellbenders in the Niangua River, Missouri. Copeia 1975: 633-639. Tarkhnishvili, D.N. 1994. Interdependences between populational, developmental and morphological features of the Caucasian salamander, Mertensiella caucasica. 524 Reproductive Biology and Phylogeny of Urodela Pp. 315-325. In H. Greven and B. Thiesmeier (eds), Biology of Salamandra and Mertensiella. Mertensiella Supplement 4, DGHT, Bonn, Germany. Tarkhnishvili, D.N. and Gokhelashvili, R.K. 1994. Preliminary data of the age structure of a Mertensiella caucasica population. Pp.327-334. In H. Greven and B. Thiesmeier (eds), Biology of Salamandra and Mertensiella. Mertensiella Supplement 4, DGHT, Bonn, Germany. Thiesmeier, B. 1990. Untersuchungen zur phänologie und populationsdynamik des feuersalamanders (Salamandra salamandra terrestris Lacépède, 1788) im Niederbergischen Land (BRD). Zoologische Jahrbuecher Systematik 117: 331-353. Thiesmeier, B. 1994. Aspects of larval ecology of fire salamanders (Salamandra salamandra) in middle Europe. Pp. 335-345. In H. Greven and B. Thiesmeier (eds), Biology of Salamandra and Mertensiella. Mertensiella Supplement 4, DGHT, Bonn, Germany. Thiesmeier, B. and Haker, K. 1990. Salamandra salamandra bernardezi Wolterstorff, 1928 aus Oviedo, Spanien, nebst bemerkungen zur viviparie in der gattung Salamandra. Salamandra 26: 140-154. Thiesmeier, B. and Kupfer, A. 2000. Der Kammmolch: ein Wasserdrache in Gefahr. Laurenti-Verlag, Bochum, Germany. 158 pp. Thiesmeier, B., Sauer, W. and Mutz, T. 1994. Birth and dates of offspring of Salamandra salamandra bernardezi from Oviedo. Pp. 347-354. In H. Greven and B. Thiesmeier (eds), Biology of Salamandra and Mertensiella. Mertensiella Supplement 4, DGHT, Bonn, Germany. Thiesmeier, B. and Schuhmacher, H. 1990. Causes of larval drift of the fire salamander, Salamandra salamandra terrestris, and its effects on population dynamics. Oecologia 82: 259-263. Tilley, S.G. 1968. Size-fecundity relationships and their evolutionary implications in five desmognathine salamanders. Evolution 22: 806-816. Tilley, S.G. 1973. Life histories and natural selection in populations of the salamander Desmognathus ochrophaeus. Ecology 54: 3-17. Tilley, S.G. 1974. Structures and dynamics of populations of the salamander Desmognathus ochrophaeus Cope in different habitats. Ecology 55: 808-817. Tilley, S.G. 1980. Life histories and comparative demography of two salamander populations. Copeia 1980:806-821. Tilley, S.G. and Bernardo, J. 1993. Life history evolution in plethodontid salamanders. Herpetologica 49: 154-163. Ultsch, G.R. and Arceneaux, S.J. 1988. Gill loss in larval Amphiuma tridactylum. Journal of Herpetology 22: 347-348. Valentine, B.D. and Dennis, D.M. 1964. A comparison of the gill-arch system and fins of three genera of larval salamanders, Rhyacotriton, Gyrinophilus, and Ambystoma. Copeia 1964: 196-201. Vandel, A. and Bouillon, M. 1959. La reproduction du protée (Proteus anguinus Laurenti). Comptes Rendus de l’Académie des Sciences, Paris 248: 1267-1272. Vandel, A., Durand, J. and Bouillon, M. 1964. Observations sur le développement du protée, Proteus anguinus Laurenti (Batraciens, Urodèles). Comptes Rendus de l’Académie des Sciences, Paris 259: 4801-4804. Via, S. 1993. Adaptive phenotypic plasticity: target or by-product of selection in a variable environment? American Naturalist 142: 352-365. Vial, J.L. 1968. The ecology of the tropical salamander, Bolitoglossa subpalmata, in Costa Rica. Revista de Biologia Tropical 15: 13-115. Life Histories 525 Vilter, V. 1986. La reproduction de la salamandre noire (Salamandra atra). Pp. 487-495. In P. P. Grasse and M. Delsol (eds), Traité de Zoologie, Amphibiens, Tome 14. Masson, Paris, France. Voss, S.R. 1993. Relationship between stream order and length of larval period in the salamander Eurycea wilderae. Copeia 1993: 736-742. Voss, S.R. 1995. Genetic basis of paedomorphosis in the axolotl, Ambystoma mexicanum: a test of the single-gene hypothesis. Journal of Heredity 86: 441-447. Wake, D.B. 1966. Comparative osteology and evolution of the lungless salamanders, family Plethodontidae. Memoirs of the Southern California Academy of Sciences 4: 1-111. Wake, D.B. 1991. Homoplasy: the result of natural selection, or evidence of design limitations? American Naturalist 138: 543-567. Wake, D.B. and Castanet, J. 1995. A skeletochronological study of growth and age in relation to adult size in Batrachoseps attenuatus. Journal of Herpetology 29: 60-65. Wake, D.B. and Hanken, J. 1996. Direct development in the lungless salamanders: what are the consequences for developmental biology, evolution and phylogenesis? International Journal of Developmental Biology 40: 859-869. Wake, M.H. 1993. Evolution of oviductal gestation in amphibians. Journal of Experimental Zoology 266: 394-413. Wakely, J.F., Fuhrman, G.J., Fuhrman, F.A., Fischer, H.G. and Mosher, H.S. 1966. The occurrence of tetrodotoxin (tarichatoxin) in Amphibia and the distribution of the toxin in the organs of newts (Taricha). Toxicon 3: 195-203. Wassersug, R.J. 1974. Evolution of anuran life cycles. Science 185: 377-378. Werner, E.E. 1986. Amphibian metamorphosis: growth rate, predation risk, and the optimal size at transformation. American Naturalist 128: 319-341. Whipple, I.L. 1906. The ypsiloid apparatus of urodeles. Biological Bulletin 10: 255-297. Whiteman, H.H. 1994. Evolution of facultative paedomorphosis in salamanders. Quarterly Review of Biology 69: 205-221. Whittier, J.M. and Crews, D. 1987. Seasonal reproduction: patterns and controls. Pp. 385-409. In D. O. Norris and R. E. Jones (eds), Hormones and Reproduction in Fishes, Amphibians, and Reptiles. Plenum Press, New York, NY. Wilbur, H.M. 1972. Competition, predation, and the structure of the Ambystoma-Rana sylvatica community. Ecology 53: 3-21. Wilbur, H.M. 1980. Complex life cycles. Annual Review of Ecology and Systematics 11: 67-93. Wilbur, H.M. and Collins, J.P. 1973. Ecological aspects of amphibian metamorphosis. Science 182: 1305-1314. Wilder, I.W. 1913. The life history of Desmognathus fusca. Biological Bulletin 24: 251-342. Wilder, I.W. 1924. The relation of growth to metamorphosis in Eurycea bislineata (Green). Journal of Experimental Zoology 40: 1-112. Wilder, I.W. 1925. The Morphology of Amphibian Metamorphosis. Smith College, Northampton, Massachusetts. 161 pp. Zalisko, E.J., Brandon, R.A. and Martan, J. 1984. Microstructure and histochemistry of salamander spermatophores (Ambystomatidae, Salamandridae and Plethodontidae). Copeia 1984: 739-747. Zardoya, R. and Meyer, A. 2001. On the origin of and phylogenetic relationships among living amphibians. Proceedings of the National Academy of Sciences, USA 98: 7380-7383.