Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
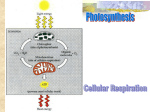
UNIT 3 NOTES CHAPTER 8 SECTIONS 1-5 All living systems require constant input of free energy, mainly to keep them ordered and organized. There is order to metabolic pathways, organ systems, and organisms. Metabolism is Energy is the capacity to cause change which often results in work being done. Some of the types of work done by cells that require energy: Building macromolecules Cell division Pumping ions and other materials across cell membranes Moving: flagella, amoeboid movement, muscles, etc. Making light- bioluminescence Heating the cell Some types of energy are: Kinetic energy is energy associated with motion Heat (thermal energy) is kinetic energy associated with random movement of atoms or molecules Potential energy is energy that matter possesses because of its location or structure Chemical energy is potential energy available for release in a chemical reaction Thermodynamics is the study of energy transformations, where energy is changed from one form to another. You have probably heard of the first law of thermodynamics. It is the conservation of energy- energy can be neither created nor destroyed, just changed to a different form. So that means that chemical energy can be changed into heat or kinetic energy, light energy can be changed to heat or chemical energy, etc. The second law of thermodynamics is a bit more complicated. It states that every energy transfer or transformation increases the entropy (disorder) of the universe. Cells and organisms transform energy all the time in a multitude of reactions to do work to keep them organized. How does that not violate the second law? Even though living systems need to maintain order which decreases entropy, cellular processes such as respiration break down larger molecules into smaller ones that have less energy. This lower energy level is accounted for by heat generated and given off to the environment (universe) which increases entropy. There may be a decrease in entropy in an organism, but there is an overall increase in entropy in the system as a whole. 1 UNIT 3 NOTES So cells and organisms live and grow everyday taking in energy (food or light), breaking large molecules apart, and reorganizing small molecules into larger ones it needs, always giving off heat. Metabolic pathways can be separated into two categories: breaking and building. Pathways that release energy by breaking large molecules apart (breaking bonds) are called __________________________. Respiration is an example. Pathways that use energy to make bonds and build molecules are called _____________________. Protein synthesis is an example. Since an organism never gets to use all the energy it acquires, there must be a constant supply or it will die. The energy coming into a system, or organism, must be greater than the heat lost to the environment in order for the organism to have enough energy left over for metabolic processes. Now let’s talk about free energy, G. This doesn’t mean there’s no cost; it means the energy is available to do work. Cells need that. But where does it come from? Let’s say it’s in that Snickers bar you just ate. (Yeh, get me one too!) Eventually the sugar in there will get to your cells and the sugar has energy stored in its chemical bonds. That stored energy is free energy. When the sugar is broken down the energy in those bonds holding the atoms together will be released and become free to do some work. Some energy will also be released as heat, remember. Some kinds of work the cell does were mentioned above. Now that the cell has some energy it can, oh I don’t know, build a protein. Building things, or creating order (decreasing entropy), requires energy while breaking things (increasing entropy) releases it (generally). So for the most part, catabolic reactions release energy and anabolic reactions require energy. These are actual things that cells do. As you can see some things need free energy to get done and others don’t. Those things that don’t need free energy added, like the breakdown of sugar, happen spontaneously. Somewhere some scientists with A LOT of time on their hands figured out that these energy changes could be measured and they could tell what reactions would be spontaneous and which would not according to this formula: Free energy change = change in enthalpy – (temperature K)(change in entropy) ∆G = ∆H - T ∆S If ∆G is negative then the reaction will occur spontaneously because it does not require an input of free energy, but actually releases energy and is termed an exergonic reaction. An endergonic reaction has a positive ∆G, requires the input of free energy, and is not spontaneous. 2 UNIT 3 NOTES If you recall, entropy, S, is a measure of disorder and the universe prefers disorder, randomness. Reactions that favor disorder will have a positive change in S or, ∆S will be greater than zero. A high ∆S will usually result in a negative ∆G. Order is maintained by coupling cellular processes that increase entropy (and so have negative changes in free energy) with those that decrease entropy (and so have positive changes in free energy). Since cells use both kinds of reactions, exergonic reactions that release energy, such as ATPADP, can be coupled with endergonic reactions that need energy so that the energy released from the former is used for the latter. If slightly more energy is released than is used the overall ∆G will be negative and therefore, the whole thing will happen spontaneously. POOF!! Just like magic. When reactions are coupled and the overall ∆G is negative then the system as a whole is exergonic and will occur spontaneously. This is how much work gets done in a cell. This does not mean that these reactions have to occur at the same time or place. When ATP is produced by one reaction it can ‘hold’ its energy until needed and can move about the cell to where it is needed. What is ATP? Write its complete name, its job, and draw a basic outline (Fig. 8.8): When is the energy released from ATP? And then ATP becomes what? ATP drives endergonic reactions by phosphorylation. What is phosphorylation? Refer to Figure 8.10 pg.150, slide 30, for an example of energy coupling using ATP. One reason ATP is used as ‘the energy molecule’ is because it is renewable. Yes, all cells are green, energy responsible, and recycle everyday. 3 UNIT 3 NOTES ADP is phosphorylated using energy from catabolic reactions like cellular respiration. So ATP ADP ATP happens over and over and over and … That’s the story of “How the Cell Gets Its Energy”. Read it every night for a week before you go to bed to help you sleep. Pretend it’s one of Grimm’s fairy tales. When you get tired of that story (no pun intended) here’s another one: the story of “The Enzyme and the Substrate”. Once upon a time there was a catalyst. What is a catalyst? Biological catalysts are called enzymes. Enzymes help reactions that would normally happen anyway to happen faster, that is, more often in the same amount of time. Almost all enzymes are proteins and most of their names end in –ase. Quick- what’s the most important thing you learned about proteins in the last unit? That’s right, they work by shape. Shape determines function and for a biological catalyst shape is the most important thing. An enzyme provides a space with the right conditions and just the right fit for the reactants of a reaction. Everything is already perfect when the reactants get there so the reaction can happen speedyquick, the product is released and more reactants can come and react, product released, more come, etc. It’s like a Las Vegas wedding chapel compared to Westminster Abbey. Las Vegas is faster and waaaay cheaper. Speaking of cheaper, for reactions to happen at all there is a cost. It is called the activation energy. All chemical reactions require activation energy to get them started, even those that occur spontaneously. Look at slide 36, Fig. 8-14 pg. 152. Notice that it shows that ∆G for that reaction is <0 so it is spontaneous, but it still requires activation energy. That’s the Westminster Abbey cost. How can we get that lowered to the Las Vegas cost? If you said “With an enzyme!” you’re right! So, how does an enzyme work? Make sure to check out slide 38, Fig. 8-15 pg. 153. As you can see, the activation energy is lower when there is an enzyme to help the reaction to occur. And that’s the way you need to think about it: enzymes can only help reactions to happen faster but otherwise do not participate in the reaction- they don’t go on the honeymoon. Now that you know all about Enzyme, let’s learn about Substrate. It was said that an enzyme is a protein (95% of the time) and the most important thing about 4 UNIT 3 NOTES proteins is their shape which provides a perfect space for a reaction. Since every protein has its own shape each enzyme is shaped differently, therefore, it can only help one specific reaction. This is why you have a ton of enzymes. Remember that Snickers bar you ate? To digest it you need a different enzyme for each ingredient because each type of molecule in the Snickers is shaped differently. That means enzymes are very specific and can act on only one substrate. For our wedding analogy that means that only one couple can get married per chapel, but it is still cheaper than Westminster Abbey, and they can get married over and over and over and over…… really fast. So a substrate is the reactant an enzyme acts on. When the two get together what is it called? What do we call the place the substrate binds to? Sometimes when the enzyme binds to the substrate the enzyme’s shape will change just slightly making a more snuggly fit with the substrate so it works better. This is called induced fit- the final shape is produced only when the substrate binds to the enzyme. You can see this in slide 40, Fig. 8.17 pg. 154. Also notice in the figure that there are two reactants for the substrate. Chemical reactions involve breaking things apart, putting things together, or rearranging things. Most chemical reactions are also reversible and so enzymes go both ways; they can break one substrate molecule into two or put those two back together depending on the needs of the cell. (The wedding chapel could also grant a divorce, which could be possible in Las Vegas.) Therefore, an enzyme may actually have two substrates but both will be needed for the reaction to take place. We learned a lot so let’s review: Every protein has a specific shape A enzyme contains an active site that is complementary in shape to the substrate An enzyme is a catalyst which lowers the activation energy of the reaction Enzymes are substrate specific Enzymes are necessary for life. Otherwise the reactions wouldn’t happen fast enough. Enzymes are so specific that even enantiomers are not acted upon. Remember, these are like mirror images or left hand-right hand shapes. If the thumb is on the wrong side the glove doesn’t fit. It’s the same with an enzyme. It only works when the substrate fits. Now you have another bedtime story so you can alternate for a while so you don’t get bored. 5 UNIT 3 NOTES Cofactors and coenzymes affect enzyme function; this interaction relates to a structural change that alters the activity rate of the enzyme. The enzyme may only become active when all the appropriate cofactors or coenzymes are present and bind to the appropriate sites on the enzyme. Some enzymes need these extra parts to get the correct final shape to work. Many of these are vitamins and minerals required in your diet. You need to eat those so your enzymes will have all the parts needed for proper function. So eat your broccoli. All this is well and good, but as in every fairy tale there is a villain. Yup, there is always a wicked queen or stepmother. And why is it always a woman? Anyway, for enzymes to work properly they must have the right conditions. They’re picky. What are some conditions that affect enzymes? As you can see from Fig. 8.18 pg. 155, slide 44, the two enzymes in each graph work best in very different temperatures or pH. Outside those ranges the enzymes cease to function. This also goes hand in hand with the idea of compartmentalization. Cells have membrane bound areas (organelles) that can have different conditions and organisms like yourselves have organs with different conditions. Your stomach has a pH of around 2, but your small intestine has a pH of about 8. Chemical concentrations can also be different in organs and organelles. Temperature also has an effect on enzymes. Looking at those graphs you can see that each enzyme has an optimum that it likes best. For human enzymes, that is going to be around 37o C because that is body temperature. Below that enzymes get slow, but when you start to go above that bad things happen (cue the music and the wicked queen!). At high temperatures proteins, and hence enzymes, start to fall apart and lose their shape. No shape, no function. This is called denaturization. Once a protein is denatured it is just a pile of amino acids, and like Humpty Dumpty it’s not getting put back together. Thought question: if high temperatures will denature enzymes is a fever good or bad? Think hard before answering and cue the Jeopardy theme. Those are environmental conditions that affect enzymes. There are also some chemicals that affect enzymes called inhibitors (some are very, very, evil, wicked, bad stepmothers). What does inhibit mean? If you don’t know, get a dictionary. That’s the big book on the shelf in the corner covered in dust that has just a bunch of words and their meanings in it. That’s all we had before Google. (sigh) 6 UNIT 3 NOTES Slide 45 tells you there are two kinds of inhibitors: Competitive inhibitors that bind where? Noncompetitive inhibitors that work how? Notice the evil stepmothers, I mean, examples of inhibitors: toxins, poisons, pesticides, and antibiotics. What’s the common theme? Yes, they all tend to sicken or cause death. While most inhibitors are reversible these are not. Once they bind they stay and the enzyme is rendered useless. If all of the enzyme an organism has is out of commission then that reaction can no longer be performed and the organism will suffer and/or die. Literally a poison apple. Chemical chaos would result if a cell’s metabolic pathways were not tightly regulated. We can’t have cells running amok just doing all kinds of reactions willynilly. So there are two ways to regulate enzymes. One is by switching on or off the gene that has the code to make the enzyme. This will be discussed in Unit 6. For now suffice it to say that when genes are “on” the proteins they code for can be made, so the cell would be making many copies of that enzyme. Turn the gene “off” and production stops- no more enzyme, no more reactions. The reaction would eventually stop if the gene was not turned back on because enzymes, like other molecules in the cell, don’t last forever but eventually degrade. OK, that’s one way to control enzymes. What’s the other, you ask? It’s called allosteric regulation – a protein’s function at one site is affected by the binding of a regulatory molecule to a separate site. The protein can be either activated or inhibited. What does the binding of an activator do? What does the binding of an inhibitor do? Cooperativity is a special type of allosteric regulation. Cooperativity occurs in multisubunit enzymes when the binding of a substrate molecule to one of the subunits stabilizes the whole unit and allows other substrate molecules to bind more easily. This is seen in hemoglobin which is not an enzyme, but it is a protein. Because some enzymes can be regulated allosterically, if scientists can determine what enzyme controls a pathway and what molecule can inhibit that enzyme, then there is a chance a drug can be made to control the pathway. Powerful stuff. 7 UNIT 3 NOTES Some enzymatic reactions are regulated by their own products and/or substrates. Feedback inhibition occurs when the product of a metabolic pathway acts as an inhibitor by binding to one of the enzymes in the pathway. This has the effect of slowing or stopping the production of product when enough has been produced. This is a common method of metabolic control. As product is used production would start up again. See Figure 8.22 pg. 159, slide 56, for the isoleucine pathway as an example. Some metabolic pathways are regulated by the amount of substrate. Production of product may begin when there is a build up of substrate to activate the enzymes. The production of ATP is regulated this way. CHAPTER 9 SECTIONS 1-6 **REMINDER: pH is a measure of H+ concentration. A lower pH is more acidic- it has more H+ ions. Remember back on pg. 2 where we had the story about energy? And we said that cells get their energy from a Snickers bar? Or more accurately the chemical bonds in the sugar molecules in the Snickers bar? In this story chapter you will learn, in great, gory detail exactly how that happens. Cells can actually get energy to use from fats and proteins as well as sugar, but sugar, namely glucose, is always used as the example so we will stick with that. The chemical energy that is in food can be stored or converted to ATP for use. Before we begin (because this is not in the PowerPoint) let’s talk about calories, those things in food you don’t want too many of. A calorie in food is actually a measure of the energy it contains. If you were to take a gram of glucose and burn it completely it would release 4000 calories or 4 kcal or 4C. The C means Calories and that is how energy is measured in food. They say calories but they mean Calories (kilocalories). Proteins also have about 4kcal per gram, but fats have 9kcal per gram. This is why fat is used as storage. You get more kcal per gram. This is also why it is hard to get rid of once you store it. It takes more than twice as much work to burn off a gram of fat than sugar. Check it out on a food label. If it says the food has 3 grams of fat then the total Calories from fat in that food should be 3gm X 9kcal/gm = 27kcal or 27 C. So, what is our cell going to do with the glucose molecule? Burn it? Yes, sort of. But not all at once. That would release all the energy at one time and destroy the cell like a nuclear bomb. Not helpful. Instead the cell is going to break it down slowly, step by step. There are several ways a cell can do this and we will discuss two of them: fermentation and aerobic cellular respiration. 8 UNIT 3 NOTES Fermentation partially breaks down glucose and stores some energy in ATP, but not much. We’ll talk about that later. Aerobic cellular respiration is commonly called cellular respiration and is usually what is meant by just respiration, so that’s what we’ll call it. Respiration makes waaay more ATPs than fermentation. What does respiration use that fermentation does not? In what organelle does respiration take place? Summarize the overall process of cellular respiration: Note: This is a catabolic reaction so it is exergonic and will happen spontaneously. Always try to relate new learning to old learned stuff. Everything is connected. Let’s take a quick trip back to chemistry and review REDOX reactions. I didn’t like them either so that’s why we need to review just a little because where the electrons go is very important when studying respiration. I believe it goes like this: LEO – Lose Electrons Oxidation, becomes more positive says GER – Gain Electrons Reduction, becomes more negative Chemical bonding involves electrons so breaking bonds involves moving electrons around. This is really where the energy is. During respiration, the electrons in the bonds of the glucose are transferred from one atom to another as it is broken down. The glucose Loses Electrons and is therefore, Oxidized. Energy is released and ATP is made to store the energy. The electrons are moved around a lot until they reach their final acceptor, our good friend, oxygen. Since oxygen is Gaining Electrons it gets Reduced. Remember: oxygen is the final electron acceptor. ***There are A LOT of steps to respiration. Fortunately, you DO NOT need to learn each tiny little step or the names of most molecules. Your book has all the really gory details if you must know, but if I were you I would just stick to what is talked about here, in class, and the PowerPoint. *** Before the electrons get to oxygen they are passed around like a hot potato. During the breakdown of glucose (C6H12O6) the six carbons that are all attached must be separated to form carbon dioxide (CO2) and the hydrogens are removed. As these bonds are all broken electrons are released and passed to electron carriers, or acceptors, that will take them to something called the electron transport chain. The 9 UNIT 3 NOTES main electron acceptor is NAD+. When it accepts an electron it becomes NADH. In your previous class you probably learned of the electron transport chain as something mysterious that looked like a staircase. It’s usually drawn that way to emphasize that it is a process comprised of many little steps, where at each step a tiny bit of energy is released rather than one big burst. See slide 70, Fig. 9.5 pg. 165. What is at the bottom? OK, get ready, set, GO! This is cellular respiration! (imagine bells, whistles, fireworks) Overall, there are three stages you need to know. List them here: 1. 2. 3. Slides 72-74, Fig. 9.6. pg. 166, shows the order of these stages, where they take place in the cell, and whether it is substrate-level or oxidative phosphorylation. (I’ll explain that later.) This is one of the most important figures in the whole book so learn it. Notice each stage produces ATP. Where does most of the ATP come from? Why? What percentage of all the ATP is made here? Slide 77, Fig. 9.7 pg. 167, shows what substrate-level phosphorylation is. Some substrate (could be any number of things) that has a phosphate group is acted upon by an enzyme to remove the phosphate group and attach it to ADP making ATP. Yup, that’s it. Simple. This happens a few times during the first two stages of cellular respiration just as a by-product of the actual reaction going on as glucose is changed to carbon dioxide. The first stage of respiration is glycolysis. Glyco- means a carbohydrate, in this case glucose. Lysis means to split or break apart. Put them together and you have the splitting of glucose. What does the glucose get split into? 10 UNIT 3 NOTES Where goes this reaction take place? Slide 79, Fig. 9.8 pg. 167, is a summary of glycolysis. That’s all you need to learn. Unless you are planning to get a doctorate degree in biochemistry you DO NOT need to learn ANY of Fig. 9.9 on pgs. 168-9. Those are gory details we don’t need. What you DO need to know is the NET summary: Glucose is oxidized through a series of steps to 2 pyruvates Takes place in cytosol 2 ATPs go in and 4 ATPs are made for a gain of 2 ATPs (substrate level phosphorylation) 2 NAD+ go in and accept electrons to make 2NADH and 2H+ (these H+s dissolve into the cytoplasm) Glycolysis has a net yield of 2 ATPs and 2 NADHs per glucose molecule. We now have 2 pyruvate molecules that need to be further oxidized to get all the energy out. If there is oxygen available to the cell those pyruvates will be transported to the mitochondria where oxidation will continue. If oxygen is not available then the pyruvate will suffer a different fate depending on what kind of cell it is in. We’ll discuss this later. At the mitochondrion pyruvate goes through a transitional step before entering the Kreb’s cycle or what is now called the citric acid cycle. (I don’t know what Kreb did to make scientists drop his name, but it couldn’t have been good. Probably blew up somebody’s lab.) Each pyruvate is converted to acetyl-CoA. This occurs in 3 steps (slide 81, Fig. 9.10 pg. 170) which yield one CO2 and one NADH. The CO2 is removed as waste, the NADH will go to the ETC (electron transport chain) and the acetyl-CoA can now enter the citric acid cycle. Keep in mind that each glucose produces two pyruvates, therefore, this transition reaction and the citric acid cycle will occur twice for each molecule of glucose. Where does the citric acid cycle take place? The cycle oxidizes organic fuel derived from pyruvate, generating 1 ATP, 3 NADH, and 1 FADH2 per turn. Since there are two turns per glucose, what do you get per glucose? 11 UNIT 3 NOTES Here you are introduced to another electron acceptor, FAD. It does the same job as NAD+ and takes electrons to the ETC, but not quite as efficiently as NAD+. During oxidative phosphorylation, chemiosmosis couples electron transport to ATP synthesis. This is the title to section 9.4 and I know what you’re thinking: What the h*** does that mean? By using all these big scientific terms the writers were able to say in one sentence how the cell makes most of its ATP. We will now take this one step at a time and break it down so you too, can tell your friends, “During oxidative phosphorylation, chemiosmosis couples electron transport to ATP synthesis.” and actually know what you’re talking about. Your friends will think you’re a nerd, but that’s OK. In the immortal words of Mr. Palczak, “It’s OK to be a nerd because if you’re not, someday you will work for one.” Alright, boss, let’s get started. The 2 ATPs generated by two turns of the citric acid cycle will be used by the cell to do work. The 2 FADH2 and the 6 NADH will transport their electrons to the ETC. The energy in those electrons will be used to make most of the ATP the cell needs, but not directly by the ETC. The ATP is made by a process called oxidative phosphorylation which uses chemiosmosis. Where is the ETC located? The ETC is just a bunch of proteins that are imbedded next to each other in a membrane. This is not the only ETC cells have. There is also one in chloroplasts that is used during photosynthesis. When somebody mentions the ETC, though, this is the one they mean. Look at slide 87, Fig. 9.13 pg. 173. You can see all the proteins lined up on a slant just like stairs. At the top is NADH. It donates its electrons to the first protein in the chain and is converted back to NAD+. (Recycling again) Those donated electrons then go to the next protein and the next, etc. until they get to the bottom where they are given to oxygen, who grabs a couple hydrogen ions and makes water. Now, the ETC is shown here going down because it’s on a kind of graph. Each protein molecule in the ETC is a bit more electronegative than the one before it and the electrons kind of get pulled along until oxygen, the most electronegative, gets the electrons. In real life they don’t really look like steps. Along the way, at each step, the electrons lose a bit of their energy ‘cause something else is also going on. Recall what we said earlier: if all the energy that was in a glucose molecule was released all at once, it would blow up the cell. So to harness that energy and make it useful, the cell has to break the glucose down in lots of steps. Glycolysis, the citric acid cycle, and now the ETC are those steps. So, what else is going on in this ETC? 12 UNIT 3 NOTES Electron transfer in the electron transport chain causes proteins to pump H+ from the mitochondrial matrix to the intermembrane space. As those electrons go from one protein to another, hydrogen ions, H+, get pumped out to the space between the inner and outer membranes of the mitochondrion. That’s what uses up a bit of the electron’s energy. Pushing all those H+s to one side of a membrane establishes an electrochemical gradient called a proton-motive force. It’s called that because it can do work. There is now a greater concentration of H+, as well as a higher positive charge, on one side of the membrane. What happens when there is a concentration gradient? Yup, diffusion, but in this case it is called chemiosmosis because the cell is going to use the H+ gradient to do work, the work of making ATP. No work is done in plain old diffusion. The H+s now want to come back across the membrane into the matrix to even out. When they do, they go through a special channel protein called ATP synthase. The ‘force’ of the H+s flowing through the ATP synthase in the membrane has enough energy to power the phosphorylation of ADP ATP. It even looks like a little motor. (slide 90, Fig.9.14 pg. 174) This is oxidative phosphorylation. The energy to do this came from the complete oxidation of glucose using an ETC. If there is no ETC involved then it is not oxidative phosphorylation. 13 UNIT 3 NOTES So what did the cell get for all this trouble? I understand that you have a headache now, but this isn’t about you, it’s about the cell. Only about half of the energy that was stored in the glucose gets transferred to ATP so the cell can use it to do work. Where did the rest go? Read the beginning of these notes again if you don’t remember. Respiration generates about 38 ATPs per glucose molecule. Why do we say about? Well, because that is an average. Look back at slide 87, Fig. 9.13 for a minute. Notice where FADH2 joins the ETC. It is not quite at the top. Because FADH2 joins at a lower level it is not able to pump as many H+s to the intermembrane space as NADH. The cell usually gets 3 ATP for each NADH and 2 ATP for each FADH2. Overall from one molecule of glucose, here is what the cell gets for a net total: ATPs ATPs NADHs FADH2s From ETC Total ATPs Glycolysis 2 2 0 4 6 Decarboxylation of Pyruvate 0 2 0 6 6 Krebs Cycle 2 6 2 22 24 Total 4 10 2 32 36 Why is the total 36 when I said respiration generates 38 ATP? Where did those two ATP go? As I said way back on pg. 8, the cell can use all kinds of nutrient molecules to get energy, not just glucose. Things like proteins, fats, and other carbs. When you eat all the molecules of nutrients are broken down into their building block molecules and enter into either glycolysis or the citric acid cycle at some point so ATP can be made. Slide 97, Fig. 9.20 pg. 180, shows you where different things would enter this pathway. This all depends on the needs of the cell at the time. Some of these building block molecules will be used as building blocks to make new stuff; some might just be put into storage. Whether the cell makes ATP or new proteins is regulated by negative feedback. If there is enough ATP, no need to make more, so the rate of respiration would be slowed. Not enough ATP, then the rate would increase. Remember those enzyme inhibitors from chapter 8? All of these reactions 14 UNIT 3 NOTES in this entire pathway are controlled by enzymes. Several of them would be points of regulation in this pathway where the product would act as the inhibitor by binding to the enzyme when there is plenty and shutting off the pathway temporarily. That’s negative feedback. So what does the cell do if there isn’t any oxygen? We know what it does NOT do and that’s aerobic respiration. If oxygen is not available NADH and FADH2 do not take their electrons to the ETC. If NADH can’t go to the ETC and be recycled to NAD+ then things are going to grind to a halt unless NADH can get rid of its H and electrons somewhere else. Where else can that be? Let’s find out. Glycolysis is very old and almost all cells use that to get some ATP. This reaction happens with (aerobic) or without (anaerobic) oxygen. We spent pages talking about the aerobic version of respiration. Anaerobic respiration is performed by critters that live in really, really harsh nasty places like hydrothermal vents at the bottom of the ocean and hot springs in Yellowstone where the temperature is like 600o F. They also use an ETC, but the final electron acceptor is not oxygen but sulphur, which is right below oxygen on the periodic table. Another way to get ATP and recycle NADH without oxygen is to couple glycolysis with fermentation which takes place in the cytoplasm, no mitochondria needed. Lots of bacteria, yeast, and other organisms use this. There are two types of fermentation: alcohol and lactic acid. Ring a bell? I hope so. Alcohol fermentation, yes, is used to make beer, wine, bread, etc. (In bread making the alcohol cooks away so, no, you can’t get drunk on bread.) The pyruvates generated by glycolysis do not go to the citric acid cycle but instead are converted to ethanol in two steps. The first step releases CO2 (which makes bread rise) and the second step recycles NAD+. This allows glycolysis to continue so the cell can make more ATP. How many net ATPs does the cell get from fermentation of one glucose molecule? Not many is it. That’s because the only place ATP is made is during glycolysis by substrate level phosphorylation. Lactic acid fermentation isn’t any more efficient. Here, pyruvate is reduced by NADH to become lactic acid (lactate). This one step regenerates the NAD+ so glycolysis can keep going, but no more ATPs are made. This process can be used to make yogurt. It is also used by your muscle cells when you are exercising so much oxygen gets scarce. Fermentation is OK for generating ATP if you are a bacteria or yeast, but if you are a large mammal with 50 trillion cells and you need to move around, you’re gonna 15 UNIT 3 NOTES need A LOT more ATP than that. That’s why we need to breathe oxygen. You and I can’t get enough energy without it. Plus, ever smell sulphur? That rotten egg smell? I don’t know about you, but I’d rather stick to breathing oxygen than have to start taking in sulphur- groooossssss! Even though aerobic respiration is much more efficient there are some organisms that can’t use it. Oxygen is toxic to them and they will die. They are called obligate anaerobes. If you are obligated to do something that means you have to and you have no choice, like going to school. These organisms have no choice but to not use and stay away from oxygen. Other organisms called facultative anaerobes can do both. If oxygen is available they’ll use; it if not they don’t. Glycolysis is one of the oldest known biological processes. It is performed by almost all organisms. Even most lowly prokaryotes perform glycolysis, as part of fermentation. Due to its importance in providing the cell with ATP it has been kept as organisms have evolved. Aerobic bacteria, that do not have mitochondria, can also use oxygen to complete respiration. The citric acid cycle happens in the cytoplasm and the ETC and ATP synthase are imbedded in their cell membrane. (another clue to evolution!) You have just completed two very challenging chapters. They are long and complicated. Other than the most basic stuff everything here is new to you. I expect you to be lost, confused, dumbfounded, crying, or at the very least in awe of the wonder that is life. There will be a lot more complicated stuff coming in future units. I’m telling you this now not to discourage you, but to make the point that if you aren’t getting it ASK!! I will not bite your head off. Ask in class, 10th period, whenever, but ask. The only stupid question is the one not asked. The more we all talk the more we’ll all learn. Now get ready for photosynthesis! CHAPTER 10 SECTIONS 1-3 What does photosynthesis do? Autotrophs, which are producers, make organic compounds from inorganic ones. Which ones? Plants are the main Autotrophs we will talk about. Where do they get their energy from? What makes plants green? I know, not all plants are green, but they all have chlorophyll of some kind. It comes in three flavors. (a, b, and c, really. That’s what they’re called.) Those that don’t 16 UNIT 3 NOTES look green have more of another kind of pigment that makes them a different color, like red. Red is caused by a pigment called anthocyanin. A pigment is a molecule that can absorb light energy to excite its electrons. Excited electrons move to a higher orbital because they have that extra energy. They think they’re better than the other electrons in their ground states. Different kinds of pigments can absorb light at different wavelengths or colors. Different colors have different amounts of energy. Red has less energy than blue, ROYGBIV. Pigments can only absorb specific amounts of energy which correspond to certain colors. Colors that are absorbed are used for their energy and colors that are not, are reflected back for you to see. So those green leaves are green because green is not absorbed but reflected. Chlorophyll doesn’t use green light. Most photosynthesis takes place in the leaves. Leaves are usually oriented on plants in such a way as to maximize the surface area exposed to the sun. If you were to stand over a plant and look down, you would see smaller leaves on top, larger on the bottom, and the leaves tend to spiral around the stem so that they are not directly on top of each other blocking the light. Smart aren’t they? It makes sense. If you put solar panels on your roof, you don’t stack them up; you spread them out so they all get sunlight, not just the one on top. Leaves also have little teeny tiny holes for gas exchange called stomata or stomates (one is a stoma). What gas that plants use goes in? What gas goes out? From chapter 6 you should know what a chloroplast looks like. It has a bunch of things that look like stacks of pancakes inside. These stacks are called grana (one is a granum) and each ‘pancake’ is called a thylakoid. Each thylakoid has a space inside, and the chlorophyll pigments are in the membranes. They are all surrounded by a thick fluid called stroma. (Pay attention to detail here this word has an r in it. Compare to one stomata above!) Like the mitochondrion, the chloroplast has a double membrane, and probably has evolved from some type of photosynthetic bacteria. (endosymbiotic theory) Respiration has three stages. Photosynthesis only has two: the light reactions and the Calvin cycle. There are lots of gory details to both of these, but we are going to learn the PG version just like we did for respiration, and not look at the R version. But, if you want that doctorate in biochem, or you are tired of counting sheep, go ahead and look. Just don’t blame me if you have nightmares. What, do you suppose, plants need for the light reactions? (Yes, it is that obvious) They are called the light reactions for a reason. They only take place when the sun 17 UNIT 3 NOTES is shining. The light shines on the chlorophyll and starts a whole chain reaction of stuff. (Stuff is a very scientific term. I use it a lot.) Where do the light reactions take place? The Calvin cycle is how the sugar, or food, the plant makes gets made. Where does that take place? Let’s take a closer look at the light reactions. The light reactions convert solar energy to the chemical energy of ATP and NADPH. The first thing that happens is that light has to shine on the photosystem in the thylakoid membrane. A photosystem has a reaction-center complex in the middle that contains chlorophyll a and the primary electron acceptor. It’s so special they don’t even tell us what it is. The reaction-center complex is surrounded by lightharvesting complexes that have a bunch of pigments to absorb solar energy. These pigments funnel energy to the chlorophyll a molecules which get excited and give an electron to the primary electron acceptor. For a visual look at slide 123, Fig. 10.12 pg. 193. This is how a photosystem works. You will NOT be asked to explain this on a test. There are two photosystems in the thylakoid membrane and, they usually work together. The first one is actually Photosystem II, or PS II. It is best at absorbing light of wavelength 680 nm, so its reaction-center chlorophyll a molecule is called P680. It’s just a real shorthand way for scientists to know which one they are talking about. Photosystem I (PS I) is best at absorbing a wavelength of 700 nm. The reaction-center chlorophyll a of PS I is called P700. During the light reactions, there are two possible routes for electron flow: cyclic and linear. The main route is linear electron flow because that produces ATP and NADPH, which are needed to make the sugar. Both photosystems are used. Here’s how it goes. You can follow along step by step in the PowerPoint starting with slide 127 or your book using Fig. 10.13. This is the part you really need to know. If you understand this paragraph then you understand photosynthesis: The sun shines. Photons hit the pigment molecules in PS II and they pass their energy around until P680 gets excited. The excited electron is given to the primary electron acceptor. Now P680 is missing an electron and we can’t have that. Some enzymes come along, split a water molecule, and strip the hydrogens of their electrons, giving them to P680, and releasing O2. P680 is now ready to be excited again. The primary electron acceptor now gives its electron to another ETC. As the electron travels down the ETC to PS I, protons (H+) will be pumped across the thylakoid membrane into the space setting up a PMF (proton-motive force). These protons will eventually flow back across the membrane to the stroma through ATP 18 UNIT 3 NOTES synthase and make ATP just like we saw in respiration (chemiosmosis). Meanwhile, the sun is still shining and PS 700 gets excited, gives its electron to the primary electron acceptor, and then accepts the electron coming in from the ETC. The electron from the primary electron acceptor goes to ANOTHER ETC and ends up in a protein called ferredoxin (Fd). Ferredoxin then gives the electron to an electron carrier called NADP+ reducing it to NADPH which will then go to the Calvin cycle. ***NOTE: the electron carrier here is NADP+ as opposed to the carrier we saw in respiration, NAD+. Try to remember P for photosynthesis so the correct carrier for photosynthesis is the one with the P in it. Before we look at the Calvin cycle, let’s compare chemiosmosis in chloroplasts and mitochondria: Both make ATP but use different energy sources: mitochondria get energy from the chemical bonds in food, and chloroplasts use light energy. In mitochondria, protons are pumped to the intermembrane space and drive ATP synthesis as they diffuse back into the mitochondrial matrix In chloroplasts, protons are pumped into the thylakoid space and drive ATP synthesis as they diffuse back into the stroma Slide 139, Fig. 10.16 pg. 196 is an excellent reference for comparison. Ok where are we? Oh, yah. What were the products we got from the light reactions and where are they? This is kinda confusing so I’ll help you out. Early on we had to split water to get electrons. That produced O2 that diffused out and will eventually end up in the air you are breathing. Then those pesky electrons traveled through some ETCs and helped make ATP which is in the stroma along with the electron carrier NADPH, both of which are needed by the Calvin cycle which, by coincidence, takes place…. In the stroma! Slide 141, Fig. 10.17 pg. 197 shows the whole thing in splendid Technicolor. So, on to the Calvin cycle. This part is simple. It is a cycle. Therefore, it ends where it begins just like the citric acid cycle, and you DO NOT need to know any of the steps around the cycle. You do need to know what goes in and what comes out. The cycle builds sugar from smaller molecules by using ATP and the reducing power of electrons carried by NADPH. The cycle gets the carbon for the sugar from CO2. Since CO2 only has one carbon and glucose has six, the cycle needs to operate six times to make one glucose. After 3 turns one molecule of G3P is made which is either combined with another one to make glucose, or made into starch for storage. The beginning of the cycle is carbon fixation. This is where the carbon in the carbon dioxide is taken into the cycle to be processed and its fate is sealed. It’s fixed. The enzyme responsible for this is rubisco, which is thought to be the most abundant protein 19 UNIT 3 NOTES on Earth. Since no light is required for this cycle it is often performed by plants at night. The last slide 145, Fig. 10.21 pg. 203 is a great summary that I’ll leave you with. In summary: Photosynthesis has 2 sets of reactions: o Light reactions split water Hydrogens are used to produce a reduced coenzyme, NADPH, and ATP Oxygen is given off o Once the NADPH and ATP are formed the rest of the reactions do not require light CO2 is reduced to glucose in a set of cyclic reactions: the Calvin cycle Fig. 10-21 H2O CO2 Light NADP+ ADP + P i Light Reactions: Photosystem II Electron transport chain Photosystem I Electron transport chain RuBP ATP NADPH 3-Phosphoglycerate Calvin Cycle G3P Starch (storage) Chloroplast O2 Sucrose (export) 20