Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
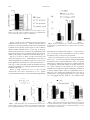
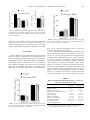
56, 150 –155 (2000) Copyright © 2000 by the Society of Toxicology TOXICOLOGICAL SCIENCES Effects of Acetaminophen on Preimplantation Embryo Glutathione Concentration and Development in Vivo and in Vitro Delia N. Laub, Nawal O. Elmagbari, Nura M. Elmagbari, Melissa A. Hausburg, and Catherine S. Gardiner 1 Department of Biology, University of Northern Colorado, Greeley, Colorado 80639 Received December 28, 1999; accepted February 25, 2000 This study investigated the effects of high doses of acetaminophen (APAP) on preimplantation embryos. Previous studies indicate that cleavage-stage embryos cannot synthesize reduced glutathione (GSH) de novo and may be sensitive to GSH-depleting toxicants. Alternatively, there may be maternal mechanisms that protect the embryos from the adverse effects of these toxicants. To address these possibilities, we cultured two-cell stage embryos in 0, 375, 750, or 1500 M APAP and evaluated GSH concentration and development. APAP depressed embryo development to the morula and blastocyst stages in vitro, but a decrease in embryo GSH concentration was not detected. Furthermore, administration of 800 or 1430 mg/kg APAP to female mice 12 h prior to embryo collection on day 2 of gestation, or administration of 800 mg APAP/kg/day from day – 8 to day 1 or day 3 of gestation, did not significantly affect ovary or embryo GSH concentration or embryo development. Liver GSH, however, was significantly decreased. Moreover, no adverse effects on embryo development to term were observed after treatment of female mice with 1430 mg APAP/kg/day from day – 8 to day 3 of gestation. In summary, in vitro embryos were adversely affected, in terms of development, by APAP. In vivo, large doses of APAP depleted liver GSH but did not affect development of preimplantation embryos. In conclusion, preimplantation embryos appear to be protected from GSH-depleting toxicants such as APAP in vivo. Key Words: preimplantation; glutathione; acetaminophen; embryo. Acetaminophen (APAP) is commonly used for its analgesic and antipyretic properties and is very safe under normal conditions (Sipes and Gandolfi, 1991). In the liver, glucuronidation and sulfation metabolize APAP, with only a small amount entering the P-450 oxygenase system. The P-450 system converts APAP into a highly reactive intermediate, N-acetylbenzoquinone imine (NAPQI), which can conjugate with reduced glutathione (GSH). A high dose of APAP will saturate the glucuronidation and sulfation pathways and increase the amount of APAP metabolized by the P-450 oxygenase system. Consequently, NAPQI levels increase, leading to depletion of hepatic stores of GSH. Eventually, NAPQI will bind with 1 To whom correspondence should be addressed. Fax: (970) 351-2335. Email: [email protected]. cellular macromolecules, causing tissue necrosis and death. Mitchell et al. (1973) administered one dose of 375 mg/kg APAP ip to mice and found liver necrosis in 45% of the animals examined. Furthermore, it has been demonstrated that hepatoxicity increases when female mice are ovariectomized (Raheja et al., 1983). Finally, the LD50 of APAP in mice has been measured as 4.11% APAP or 5877 mg APAP/kg/day for a 14-day exposure to APAP mixed with mouse chow (Reel et al., 1992). Earlier studies have shown that APAP can cause adverse effects on embryos or fetuses at high doses. Mice exposed to doses up to 1430 mg APAP/kg/day in their food, using a continuous breeding protocol, exhibited a decrease in the number of litters per pair, post-natal weight gain, and a decrease in birth weight of F2 pups (Reel et al., 1992). Stark et al. (1989a) found open neural tubes in day 9 rat embryos cultured for 24 h with 0.50 mM APAP, while Weeks et al. (1990) found open neural tubes as well as incomplete body curvature in rat embryos cultured with 300 –750 M APAP from days 9.5 to 11.5 of gestation. Furthermore, APAP overdoses in pregnant women have led to birth defects and fetal death (Haibach et al., 1984). However, the sensitivity of the preimplantation embryo to APAP, both in vitro and in vivo, has not been previously examined. In the reduced form, the endogenous tripeptide, glutathione (GSH, ␥-glutamycysteinylglycine), serves to protect cells from oxidative stress, aids in metabolism of toxicants, and functions as a transport form of cysteine (Reed, 1990). GSH also has an important functional role in reproduction and early development. During normal fertilization, high levels of GSH must be present in an oocyte for sperm nuclear decondensation to occur (Perreault et al., 1988). When mice are treated with a GSH synthesis inhibitor, L-buthionine S,R-sulfoximine, oocytes showed a decrease in GSH content from 1.8 pmol to 0.20 pmol GSH per ovum (Calvin et al., 1986). In addition, depletion of GSH in mouse oocytes can adversely affect sperm nuclear decondensation and microtubule function, which consequently impairs embryonic development (Zuelke et al., 1997). During normal embryo development, GSH concentration decreases 90% during the period from the unfertilized oocyte to the blastocyst stage (Gardiner and Reed, 1994). In addition, em- 150 EFFECTS OF ACETAMINOPHEN ON PREIMPLANTATION EMBRYOS bryos cannot synthesize GSH de novo until the blastocyst stage, which may leave them particularly susceptible to the effects of toxicants (Gardiner and Reed, 1995). Recently, GSH was discovered in the reproductive tract fluid of mice and was found to improve development of preimplantation embryos after chemically induced depletion of GSH (Gardiner et al., 1998). Because early embryos cannot synthesize GSH (Gardiner and Reed, 1995), we hypothesize that they may be very sensitive to the effects of glutathione-depleting toxicants such as APAP. An alternative hypothesis is that there are maternal mechanisms that function to decrease exposure of embryos to toxicants and to protect the embryos during preimplantation development. We conducted 4 experiments to begin to address both of these hypotheses. In the first experiment, we investigated the direct effect of APAP supplementation in culture medium on preimplantation embryos. We also examined the effects of one dose of APAP on the embryos, ovaries, and liver of pregnant mice allowed to recover for 12 h in vivo. Next, we determined the effects of daily APAP exposure in vivo throughout preimplantation development on liver, ovary, and embryo GSH, as well as embryo development at day 3 of gestation. Finally, we investigated the effect of APAP on fetal development to term when the dam was given APAP daily from 8 days prior to ovulation until day 3 of gestation. MATERIALS AND METHODS Materials. Chemicals necessary for our experiments were purchased from Sigma (St. Louis, MO) and mice were bred from NSA mice from Harlan (Indianapolis, IN). Treatment of mice with acetaminophen. The acetaminophen was suspended in a 0.5% tragacanth solution and given to female pubertal mice intragastrically at 800 or 1430 mg/kg of body weight. The control group received only the 0.5% tragacanth solution. Embryo collection and evaluation. Female pubertal NSA mice were synchronized and superovulated by intraperitoneal injection with 10 IU equine chorionic gonadotropin and 44 – 48 h later with 5 IU of human chorionic gonadotropin. Females were bred with proven breeder males and were checked the next day for a copulation plug (designated as day 0 of gestation). Females were euthanized on day 1 (d1), day 2 (d2), or day 3 (d3), and 2-cell, morula, or blastocyst stage embryos were collected by flushing M16 medium through the oviducts and/or uteri (Hogan et al., 1994). Embryos were then washed by sequential passage though drops of M16 medium with 4 mg/ml bovine serum albumin (BSA). Embryos cultured in vitro were placed into 10 l drops (10 embryos per drop) of M16 medium with 4 mg/ml BSA and covered with mineral oil. Embryos were cultured in a humidified atmosphere of 5% CO 2 in air at 37°C. Daily microscopic evaluation of embryos consisted of noting fertilization, classifying embryos into their respective developmental stages, and identifying degenerate embryos. Degenerate embryos were not used for GSH analysis. Liver and ovary collection and evaluation. Portions of the liver and both ovaries were dissected from euthanized female mice and weighed. The overall morphology of the tissues was assessed, and the samples were prepared for quantification of GSH. Fetus collection and evaluation. Female pubertal mice were bred with proven breeder males on day –1. On d17 of gestation, dams were euthanized and fetuses were removed via cesarean section. Maternal and fetal evaluation 151 FIG. 1. Embryo development in vitro when cultured with APAP from the 2-cell stage (d1). Developmental stages are indicated as follows: deg (degenerate), 4 – 8 (4-cell to 8-cell), M (compacted morula), B (blastocyst), XB (expanded blastocyst), IH (initiating hatching blastocyst), and H (hatched blasto cyst). Within a developmental stage, columns not accompanied by the same letter (a,b,c,d) are significantly different (p ⬍ 0.05). Values are means ⫾ standard error. consisted of recording the number of resorptions in the uterus, weighing the intact uterus with the fetuses, weighing the pregnant dam, examining each fetus for gross malformations, weighing individual fetuses, and noting the number of pups per litter. Detection of GSH and GSSG by HPLC. Pools of 20 –30 embryos, liver samples, and ovary samples were homogenized in perchloric acid-diethylenetriaminepentaacetic acid solution and prepared for GSH and GSSG detection via high performance liquid chromatography (Martin and White, 1991). Each sample was fluorescently labeled with dansyl chloride and subsequently analyzed on an amino propyl silica column using a methanol-sodium acetate gradient system (Reed et al., 1980). The detection limit for this method is 1 pmol of GSH. Effects of APAP in culture medium on embryo development and GSH concentration in vitro. Embryos were collected on d1, washed, and placed into culture drops at the 2-cell stage with 0, 375, 750, or 1500 M APAP in M16 with 4 mg/ml BSA. Embryos were derivatized on d2 (at the 4-cell to compacted morula stage) for GSH quantification, and development was assessed on d2, d3, d4, and d5. Effects of one dose of APAP on GSH concentration and embryo development in vivo. Mice found to have copulation plugs on d0 were treated on d2 with 0, 800, or 1430 mg APAP/kg. Mice were euthanized, and samples were collected 12 h after dosing. Effects of APAP treatment for 10 or 12 days on GSH concentration and embryo development in vivo. Female mice were treated with 0 or 800 mg/kg APAP daily from 8 days prior to ovulation until the morning of d1 or d3 of gestation. Embryos and other tissues were collected on the afternoon of d1 or d3. Effects of preimplantation exposure with APAP on development to term. Female mice were treated with 0 or 1430 mg/kg APAP daily starting 8 days prior to ovulation and continuing until 3 days after copulation plugs were detected. Fetuses were removed by cesarean section on d17 of gestation. Statistical analysis. Morphological development was analyzed using Kruskal-Wallis analysis of variance. Analysis of variance and least-significantdifference procedures were used to determine differences in GSH and GSSG content of embryos, liver, and ovaries. A t-test was used to analyze the number of fetuses per dam, number of resorptions per dam, individual fetal weight, total fetal weight, uterine weight, liver weight, and weight of dam on d17. Chi-square analysis was used to analyze the number of mice with copulation plugs and the number of deceased mice. Statistica software was used for statistical analysis. All experiments were repeated at least twice with a minimum of 3 replications for each treatment each time. 152 LAUB ET AL. FIG. 2. Embryo GSH after treatment in vitro with APAP for 24 h. Embryos were at the 4-cell to compacted morula stage. No significant differences were detected. Values are means ⫾ standard error. RESULTS Effects of APAP in culture medium on embryo development and GSH concentration in vitro. Embryos collected on d1 of gestation (2-cell stage) and cultured with varied concentrations of APAP did not develop to later stages as well as embryos cultured in M16 with BSA alone (p ⬍ 0.05, Fig. 1). However, GSH concentration on day 2 (at the 4-cell to compacted morula stage) did not decrease to correspond with the lack of development in the groups of embryos treated with APAP (Fig. 2). Effects of one dose of APAP on GSH concentration and embryo development in vivo. Administration of one dose of APAP intragastrically to pregnant female mice on d2 of gestation significantly decreased liver GSH concentrations (p ⬍ 0.05, Fig. 3). Ovary and embryo GSH concentrations as well as embryo development (Fig. 4) did not significantly differ due to treatment with APAP. Effects of APAP treatment for 10 or 12 days on GSH concentration and embryo development in vivo. Figures 5 and 6 demonstrate that liver GSH concentration significantly FIG. 3. GSH concentrations 12 h after treatment with APAP on d2 of gestation. Within a tissue type, columns not accompanied by the same letter (a,b) are significantly different (p ⬍ 0.05). Values are means ⫾ standard error. FIG. 4. Embryo development in vivo 12 h after treatment with APAP on d2 of gestation. No significant differences were detected. Values are means ⫾ standard error. decreased due to treatment with APAP (p ⬍ 0.05); however, ovary and embryo GSH concentrations as well as embryo development (Figs. 7 and 8) were not affected. Effects of preimplantation exposure with APAP on development to term. To assess how exposure during preimplantation development affects fetal development to term, we exposed female mice to APAP from 8 days prior to ovulation until day 3 of pregnancy and allowed embryos to develop until d17 in vivo. Upon evaluation of the females on d17 of gestation, maternal body weight did not differ significantly, nor did liver or uterine weight (Table1). However, significantly fewer female mice were found with copulation plugs each day of breeding in the 1430 mg/kg APAP group (p ⬍ 0.05; Control 18 ⫾ 4% plugged each day, APAP 4 ⫾ 2%). While no mice FIG. 5. GSH concentration after daily APAP treatment from 8 days prior to ovulation until the first day of gestation. Within a tissue type, columns not accompanied by the same letter (a,b) are significantly different (p ⬍ 0.05). Values are means ⫾ standard error. 153 EFFECTS OF ACETAMINOPHEN ON PREIMPLANTATION EMBRYOS FIG. 6. GSH concentration after daily APAP treatment from 8 days prior to ovulation until d3 of gestation. Within a tissue type, columns not accompanied by the same letter (a,b) are significantly different (p ⬍ 0.05). Values are means ⫾ standard error. died in the control group, 9 mice treated with APAP died during the course of the study. No gross malformations were found, but individual fetuses from females treated with APAP weighed significantly more than fetuses from the control dams. DISCUSSION Previous studies have shown that embryos do not possess the capacity to synthesize glutathione de novo until the blastocyst stage of development (Gardiner and Reed, 1995). Furthermore, preimplantation embryos exhibit a 90% decrease in glutathione concentration from the unfertilized oocyte to the blastocyst stage (Gardiner and Reed, 1994). This may leave preimplantation embryos susceptible to the effects of toxicants such as APAP that are normally detoxified by GSH. Alternatively, FIG. 8. Embryo development after treatment with APAP from 8 days prior to ovulation until day 3 of gestation. No significant differences were detected. Values are means ⫾ standard error. there may be maternal mechanisms present to protect the embryos during this sensitive period. Our first experiment demonstrated that development to the morula and blastocyst stages was depressed in embryos cultured in 375, 750, and 1500 M APAP. Previous studies have demonstrated that APAP inhibits Na ⫹/K ⫹-ATPase (Corcoran et al., 1987), an enzyme important in blastocoel formation (Gardiner and Menino, 1993). This may partially explain our observed decrease in embryo development to later stages. Moreover, our results are consistent with studies of the effects of APAP on postimplantation development of rat embryos. Rat embryos cultured in APAP (0.50 mM) on d9, d10, or d11 of gestation exhibit an increased incidence of neural tube defects (Stark et al., 1989b) and decreases in embryonic length and yolk sac protein (Stark et al., 1989a). Furthermore, previous TABLE 1 Effects of Daily Dosing with APAP from Day ⴚ8 to Day 3 of Gestation on Development to Term Number of females Weight of dam on d17 (g) Number of mice with copulation plugs Weight of liver (g) Uterine weight (g) Number of fetuses/dam Number of resorptions/dam Individual fetal weight (g) Total fetal weight (g) Number of malformations FIG. 7. Embryo development after treatment with APAP from 8 days prior to ovulation until the first day of gestation. No significant differences were detected. Values are means ⫾ standard error. 0 mg/kg/day 1430 mg/kg/day 36 50.1 ⫾ 1.2 25* 3.0 ⫾ 0.1 15.6 ⫾ 1.1 11.7 ⫾ 0.9 1.5 ⫾ 0.6 0.90* ⫾ .01 10.5 ⫾ 0.9 0 36 49.7 ⫾ 2.6 8* 3.2 ⫾ 0.1 13.8 ⫾ 2.2 9.7 ⫾ 1.9 2.8 ⫾ 1.0 1.01* ⫾ 0.02 9.7 ⫾ 1.6 0 Note. Values are means ⫾ standard error unless otherwise indicated. * Values significantly different from each other ( p ⬍ 0.05). 154 LAUB ET AL. studies indicate that depletion of embryonic GSH concentration prior to APAP treatment causes an increase in the incidence of embryonic defects (Stark et al., 1989b), while N-acetylcysteine protects cultured d9 rat embryos from the embryotoxic effects of 0.30 mM APAP in vitro (Weeks et al., 1990). At high doses, cytochrome P-450 monooxygenases convert APAP to its reactive metabolite, NAPQI, which subsequently depletes GSH, leading to toxicity. Previous studies have detected cytochrome P-450 activity in preimplantation embryos at the blastocyst stage (Pedersen et al., 1985). Cytochrome P4501A1 gene expression has also been detected as early as the oocyte stage (Dey and Nebert, 1998). If the monooxygenase system is functioning during preimplantation development, APAP could be metabolized to NAPQI, causing a decrease in GSH concentration. Our data did not show a significant decrease in embryo GSH at d2 of gestation after 24 h of culture in APAP when embryos were at the morula stage. This may be evidence of APAP toxicity without GSH depletion. It is also possible that GSH levels had already recovered or GSH was not yet depleted at the time of our measurement. Alternatively, differences in GSH concentration due to APAP may have been present but were not detectable due to variation in samples. Previous studies show that toxicants can induce GSH synthesis by increasing transcription of the catalytic subunit of the ratelimiting enzyme in GSH synthesis (Shi et al., 1994). APAP could have induced early GSH synthesis, which would have allowed the embryos to recover their intracellular GSH. Alternatively, embryo development could have been adversely affected by means other than depletion of GSH pools. After the direct effects of APAP were assessed, we examined the effects of APAP in vivo. APAP was administered to female mice, and effects were examined 12 h after one dose or after 10 to 12 days of daily dosing. No significant differences in ovary GSH concentration, embryo GSH concentration, or embryo development were observed. Similar to previous studies (Mitchell et al., 1973), however, liver GSH concentration decreased significantly following one or several doses of APAP. Earlier studies indicate that hepatoxicity increases in pregnant mice (Larrey et al., 1986). These data confirmed that the doses of APAP administered were high enough to deplete GSH. These results suggest that the preimplantation embryo is at least partially protected from APAP in vivo and that the reproductive tract does not experience significant GSH depletion due to the doses examined. One likely mechanism of protection is the hepatic metabolism of a significant amount of the APAP, which resulted in decreased exposure of the preimplantation embryo to the APAP. If the concentration of APAP in the reproductive tract fluid were the same as in the in vitro studies, the embryos may have exhibited similar adverse effects in vivo as were seen in vitro. To determine whether exposure of embryos to APAP in vivo during the preimplantation period leads to toxic effects on development to term, female mice were treated with 1430 mg/kg/day APAP 8 days prior to ovulation and throughout development to the blastocyst stage. Subsequently, effects on fetuses at term were examined. Table 1 shows that weight of dams on d17, liver weight, uterine weight, number of fetuses per dam, number of resorptions per dam, and total fetal weight were not significantly different. However, individual fetal weights were higher in dams treated with APAP. This effect may be due to the presence of a smaller number of fetuses per dam and a higher number of resorptions in the uterus. Although there was a trend for an increased number of resorptions and a decreased number of fetuses in APAP-treated dams, the effect was not significant. Furthermore, we did not detect any gross malformations of fetuses in either group. Similarly, Lum and Wells (1986) found that administration of one ip dose of APAP does not cause cleft palate in mice. However, 9 out of 36 mice in our APAP group died during the course of treatment, indicating that a toxic dose of APAP was administered. We found significantly fewer female mice with copulation plugs in the APAP group. It is not clear whether this was a physiological effect causing a delay or absence of estrus and ovulation or an effect on copulation behavior. Reel et al. (1992) also treated mice with doses of APAP up to 1430 mg/kg administered in mouse chow, using a continuous breeding protocol, and found a decrease in the number of litters per pair, an increase in the presence of abnormal sperm, and a decrease in post-natal weight of the pups. Our results would suggest that these effects were not due to toxicity at the preimplantation stages of development. We conclude that preimplantation embryos are very sensitive to APAP toxicity in vitro. In vivo, however, there appear to be maternal mechanisms protecting the embryos or decreasing the exposure of the embryos to APAP. The liver may metabolize and conjugate enough APAP such that only very low levels reach the reproductive tract. In support of this hypothesis, our study did not detect a significant decrease in ovarian GSH concentration. Furthermore, toxic concentrations of APAP may not enter the reproductive tract fluid in the lumen of the uterus where the preimplantation embryos are developing. Another possibility is that uterine fluid may harbor substances that protect the preimplantation embryo, as demonstrated by a previous study indicating that GSH is present in uterine and oviductal fluid and has beneficial effects on preimplantation development (Gardiner et al., 1998). Further studies are required to elucidate the maternal mechanisms that are protecting the embryos from the adverse effects of high doses of APAP. ACKNOWLEDGMENTS The authors would like to thank James Salmen, Gene Gushansky, Shawn Stover, Analouise Matt, and Leslie Calaustro for their assistance. This work was supported by the National Institute of Environmental Health Sciences, NIH grant ES08818. EFFECTS OF ACETAMINOPHEN ON PREIMPLANTATION EMBRYOS REFERENCES Calvin, H. I., Grosshans, K., and Blake, E. J. (1986). Estimation and manipulation of glutathione levels in prepuberal mouse ovaries and ova: Relevance to sperm nucleus transformation in the fertilized egg. Gamete Res. 14, 265–275. Corcoran, G. B., Chung, S. J., and Salazar, D. E. (1987). Early inhibition of the Na ⫹/K ⫹-ATPase ion pump during acetaminophen-induced hepatotoxicity in rat. Biochem. Biophys. Res. Commun. 149, 203–207. Dey, A., and Nebert, D. W. (1998). Markedly increased constitutive CYP1A1 mRNA levels in the fertilized ovum of the mouse. Biochem. Biophys. Res. Commun. 251, 657– 661. Gardiner, C. S., and Menino, A. R., Jr. (1993). Development of Na ⫹/K ⫹ATPase activity and blastocoel formation. In Preimplantation Development (B. D. Bavister, Ed.), pp. 200 –211. Springer-Verlag, New York. Gardiner, C. S., and Reed, D. J. (1994). Status of glutathione during oxidantinduced oxidative stress in the preimplantation mouse embryo. Biol. Reprod. 51, 1307–1314. Gardiner, C. S., and Reed, D. J. (1995). Synthesis of glutathione in the preimplantation mouse embryo. Arch. Biochem. Biophys. 318, 30 –36. Gardiner, C. S., Salmen, J. J., Brandt, C. J., and Stover, S. K. (1998). Glutathione is present in reproductive tract secretions and improves development of mouse embryos after chemically induced glutathione depletion. Biol. Reprod. 59, 431– 436. Haibach, H., Akhter, J. E., Muscato, M. S., Cary, P. L., and Hoffmann, M. F. (1984). Acetaminophen overdose with fetal demise. Am. J. Clin. Pathol. 82, 240 –242. Hogan, B., Beddington, R., Costantini, F, and Lacy, E. (1994). Manipulating the mouse embryo: A Laboratory Manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York. Larrey, D., Letteron, P., Foliot, A., Descatoire, V., Degott, C., Geneve, J., Tinel, M., and Pessayre, D. (1986). Effects of pregnancy on the toxicity and metabolism of acetaminophen in mice. J. Pharmacol. Exp. Ther. 237, 283–291. Lum, J. T., and Wells, P. G. (1986). Pharmacological studies on the potentiation of phenytoin teratogenicity by acetaminophen. Teratology 33, 53–72. Martin, J., and White, I. N. (1991). Fluorimetric determination of oxidised and reduced glutathione in cells and tissues by high-performance liquid chromatography following derivatization with dansyl chloride. J. Chromatogr. 568, 219 –225. Mitchell, J. R., Jollow, D. J., Potter, W. Z., Gillette, J. R., and Brodie, B. B. (1973). Acetaminophen-induced hepatic necrosis: IV. Protective role of glutathione. J. Pharmacol. Exp. Ther. 187, 211–217. 155 Pedersen, R. A., Meneses, J., Spindle, A., Wu, K., and Galloway, S. M. (1985). Cytochrome P-450 metabolic activity in embryonic and extraembryonic tissue lineages of mouse embryos. Proc. Natl. Acad. Sci. U.S.A. 82, 3311– 3315. Perreault, S. D., Barbee, R. R., and Slott, V. L. (1988). Importance of glutathione in the acquisition and maintainence of sperm nuclear decondensing activity in maturing hamster oocytes. Dev. Biol. 125, 181–186. Raheja, K. L., Linscheer, W. G., and Cho, C. (1983). Hepatotoxicity and metabolism of acetaminophen in male and female mice. J. Tox. Environ. Health. 12, 143–158. Reed, D. J. (1990). Chemical mechanisms of drug-induced liver injury. In Hepatology: A Textbook of Liver Disease. (D. Zakim and T. D. Boyer, Eds.), pp. 737–753. W. B. Saunders, Philadelphia. Reed, D. J., Babson, J. R., Beatty, P. W., Brodie, A. E., Ellis, W. W., and Potter, D. W. (1980). High-performance liquid chromatography analysis of nanomole levels of glutathione, glutathione disulfide, and related thiols and disulfides. Anal. Biochem. 106, 55– 62. Reel, J. R., Lawton, A. D., and Lamb, J.C., IV (1992). Reproductive toxicity evaluation of acetaminophen in Swiss CD-1 mice using a continuous breeding protocol. Fundam. Appl. Toxicol. 18, 233–239. Shi, M. M., Kugelman, A., Iwamoto, T., Tian, L., and Forman, H. J. (1994). Quinone-induced oxidative stress elevates glutathione and induces ␥-glutamylcysteine synthetase activity in rat lung epithelial L2 cells. J. Biol. Chem. 269, 26512–26517. Sipes, I. G., and Gandolfi, A. J. (1991). Biotransformation of toxicants. In Casarett and Doull’s Toxicology: The Basic Science of Poisons, 4th ed. (M. O. Amdur, J. Doull, and C. D. Klaassen, Eds.), pp. 88 –126. Pergamon Press, New York. Stark, K. L., Harris, C., and Juchau, M. R. (1989a). Modulation of the embryotoxicity and cytotoxicity elicited by 7-hydroxy-2-acetylaminofluorene and acetaminophen via deacetylation. Toxicol. Appl. Pharmacol. 97, 548 –560. Stark, K. L., Harris, C., and Juchau, M. R. (1989b). Influence of electrophilic character and glutathione depletion on chemical dysmorphogenesis in cultured rat embryos. Biochem. Pharmacol. 38, 2685–2692. Weeks, B. S., Gamache, P., Klein, N. W., Hinson, J. A., Bruno, M., and Khairallah, E. (1990). Acetaminophen toxicity to cultured rat embryos. Teratog. Carcinog. Mutagen. 10, 361–371. Zuelke, K. A., Jones, D. P., and Perreault, S. D. (1997). Glutathione oxidation is associated with altered microtubule function and disrupted fertilization in mature hamster oocytes. Biol. Reprod. 57, 1413–1419.