Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
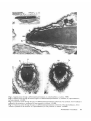
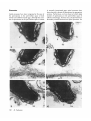
Fertilization in ascidians: apical processes and gamete fusion in Ciona intestinalis spermatozoa MAKOTO FUKUMOTO Biological Institute, College of General Education, Nagoya City University, Nagoya 467, Japan Summary This paper describes the fine structure of the spermatozoan apex and its morphological changes during the process of fertilization in Ciona intestinalis. At the apex of the head an acrosome is present in the form of a flattened vesicle containing moderately electron-dense material with an electron-dense plate in its centre. Contiguous to the acrosome, an apical substance is located in the anterior-most tip of the sperm head. The spermatozoa that bind to and penetrate the chorion of both normal and caffeine-treated eggs have an intact acrosome and do not show an observable alteration of the plasmalemma enclosing the apex of the head. On the other hand, spermatozoa in the perivitelline space of both normal and caffeine-treated eggs lack an acrosome. Instead of an acrosome, apical processes are observed at the apex of the spermatozoa in the perivitelline space of both normal and caffeinetreated eggs. Gamete fusion seems to occur between the egg membrane and some of these apical processes at the tip of the sperm head. The apical process reported here is thought to be analogous to the acrosomal process in other marine invertebrates. The apical processes that are induced in the perivitelline space have not been previously described in ascidians. Introduction through the chorion in Halocynthia mretzi (Ishikawa et al. 1978; Fuke-Tsukamoto, 1983). Test cells are located in the perivitelline space between the egg surface and the chorion. The role of the test cells in fertilization still remains enigmatic. Such an elaborate set of egg investments leads to the supposition that spermatozoa of ascidians may be highly modified to effect penetration through the envelope and achieve successful fusion with the female gamete. Recent studies have shown that ascidian spermatozoa have a small acrosome (Cloney & Abbott, 1980; Fukumoto, 1985, 1986). Furthermore, there is evidence that spermatozoa of ascidians contain proteases that act as lysins (Woollacott, 1977; Hoshi et al. 1981; Sawada et al. 1982, 1984a,b). It is not clear how the acrosome participates in the process of fertilization and where the lysins are located in the spermatozoa. In order to resolve these issues it is necessary to investigate the morphological changes at the apex of spermatozoa during the process of fertilization. In spite of a fairly large number of descriptive and experimental studies on ascidian fertilization, our knowledge of the events associated with fertilization is still poor and fragmentary in both biochemical and morphological aspects. The ascidian egg is enclosed by a relatively thick and tough chorion (vitelline coat) to which spermatozoa bind as a prerequisite for fertilization (Rosati & DeSantis, 1978; DeSantisei al. 1980; Honegger, 1982; FukeTsukamoto, 1983; Hoshi et al. 1985) and which might participate in the block to polyspermy in Ascidia nigra (Lambert, 1986). The chorion, which is externally decorated by a single layer of highly vacuolated follicle cells, presents a barrier to successful penetration by the spermatozoa. The follicle cells are thought to be involved in sperm attraction in Ciona intestinalis (Miller, 1975), in egg flotation in Corella willmeriana (Lambert & Lambert, 1978) and in sperm penetration Journal of Cell Science 89, 189-196 (1988) Printed in Great Britain © The Company of Biologists Limited 1988 Key words: fertilization, ascidians, Ciona intestinalis, spermatozoa. 189 With the electron microscope it is difficult to detect gamete fusion and morphological changes in spermatozoa during the process of fertilization, especially in the perivitelline space, because of the limited number of spermatozoa that can pass through the chorion, even if inseminated at high concentrations of sperm. Recently the author has found that many spermatozoa penetrate the chorion and reach the perivitelline space when caffeine- or theophylline-treated eggs are inseminated by fairly high concentrations of spermatozoa, resulting in polyspermic fertilization. This has provided us with useful material for observing how gamete fusion occurs and any structural changes taking place in C. intestiualis spermatozoa prior to gamete fusion in the perivitelline space. This paper describes the apical processes of C. intestinalis spermatozoa in the perivitelline space in both normal and caffeine-treated eggs and gamete fusion in caffeine-treated polyspermic eggs. Materials and methods Ciona intestinalis was collected on the coast of Chita Peninsula, Aichi Prefecture, Japan. For fine-structural observations on differentiated spermatozoa, the sperm duct, which is filled compactly with differentiated spermatozoa, was fixed in 3 % glutaraldehyde in 0-1 M-cacodylate buffer (pH 7-4) for 1 h at room temperature. For observations on the spermatozoa suspended in both normal sea water and caffeine-containing sea water (S mMcaffeine), sperm that were carefully obtained from the sperm duct by pipetting were suspended for lOmin in normal sea water or caffeine-containing sea water. They were centrifuged and fixed in 3 % glutaraldehyde in 0-1 M-cacodylate buffer (pi I 7-4) for 30min at room temperature. For observations on both normally fertilized and polyspermic eggs, the eggs obtained from the oviduct by careful puncture with a razor were divided into two groups. One group was suspended in normal sea water for 5 min and then cross-fertilized with a fairly high concentration of spermatozoa (approx. 108 sperm ml"'). The other group was suspended in caffeinecontaining sea water (5 mM-caffeine) for 5 nun (caffeine pretreatment) and cross-fertilized, after removal of surplus caffeine-containing sea water, with the same concentrations of spermatozoa. These eggs were fixed at 3-5 min after insemination in 3 % glutaraldehyde in 0-1 M-cacodylate buffer (pH 7-4) for 30 min at room temperature. These specimens were prepared for electron microscopy according to a method described by Fukumoto (1981). Electron micrographs were taken on a Hitachi H-300 electron microscope operated at 75 kV. Results Fine structure of the apex of spermatozoa The spermatozoon of C. intestinalis has architectural features that are characteristic of the ascidian spermatozoa described by Cloney & Abbott (1980) and 190 M. Fukumoto Fukumoto (1986). It has a moderately elongated head, approximately 5 jitm long, with a wedge-shaped tip, and a single mitochondrion, which is closely applied laterally to the nucleus (Fig. 1). In appropriate sagittal (Fig. 2) and transverse (Fig. 3) sections through the apex of the head an acrosome is seen. The acrosome is a flattened vesicle, about 150 nm X 160 nm X 60 nm, filled with moderately electron-dense material with an electron-dense plate in its central region. An accumulation of granular material, approximately 5-7 nm in diameter, is observed at the anterior tip of the head contiguous to the acrosome (Figs 2, 4), and is referred to here as apical substance. Such an accumulation of electron-dense substance was first reported at the apex of the differentiated spermatozoa in Ascidia callosa, where it was assumed to correspond to the periacrosomal substance found in some animal species (Cloney & Abbot, 1980). In C. intestinalis, Rosati et al. (1985) found similar electron-dense material at the apex of the sperm head. The inner and the outer nuclear membranes under the acrosome make close contact with each other to form a specialized structure that appears to form a pedestal for the acrosome (Figs 3, 4, arrows). A fuzzy material decorates the external surface of the plasmalemma (Figs 2, 3, 4). Apical processes and gamete fusion The binding of the spermatozoa to the chorion seems to be established by the fuzzy material on the plasmalemma of the apex adhering to the outer fibrous structures of the chorion (Figs 7, 8). There is evidence that this binding is mediated by an enzyme-substrate reaction between a-L-fucosidase on spermatozoa and fucosylated glycoproteins located on the fibrous structures of the chorion (Hoshi et al. 1985). An intact acrosome has been recognized at the apex of spermatozoa that are suspended in both normal and caffeinecontaining sea water (Figs 5, 6), and it binds to and passes through the chorion in both normal and caffeine-treated eggs (Figs 7, 8, 9, 10). Several processes that have been designated apical processes in the present paper have been found protruding from the apical region of the spermatozoa in the perivitelline space of both normally fertilized (Figs 11, 12) and caffeine-treated eggs (Figs 13, 14). These processes typically are about 100 nm in length and 40 nm in diameter (Figs 12, 14). It is also interesting that gamete fusion probably occurs between the egg plasmalemma and one or more of the apical processes of the spermatozoa (Figs 15, 16, 17, arrows). Fig. 1. Sagittal section through a differentiated spermatozoon. »i, mitochondrion; n, nucleus. X9500. Fig. 2. Sagittal section through the anterior region of a differentiated spermatozoon, a, acrosome; as, apical substance; fu, fuzzy material. X 114000. Fig. 3. Transverse section through the apex of a differentiated spermatozoon at the level of an acrosome. Arrow indicates a pedestal for the acrosome. a, acrosome;/«, fuzzy material; n, nucleus. XI14000. Fig. 4. Transverse section through the apex of a differentiated spermatozoon at the level of the apical substance. Arrow indicates a pedestal for the acrosome. as, apical substance;fu, fuzzy material; n, nucleus. X 114000. Fettilization in ascidians 191 Discussion Apical processes have been recognized at the apex of the spermatozoa in the perivitelline space of both normal and caffeine-treated eggs. Although the number of spermatozoa in the perivitelline space is limited in normally inseminated eggs, apical processes have been observed in almost all spermatozoa in appropriate sections. Furthermore, we have observed a fairly large number of spermatozoa in the perivitelline space of caffeine-treated eggs. In these cases, all spermatozoa in favourably oriented sections have apical processes. On ---2. r - : as 192 M. Fukumoto the other hand, these processes have never been observed at the apex of spermatozoa that have been suspended for 10 min in normal sea water (Fig. 5) or in caffeine-containing sea water (Fig. 6). Moreover, 100% of caffeine-treated eggs can be fertilized and develop normally if they are inseminated by 106 sperm ml" 1 , a concentration of spermatozoa that can fertilize 100 % of normal eggs. These facts suggest that the apical processes reported here have not been induced by caffeine and that caffeine does not have a harmful effect, at this concentration (5mM), on the process of fertilization and development in C. intestinalis. Because these processes are observed in many spermatozoa in both normal and caffeine-treated eggs, these structural changes most probably occur in C. intestinalis spermatozoa as one of the prerequisite steps during the process of normal fertilization. As the processes are believed to function in fusion with the egg plasmalemma, they might be analogous to the acrosomal processes of other marine invertebrates. A vesicle is found contiguous to the region where gamete fusion might occur (Figs 16, 17). At present, the significance of this vesicle is unknown. In spite of intensive observations of many spermatozoa in the perivitelline space in both normal and caffeine-treated eggs, an acrosome has not been found, except for a possible residue of an acrosome in a few cases (Fig. 13, arrow). Furthermore, the apical substance sometimes has a fibrillar appearance (Figs 10, 14), which suggests that a molecular reorganization, such as, for example, polymerization of actin, might have occurred. These facts suggest that the formation of the apical processes occurs as changes are taking place in the acrosome and apical substance. In this Fig. 5. Sagittal section through the apex of a spermatozoon suspended in normal sea water, a, acrosome; as, apical substance;/^, fuzzy material. X120000. Fig. 6. Sagittal section through the apex of a spermatozoon suspended in caffeine-containing sea water. a, acrosome; as, apical substance;fu, fuzzy material, x 120 000. Fig. 7. Sagittal section through the apex of a spermatozoon that is binding to the chorion of a normal egg (non-caffeine-treated egg), a, acrosome; as, apical substance; ch, chorion. X120000. Fig. 8. Sagittal section through the apex of a spermatozoon that is binding to the chorion of a caffeinetreated egg. a, acrosome; as, apical substance; ch, chorion. X120000. Fig. 9. Sagittal section through the apex of a spermatozoon that is penetrating the chorion of a normal egg (non-caffeine-treated egg), a, acrosome; as, apical substance; ch, chorion. X 120000. Fig. 10. Sagittal section through the apex of a spermatozoon that is penetrating the chorion of a caffeinetreated egg. a, acrosome; as, apical substance; ch, chorion. X120000. context it is of particular interest to note the finding that actin is probably present at the apex of the sperm head in Boltenia and Cnemidocarpa (Lambert & Lambert, 1984), although negative results have been obtained in Ascidia ceratodes (Lambert & Lambert, 1984) and in Phallusia mammillata (Honegger, 19866). Although we have no supporting evidence, it seems safe to assume that the factor(s) that elicit(s) the structural changes at the apex of the spermatozoa exist(s) in the perivitelline space of C. intestinalis. The claim that the acrosome reaction occurs at the surface of the chorion after the specific binding of spermatozoa has been made repeatedly for C. intestinalis (Rosati & DeSantis, 1978; DeSantis et al. 1980, 1983; Monroy & Rosati, 1983; Rosati et al. 1985). In spite of intensive observations on a large number of spermatozoa that are bound to the chorion, the acrosome reaction has not been observed, as has been reported (Fukumoto, 1984a). In Phallusia mammillata, Honegger (1986a,b) found several vesicles (up to eight) at the apex of the sperm head. He reported that some of these vesicles fuse with the sperm membrane and release their contents prior to the passage of the sperm through the chorion and that their remains can still be recognized in the spermatozoa in the perivitelline space. He has speculated that these vesicles contain chorion lysin(s) and /or additional enzymes that participate in the process of sperm-egg fusion. Honegger (1986a,b) has suggested that sperm-egg fusion in P. mammillata occurs between the plasma membrane of the post-acrosomal region of the sperm head and the egg membrane, as observed in mammalian fertilization. In contrast to the findings in P. mammillata, gamete fusion between the sperm and the egg plasma membranes in C. intestinalis most probably occurs at the anterior tip of the sperm head by means of the apical process(es) that protrude(s) from the apical region of the spermatozoa. In C. intestinalis, I have reported acrosome fragmentation, which has been referred to as an acrosome reaction. The acrosome transforms into smaller vesicles immediately after spawning or suspension in sea water (Fukumoto, 1984a). In the present study, however, such a change has not been recognized in acrosomes of spermatozoa that are suspended in normal or caffeine-containing sea water, or binding to and passing through the chorion. The reason for this discrepancy is not clear, but it is probable that I mistook the apical substance, which sometimes appears as small vesicles (Fig. 5) and figs 4, 6, 8, 9, 14 (Fukumoto, 1984a), for acrosome fragmentation because I overlooked the apical substance at that time. Further studies to clarify the precise reason for this discrepancy are in progress. In any case, the spermatozoa can pass through the chorion without releasing the contents of the acrosome and without any observable changes in the plasmalemma Fertilization in ascidians 193 Fig. 11. Transverse section, probably at the level of the apical substance, at the apex of a spermatozoon that is in the perivitelline space of a normal egg. as, apical substance", es, egg surface; n, nucleus; pr, processes. X 120000. Fig. 12. Longitudinal section through the apex of a spermatozoon that is in the perivitelline space of a normal egg. pr, processes. X 120000. Fig. 13. Sagittal section through the apex of a spermatozoon that is in the perivitelline space of a caffeine-treated egg. Arrow indicates probable remnant of the acrosome. ep, egg plasmalemma; pr, processes. X120000. Fig. 14. Sagittal section through the apex of a spermatozoon that is in the perivitelline space of a caffeine-treated egg. Electron-dense material is present at the apex, pr, processes. X 120 000. enclosing the apex of the sperm head (Fukumoto, 1984fl, and the present study). The findings in the present study are in good agreement with the hypotheses that ascidian spermatozoa contain a poorly developed acrosome that reacts at an appropriate step in fertilization and participates 194 M. Fukumoto mainly in the fusion of the gamete plasma membranes, and that the chorion lysin(s) are intercalated into the plasmalemma enclosing the sperm head (Fukumoto, 1983, 19846). The author is most grateful to Professor Gary Freeman and Dr Judy Lundelius, of the University of Texas at Austin, for V? 17 Fig. IS. Sagittal section through the head of a spermatozoon that is in contact with the egg surface in a caffeine-treated egg. es, egg surface. X 10 000. Fig. 16. Enlargement of the apical region of the sperm head shown in Fig. IS. Arrow indicates the region where the egg and the sperm plasma membranes are probably confluent, ep, egg plasmalemma; v, vesicle. X 120000. Fig. 17. Longitudinal section through the apex of a spermatozoon whose plasmalemma is probably confluent with the egg membrane at two places (arrows) in a caffeine-treated egg. ep, egg plasmalemma; v, vesicle. X120000. Fertilization in ascidians L95 their valuable suggestions and for reading the manuscript. He also thanks Miss Mieko Matsushima and Mr Toshihiro Nagao, medical students of Nagoya City University, for their assistance in preparing the manuscript. References CLONEY, R. A. & ABBOTT, L. C. (1980). The spermatozoa of ascidians: acrosome and nuclear envelope. Cell Tiss. Res. 206, 261-270. DESANTIS, R., JAMUNNO, G. & ROSATI, F. (1980). A study of the chorion and the follicle cells in relation to the sperm-egg interaction in the ascidian, Ciona intestinalis. Devi Biol. 74, 490-499. DESANTIS, R., PINTO, M. R., COTELLI, F., ROSATI, F., MONROY, A. & D'ALESSIO, G. (1983). A fucosyl glycoprotein component with sperm receptor and spermactivating activities from the vitelline coat of Ciona intestinalis eggs. Expl Cell Res. 148, 508-513. FUKE-TSUKAMOTO, M. (1983). Self and non-self recognition between gametes of the ascidian, Halocynthia roretzi. Roux's Arch. Devi Biol. 192, 347-352. FUKUMOTO, M. (1981). The spermatozoa and spermiogenesis of Perophora forniosana (Ascidia) with special reference to the striated apical structure and the filamentous structures in the mitochondrion. J. Ultrastruct. Res. 77, 37-53. FUKUMOTO, M. (1983). Fine structure and differentiation of the acrosome-like structure in the solitary ascidians, Pynra haustor and Styela plicata. Dev. Growth Differ. 25, 503-513. FUKUMOTO, M. (1984a). Fertilization in ascidians: acrosome fragmentation in Ciona intestinalis spermatozoa. J. Ultrastruct. Res. 87, 252-262. FUKUMOTO, M. (19846). The apical structure in Perophora annectens (Tunicata) spermatozoa: fine structure, differentiation and possible role in fertilization. J. Cell Sa. 66, 175-187. FUKUMOTO, M. (1985). Acrosome differentiation in Molgula manhattensis (Ascidiacea, Tunicata)..?. Ultrastruct. Res. 92, 158-166. FUKUMOTO, M. (1986). The acrosome in ascidians. 1. Pleurogona. Int. J. Invert. Reprod. Dev. 10, 335-346. HONEGGER, T. G. (1982). Effect on fertilization and localized binding of lectins in the ascidian Phallusia mammillata. Expl Cell Res. 138, 446-451. HONEGGER, T . G. (1986a). Ultrastructural and experimental investigations of sperm-egg interactions in the ascidian, Phallusia mammillata. In Progress in Developmental Biology, part B, pp. 85-89. New York: Alan R. Liss. HONEGGER, T. G. (19866). Fertilization in ascidians: studies on the egg envelope, sperm and gamete 196 M. Fukumoto interactions in Phallusia mammillata. Devi Biol. 118, 118-128. HOSHI, M., DESANTIS, R., PINTO, M. R., COTELLI, F. & ROSATI, F. (1985). Sperm glycosidases as mediators of sperm-egg binding in the ascidians. Zool. Sci. 2, 65-69. HOSHI, M., NUMAKUNAI, T . & SAWADA, H. (1981). Evidence for participation of sperm proteinases in fertilization of the solitary ascidian, Halocynthia roretzi: effects of protease inhibitors. Devi Biol. 86, 117-121. ISHIKAWA, M., NUMAKUNAI, T. & KUBO, M. (1978). Fertilization in ascidians: interaction between spermatozoa and eggs. In Abstr. Ilth Ann. Meet. Japanese Soc. Devi Biol. 74 (in Japanese). LAMBERT, C. C. (1986). Fertilization-induced modification of chorion Ar-acetylglucosamine groups blocks polyspermy in ascidian eggs. Devi Biol. 116, 168-173. LAMBERT, C. C. & LAMBERT, G. (1978). Tunicate eggs utilize ammonium ions for flotation. Science 200, 64—65. LAMBERT, C. C. & LAMBERT, G. (1984). The role of actin and myosin in ascidian sperm mitochondrial translocation. Devi Biol. 106, 307-314. MILLER, R. L. (1975). Chemotaxis of the spermatozoa of Ciona intestinalis. Nature, Land. 254, 244—245. MONROY, A. & ROSATI, F. (1983). Review article: A comparative analysis of sperm-egg interaction. Gamete Res. 7, 85-102. ROSATI, F. & DESANTIS, R. (1978). Studies on fertilization in the ascidians. I. Self-sterility and specific recognition between gametes of Ciona intestinalis. Expl Cell Res. 112, 111-119. ROSATI, F., PINTO, M. R. & CASAZZA, G. (1985). The acrosomal region of the spermatozoon of Ciona intestinalis: its relationship with the binding to the vitelline coat of the egg. Gamete Res. 11, 378-389. SAWADA, H., YOKOSAWA, H., HOSHI, M. & ISHII, S. (1982). Evidence for acrosin-like enzyme in sperm extract and its involvement in fertilization of the ascidian, Halocynthia roretzi. Gamete Res. 5, 291-301. SAWADA, H., YOKOSAWA, H. & ISHII, S. (1984a). Purification and characterization of two types of trypsinlike enzymes from sperm of the ascidian (Prochordata) Halocynthia roretzi. J. biol. Chem. 259, 2900-2904. SAWADA, H., YOKOSAWA, H., SOMENO, T., SAINO, T . & ISHII, S. (19846). Evidence for the participation of two sperm proteases, spermosin and acrosin, in fertilization of the ascidian, Halocynthia roretzi: inhibitory effects of leupeptin analogs on enzyme activities and fertilization. Devi Biol. 105, 246-249. WOOLLACOTT, R. M. (1977). Spermatozoa of Ciona intestinalis and analysis of ascidian fertilization.^. Morph. 152, 77-88. (Received 9 jfulv 1987 -Accepted, in revised form, 22 October 1987)