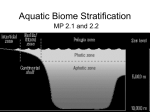
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
J. Great Lakes Res. 26(1):31–54 Internat. Assoc. Great Lakes Res., 2000 Benthic and Pelagic Secondary Production in Lake Erie after the Invasion of Dreissena spp. with Implications for Fish Production Ora E. Johannsson1,*, Ron Dermott1, Donna M. Graham1, Julie A. Dahl2, E. Scott Millard1, Debra D. Myles3, and Jennifer LeBlanc4 1Great Lakes Laboratory for Fisheries and Aquatic Sciences Canada Centre for Inland Waters 867 Lakeshore Rd. P.O. Box 5050 Burlington, Ontario L7R 4A6 2INAC-Environment and Conservation P.O. 2310 Yellow Knife, Northwest Territory X1A 2R3 3Dept. of Fisheries and Oceans Box 1500, 4914-50th St P.O. 2310 Yellowknife, Northwest Territory X1A 2P7 44072 Cid Way Pleasanton, California 94566 ABSTRACT. Benthic and pelagic secondary production were measured at nearshore and offshore sites in the western, west-central, and eastern basins of Lake Erie in 1993 to determine the relative importance of benthic and pelagic foodwebs to the fish community after dreissenid colonization. Benthic biomass increased greatly between 1979 and 1993 because of the presence of dreissenids, and > 90% of benthic production in 1993 came from dreissenids. Biomass of “other” benthos (excluding unionids and dreissenids) did not decline. Dreissenid production was in addition to, and not at the expense of, “other” benthic production. Zooplankton production was close to or within the 95% confidence interval of that predicted from primary production (photosynthesis) based on the relationship described by Makarewicz and Likens (1979). Deviations from this relationship were correlated with summer zooplankton mean length, suggesting that planktivory was an important factor in the regulation of zooplankton production in the lake. Dreissenids therefore impact zooplankton production by reducing algal biomass and primary production particularly in unstratified regions, by decreasing rotifer abundance and hence biomass and production, and by producing veligers which contribute 10% to 25% to zooplankton production. Potential fish biomass which could be supported by the benthic and pelagic foodwebs was estimated from empirical equations. The benthic food chain could potentially support more fish biomass than the pelagic food chain in all basins in 1993, even if dreissenids were excluded from the calculations. INDEX WORDS: Benthos, zooplankton, Dreissena, fish, production, planktivory. INTRODUCTION Zebra (Dreissena polymorpha) and quagga mussels (D. bugensis) are re-organizing community structure and the flow of energy through freshwater communities in North America (Leach 1993; Hol- land et al. 1995; Dahl et al. 1995; Dermott and Kerec 1997; Fahnenstiel et al. 1995a, b; Madenjian 1995; Nalepa and Fahnenstiel 1995; Graham et al. 1996). In so doing, they are changing the relative availability of pelagic and benthic food sources for the fish community. Dreissenids may affect zooplankton both directly through predation (Shevtsova et al. 1986 in *Corresponding author. E-mail: [email protected] 31 32 Johannsson et al. MacIsaac et al. 1995, MacIsaac et al. 1991) and indirectly through competition for food (Ten Winkel and Davids 1982, Cotner et al. 1995, Lavrentyev et al. 1995). Common food sources of dreissenids include rotifers, ciliates, small nauplii (also eaten by omnivorous copepods, particularly cyclopoids), large algae (also eaten by copepods, particularly calanoids), and to a lesser extent, small algae and bacteria (also eaten by cladocerans) (Ten Winkel and Davids 1982, Sprung and Rose 1988, Cotner et al. 1995, Lavrentyev et al. 1995, MacIsaac et al. 1995). In addition to physically consuming the same food as zooplankton, dreissenids may also reduce primary production below the potential set by the phosphorus concentration (Millard et al. 1999), and in some instances, they may decrease the pelagic phosphorus pool, at least temporarily (Mellina et al. 1995, Fahnenstiel et al. 1995b, Millard unpubl. data). In phosphorus-limited systems, such as Lake Erie, phosphorus levels set the level of primary production which in turn should set the maximum level of zooplankton production (Makarewicz and Likens 1979; Lean et al. 1983; McQueen et al. 1986; Millard et al. 1996a,b). While the impact of dreissenids on the pelagic community is predominantly negative, their effects on the benthic community are mixed. The benthic community benefits from increased water clarity, the extension of littoral habitat, and increased habitat structure in the presence of dreissenids (Dermott et al. 1993, Griffiths 1993, Stewart and Haynes 1994, Lowe and Pillsbury 1995, Skubinna et al. 1995). However, mussels are responsible for the decimation of unionid and larger sphaeriid clams (Mackie 1993, Gillis and Mackie 1994, Dahl et al. 1995) and are thought to be largely responsible for the disappearance of the benthic amphipod Diporeia in eastern Lake Erie (Dermott and Munawar 1993, Dermott and Kerec 1997). Lake Erie is a focal point for research on dreissenids and was chosen for this study because it was near the epicenter of the invasion, a range of trophic and substrate conditions exist within the lake, and most importantly, the potential impact of dreissenids on the fishery was of economic significance. The fishery in Lake Erie is one of the more profitable freshwater fisheries in the world (T. Cowan, DFO, pers. comm.) with a long-term commercial catch of 11 to 20 million kg/yr in Canadian waters (OMNR 1995) and with a valuable sports fishery for walleye and salmon in American waters. In the present study lower trophic level productivity in the presence of dreissenids in nearshore and offshore environments in Lake Erie was investigated. The production and community structure of benthos and zooplankton, and the relative importance of the benthic and pelagic food chains in supporting the fish community were examined. METHODS AND MATERIALS Description of Study Site Morphometrically, Lake Erie consists of three basins with distinct structural and trophic characteristics (Fig. 1) (Burns 1985). The western basin is shallow (mean depth = 7.4 m; Mortimer 1987), rarely stratifies, and is separated from the central basin by the Lorain Ridge southeast from Point Pelee. The offshore of the central basin is a plain 20 m to 24 m deep (mean basin depth = 18.3 m). The basin stratifies in summer and extensive areas of the hypolimnion on the western side of the plain go anoxic or experience low oxygen conditions by the end of the summer. The Pennsylvania Ridge, south of Long Point, divides the central and eastern basins. The deepest regions of the east basin are off Long Point (mean basin depth = 25.0 m, maximum depth = 64 m). Strong gradients in temperature and productivity exist along the east-west axis of the lake. In 1993, the mean May to October temperature of the mixed layer was 2°C to 3°C lower in the eastern and west-central basins than in the western basin, average total phosphorus ranged from 7 µg/L to 10 µg/L in the eastern basin to 17 µg/L in the western basin, and chlorophyll a increased from 1.06 µg/L to 2.24 µg/L in the eastern basin to 4.50 µg/L in the western basin (Dahl et al. 1995). Field Program Two stations in the western basin, two in the west-central basin, and three in the eastern basin were sampled as part of the Lake Erie Biomonitoring Program (LEB) (Fig. 1). Sites were chosen to include both nearshore and offshore regions, and in the eastern basin, areas which did and did not stratify during the summer (Table 1). Sampling extended from 3 May 1993 to 25 October 1993 in the western and west-central basins, and from 10 May to 18 October 1993 and 10 May to 8 November 1994 in the eastern basin. All parameters were sampled every 2 weeks except benthic samples were taken monthly from May to October 1993 and once in May 1994 from all sites, except E1 in the eastern basin. Epilimnetic, metalimnetic, and hypolimnetic or Benthic and Pelagic Production in Lake Erie: Post-Dreissena 33 FIG. 1. Morphometric map of Lake Erie indicating the sampling locations and station names. whole-water-column zooplankton samples were collected depending on the thermal stratification of the water column. The water column was considered stratified when the density gradient caused by the difference in temperature exceeded 0.08/m3/m at a depth > 4 m (Reynolds and Wiseman 1982, Reynolds et al. 1984). Zooplankton were collected with a 0.5-m diameter, 110-µm mesh Wisconsin closing net, 3-m in length, which was metered to allow correction for filtering efficiency. The net was raised at 0.8 to 1.0 m/s. A 2-m gap was left between samples from different strata to prevent overlap of the samples caused by movement of the boat Two replicate samples were collected, preserved in 4% phosphate-buffered, sugared formalin, and combined before sample analysis. In 1994 total water column samples were also collected at E2 to assess the ability of the stratified sampling method to provide seasonally reliable biomass estimates. The seasonally-weighted (weighted for intersampling interval) mean biomass of zooplankton in the total water column during the stratified period was 26.9 µg/L as estimated from the stratified samples and 26.6 µg/L as estimated from the total water column samples. To sample rotifers, integrated, whole-water samples were collected over the same depth ranges as the zooplankton net hauls using a Shurflo diaphragm pump. Ten liters of this water was passed through a 20-µm sieve and preserved with 4% phosphate-buffered, sugared formalin. In 1994, a 20 L sample was taken from the epilimnion where ro- tifers were most abundant, otherwise 50 L were filtered. The rotifers were anaesthetized for 2 minutes in soda water before being preserved. Five replicate petit-Ponar samples (0.0232 m 2 each) were taken every month for benthos. Each sample was sieved through a 0.25-mm screen and preserved in 10% neutral formalin. Additional information on the other parameters measured and results of the Lake Erie Biomonitoring Program are given in Dahl et al. (1995) and Graham et al. (1996). Sample Analysis A minimum of 400 individual zooplankton were enumerated and measured using a stratified, random, counting procedure (Cooley et al. 1986) ensuring that at least 100 individuals of each major group were included. If animal density was low, 20% of the sample was counted. De Milo (1993) found that the genus Bosmina in the Great Lakes consists of two new species (B. liederi and B. freyi) and not B. longirostris. The two species are difficult to separate; consequently, Bosmina were identified only to genus. Lengths of cladocerans were measured from the top of the helmet to the base of the tail spine, copepods from the anterior end of the cephalothorax to the end of the caudal rami, and veligers across the widest section of the shell. Body mass was estimated from length-weight regressions from the literature: the equations for the dominant groups were verified with field data (Appendix 1a). 9.6 Fine silt Station Depth (m) Substrate Type Fine silt 10.4 none W3 1993 May 6– Oct. 27 Coarse sand 16.3 Jul. 15– Aug. 17 WC1 1993 May 7– Oct. 26 Temperature of SWM* 18.9 19.6 16.7 Mixed Depth Max. 25.5 24.6 23.3 (oC) Min 10.5 10.1 6.6 *SWM is the seasonally weighted mean over the period sampled. none Period of Stratification Sampling Date Range W1 1993 May 7– Oct. 26 16.9 23.5 6.8 Fine silt 21.6 Jun. 30– Sept. 17 WC2 1993 May 7– Oct. 26 17.5 23.7 10.6 Fine sand 5.9 none E1 1993 May 14– Oct. 20 16.6 22.5 8.0 5.9 none E1 1994 May 10– Oct. 18 16.8 23.1 7.0 Fine silt 9.2 none E3 1993 May 12– Oct. 20 Station and water quality parameters for the LEB sites sampled in 1993 and 1994. Parameter/Station TABLE 1. 16.4 22.1 5.2 9.2 none E3 1994 May 10– Oct. 18 17.0 23.3 4.3 Fine silt 38.0 Jun. 24– Sept. 24 E2 1993 May 14– Oct. 5 16.2 23.0 2.9 38.0 Jun. 16– Oct. 18 E2 1994 May 10– Oct. 18 34 Johannsson et al. Benthic and Pelagic Production in Lake Erie: Post-Dreissena A minimum of 400 rotifers were enumerated in a Sedgewick-Rafter chamber. Body mass was estimated according to the formulae of Ruttner-Kolisko (in Bottrel et al. 1976). Polyarthra biomass was estimated from a simplified formula, however, the literature equation was not applicable for Lake Erie rotifers. A common equation was developed for P. remata, P. vulgaris, and P. major: v = 0.158a3 (95% confidence interval: 0.153 to 0.163) where v = volume in µ3 (or wet body mass in µg × 10–6 assuming a density of one) and a = longest dimension in microns. The equation for P. dolichoptera was v = 0.205a3 (95% confidence interval: 0.195 to 0.215). Three or four of the preserved benthic samples were analyzed from each station on each sample date, while the remaining samples were archived. All organisms retained on a 580-µm screen (#30 mesh) were enumerated. The finer material retained on a 250-µm screen was subsampled by volume, and a minimum of 1/4 of this material was sorted under a microscope at a magnification of 20X. The counts were converted to number per sample. Invertebrates were identified and their blotted wet weights were measured. The organisms in each taxonomic group were placed into distilled water for exactly 60 s before weighing to ensure wet weights were consistent. Common taxa were identified to genus or species where possible, but only to family level for the Nematoda, Oligochaeta, Ostracoda, and Harpacticoida. The Chironomus species, Procladius species, Gammarus fasciatus, and the zebra (Dreissena polymorpha) and quagga (D. bugensis) mussels were divided into size classes and enumerated. Average density and wet biomass (g/m2) of each taxon were calculated per m2 for each station. Wet weight of the mollusks was converted to shell free wet weight using calculated ratios of soft tissue to total wet weight: D. polymorpha 0.56, D. bugensis 0.58, Sphaeriidae 0.3. Production Calculations All zooplankton production was calculated on a dry-weight basis unless otherwise indicated. All benthic production was calculated on a shell-free, wet-weight basis. All comparisons between the production of the benthos and plankton were done on a wet-weight basis assuming zooplankton dry body mass was 10% of wet weight. This is consistent with the associated literature. Sufficient error is associated with the numerous steps in production (sampling, enumeration, length-weight regressions, Production/Biomass approximations), that small 35 differences in production should not be considered significant. Zooplankton Seasonal (1 May to 31 October) zooplankton production was calculated for the total water column at each station. Production estimates calculated using the egg-ratio method are based on changes in abundance and egg ratios between dates and on mean temperature. Production estimates calculated from P/B ratios depend on seasonally-weighted mean body size, seasonally-weighted mean population biomass, and median temperature. Therefore, production was calculated for each species within individual thermal layers over the temporal duration of that layer. For example, in 1993 production in the metalimnion of station E2 was calculated for the stratified period: 18 June to 29 September. In the offshore of the eastern basin, the data from the preand post-stratified periods were combined with the hypolimnetic data from the stratified period to create a single continuous season because water temperatures were similar (> 4 and < 10°C) and warm water species were not abundant until after stratification. Similarly, in the west-central basin, the preand post-stratified data were combined with the epilimnetic data because the systems became unstratified while temperatures were still high (17 and 23°C). For each station, the species productions for each thermal layer were summed to generate seasonal water column production. The egg-ratio method of Paloheimo (1974), as described in Cooley et al. (1986), was used to calculate production of cladocerans, cyclopoids, and calanoids if the seasonally-weighted, mean biomass of the species was > 50 mg/m 2 /season and the species carried its eggs. Development coefficients are listed in Appendix 1b. When these conditions were not met, production was estimated using P/B relationships. P/B estimates for non-predatory cladocerans were taken from Stockwell and Johannsson (1997): when temperature was > 10°C, P/B = 0.162/d and when it was < 10°C, P/B = 0.042/d. For the larger, predatory cladocerans, Bythotrephes cederströemi and Leptodora kindtii, the size-dependent equations of Stockwell and Johannsson (1997) were used to estimate daily P/B: mean seasonal temperature > 10°C log (daily P/B) = –0.23 * log (dry wt (µg)) –0.73 (1) 36 Johannsson et al. mean seasonal temperature < 10°C log (daily P/B) = –0.26 * log (dry wt(µg)) –1.36 (2) The P/B estimates for cyclopoid, calanoid and rotifer production were taken from the seasonal daily P/B equations of Shuter and Ing (1997): log(median daily P/B) = A + 0.04336 * (median temperature (°C)) (3) where A = –1.844 (cyclopoids) = –2.294 (calanoids) = –1.631 (rotifers) Comparisons between production estimates using the P/B equations of Stockwell and Johannsson (1997) and Shuter and Ing (1997) and egg-ratio production estimates from a long-term monitoring site in Lake Ontario (Johannsson, unpubl. data), indicated that the equations of Stockwell and Johannsson (1997) came closer to predicting egg-ratio estimates of seasonal zooplankton production for cladocerans while the seasonal daily P/B equations of Shuter and Ing (1997) were better for cyclopoid species. No calanoid species were present in Lake Ontario in sufficient biomass to calculate production using the egg-ratio method, therefore comparisons were not possible for this group. Dreissenid veligers grow from approximately 60 to 250 µm in length, or from 0.033 to 1.711 µg dry weight over a 44 to 70 day period (Leach 1993, Doka 1994), which equates to a daily P/B of 0.160 to 0.101/d. A daily P/B of 0.1/d was used to be conservative and to allow for slower growth during the colder portions of the season. This rate was applied to veligers in the epilimnion. A rate of 0.04/d was applied when the median temperatures was < 10°C. The daily P/B of a species in a specific thermal layer was multiplied by the seasonally-weighted mean biomass of that species and by the length of the season of that thermal layer in days in order to calculate seasonal production. Egg-ratio and P/B estimates of production for individual species were compared, where possible, in order to test the validity of using the above P/B values to calculate the production of the less abundant species. Where the average egg-ratio estimates were more than 25% different from the P/B estimates, the P/B estimates of all similar species were adjusted by the average percent difference. Benthos Annual production of the common benthic species: Dreissena spp., Chironomus spp., Procladius spp., and Gammarus fasciatus was calculated using the size-frequency method of Krueger and Martin (1980). These calculations used monthly mean densities in four size classes, average wet weight (shell-free) per individual in each size class, and the geometric weight between neighboring size classes j and j+1. A cohort generation time of 1 year was used at all sites except E2. Data at E2 suggested that in deep water, growth of D. bugensis was half that at the other stations, so a generation time of 2 years was used at E2. Where numbers of the common species were insufficient to calculate size frequencies, and for all other benthic taxa, production was calculated from the mean annual wet biomass and published values for the turnover (P/B) ratios for each taxon (Appendix 2). RESULTS Dreissena spp. represented between 90% and 99% of the benthic biomass at all stations except WC2 in the west-central basin, where the biomass was dominated by Chironomus spp. (Table 2). As a result, the biomass and production of the benthic fauna in Lake Erie were dominated by Dreissena spp., which contributed over 90% of the benthic production at 5 of the 6 stations (Table 3). Dreissenid veliger production was not as significant in the pelagic system as adult dreissenid production was in the benthic system. In most instances, it was responsible for 10% to 25% of pelagic production. No clear spatial pattern was evident except for the low production at WC2 (Table 4). Zooplankton Spatial gradients of zooplankton production within the lake were dissimilar among the three subgroups of zooplankton: macrozooplankton, rotifers, and veligers (Table 4). Macrozooplankton production was higher in the west and west-central basins than in the eastern basin. Rotifer production was greatest in the west-central basin and offshore of the eastern basin, and least in the nearshore of the eastern basin. Veliger production was lowest in the nearshore of the eastern basin and in the offshore of the west-central basin. Production may best be understood if examined on a basin basis. In the eastern basin, total water column, May to October zooplankton production (TZP) was much Benthic and Pelagic Production in Lake Erie: Post-Dreissena 37 TABLE 2. Annual benthic biomass at selected Lake Erie stations, 1993. Biomass is wet, shell-free weight (g/m2). Averages were calculated from 7 sampling dates, with the exception of Station W3 (n = 5) and E2 (n = 6). W1 Species Biomass Nematoda 0.064 Platyhelminthes/ Nemertea 0.001 Oligochaeta 4.002 Hirudinae 0.094 Amphipoda 0.050 Isopoda 0.000 Ostracoda 0.286 Harpacticoida 0.222 Ephemeroptera 0.009 Trichoptera 0.031 Chironomidae 1.630 Gastropoda 0.014 Sphaeriidae 0.188 Dreissena polymorpha 64.800 Dreissena bugensis 0.000 W3 S.E. Biomass WC1 S.E. Biomass WC2 S.E Biomass E2 E3 S.E. Biomass S.E. Biomass S.E. 0.030 0.081 0.018 0.022 0.010 0.129 0.028 0.155 0.020 0.171 0.030 0.001 0.989 0.033 0.025 0.000 0.046 0.031 0.006 0.031 0.781 0.009 0.046 0.187 4.068 0.752 0.925 0.000 0.076 0.109 0.001 0.038 0.299 0.655 0.148 0.104 0.831 0.153 0.585 0.000 0.018 0.019 0.001 0.021 0.081 0.273 0.057 0.050 1.649 0.010 0.112 0.066 0.105 0.005 0.000 0.000 1.445 0.291 0.008 0.031 0.426 0.007 0.106 0.059 0.053 0.002 0.000 0.000 0.827 0.125 0.005 0.075 6.647 0.022 0.002 0.000 0.129 0.070 0.000 0.000 12.311 0.000 1.206 0.055 1.185 0.022 0.002 0.000 0.022 0.013 0.000 0.000 1.583 0.000 0.127 0.074 12.593 0.004 0.000 0.000 0.143 0.021 0.000 0.000 0.114 0.001 0.091 0.029 1.555 0.004 0.000 0.000 0.021 0.006 0.000 0.000 0.054 0.000 0.048 0.392 3.906 0.090 3.573 0.030 0.055 0.177 0.001 1.669 2.653 1.623 0.039 0.072 0.440 0.064 0.926 0.030 0.016 0.047 0.001 0.553 0.479 0.139 0.011 64.416 624.100 249.577 347.633 161.073 0.263 0.218 0.552 0.366 115.382 52.370 0.001 416.490 171.374 5.147 3.875 264.005 81.157 246.287 63.893 0.000 0.001 Total Dreissena % Quagga 64.800 0.0 65.599 624.101 254.033 764.120 244.812 — 0.0 — 54.5 — 5.410 95.1 3.995 — 264.557 99.8 79.321 361.669 111.460 — 68.1 — Total Benthos % Dreissena 71.393 90.8 66.669 631.440 250.533 767.884 247.250 8.2 98.8 6.9 99.5 9.4 26.002 20.8 5.630 5.9 277.753 95.2 83.414 376.050 113.823 0.9 96.2 1.6 lower in the nearshore (stations E1, E3) than in the offshore (E2) (Table 4). Offshore epilimnetic production during the stratified period exceeded nearshore production for the season. Macrozooplankton production in the nearshore ranged from 0.72 g dry wt/m 2 to 1.72 g dry wt/m2. In order of decreasing contribution, the majority (> 85%) of this production came from Epischura lacustris, Leptodiaptomus minutus, Skistodiaptomus oregonensis, Tropocyclops extensus, and Bosmina spp., in 1993, and from E. lacustris, Diacyclops thomasi, S. oregonensis, Daphnia retrocurva, Bosmina spp. and L. minutus in 1994. Offshore TZP was 20.03 g dry wt/m2 in 1993 and 10.75 g dry wt/m2 in 1994. Over 85% of macrozooplankton production came from E. lacustris, Bosmina spp., T. extensus, D. thomasi, Mesocyclops edax, S. oregonensis, Eurytemora affinis, and Bythotrephes cederstroemi in 1993, and from D. thomasi, D. retrocurva, Bosmina spp., E. lacustris, Leptodiaptomus ashlandi, and B. cederstroemi in 1994. Total zooplankton production in 1993 was elevated compared with that in 1994 pre- dominantly because of the high level of veliger production, 9.80 g dry wt/m2 compared with 1.69 g dry wt/m2. The high densities of veligers in the metaand hypolimnion and the large size of these individuals compared with those in the epilimnion suggests that the pediveliger stage may have postponed settlement (Peterson 1984, Boudreau et al. 1993, Sprung 1993). Veligers in the hypo- and metalimnion were 64 µm and 27 µm longer, respectively, than veligers in the epilimnion in 1993, while the length difference was much less in 1994 - 9 µm. In normal circumstances when the pediveligers settle out more rapidly, their production would be associated with the benthic food chain. If the relative production of veligers in the three thermal layers were similar in 1993 and 1994, then total veliger production at E2 would drop from 9.80 g dry wt/m 2 to 2.48 g dry/m2 in 1993 and TZP would drop to 12.98 g dry wt/m2. Rotifer production contributed a significant proportion to total zooplankton production in the eastern basin and tended to be higher in 1994 than in 1993 except in the meta- and hypolimnion 38 Johannsson et al. TABLE 3. Annual benthic production estimates for different taxonomic groups at the sampled Lake Erie stations, 1993. Production is given in wet, shell-free weight (g/m2/yr). Species Nematoda Platyhelminthes/Nemertea Oligochaeta Hirudinae Amphipoda Isopoda Ostracoda Harpacticoida Ephemeroptera Trichoptera Chironomidae Gastropoda Sphaeriidae Dreissena polymorpha Dreissena bugensis Total Dreissena Total Production % Dreissena W1 0.91 0.01 18.70 0.32 0.10 0.00 1.16 1.12 0.02 0.16 7.07 0.02 0.51 448.50 0.00 448.50 478.60 93.71 W3 1.14 1.34 18.99 2.25 4.87 0.00 0.31 0.55 0.00 0.19 1.11 1.18 0.32 3,010.67 0.01 3,010.68 3,042.93 98.98 of the offshore station. Inshore, rotifers contributed 30% (1993) and 40% to 60% (1994) toward TZP. Offshore, the contribution was approximately 35% in both years; however, rotifer production was more important in the epilimnion in 1994 (52%) than in 1993 (36%). In the west-central basin, total water column production was greater at the nearshore station (WC1) than in the offshore (WC2): 18.71 g dry wt/m2 as compared with 13.92 g dry wt/m2 (Table 4). This difference was mainly because of higher levels of production of Bosmina spp. and veligers in the nearshore. Bosmina spp. production was 6.08 g dry wt/m2 at WC1 and 2.35 g dry wt/m2 at WC2, while veliger production was 4.13 g dry wt/m2 at WC1 and 0.24 g dry wt/m2 at WC2. Together Bosmina spp., B. cederstroemi, D. thomasi, M. edax, L. minutus, Eubosmina spp., and calanoid and cyclopoid copepodids contributed > 85% of macrozooplankton production at WC1, and Bosmina spp., D. thomasi, B. cederstroemi, M. edax, Leptodiaptomus sicilis, E. lacustris, Leptodiaptomus siciloides, T. extensus, D. retrocurva, and Eubosmina spp. contributed > 85% at WC2. Rotifers contributed 23% toward TZP at WC1, and 46% at WC2. The majority of rotifer production occurred in the epilimnion where they were the principal grazers at WC2 contributing 66% of TZP. WC1 0.32 0.30 7.93 0.03 0.18 0.21 0.47 0.03 0.00 0.00 7.44 0.48 0.02 746.64 1,161.54 1,908.18 1,925.59 99.10 WC2 1.86 0.61 32.66 0.07 0.01 0.00 0.54 0.37 0.00 0.00 24.58 0.00 3.04 1.33 32.74 34.07 97.81 34.83 E2 2.19 0.40 53.81 0.01 0.00 0.00 0.64 0.11 0.00 0.00 0.39 0.00 0.18 1.79 532.55 534.34 592.07 90.25 E3 2.42 2.60 15.61 0.25 20.46 0.10 0.27 0.89 0.00 8.42 17.34 5.25 0.07 492.35 2,484.85 2,977.20 3,050.88 97.58 Total zooplankton production at the two sites in the western basin were markedly different. At the northern nearshore site (W1), TZP was 12.09 g dry wt/m2, while in the center of the basin at site W3 it was 23.77 g dry wt/m2 (Table 4). The difference was principally a consequence of the much greater productivity of Daphnia retrocurva in the offshore: 15.42 g dry wt/m 2 as compared with 2.40 g dry wt/m2. Daphnia were 12% more abundant and 13% longer at W3 than W1 in 1993. This translated into a 66% increase in biomass and 543% increase in production. The marked increase in production was a result of an exponential increase in the number of eggs per female with increasing body size. At W3, Daphnia alone contributed 88% of macrozooplankton production. At W1, D. retrocurva, E. affinis, Bosmina spp., Diaphanosoma spp., D. thomasi, and E. lacustris together contributed just > 85% to total macrozooplankton production. Veliger production was similar at the two stations, approximately 2.50 g dry wt/m2. Rotifer production was slightly higher at W3 ( 3.73 g dry wt/m2) than at W1 (2.22 g dry wt/m2). At both sites, rotifers contributed approximately 17% toward TZP. The empirical relationship between primary production (PP), measured as photosynthesis, and zooplankton production, developed by Makarewicz and Likens (1979), can be used to evaluate whether ob- Benthic and Pelagic Production in Lake Erie: Post-Dreissena 39 TABLE 4. Seasonal zooplankton production at the LEB sites by thermal layer (non-bolded numbers) and for the whole water column (bolded numbers). Species/Stations Cladocerans Bosmina spp. Eubosmina spp. Daphnia retrocurva D. galeata mendotae D. longiremis Daphnia spp. Ceriodaphnia spp. Holopedium gibberum Diaphanosoma sp. Bythotrephes Leptodora kindtii Total Cladocerans W1-t 0.95 0.05 2.40 0.00 0.15 0.00 0.00 0.00 0.71 0.00 0.03 4.31 Zooplankton Production (g/m2/May-Oct. season) W3-t WC1-te WC1-m WC1-h 0.13 3.22 1.52 1.34 0.29 0.31 0.03 0.01 15.42 0.02 0.00 0.00 0.00 0.00 0.00 0.00 0.19 0.02 0.02 0.01 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.10 0.01 0.06 0.03 0.00 0.10 0.52 0.01 0.32 0.00 0.02 0.00 16.45 3.69 2.17 1.41 WC1-t 6.08 0.36 0.02 0.00 0.05 0.00 0.00 0.00 0.10 0.62 0.02 7.26 Cyclopoids Diacyclops thomasi Tropocyclops extensus Mesocyclops edax Cyclops vernalis Cyclopoid copepodids Cyclopoid nauplii Total Cyclopoids 0.32 0.01 0.11 0.05 0.09 0.04 0.62 0.51 0.00 0.06 0.01 0.05 0.01 0.65 0.08 0.18 0.21 0.02 0.12 0.02 0.63 0.16 0.02 0.11 0.02 0.10 0.02 0.41 0.35 0.00 0.09 0.05 0.06 0.00 0.56 0.59 0.20 0.41 0.10 0.28 0.04 1.61 Calanoids Leptodiaptomus minutus L. ashlandi Skistodiaptomus oregonensis L. sicilis L. siciloides Limnocalanus macrurus Eurytemora affiinis Senecella calanoides Epischura lacustris Epischura copepodids Calanoid copepodids Calanoid nauplii Total Calanoids Total Macrozooplankton 0.04 0.17 0.00 0.10 0.02 0.03 1.84 0.04 0.10 0.12 0.04 0.02 2.51 7.44 0.03 0.05 0.01 0.03 0.00 0.06 0.04 0.00 0.04 0.00 0.12 0.02 0.40 17.50 0.02 0.01 0.02 0.01 0.01 0.00 0.04 0.00 0.10 0.02 0.10 0.01 0.35 4.66 0.13 0.05 0.06 0.01 0.01 0.00 0.08 0.00 0.04 0.02 0.05 0.01 0.46 3.04 0.23 0.06 0.03 0.05 0.02 0.00 0.02 0.00 0.02 0.00 0.12 0.00 0.56 2.53 0.38 0.13 0.11 0.07 0.05 0.00 0.13 0.00 0.16 0.04 0.28 0.02 1.36 10.23 Dreissenids Veliger 2.43 2.55 2.58 1.46 0.09 4.13 3.90 11.14 0.38 4.87 0.08 2.70 4.35 18.71 Rotifers Rotifers 2.22 3.73 Total Production 12.09 23.77 t = total water column, e = epilimnion, m = metalimnion, h = hypolimnion (Continued next page) served zooplankton production achieved potential levels set by PP in Lake Erie (Fig. 2). Zooplankton production in Makarewicz and Likens’ tabulation included both rotifers, collected with at least a 45-µm mesh net, and crustacean zooplankton. A similar comparison in the present study would include rotifers, veligers, and macrozooplankton (TZP). Their PP included all sources of PP including benthic and macrophyte. Only pelagic algal production was measured in the present study which may underestimate PP at the shallower stations (E1, E3, W1, and W3). Seasonal TZP was predicted for each station from seasonal PP estimated during the present study (Millard et al. 1999). Observed TZP 40 TABLE 4. Johannsson et al. (Continued). Species/Stations Cladocerans Bosmina spp. Eubosmina spp. Daphnia retrocurva D. galeata mendotae D. longiremis Daphnia spp. Ceriodaphnia spp. Holopedium gibberum Diaphanosoma sp. Bythotrephes Leptodora kindtii Total Cladocerans Zooplankton Production (g/m2/May-Oct. season) WC2-te WC2-m WC2-h WC2-t 1.18 0.10 1.07 2.35 0.22 0.01 0.01 0.24 0.12 0.02 0.11 0.25 0.00 0.00 0.00 0.00 0.00 0.02 0.18 0.20 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.02 0.00 0.02 0.00 0.00 0.02 0.82 0.01 0.02 0.00 0.03 1.88 0.63 1.38 3.90 Cyclopoids Diacyclops thomasi Tropocyclops extensus Mesocyclops edax Cyclops vernalis Cyclopoid copepodids Cyclopoid nauplii Total Cyclopoids 0.13 0.25 0.17 0.01 0.03 0.01 0.61 0.09 0.01 0.11 0.01 0.04 0.02 0.26 0.68 0.00 0.29 0.01 0.02 0.01 1.00 0.90 0.26 0.56 0.02 0.09 0.04 1.87 Calanoids Leptodiaptomus minutus L. ashlandi Skistodiaptomus oregonensis L. sicilis L. siciloides Limnocalanus macrurus Eurytemora affinis Senecella calanoides Epischura lacustris Epischura copepodids Calanoid copepodids Calanoid nauplii Total Calanoids Total Macrozooplankton 0.02 0.01 0.00 0.09 0.00 0.00 0.02 0.00 0.06 0.03 0.02 0.01 0.28 2.76 0.03 0.03 0.07 0.09 0.01 0.00 0.01 0.00 0.25 0.02 0.03 0.02 0.55 1.45 0.03 0.06 0.06 0.20 0.32 0.00 0.00 0.00 0.03 0.00 0.01 0.00 0.70 3.09 0.08 0.09 0.15 0.38 0.33 0.00 0.03 0.00 0.34 0.05 0.05 0.03 1.53 7.30 Dreissenids Veliger 0.16 0.06 0.02 0.24 0.35 1.85 0.35 3.45 6.38 13.92 Rotifers Rotifers 5.69 Total Production 8.62 t = total water column, e = epilimnion, m = metalimnion, h= hypolimnion (Continued next page) from the LEB sites all fell within or close to the 95% confidence interval surrounding the relationship developed by Makarewicz and Likens (Fig. 2). Zooplankton mean length (ZML) was calculated for the summer season, 27 June to 30 September in the eastern and west-central basins, and 15 May to 30 September in the western basin, as an index of planktivory. Shorter ZMLs are thought to be associated with more intense planktivory (Brooks and Dodson 1965, Lynch 1979, Mills et al. 1987). Mills et al. (1987) suggested that a ZML of 0.8 mm, as determined with a 153-µm mesh net, was characteristic of a well balanced fish community. Zooplankton mean lengths were converted to 153-µm Benthic and Pelagic Production in Lake Erie: Post-Dreissena TABLE 4. 41 (Continued). Species/Stations Cladocerans Bosmina spp. Eubosmina spp. Daphnia retrocurva D. galeata mendotae D. longiremis Daphnia spp. Ceriodaphnia spp. Holopedium gibberum Diaphanosoma sp. Bythotrephes Leptodora kindtii Total Cladocerans Zooplankton Production (g/m2/May-Oct. season) E1-t93 E3-t93 E2-e93 E2-m93 E2-th93 0.08 0.15 0.33 0.10 0.56 0.00 0.00 0.00 0.00 0.00 0.01 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 0.00 0.00 0.00 0.00 0.00 0.10 0.05 0.01 0.01 0.01 0.10 0.03 0.00 0.12 0.22 0.54 0.20 0.57 E2-t93 0.99 0.01 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.16 0.00 1.18 Cyclopoids Diacyclops thomasi Tropocyclops extensus Mesocyclops edax Cyclops vernalis Cyclopoid copepodids Cyclopoid nauplii Total Cyclopoids 0.01 0.07 0.00 0.00 0.01 0.00 0.09 0.04 0.18 0.05 0.00 0.00 0.00 0.28 0.02 0.70 0.02 0.00 0.01 0.00 0.74 0.09 0.01 0.11 0.00 0.01 0.00 0.21 0.37 0.08 0.14 0.01 0.01 0.00 0.60 0.47 0.78 0.26 0.01 0.02 0.00 1.55 Calanoids Leptodiaptomus minutus L. ashlandi Skistodiaptomus oregonensis L. sicilis L. siciloides Limnocalanus macrurus Eurytemora affinis Senecella calanoides Epischura lacustris Epischura copepodids Calanoid copepodids Calanoid nauplii Total Calanoids Total Macrozooplankton 0.41 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.10 0.11 0.01 0.00 0.64 0.86 0.54 0.00 0.19 0.02 0.00 0.00 0.02 0.00 0.30 0.06 0.07 0.02 1.23 1.72 0.35 0.05 0.00 0.01 0.03 0.00 0.01 0.00 0.19 0.11 0.02 0.02 0.79 2.06 0.20 0.04 0.12 0.01 0.00 0.00 0.07 0.00 1.05 0.00 0.03 0.00 1.52 1.93 0.08 0.06 0.09 0.02 0.01 0.00 0.09 0.00 0.06 0.00 0.00 0.00 0.41 1.58 0.63 0.15 0.21 0.04 0.04 0.00 0.16 0.00 1.30 0.11 0.06 0.03 2.72 5.45 2.20 4.26 3.35 9.80 2.38 6.64 1.99 8.18 4.16 0.68 5.60 2.31 5.05 20.30 13.98 Dreissenids Veliger 0.48 1.00 modified as per 1994(see text) Rotifers Rotifers 0.53 1.29 Total Production 1.86 4.01 Total Production (modified veligers) t = total water column, e = epilimnion, m = metalimnion, h = hypolimnion (Continued next page) equivalent ZMLs and ranged from 0.49 mm at WC2 to 0.67 mm at W3 (Table 5). The lack of larger species of Daphnia, normally D. galeata mendotae in the Great Lakes, was also indicative of intense fish predation (Wells 1970, Johannsson et al. 1991). In 1993, D. g. mendotae was observed briefly at only one site (E1). In 1994, D. g. mendotae was again observed briefly at E1 and was less abundant than D. retrocurva at E2. Regression analysis indicated that the percent deviation of observed TZP from predicted TZP [(predicted TZP—observed TZP)/predictedTZP*100] was negatively related to summer ZML (p < 0.001, r 2 = 0.88), as were the percent deviations of the 42 TABLE 4. Johannsson et al. (Continued). Species/Stations Cladocerans Bosmina spp. Eubosmina spp. Daphnia retrocurva D. galeata mendotae D. longiremis Daphnia spp. Ceriodaphnia spp. Holopedium gibberum Diaphanosoma sp. Bythotrephes Leptodora kindtii Total Cladocerans Zooplankton Production (g/m2/May-Oct. season) E1-t94 E3-t94 E2-e94 E2-m94 E2-th94 0.04 0.05 0.53 0.14 0.07 0.00 0.00 0.01 0.00 0.00 0.02 0.08 0.08 0.52 0.43 0.00 0.00 0.02 0.02 0.01 0.00 0.00 0.01 0.01 0.01 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 0.02 0.08 0.02 0.01 0.02 0.01 0.02 0.00 0.00 0.10 0.17 0.74 0.71 0.53 E2-t94 0.74 0.02 1.03 0.05 0.03 0.00 0.00 0.00 0.00 0.10 0.00 1.96 Cyclopoids Diacyclops thomasi Tropocyclops extensus Mesocyclops edax Cyclops vernalis Cyclopoid copepodids Cyclopoid nauplii Total Cyclopoids 0.24 0.00 0.00 0.00 0.01 0.00 0.26 0.33 0.01 0.01 0.00 0.02 0.00 0.37 0.16 0.02 0.02 0.00 0.04 0.01 0.25 1.44 0.00 0.05 0.00 0.02 0.00 1.51 0.28 0.00 0.02 0.00 0.01 0.00 0.32 1.88 0.03 0.08 0.00 0.07 0.01 2.08 Calanoids Leptodiaptomus minutus L. ashlandi Skistodiaptomus oregonensis L. sicilis L. siciloides Limnocalanus macrurus Eurytemora affinis Senecella calanoides Epischura lacustris Epischura copepodids Calanoid copepodids Calanoid nauplii Total Calanoids Total Macrozooplankton 0.04 0.00 0.09 0.00 0.00 0.00 0.01 0.00 0.07 0.02 0.10 0.03 0.36 0.72 0.06 0.01 0.04 0.01 0.00 0.00 0.00 0.00 0.14 0.06 0.06 0.04 0.42 0.96 0.12 0.06 0.05 0.02 0.00 0.01 0.01 0.00 0.25 0.11 0.05 0.02 0.70 1.69 0.16 0.08 0.03 0.02 0.00 0.00 0.00 0.00 0.23 0.01 0.02 0.00 0.55 2.77 0.01 0.01 0.01 0.01 0.00 0.00 0.00 0.00 0.03 0.00 0.01 0.00 0.09 0.93 0.29 0.15 0.08 0.05 0.00 0.01 0.01 0.00 0.51 0.12 0.07 0.02 1.33 5.37 Dreissenids Veliger 0.44 0.37 1.47 0.15 0.06 1.70 3.34 6.51 0.27 3.20 0.08 1.07 3.69 10.75 Rotifers Rotifers 0.79 2.18 Total Production 1.95 3.51 T = total water column, e = epilimnion, m = metalimnion, h = hypolimnion macrozooplankton + veliger production (p = 0.001, r2 = 0.85), and macrozooplankton alone (p = 0.003, r2 = 0.78): one outlier (WC1) was removed from these analyses (Fig. 3). The relationships were still significant if WC1 were included but the explained variances were lower (r2 = 0.45, 0.43, and 0.47 respectively). The slopes of the three relationships were not significantly different (ANCOVA p = 0.33): the adjusted means were significantly differ- ent from one another (p < 0.001). Neither variation in dreissenid seasonally-weighted mean biomasses in 1993 nor dreissenid clearance rates in May of 1993 and 1994 could explain the percent deviation in observed versus predicted zooplankton production. The estimated clearance rates were taken from Table 17 of Graham et al. (1996). Thus zooplankton production in Lake Erie appears to be controlled by a combination of the level Benthic and Pelagic Production in Lake Erie: Post-Dreissena FIG. 2. Relationship between seasonal zooplankton production (TZP) and primary production (PP): the predictive relationship and 95% confidence intervals are presented for the equation developed by Makarewicz and Likens (1979) where log(TZP mg C/m2) = 3.032 + 0.0063 (PP g C/m2), and (●) data from Lake Erie in 1993 and 1994. Carbon data were converted to dry weight assuming a 50% C composition. *Data for E2 1993 where the veliger production was modified (see text for explanation). of PP during the growing season (May to October) and the intensity of predation. These factors could be expressed jointly in multiple regression relationships where zooplankton production is expressed in g dry wt/m2/season, ZML is in mm, and PP is in g C/m2/season: MP = macrozooplankton production, and MVP = macrozooplankton + veliger production: TZP = –36.5 + 62.7 * ZML + 0.111 * PP F = 51.1, p < 0.001, n = 8, adjusted R2 = 0.94 (4) MVP = –34.0 + 57.7 * ZML + 0.083 * PP F = 16.4, p = 0.006, n = 8, adjusted R2 = 0.82 (5) MP = –28.1 + 46.2 * ZML + 0.076 * PP F = 11.8, p = 0.013, n = 8, adjusted R2 = 0.76 (6) The statistical procedure identified WC1 as an outlier and it was removed from these relationships. Tolerance levels were greater than 0.95 indicating no correlation among the independent variables. Log transforming the zooplankton production esti- 43 FIG. 3. Percent deviation of the Lake Erie total zooplankton production estimates from the Makarewicz and Liken (1979) relationship (see Fig. 2) with respect to the summer zooplankton mean length. See text for definition of summer period. Open symbols are station WC1. TABLE 5. Whole-water column, summer zooplankton mean length (ZML) as determined by a 153-um net. The 1993 data were taken from Figure 4 of Johannsson et al. (1999). The 1994 and E1 1993 data were converted from 64-µm to 153µm equivalent data using the equation of Johannsson et al. (1999): ZML153 = 0.042 + 1.330 ZML64 . The season was determined by temperature. June 27 to Sept. 30 in the eastern and westcentral basins, May 15 to Sept. 30 in the western basin. Station/Year E1 E3 E2 WC1 WC2 W1 W3 Mean Body Length (mm) 1993 1994 0.56 0.54 0.54 0.52 0.64 0.61 0.52 0.49 0.53 0.67 mates did not improve the relationships. The veliger distribution and size structure at E2 in1993 were unusual; therefore, the adjusted values described above were used for all the percent deviation and regression analyses. Benthos Benthic production was higher at the nearshore stations of the central and eastern basins than at the 44 Johannsson et al. offshore stations (Table 3). The eastern and western basins had similar ranges of benthic production. The zebra mussel, D. polymorpha, contributed most of the production in the western basin, D. bugensis contributed the majority of the benthic production at the other stations. The calculated P/B ratios of D. polymorpha averaged 4.4 (SE = 0.7) while those for D. bugensis averaged 5.3 (SE = 1.8), and were greatest at the nearshore stations, and lowest at the deep offshore station of the eastern basin (P/B = 2.0). After the mussels, oligochaetes contributed the most to benthic production in the lake. Annual production by the oligochaetes ranged between 8 and 54 g/m2/y (Table 3), with their greatest production at the two offshore hypolimnetic stations, WC2 and E2. Chironomids, mostly Chironomus thummi, were important contributors to the benthic production at the inshore stations and less important offshore in the eastern basin, but total chironomid production was greatest in the offshore of the west-central basin. Benthic production was much lower (97.8 g/m2/y) at the offshore west-central station (WC2) than at the other stations, due to the low density of Dreissena at that station. The benthic community at this station was comprised of oligochaete worms, chironomids, Pisidium, and a few Sphaerium clams, typical of the hypolimnetic benthic community of Lake Erie prior to the arrival of Dreissena (Dermott 1994). The low proportion of Dreissena and relatively high biomass of Sphaeriidae (1.2 g/m2, wet shell-free) and Chironomidae at station WC2 probably reflects the low mid-summer oxygen conditions in the hypolimnion of the central basin. The oligochaetes, Chironomus spp., and several of the Pisidium species are adapted to survive in water that experiences short periods of low oxygen (Jonasson 1972). At this station, calculated production by the Tubificidae was 32.6 g/m2/y, size-frequency production by the Chironomus semireductus group was 17.1 g/m2/y (P/B = 1.7) and that by C. thummi group was 7.2 g/m2/y (P/B = 2.9). Together, production by the Chironomidae represented 25% of the total benthic production at this hypolimnetic station. Amphipods were only common at the nearshore stations. No specimens of the deep-water amphipod Diporeia hoyi were found in any of the samples collected for this study. Gammarus fasciatus was present at all the nearshore stations, its biomass ranged from 0.05 g/m2 in the west basin to was 0.11 g/m2 at WC1 in the central basin. The biomass (3.6 g/m2) and production (20.5 g/m2/y) of this species were greatest at the nearshore station E3 in the eastern basin, where a large population of Gammarus was associated with the Dreissena colonies (Table 2). However, the high production of amphipods at E3 represented only 0.7% of the total annual production at E3 because of the high biomass of Dreissena spp. at that station. Gammarus also contributed a minor amount to the production at the nearshore, west-central basin station. The calculated P/B ratios of Gammarus averaged 3.9 (SE = 0.9) in Lake Erie, ranging between 1.1 and 5.7. Sphaeriidae clams contributed little to the benthic production of Lake Erie, except at station WC2. Although numerous, the small Nematoda contributed a minor share of the production in the eastern basin (2.2 to 2.4 g/m2/y). The contributions of the other members of the benthic community were generally less than 1 g/m2/y. Both abundance and biomass of the benthic fauna were dominated by the vast number of settled Dreissena spp. (Dahl et al. 1995). In 1993, the quagga mussel (D. bugensis) was not present in the western basin but represented 95% to 99% of the mussels present at the two stations > 20 m, WC2 and E2. During 1993, D. bugensis became increasingly more common in the central and eastern basins. Dreissena bugensis was able to colonize WC2 during the summer of 1993 when the hypolimnion of the central basin did not go anoxic (M. Charlton, pers. com.). Biomass of D. bugensis at station WC2 increased steadily from 0.05 g/m2/ (wet shell-free) in May, 1993 to 28.12 g/m2 in May 1994, as the settled mussels grew over the year. Only one cohort existed at this site, individuals grew from an average of 0.0002 g/ind on 20 May 1993 to 0.2917 g/ind (wet shell-free) by 20 May 1994. DISCUSSION Benthic filtering bivalves, such as Dreissena, can sequester much of the pelagic primary production (Officer et al. 1982, Stanczykowska and Lewandowski 1993), thereby altering the balance between pelagic and benthic foodwebs. The biomass and production of the benthic and pelagic fish communities will shift to reflect these changes. Impacts of dreissenids were observed on both the pelagic and benthic foodwebs in Lake Erie. The resultant impact on potential fish biomass and production is discussed in the concluding section. Conditions that favor bivalve dominance include Benthic and Pelagic Production in Lake Erie: Post-Dreissena a high proportion of suitable substrate, lack of stratification of the overlying water, and good water flow. Within Lake Erie, these areas can be found year round along the north and south shores of the eastern basin and among the rocky islands of the western basin. In the offshore of the central and eastern basins conditions are favorable during isothermal periods. Areas less favorable for dreissenids include those regions which stratify in the summer separating the mussels from the productive surface layers, and the western portion of the central basin which has periodic hypolimnetic anoxia or low oxygen levels in the late summer (Bertram 1993, Charlton et al. 1993). Thus the impacts of dreissenids would be expected to vary both spatially and temporally within the lake. Zooplankton Production No estimates of zooplankton production were made in Lake Erie immediately prior to the invasion of dreissenids. Therefore their impact can not be assessed directly. The energy pathways leading to zooplankton production consists of several links which could be altered by dreissenids. They could decrease the levels of chlorophyll a and photosynthesis (Leach 1993; Fahnenstiel et al. 1995a, b; Millard et al. 1999), alter the level of detrital material and bacteria in the water column, consume small zooplankton (Shevtsova et al. 1986 in MacIsaac et al. 1995, MacIsaac et al. 1991), and compete with zooplankton for the available primary production. Chlorophyll a concentrations are below those predicted from total phosphorus concentrations and Daphnia composition in the nearshore of the eastern basin (Graham et al. 1996). Millard et al. (1999) also documented a decrease in seasonal primary production in this region compared with that predicted from total phosphorus. The declines in chlorophyll a, and consequently primary production, were attributed to dreissenids. Zooplankton production is partially dependent on the level of primary production (Makarewicz and Likens 1979, present study). Therefore, to the extent that dreissenids have depressed primary production, they have also depressed potential zooplankton production. The second link between observed primary production and total zooplankton production was not affected by the presence of adult dreissenids; however, 88% of the variability in the LEB TZP estimates from the Makarewicz and Likens relationship could be accounted for by zooplankton mean length 45 (ZML). Planktivory may play a strong role in regulating zooplankton production in Lake Erie as TZP was reduced by an average of 36% (range 15% to 50%, n = 5) at sites with ZMLs of 0.49 to 0.56, excluding WC1. This modification of bottom-up control of zooplankton production by top-down influences agrees with the top-down bottom-up theory of trophic regulation (McQueen et al. 1986, 1989). An impact of direct competition for algal food resources could not be demonstrated, although zooplankton and dreissenids must both be consuming the phytoplankton produced in the unstratified regions throughout the season. This competition is reduced as zooplankton feed throughout the water column, while dreissenids feed predominantly from the bottom waters (Fréchette et al. 1989, MacIsaac et al. 1999). Dreissenid impact on chlorophyll a and competition with zooplankton increases as the water column shortens and as the level of turbulence increases. Total zooplankton production now consists of veliger larvae as well as the traditional crustacean and rotifer species. Veligers can frequently contribute 10% to 25% to total zooplankton production. If veligers were excluded from the calculation of TZP, TZP would fall below the 95% confidence interval for the Makarewicz and Likens relationship at stations E1, E3, and W1. Veligers may be replacing some of the traditional zooplankton production. If the potential capacity for zooplankton production is set from the bottom up and modified by planktivory, then the potential production can be estimated knowing the relationship between total phosphorus and PP (Millard et al. 1999) and the relationship between PP, ZML, and TZP developed above (equation 4). The Lake Erie data set is restricted in the size range of zooplankton communities encountered. A ZML of 0.67 mm (observed at W3) was assumed as the optimum size, instead of the 0.8 mm suggested by Mills et al. (1987) in order to stay within the bounds of the regression equations. Comparing observed TZP with potential predicted TZP suggested that TZP was particularly depressed in the nearshore of the eastern basin (Table 6). TZP at sites E1 and E3 was 9% to 23% of capacity. The middle of the western basin (W3) and the nearshore of the west-central basin (WC1) were operating at 85% of capacity, and the remaining sites at 45% to 65% of capacity. The decline in observed TZP can be divided roughly into that attributable to predation and that to decreased PP, keeping in mind that there is error 46 Johannsson et al. TABLE 6. Calculation of potential productive capacity of zooplankton at the LEB sites assuming a summer zooplankton mean length of 0.67 mm. Primary production (PP (gC/m2/season)) was predicted from May to October seasonally-weighted mean total phosphorus (TP (ug/L)) using the equation of Millard et al. (1999) (PP = 390.5TP/(18.45+TP)). Potential total zooplankton production (Pot-TZP (g dwt./m2/season)) was estimated from equation (4). Similarly, Pot-TZP* was estimated using the observed ZMLs. Observed production is expressed as a percentage of potential production. Pot-TZP* ZML = Observed TP PP Pot-TZP (ZML = 0.67) TZP Observed Percent Obs/Pot 7.8 8.5 6.6 10.8 12.8 17.5 19.1 116 123 103 144 160 190 199 18.83 19.62 17.37 21.95 23.70 27.05 27.99 1.86 12.98 4.01 18.71 13.91 12.09 23.77 10 66 23 85 59 45 85 11.66 17.66 8.90 12.18 11.98 17.93 27.99 16 73 45 154 116 67 85 1994 E1* 10.1 E2 8.1 E3* 8.5 * summer ZML ≤ 0.56 138 119 123 21.28 19.17 19.62 1.95 10.71 3.51 9 56 18 12.82 15.26 9.85 15 70 36 Station/Year 1993 E1* E2 E3* WC1* WC2* W1* W3 associated with the predictive regression relationships, particularly the latter which is based on a limited number of points. The difference between the potential TZP calculated at optimal ZML and that calculated at observed ZMLs is a measure of the impact of planktivory. The difference between the potential TZP calculated at observed ZMLs and the observed TLP is a measure of changes in PP. The depression in zooplankton production in the nearshore region of the eastern basin was principally because of the impact of dreissenid filtering on chlorophyll a levels and consequently PP (Millard et al. 1999, Table 6). After removing the effect of changes in PP, TZP was depressed by 40% to 50% at sites with ZMLs < 0.56. A similar analysis could be done using macrozooplankton + veliger production or macrozooplankton production using equations (5) and (6). The slopes of equations (4), (5) and (6) were not significantly different, thus the results of such an analysis would be similar. Benthic Production Western Basin Production was almost exclusively dominated by Dreissena which accounted for over 95% of the annual production and wet shell-free biomass in the basin during 1993. Lack of benthic production esti- Percent Obs/Pot* mates for Lake Erie prior to the colonization of the lake by Dreissena prevent direct comparison of production. Thus benthic biomass was used as a surrogate for production, to compare pre- and postmussel periods. Annual wet benthic biomass in the western basin averaged 351.4 g/m2 (shell-free) during 1993. Average biomass in the western basin during autumn 1979 (without Unionidae), 9 years prior to the invasion by the zebra mussels, was 6.7 (S.E. 1.47; N = 52) g/m 2 (Dermott 1994). This value was comparable to the benthic biomass (7.0 g/m2, excluding the mussels) in the west basin during 1993, after the invasion of Lake Erie by D. polymorpha. This similarity suggests that the huge biomass in the mussel populations was not at the expense of the biomass of the remaining benthic community. In the nearshore areas of Lake Erie, the populations of Gammarus fasciatus increased in association with the newly established mussel colonies. The mussel colonies provide interstitial spaces and a probable increased food supply in the form of pseudofaeces (Dermott et al. 1993, Stewart and Haynes 1994). Biomass of Gammarus in the western basin increased from an average of 0.002 (S.E. 0.0008 g/m2) at 52 sites sampled during 1979 (Dermott 1994) to 0.487 g/m2 (wet weight) at the two stations sampled during 1993. Benthic and Pelagic Production in Lake Erie: Post-Dreissena West-Central Basin Dreissena have altered production of the benthic community more at the shallower station (WC1) where they accounted for > 99% of the total production, than at the deeper station (WC2) where they accounted for only 27% of production. This is consistent with the low mussel density at the deeper station. The average wet benthic biomass in the central basin during 1979 was 10.6 (S.E. 1.43; N = 69) g/m2 (Dermott 1994), compared with 12.2 g/m2 (without Dreissena) in 1993. Again, this indicates that there has been little change in biomass of endemic benthic fauna, but Dreissena have increased the benthic biomass of the central basin sites to 396 g/m2 (shell-free). Eastern Basin In 1993, total abundance of the benthic fauna was greatest in the eastern basin, because of the very high density of recently settled quagga mussels (D. bugensis) of < 1 mm length (Dahl et al. 1995). As in the western basin, benthic production and biomass were dominated by Dreissena and the huge amount of biomass in the mussel population was in addition to the pre-mussel biomass of the benthic community. In 1979, average wet biomass in the eastern basin was 7.6 (S.E. 1.03; N = 38) g/m 2, compared to 13.8 g/m 2 (without Dreissena) in 1993. The total benthic biomass including Dreissena in 1993 was 326.9 g/m2. Thus, energy transferred to the mussels was presumably removed from the pelagic foodweb or derived from more efficient use of organic material which was settling to the bottom. Although the biomass of the benthic community had not declined, community composition in the eastern basin had been altered. Historically, the burrowing amphipod, Diporeia, was common in the profundal zone (> 30 m) in the eastern basin, contributing up to 20% of the biomass (Barton 1988, Flint and Merckel 1978, Dermott 1994). The wet biomass of this species averaged over all the eastern basin during 1979 was 1.7 (S.E. 0.5; n = 38) g/m2. No specimens of Diporeia were collected in this study at the deep 38 m station (E2) between May 1993 and May 1994. Populations of this amphipod have disappeared, possibly because of competition for settling algae, in particular the large diatoms with the large populations of quagga mussels (D. bugensis) now present in Lake Erie at depths beyond 30 m (Dermott and Munawar 1993, Dermott and Kerec 1997). In all the other Great Lakes, Diporeia accounts for the majority of the 47 benthic production, and is a key component of the foodweb (Hurley 1986, McDonald et al. 1990). The energy to support the high biomass and production of Dreissena in Lake Erie appears not to be at the expense of the benthic community, except for the physical competition with the native Unionidae and Sphaeriidae clams (Ricciardi et al. 1996). Rather the energy is being diverted from the microbial or pelagic foodwebs. It might be expected that the uncoupling of the benthic-pelagic link to fish production through the deep-water amphipod Diporeia in the hypolimnion of the eastern basin may be causing a reduction in condition and commercial catch of pelagic rainbow smelt (Osmerus mordax, Witzel et al. 1994). However, the increase in deepwater Dreissena may favor benthic feeding fish species such as whitefish (Coregonus clupeaformis), a native coregonid, and freshwater drum (Aplodinotus grunniens) (French and Bur 1993). Comparison of Benthic and Pelagic Production Does the dominance of pelagic production over non-dreissenid benthic production in 1993 (Table 7) mean that fish production in Lake Erie was more dependent on the pelagic foodweb? That will depend on the relative mean size of the plankton and benthos and on the amount of dreissenid production which went into higher trophic levels. Borgmann (1982) argued that an ecosystem has a characteristic efficiency of biomass or energy transfer from smaller to larger organisms (particle-size efficiency hypothesis), and that the amount of biomass or energy passed to predators by prey depends on the relative size of the predator and its prey. In this way, a fish would obtain more energy from a larger organism, like an amphipod, than from a smaller organism, like a cladoceran. Thus, pelagic and benthic biomass are not equivalent in supporting fish production. Boudreau and Dickie (1992) developed an empirical relationship relating the biomass (expressed as kcal/m 2 ) and mean mass (again expressed in kcal) of organisms in an ecosystem from plankton to fish using mean mass and total biomass of phytoplankton, zooplankton + benthos, and fish. log (Biomass) = a – 0.05 * log (Mean Individual Mass) (7) If the biomass and mean individual mass of one group of organisms are known, then the intercept “a” can be calculated for that system and the bio- 48 Johannsson et al. TABLE 7. Comparison of benthic and pelagic production at the LEB sites in Lake Erie: 1993. Dreissenids 96.7 75.2 79.7 Percent Contribution Other Benthos 2.4 8.2 8.7 Zooplanktona 0.8 16.6 11.6 92.3 19.7 0.8 36.9 6.9 43.5 West W1 557 W3 3,252 a Zooplankton includes macrozooplankton and veligers. bE2 with adjusted veliger production (see text). 77.7 92.8 5.2 1.0 17.1 6.2 mass of a different group of organisms of a known mean mass calculated. The intercept “a” was determined for each site for 1993 from the observed zooplankton plus benthic data, and then for the zooplankton and benthos separately. These intercepts were inserted into the equation to determined the capacity of zooplankton and benthos both separately and together to support a fish community with a mean individual mass of 460 g wet wt (600 kcal). Fish biomass was converted to an estimate of annual production using the following equation from Boudreau and Dickie (1989): fish. The estimates of fish production derived from these equations are considered potential production because they were estimated from relationships developed from traditional ecosystems: the Lake Erie ecosystem has been disturbed recently by the invasion of dreissenids and Bythotrephes and by other exotics such as smelt and alewife during this century, and may not be functioning efficiently. Potential fish production (PFP) was determined from the combined benthos + zooplankton biomass and mean mass estimates. Boundreau and Dickie (1989) noted that the contribution of the benthic and zooplankton components varied between ecosystems. The PFP derived from the benthic and pelagic food chains were used separately to suggest their potential relative importance in supporting fish production in Lake Erie. The sum of these separate estimates was 13.5% (± 6.6% S.D.) greater than the estimate derived from benthos + zooplankton. It was found that the benthic food chain should support 75% to 95% of the PFP in Lake Erie in all the regions studied except in the nearshore of the west-central basin where the contributions from the two food chains were essentially equal (Table 8). Thus the lake, as represented by the sites studied, was dominated by the benthic foodweb irrespective of any consumption of dreissenids by higher trophic levels. Basin East Station E3 E2 E2-altb Total Production (g wet wt/m2) 3,119 710 670 West-Central WC1 WC2 2,068 173 log (Production) = 0.08 –0.32 * log (Mean Individual Mass) + 1.01 * log (Biomass) (8) where the units are the same as above. The chosen mean mass of the fish (460 g wet wt) will not influence the relative contribution from pelagic and benthic sources. The equations of Boudreau and Dickie (1989, 1992) were developed for normally functioning ecosystems and will overestimate the potential of a disturbed portion of the foodweb to support fish production. Therefore, dreissenids were excluded from the benthic estimates as it was likely that they were at least a partial energy sink. Rotifers and meiofauna were not included in these calculations. Veligers are closer to the size of small copepods and cladocerans, and therefore were included. Dry weight and wet weight were converted to kcal using the following equalities (McLaren et al. (1989) and Banse and Mosher (1980) in Boudreau and Dickie (1992)): 1 g dry wt = 6 kcal and 1 g wet weight = 1 kcal for invertebrates and 1 g wet wt = 1.3 kcal for ACKNOWLEDGMENTS Without the support and help of a number of people this project would never have been accomplished. We thank John Cooley and Vic Cairns for initiating the DFO study in Lake Erie in response to Benthic and Pelagic Production in Lake Erie: Post-Dreissena 49 TABLE 8. Potential fish production (PFP g wet wt/m2/y) in Lake Erie derived from benthic + zooplankton biomass and mean mass assuming a normally functioning ecosystem and a mean fish biomass of 460 g wet wt. Percentage of PFP derived from the pelagic and benthic food webs (May 1993 to May 1994) derived separately according to the relationships of Boudreau and Dickie (1989, 1992). See text for more detailed desription. Dreissenids were excluded. Data rounded to the nearest 5%. 1993 - Offshore Percentage of PFP Production Pelagic Benthic 20 80 Basin West PFP 1.2 West-Central 3.1 15 85 1.0 50 50 East 2.3 25 75 2.0 5 95 the dreissenid invasion. Boat time and sampling support were generously given by the Ontario Ministry of Natural Resources and Coast Guard Canada. We thank Cheryl Hopcroft for the excellent work she did enumerating and measuring the zooplankton, Beak Consultants for the benthic enumeration, and Susan Doka and Mimi Gemininc for help in analyzing the benthic data. The graph of Lake Erie is from the hands (computer) of Carolynn Bakelaar. A special thanks goes to Don Noakes who gave me a place at the DFO laboratory in Nanaimo to work on these data and to Collin Wallace who shared his office with me during that period. REFERENCES Banse, K., and Mosher, S. 1980. Adult body mass and annual production/biomass relationships of field populations. Ecol. Monogr. 50:355–379. Barton, D.R. 1988. Benthic fauna from Great Lakes Institute cruises on Lake Erie—1963, 1964, 1965. Can. Tech. Rep. Fish. Aquat. Sci. 1635. Bertram, P.E. 1993. Total phosphorus and dissolved oxygen trends in the central basin of Lake Erie, 1970–1991. J. Great Lakes Res. 19(2):224–236. Borgmann, U. 1982. Particle-size-conversion efficiency and total animals production in pelagic ecosystems. Can. J. Fish. Aquat. Sci. 39:668–674. Bottrel, H.H., Duncan, A., Gliwicz, Z.M., Grygierek, E., Herzig, A., Hillbricht-Ilkowska, A., Kurasawa, H., Larsson, P., and Weglenska, T. 1976. A review of some problems in zooplankton production studies. Norw. J. Zool. 24:419–456. Boudreau, B., Bourget, E., and Simard, Y. 1993. Effect of age, injury and predator odors on settlement and PFP 1.0 1993 - Nearshore Percentage of PFP Production Pelagic Benthic 20 80 shelter selection by lobster Homarus americanus postlarvae. Mar. Ecol. Prog. Ser. 93:119–129. Boudreau, P.R., and Dickie, L.M. 1989. Biological model of fisheries production based on physiological and ecological scalings of body size. Can. J. Fish. Aquat. Sci. 46:614–623. ——— , and Dickie, L.M. 1992. Biomass spectra for aquatic ecosystems in relation to fisheries yield. Can. J. Fish. Aquat. Sci. 49:1528–1538. Brooks, J.l., and Dodson, S.I. 1965. Predation, body size and composition of plankton. Sci.150:28–35. Burns, N.M. 1985. Erie, the Lake that Survived. Rowman and Allanheld Publishers, U.S.A. Charlton, M.N., Milne, J.E., Booth, W.G., and Chiocchio, F. 1993. Lake Erie offshore in 1990: restoration and resilience in the central basin. J. Great Lakes Res. 19(2):291–309. Cooley, J.M., and Minns, C.K. 1978. Prediction of egg development times of freshwater copepods. J. Fish. Res. Board Can. 35:1322–1329. ———, Moore, J.E., and Geiling, W.T. 1986. Population dynamics, biomass, and production of the macrozooplankton in the Bay of Quinte during changes in phosphorus loadings. In Project Quinte: point source phosphorus control and ecosystem response in the Bay of Quinte, Lake Ontario, ed. C.K. Minns, D.A. Hurley and K.H. Nicholls, pp. 167–176. Can. Spec. Publ. Fish. Aquat. Sci. 86. Cotner, J.B, Gardner, W.S., Johnson, J.R., Sada, R.H., Cavaletto, J.F., and Heath, R.T. 1995. Effects of the zebra mussel, Dreissena polymorpha, on bacterioplankton: evidence for both size-selective consumption and growth stimulation. J. Great Lakes Res. 21: 517–528. Culver, D.A., Boucherle, M.M., Bean, D.J., and Fletche, J.W. 1985. Biomass of freshwater crustacean zoo- 50 Johannsson et al. plankton from length-weight regressions. Can. J. Fish. Aquat. Sci. 42:1380–1390. Dahl, J.A , Graham, D.M., Dermott, R., Johannsson, O.E., Millard, E.S., and Myles., D.D. 1995. Lake Erie 1993, western, west-central and eastern basins: Changes in trophic status, and assessment of the abundance, biomass and production of the lower trophic levels. Can. Tech. Rep. Fish. Aquat. Sci. No. 2070. De Milo, R. 1993. Speciation in the freshwater zooplankton: Provincialism in the Bosminidae. MSc thesis, University of Guelph. Dermott, R. 1994. Benthic invertebrate fauna of Lake Erie 1979: Distribution, abundance and biomass. Can. Tech. Rep. Fish. Aquat. Sci. 2018. ———. 1995. Production and growth efficiency of two burrowing invertebrates, Hexagenia limbata and Diporeia hoyi, in Batchawana Bay, Lake Superior. Can. Tech. Report of Fisheries & Aquatic Sciences 2034. ———, and Kerec, D. 1997. Changes to the deep-water benthos of eastern Lake Erie since the invasion of Dreissena: 1979–1993. Can. J. Fish. Aquat. Sci. 54: 922–930. ———, and Munawar, M. 1993. Invasion of Lake Erie offshore sediment by Dreissena, and its ecological implications. Can. J. Fish. Aquat. Sci. 50:2298–2304. ———, Kalff, J., Leggett, W.C., and Spence, J. 1977. Production of Chironomus, Procladius, and Chaoborus at different levels of phytoplankton biomass in Lake Memphremagog, Quebec-Vermont. J. Fish. Res. Board Can. 34:2001–2007. ———, Mitchell, J., Murray, I., and Fear, E. 1993. Biomass and production of zebra mussels (Dreissena polymorpha) in shallow waters of northeastern Lake Erie. p. 399–413. In Zebra Mussels: biology impacts, and control, ed. T. Nalepa and D. Schloesser. Boca Raton, Florida: Lewis Publishers. Doka, S.E. 1994. Spatio-temporal Dynamics and Environmental Prediction of Recruitment in Dreissena Populations of Lake Erie. M.Sc. thesis. University of Guelph, Guelph, Ont. Dumont, H.J., Van de Velds, I., and Dumont, S. 1975. The dry weight estimate of biomass in a selection of Cladocera, Copepoda and Rotifera from the plankton, periphyton and benthos of continental waters. Oecolog. 19:75–97. Edmondson, W.T., and Winberg, G.G. 1969. IBP # 17 A manual on methods for the assessment of secondary productivity in fresh waters. Oxford: Blackwell Scientific Publications. Fahnenstiel, G.L., Bridgeman, T.B., Lang, G.L., McCormick, M.J., and Nalepa, T.F. 1995a. Phytoplankton productivity in Saginaw Bay, Lake Huron: effects of zebra mussel (Dreissena polymorpha). J. Great Lakes Res. 21:465–475. ———, Lang, G.L., Nalepa. T.F., and Johengen, T.H. 1995b. Effects of zebra mussel (Dreissena polymorpha) colonization on water quality parameters in Saginaw Bay, Lake Huron. J. Great Lakes Res. 21: 435–448. Flint, R.W., and Merckel, C.N. 1978. Distribution of benthic macroinvertebrate communities in Lake Erie’s Eastern Basin. Verh. Internat. Verein. Limnol. 20: 240–251. Fréchette, M., Butman, C.A., and Geyer, W.R. 1989. The importance of boundary-layer flows in supplying phytoplankton to the benthic suspension feeder, Mytilus edulis L. Limnol. Oceanogr. 34:19–36. French, J.R.P.III, and Bur, M.T. 1993. Predation of the zebra mussel (Dreissena polymorpha) by freshwater drum in western Lake Erie. In Zebra Mussels: biology, impacts and control, ed. T. Nalepa and D. Scholesser, pp. 453–464. Boca Raton, Florida: Lewis Publishers. Gillis, P.L., and Mackie, G.L. 1994. Impact of zebra mussel, Dreissena polymorpha, on populations of Unionidae (Bivalvia) in Lake St. Clair. Can. J. Zool. 72:1260–1271. Graham, D.M., Dahl, J.A., Millard, E.S., Johannsson, O.E., and White, L.L. 1996. Assessment of abundance, biomass and production of the lower trophic levels in the eastern basin of Lake Erie, 1994. Can. Tech. Rept. Fish. Aquat. Sci. No. 2110. Griffiths, R.W. 1993. Effect of zebra mussels (Dreissena polymorpha) on benthic fauna of Lake St. Clair. In Zebra mussels: biology, impacts and control, ed. T. Nalepa and D. Scholesser, pp. 415–438. Boca Raton, Florida: Lewis Publishers. Hillbricht-Ilkowska, A., and Stanczykowska, A. 1969. The production and standing crop of planktonic larvae of Dreissena polymorpha in two Mazurian lakes. Pol. Arch. Hydrobiol. 16(29):193–203. Holland, R.E., Johengen, T.H., and Beeton, A.M. 1995. Trends in nutrient concentrations in Hatchery Bay, Western Lake Erie before and after Dreissena polymorpha. Can. J. Fish. Aquat. Sci. 52:1202–1209. Hurley, D.A., 1986. Effect of nutrient reduction on the diets of four fish species in the Bay of Quinte. pp 237–246. In Project Quinte: point-source phosphorus control and ecosystem response in the Bay of Quinte, Lake Ontario, ed. Minns, Hurley & Nicholls, pp. 237–246. Can. Spec. Publ. Fish. Aquatic Sci., 86. Johannsson, O.E., Mills, E.L., and O’Gorman, R. 1991. Changes in the nearshore and offshore zooplankton communities in Lake Ontario. Can. J. Fish. Aquat. Sci. 48:1546–1557. ———, Dumitru, C., and Graham, D.M. 1999. Estimation of zooplankton mean length for use in an index of fish community structure and its application in Lake Erie. J. Great Lakes Res. 25:179–186. Johnson, M.G., and Brinkhurst, R.O. Production of benthic macroinvertebrates of Bay of Quinte and Lake Ontario. J. Fish. Res. Board Can. 28:1699–1714. Benthic and Pelagic Production in Lake Erie: Post-Dreissena Jonasson, P.M. 1972. Ecology and production of the profundal benthos in relation to phytoplankton in Lake Esrom. Oikos Suppl. 14:1–148. Krueger, C.C., and Martin, F.B. 1980. Computation of confidence intervals for the size frequency (Hynes) method of estimating secondary production. Limnol. Oceangr. 25:773–777. ———, and Waters, T.F. 1983. Annual production of macroinvertebrates in three streams of different water quality. Ecology 64:840–850. Lavrentyev, P.J., Gardner, W.S., Cavaletto, J.F., and Beaver, J.R. 1995. Effects of the zebra mussel (Dreissena polymorpha Pallas) on protozoa and phytoplankton from Saginaw Bay, Lake Huron. J. Great Lakes Res. 21:545–557. Leach, J.H. 1993. Impacts of the zebra mussel, (Dreissena polymorpha) on water quality and spawning reefs in western Lake Erie. In Zebra mussels: biology, impacts and control, ed. T. Nalepa and D. Scholesser, pp. 381–397. Boca Raton, Florida: Lewis Publishers. Lean, D.R.S., Abbott, A.P., Charlton, M.N., and Rao, S.S. 1983. Seasonal phosphate demand for Lake Erie plankton. J. Great Lakes Res. 9:83–91. Lindegaard, C. 1992. The role of zoobenthos in energy flow in deep, oligotrophic Lake Thingvallavatn, Iceland. Hydrobiol. 243:185–195. Lowe, R.L., and Pillsbury, R.P. 1995. Shifts in benthic algal community structure and function following the appearance of zebra mussels (Dreissena polymorpha) in Saginaw Bay, Lake Huron. J. Great Lakes Res. 21:558–566. Lynch, M. 1979. Predation, competition and zooplankton community structure. Limnol. Oceanogr. 26:86–102. MacIsaac, H.J., Sprules, W.G., and Leach, J.H. 1991. Ingestion of small-bodied zooplankton by zebra mussels (Dreissena polymorpha): Can cannibalism on larvae influence population dynamics? Can. J. Fish. Aquat. Sci. 48:2051–2060. ———, Lonnee, C.J., and Leach, J.H. 1995. Suppression of microzooplankton by zebra mussels: importance of mussel size. Freshwat. Biol. 34:379–387. ———, Johannsson, O.E., Ye, J., Sprules, W.G., Leach, J.H., McCorquodale, J.A., and Grigorovich, I.A. 1999. Filtering impacts of an introduced bivalve (Dreissena polymorpha) in a shallow lake: application of a hydrodynamic model. Ecosystems 2:338–350. Mackie, G.L. 1993. Biology of zebra mussel (Dreissena polymorpha) and observations of mussel colonization on unionid bivalves in Lake St. Clair of the Great Lakes. In Zebra mussels: biology, impacts and control, ed. T. Nalepa and D. Scholesser, pp. 153–166. Boca Raton, Florida: Lewis Publishers. Madenjian, C.P. 1995. Removal of algae by the zebra mussel (Dreissena polymorpha) population in western Lake Erie: a bioenergetics approach. Can. J. Fish. Aquat. Sci. 52:381–390. 51 Makarewicz, J.C., and Likens, G.E. 1979. Structure and function of the zooplankton community in Mirror Lake, New Hampshire. Ecol Monogr. 49:109–127. Malley, D.F., Lawrence, S.G., MacIver, M.A., and Findlay, W.J. 1989. Range of variation in estimates of dry weight for planktonic Crustacea and Rotifera from temperate North American lakes. Can. Tech. Rep. Fish. Aqua. Sci. No. 1666. McDonald, M., Crowder, L., and Brandt, S. 1990. Changes in Mysis and populations in southeastern Lake Michigan: A response to shifts in the fish community. Limnol. Oceanogr. 35:220–227. McLaren, I.A., Tremblay, M.J., Corkett, C.J., and Roff, J.C. 1989. Copepod production on the Scotian shelf based on life history analyses and laboratory rearings. Can. J. Fish. Aquat. Sci. 46:560–583. McQueen, D.J., Post, J.R., and Mills, E.L. 1986. Trophic relationships in freshwater pelagic ecosystems. Can. J. Fish. Aquat. Sci. 43:1571–1581. ———, Johannes, M.R.S., Post, J.R., Stewart, T.J., and Lean, D.R.S. 1989. Bottom-up and top-down impacts on freshwater pelagic community structure. Ecol. Monogr. 59:289–309. Mellina, E., Rasmussen, J.B., and Mills, E.L. 1995. Impact of zebra mussel (Dreissena polymorpha) on phosphorus cycling and chlorophyll in lakes. Can. J. Fish. Aquat. Sci. 52:2553–2573. Menzie, C.A. 1981. Production ecology of Crictopus sylvestis (Fabricius) (Diptera: Chironomidae) in a shallow estuarine cove. Limnol. Oceanogr. 26: 467–481. Millard, E.S., Myles, D.M., Johannsson, O.E., and Ralph, K.M. 1996a. Phytoplankton photosynthesis at two index stations in Lake Ontario 1987–92: assessment of the long-term response to phosphorus control. Can. J. Fish. Aquat. Sci. 53:1092–1111. ———, Myles, D.M., Johannsson, O.E., and Ralph, K.M. 1996b. Phosphorus demand of Lake Ontario phytoplankton at two index stations: light vs. phosphorus limitation of growth. Can. J. Fish. Aquat. Sci. 53: 1112–1124. ——— , Fee, E.J., Myles, D.D., and Dahl, J.A. 1999. Comparison of phytoplankton photosynthesis using 14C-incubator techniques and numerical modeling in Lakes Erie, Ontario, The Bay of Quinte and the Northwest Ontario Lake Size Series (NOLSS). In The State of Lake Erie Ecosystem (SOLE)—past, present and future, eds. M. Munawar and T. Edsall. The Netherlands: Backhuys Publishers, (in press). Mills, E.L., Green, D.M., and Schiavone Jr., A. 1987. Use of zooplankton size to assess the community structure of fish populations in freshwater lakes. North Am. J. Fish. Man. 7:369–378. Mortimer, C.H. 1987. Fifty years of physical investigations and related limnological studies on Lake Erie, 1928–1977. J. Great Lakes Res. 13(4):407– 435. Nalepa, T.F., and Fahnenstiel, G.L. 1995. Dreissena 52 Johannsson et al. polymorpha in the Saginaw Bay. Lake Huron ecosystem: overview and perspective. J. Great Lakes Res. 21:411–416. Officer, C.B., Smayda, T.J., and Mann, R. 1982. Benthic filter feeding: a natural eutrophication control. Mar. Ecol. Prog. Ser. 9:203–210. OMNR (Ontario Ministry of Natural Resources). 1995. Lake Erie Fisheries Report 1994. Lake Erie Committee Meeting, Great Lakes Fishery Commission, Buffalo, N.Y., March 22–23, 1995. Paloheimo, J.E. 1974. Calculations of instantaneous birth rate. Limnol. Oceanogr. 19:692–694. Peterson, J.H. 1984. Larval settlement behaviour in competing species: Mytilus californianus Conrad and M. edulis L. J. Exp. Mar. Biol. Ecol. 82:147–159. Reynolds, C., and Wiseman, S.W. 1982. Sinking losses of phytoplankton in closed limnetic systems. J. Plank. Res. 4(3):489–522. ——— , Wiseman, S.W., and Clark, M.S.O. 1984. Growth- and loss-rate responses of phytoplankton to intermittent artificial mixing and their potential application to the control of planktonic algal biomass. J. Appl. Ecol. 21:11–39. Ricciardi, A., Whoriskey, F.G., and Rasmussen, J.B. 1996. Impact of the Dreissena invasion on native unionid bivalves in the upper St. Lawrence River. Can. J. Fish. Aquat. Sci. 53:1434–1444. Rosen, R.A. 1981. Length-dry weight relationships of some freshwater zooplankton. J. Freshwat. Ecol. 21: 225–229. Shevtsova, L.V., Zhdanova, G.A., Movchan, V.A., and Primak, A.B. 1986. Experimental interrelationship between Dreissena and planktic invertebrates. Hydrobiological Journal 22:36–39. Shuter, B.J., and Ing, K.K. 1997. Factors affecting the production of zooplankton in lakes. Can. J. Fish. Aquat. Sci. 54:359–377. Skubinna, J.P., Coon, T.G., and Batterson, T.R. 1995. Increased abundance and depth of submersed macrophytes in response to decreased turbidity in Saginaw Bay, Lake Huron. J. Great lakes Res. 221:476–487. Sprung, M. 1993. The other life: an account of present knowledge of the larval phase of Dreissena polymorpha. pp. 39–54. In Zebra mussels: biology, impacts and control, ed. T. Nalepa and D. Scholesser, pp. 39–54. Boca Raton, Florida: Lewis Publishers. ———, and Rose, U. 1988. Influence of food size and food quantity on the feeding of the mussel Dreissena polymorpha. Oecologia 77:526–532. Stanczykowska, A., and Lewandowski, K. 1993. Effect of filtering activity of Dreissena polymorpha (Pall.) on the nutrient budget of the littoral of Lake Mikolajskie. Hydrobiol. 251:73–79. Stewart, T.W., and Haynes, J.M. 1994. Benthic macroinvertebrate communities of southwestern Lake Ontario following invasion of Dreissena. J. Great Lakes Res. 20:479–493. Stockwell, J., and Johannsson, O.E. 1997. Temperaturedependent, allometric models to estimate zooplankton production in temperate, freshwater lakes. Can. J. Fish. Aquat. Sci. 54:2351–2360. Strayer, D., and Likens, G.E. 1986. An energy budget for the zoobenthos of Mirror Lake, New Hampshire. Ecol 67:303–313. Ten Winkel, E. H., and Davids, C. 1982. Food selection by Dreissena polymorpha Pallas (Mollusca: Bivalvia). Freshwat. Biol. 12:553–558 Tudorancea, C., Green, R.H., and Huebner, J. 1979. Structure, dynamics and production of the benthic fauna in Lake Manitoba. Hydrobiol. 64:59–95. Wells, L. 1970. Effects of alewife predation on zooplankton populations in Lake Michigan. Limnol. Oceanogr. 15:556–565. Witzel, L., Burr, M., Czapla, T., Einhouse, D., Haas, R., Kenyon, R., Knight, R., Muth, K., Nepszy, S., Sztramko, L., and Thomas, M. 1994. Report of the Lake Erie Forage Task Group. Great Lakes Fisheries Commission, Lake Erie Committee. Submitted: 13 August 1998 Accepted: 12 October 1999 Editorial handling: W. Gary Sprules Benthic and Pelagic Production in Lake Erie: Post-Dreissena 53 APPENDIX 1a. Length-weight relationships used to estimate zooplankton biomass in this study. W = aLb, where length is in mm and weight is in µg dry weight. Biomass of Dreissena was determined using W = a+bL+cL2 where length is in microns and weight is in 10–3 µg dry weight. Taxon CLADOCERA Bosmina, Eubosmina Daphnia, Diaphanosoma, Sida spp. Polyphemus pediculus Holopedium gibberum Chydorus sphaericus Alona sp. Bythothrephes cederstroemi Leptodora kindti COPEPODA Calanoida Generic equation Epischura Senecella calanoides Calanoid nauplii Cyclopoida Generic Equation Mesocyclops edax Cyclopoid nauplii Harpactacoida DREISSENA VELIGERS a b c 26.6 5.00 6.93 11.21 33.23 29.70 11.13 0.44 3.13 2.84 2.15 3.04 3.21 3.48 2.77 2.67 Bottrell et al. 1976 Bottrell et al. 1976 (Dumont et al. 1975) Dumont et al. 1975 Yan (OMEE, Dorset, pers. com.) Malley et al. 1989 Dumont et al. 1975 Yan (OMEE, Dorset, pers. com.) Rosen 1981 5.50 6.50 7.70 4.20 2.46 2.63 2.33 2.48 Sprules (U. of Toronto, pers. com.) Culver et al. 1985 Culver et al. 1985 Sprules (U. of Toronto, pers. com.) 5.50 2.46 6.66 2.89 4.20 2.48 4.20 2.48 58.207 –2.636 0.037 Source Sprules (U. of Toronto, pers. com.) Culver et al. 1985 Sprules (U. of Toronto, pers. com.) Sprules (U. of Toronto, pers. com.) Hillbricht-Ilkowska and Stanczykowska 1969 APPENDIX 1b. Bëlehrádek’s co-efficients used to calculate egg development times for zooplankton production estimates by the egg-ratio method. The calanoid coefficients were derived from the relationships in Cooley and Minns (1978). Species Bosmina/Eubosmina spp. Daphnia spp. Diaphanosoma sp. Diacyclops/Tropocyclops Mesocyclops edax Cyclops vernalis Leptodiaptomus minutus L. ashlandi L.. sicilis L. siciloides Skistodiaptomus oregonensis Eurytemora affinis Epischura lacustris a 3750848 65912 1767 18901 19318 8128 40994 40994 45284 38148 156046 38148 38148 Bëlehrádek’s Coefficients α β –15.40 –3.11 –6.10 –2.12 –1.90 –1.08 –4.80 –1.77 –0.48 –1.79 –2.08 –1.67 –4.72 –2.00 –4.72 –2.00 –5.79 –2.00 –4.01 –2.00 –8.00 –2.31 –4.01 –2.00 –4.01 –2.00 54 APPENDIX 2. 1993–1994. Johannsson et al. Literature values for P/B ratios for the calculation of benthic production in Lake Erie SPECIES Coelenterata, Hydra Nematoda, Adenophorea Tardigrada Turbellaria, Tricladida Nemertea, Prostoma Oligochaeta Enchytraidae Naididae Lumbriculidae Tubificidae Hirudinae, Helobdella spp. Glossiphoria complanata Mooreobdella microstoma Mollusca Gastropoda, Physella spp. Stagnicola elodes Bithynia tentaculata Gyraulus Helisoma anceps Elimia livescens Pleurocera acuta Amnicola spp. Valvata spp. Sphaeriidae, Sphaerium spp. Sphaerium corneum Pisidium spp. Musculium securis Dreisseniidae, D. polymorpha D. bugensis Crustacea Harpacticoida, Canthocamptidae Ostracoda, Candoniidae Cyprididae Cypridopsidae Lymnocytheridae Asellidae, Caecidotea Gammaridae, Gammarus fasciatus Crangonyx Archnoidea, Hydracarina a d g Dermott 1995 Johnson and Brinkhurst 1971 Menzie 1981 L.E. this study P/B 2.7 14 5 7.3 4.6 REF. h h 4 6.1 3 4.6 3.4 2.2 1.6 f h d h e c 3.2 5 1.6 5 5 1.6 1.6 2.1 1.5 3.5 3.5 1.9 3.5 4.4 5.3 e 11 4 5 5 5 3.2 3.9 4.8 5 b e h h c i i c e d e L.E. L.E. h h SPECIES Insecta Ephemeroptera, Hexagenia Heptageniidae Tricorythodes Tricoptera, Oecetis Leptostoma Molanna Diptera Tanypodinae, Ablabesmyia Coelotanypus Procladius spp. Diamesinae, Pagastia Prodiamesinae, Monodiamesia Orthocladiinae, Crictopus spp. Heterotrissocladius changi Parakiefferiella Parametriocnemus Tanytarsini, Tanytarsus spp. Chironomini, Chironomus spp. Chironomus anthracinus grp. C. semireductus grp. C. thummi Cryptochironomus Demicryptochironomus Harnischia Parachironomus Paracladopelma Dicrotendies Microtendipes Paratendipies Polypedilum Tribelos Paralauterborniella Stictochironomus Pseudochironomus P/B REF. 2.2 4 3.7 5 5 5 a e 4.5 4 2.8 5 1.5 12 3.2 4 4 6.4 4 3.6 4 4 4.7 4.7 3.9 6.7 4.7 5 5.7 4 6.4 6 6.9 6 4 e e b m f i b d i e c h c L.E. Dermott et al. 1977 Krueger and Waters 1983 Strayer and Likens 1986 c Edmondson and Winberg f Lindegaard 1992 i Tudorancea et al. 1979 1969