Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
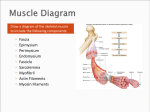
NAME: AKPANWA JR., OWOANAM MATRIC NO: 14/MHS01/024 DEPARTMENT: MEDICINE AND SURGERY LEVEL: 200 HISTOLOGY OF MUSCLE TISSUE Introduction Muscle tissue has a unique histological appearance which enables it to carry out its function. In this particle, we will examine the histology and how it relates to contractility. Muscle tissue is composed of elongated cells specialized for contraction and movement. Muscle tissue is composed of cells differentiated for optimal use of the universal cell property termed contractility. Microfilaments and associated proteins together generate the forces necessary for cellular contraction, which drives movement within certain organs and the body as a whole. Nearly all muscle cells are of mesodermal origin and they differentiate mainly by a gradual process of cell lengthening with simultaneous synthesis of myofibrillar proteins. Three types of muscle tissue can be distinguished on the basis of morphologic and functional characteristics and the structure of each type is adapted to its physiologic role Types of Muscle There are three types of muscle: Skeletal – is composed of bundles of very long, cylindrical, multinucleated cells that show cross-striations. Their contraction is quick, forceful, and usually under voluntary control. It is caused by the interaction of thin actin filaments and thick myosin filaments whose molecular configuration allows them to slide upon one another. The forces necessary for sliding are generated by weak interactions in the bridges between actin and myosin,A form of striated muscle that is under voluntary control from the somatic nervous system. Identifying features are cylindrical cells and multiple peripheral nuclei. Cardiac – A form of striated muscle that is found only in the heart. Identifying features are single nuclei and the presence of intercalated discs between the cells. it also has cross-striations and is composed of elongated, branched individual cells that lie parallel to each other. At sites of end-to-end contact are the intercalated disks, structures found only in cardiac muscle. Contraction of cardiac muscle is involuntary, vigorous, and rhythmic Smooth – Skeletal muscle consists of muscle fibers, which are long, cylindrical multinucleated cells with diameters of 10–100 m. Multinucleation results from the fusion of embryonic mesenchymal cells called myoblasts (Figure 10–2). The long oval nuclei are usually found at the periphery of the cell under the cell membrane. This characteristic nuclear location is helpful in discriminating skeletal muscle from cardiac and smooth muscle, both of which have centrally located nuclei. A form of non-striated muscle that is controlled involuntarily by the autonomic nervous system. The identifying feature is the presence of one spindle-shaped central nucleus per cell. This article will deal mainly with skeletal muscle. Composition of Skeletal Muscle A muscle cell is very specialised for its purpose. One muscle cell is known as a muscle fibre, and its cell surface membrane is known as the sarcolemma. Other organelles that are unique to muscle are T tubules, which are invaginations of the sarcolemma that conduct charge when the cell is depolarised. Muscle cells also have a specialised endoplasmic reticulum – this is known as the sarcoplasmic reticulum and contains a large store of calcium ions. Muscles also have an intricate support structure of connective tissue. Each muscle fibre is surrounded by a thin layer of connective tissue known as endomysium. These fibres are then grouped into bundles known as fascicles, which are surrounded by a layer of connective tissue known as perimysium. Many fascicles make up a muscle, which in turn is surrounded by a thick layer of connective tissue knows as the epimysium. Muscle Fibers As observed with the light microscope, longitudinally sectioned skeletal muscle fibers show cross-striations of alternating light and dark bands (Figure 10–7). The darker bands are called A bands (anisotropic or birefringent in polarized light); the lighter bands are called I bands (isotropic, do not alter polarized light). In the TEM each I band is seen to be bisected by a dark transverse line, the Z line (Ger. Zwischenscheibe, between the discs). The repetitive functional subunit of the contractile apparatus, the sarcomere, extends from Z line to Z line (Figure 10–8) and is about 2.5 m long in resting muscle. Myosin is a much larger complex (molecular mass ~500 kDa). Myosin can be dissociated into two identical heavy chains and two pairs of light chains. Myosin heavy chains are thin, rod-like molecules (150 nm long and 2–3 nm thick) made up of two heavy chains twisted together as myosin tails. Small globular projections at one end of each heavy chain form the heads, which have ATP binding sites as well as the enzymatic capacity to hydrolyze ATP (ATPase activity) and the ability to bind actin. The four light chains are associated with the head. Several hundred myosin molecules are arranged within each thick filament with their rodlike portions overlapping and their globular heads directed toward either end. Ultra structural Appearance of Skeletal Muscle The striated appearance of skeletal muscle fibres arises due to the organisation of two contractile proteins or myofilaments, actin (thin filament) and myosin (thick filament). The functional unit of contraction is in a skeletal muscle fibre is the sarcomere, which runs from Z line to Z line. A sarcomere is broken down into a number of sections: Z line – Where the actin filaments are anchored. M line – Where the myosin filaments are anchored. I band – Contains only actin filaments. A band – The length of a myosin filament, may contain overlapping actin filaments. H zone – Contains only myosin filaments. The M line is inside the H zone which is inside the A band, whilst the Z line is inside the I band. Sliding Filament Model Myosin filaments have many heads, which can bind to sites on the actin filament. Actin filaments are associated with two other regulatory proteins, troponin and tropomyosin. Tropomyosin is a long protein that runs along the actin filament, blocking the myosin head binding sites. Troponin is a small protein that binds the tropomyosin to the actin. It is made up of three parts: Troponin I - which binds to the actin filament. Troponin T - which binds to tropomyosin. Troponin C - which can bind calcium ions. The unique structure of troponin is the basis of excitation-contraction coupling: 1. When depolarisation occurs at a neuromuscular junction, this is conducted down the T tubules, causing a huge influx of calcium ions into the sarcoplasm from the sarcoplasmic reticulum. 2. This calcium binds to troponin C, causing a change in conformation that moves tropomyosin away from the myosin head binding sites of the actin filaments. 3. This allows the myosin head to bind to the actin, forming a cross-link. The power stroke then occurs as the myosin heads pivots in a ‘rowing motion, moving the actin past the myosin towards the M line. 4. ATP then binds to the myosin head, causing it to uncouple from the actin and allowing the process to repeat. Hence in contraction, the length of the filaments does not change. However, the length of the sarcomere decreases due to the actin filaments sliding over the myosin. The H zone and I band shorten, whilst the A band stays the same length. This brings the Z lines closer together and causes overall length to decrease.