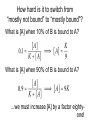Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
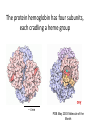
Recap from last time: perspective on DG of ATP hydrolysis, -50 kJ/mol At room temperature, kbT = 1.38 x 10-23 J/K * 298 K * 6 x 1023 mol-1 = 2.5 kJ/mol The energy required to break a typical covalent bond = 400 kJ/mol Recap from last time: enthalpies of formation for major biological macromolecules Carbohydrates: Proteins: Fatty acids: 4 kcal/g 4 kcal/g 9 kcal/g 1 lb of fat ≈ 450 g or 4000 kcal Typical burn per mile: 100 kcal Recap from last time: energy density of macromolecules Solubility limit of carbohydrates in water: 2000 g/L -- coke: 160 g/L -- glycogen: 4% of muscle mass, or 40 g/L of muscle -- “pre-diabetic” blood glucose: 1.8 g/L (10 mM) Adipose tissue density: ~1 g/mL -- lipid fraction: ~70% (nearly as good as stacked oranges) Plan for this unit Explore the energy budget of the cell: • How much energy is available? 1000 W/m2 • How do we capture it? e- excitation -> ATP • How do we store it? bio. macromolecules • How much do we need to make the components of a cell? • How efficient is the process? To double, an E. coli cell in LB uses glucose as a primary source of both carbon and energy How much sugar do we need to double an E. coli cell’s carbon #? (i.e., roughly 2 x 109 glucose molecules for raw materials) How much more sugar is needed for energy to build macromolecules? 1. Calculate the total energy needed to synthesize all of an E. coli cell’s protein – Half of the dry mass of the cell is protein, so for simplicity, we’ll just double that value to estimate the total energy needed for cell component synthesis 2. Find the energy we can obtain through respiration of one glucose molecule – Somewhat less than the energy originally required to synthesize the sugar 3. Divide to find the theoretical minimum number of sugar molecules needed for energetics How much energy is needed to synthesize amino acids and incorporate them into protein? Dry mass of protein per cell: 1.5 x 10-13 g Typical amino acid mass: 100 Da or 1.5 x 10-22 g # of amino acids per cell: 1.5E-13 / 1.5E-22 = 109 Cost to synthesize*: ~ 1 ATP/amino acid Cost to incorporate: ~ 4 ATP/amino acid Total: 5 x 109 ATP equivalents Valine 2O 5C 11 H 1N x 16 x 12 x1 x 14 Total = = = = = 32 60 11 14 117 Da How many glucose molecules do we need to get that much energy? • Can generate ~40 ATP equivalents per glucose molecule through respiration – For comparison, photosynthesis required 2700 kJ/mol or roughly 50 ATP equivalents to construct the glucose molecule in the first place How many glucose molecules do we need to build an E. coli cell? • Can generate ~40 ATP equivalents per glucose molecule through respiration – For comparison, photosynthesis required 2700 kJ/mol or roughly 50 ATP equivalents to construct the glucose molecule in the first place • 5 x 109 / 40 ≈ 1.2 x 108 glucose molecules needed for energetic purposes – x 2 to account for synthesis of NTs, lipids, etc. – Add to glucose molecules needed for raw materials • Grand total: ~ 2 x 109 glucose molecules – Dominated by need for carbon, not energy How many glucose molecules does an E. coli cell typically use to double? Fresh media: 0.1 mol glucose/L, ~ 1 cell Spent media: 0 mol glucose/L, ~ 1012 cells 0.1 mol * 6 x 1023 / 1012 cells = 6 x 1010 glucose molecules/cell Why is much more glucose used than predicted in this calculation? Predicted: 2 x 109 glucose molecules/cell Observed: 6 x 1010 glucose molecules/cell E. coli growing on LB are fermenting, not respiring • 2 ATP equivalents/glucose molecule vs. 40 • Accounts for most of the discrepancy! • Note that fermenting E. coli need more glucose for energetics than for raw carbon Plan for this unit Explore the energy budget of the cell: • How much energy is available? 1000 W/m2 • How do we capture it? e- excitation -> ATP • How do we store it? Biolog. macromolecules • How much do we need to make the components of a cell? 1-10x own dry mass in glucose! • How efficient is the process? How much of the available solar energy are plants able to capture? Of all incident light: • ≈ 37% is absorbed by a pigment molecule • ≈ 28% successfully mobilizes an e• ≈ 9% of energy is used to synthesize sugars • ≈ 7% of energy remains usable after respiration (0.3% after fermentation) How much energy do organisms really need to collect? Need 5 x 109 ATP equivalents = 4 x 10-10 J per cell division 20 minutes = 1200 s per cell division -> 3 x 10-13 W E. coli’s surface area is 1 x 10-12 m2 -> 0.3 W/m2 For a human (2000 kcal/day, 2 m2 surface area) -> 50 W/m2 Energy budget of the cell summary • • • • How much energy is available? 1000 W/m2 How do we capture it? e- excitation -> ATP How do we store it? Biolog. macromolecules How much do we need to make the components of a cell? 1-10x own dry mass in glucose! • How efficient is the process? Not very – but good enough! Where are we headed in LS 50? • Two lectures on cooperativity – Biochemical approach and stat mech approach • Four lectures on diffusion – Chemical potential differences as a driver for molecular motion • Three lectures on natural and synthetic gene regulation – Applications of cooperativity and diffusion Diffusion tends to equalize concentrations by driving molecules from regions of high conc. to regions of low conc. • For thin organisms, diffusion is enough to ensure that oxygen continually flows from the air or water into tissues (where the concentration is lower since it’s being consumed) • No circulatory system or other bells & whistles necessary • Steady-state concentration profile same as for the Bicoid problem on PS6 (falls off rapidly) – big problem for thick organisms! A circulatory system can help transport O2, but is also limited by diffusion That’s all? RB C Once the concentration of oxygen is the same inside the red blood cell as in the lung, the net flow of oxygen into the RBC Nature’s solution: molecular hoarding with hemoglobin Step one: Stash the oxygen RB C Nature’s solution: molecular hoarding with hemoglobin Step one: Stash the oxygen RB C Bound oxygen does not count toward the concentration in the red blood cell’s cytoplasm, so more oxygen can enter by Nature’s solution: molecular hoarding with hemoglobin Step one: Stash the oxygen RB C Nature’s solution: molecular hoarding with hemoglobin Step two: Advection (flow of red blood cells in the circulatory system) Nature’s solution: molecular hoarding with hemoglobin Step three: Unloading RB C Cooperativity Lecture Outline • • • • Introduction to heme and hemoglobin Limitations of simple binding Cooperativity as a potential improvement Mechanistic understanding of cooperativity in biology The iron atom in heme binds oxygen Heme The protein hemoglobin has four subunits, each cradling a heme group ~ 4 nm PDB May 2003 Molecule of the Iron oxidation state and coordination state change on oxygen binding Superoxide (O2-) Fe2+ Fe3+ Reminder: some transition metals with close d orbital spacing absorb visible light Chlorophyll Heme Oxygen binding changes the absorption spectrum, reflecting more red light Courtesy of Scott Prahl The fraction of hemoglobin bound by oxygen bound can be estimated from an absorption spectrum Courtesy of Scott Prahl Liu et al., 2012 The fraction of hemoglobin bound by oxygen bound can be estimated from an absorption spectrum Liu et al., 2012 This curve should surprise you! Recap: Simple binding curves are hyperbolic How hard is it to switch from “mostly not bound” to “mostly bound”? How hard is it to switch from “mostly not bound” to “mostly bound”? What is [A] when 10% of B is bound to A? What is [A] when 90% of B is bound to A? …we must increase [A] by a factor eightyone! Hemoglobin, an oxygen-binding protein, has a sigmoidal curve Switches from 10% to 90% bound over an ≈ four-fold change in oxygen availability Hemoglobin has four oxygen binding sites: Is this responsible for its sigmoidal curve? Suppose all four sites are identical and independent (i.e., each site is unaffected by the binding of O2 elsewhere) with binding reaction rate kon and dissociation reaction rate koff P: Protein (hemoglobin) X: Ligand (O2) Finding the binding curve when sites are independent What is the total concentration of binding sites? What is the total concentration of bound sites? What is the fraction of sites bound?