Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Danni al DNA e meccanismi di riparazione
Le mutazioni del DNA
Mutations and Consequences
Mutation refers to the change in a DNA sequence, which may involve only a few bases or
the large-scale chromosome abnormality. This section covers the small-scale mutations
(substitution, deletion, insertion) and the exon skipping that results from mutation at the
splice site.
Substitution
In the substitution mutation, one or more nucleotides are substituted by the same number of
different nucleotides. In most cases, only one nucleotide is changed. Based on the change
in the nucleotide type, the substitution mutation may be divided into transition and
transversion mutations. Based on the consequence of mutation, the substitution mutation
may be grouped into silent, missense and nonsense mutations
Deletion
The deletion mutation involves elimination of one or more nucleotides from a DNA
sequence. It may cause frameshift, producing a non-functional protein.
Note that deletion and insertion mutations often occur in the repetitive sequence. As
explained in the next section, they are usually caused by "replication slippage".
Insertion
In the insertion mutation, one or more nucleotides are inserted into a sequence. If the number
of inserted bases is not a multiple of 3, it will cause frameshift, resulting in serious
consequences. As shown in the following table, non-frameshift insertions may also cause
diseases.
The substitution mutation.
(a) Illustration of transition (blue) and transversion (red) mutations. In the transition mutation,
a pyrimidine (C or T) is substituted by another pyrimidine, or a purine (A or G) is substituted
by another purine. The transversion mutation involves the change from a pyrimidine to a
purine, or vice versa.
(b) Examples of silent, missense and nonsense mutations. The silent mutation does not
produce any change in the amino acid sequence, the missense mutation results in a different
amino acid, and the nonsense mutation generates a stop signal.
Real examples of deletion mutations which cause diseases. (a) Deletion of "T" from the
sequence "TTTTT" in the CFTR gene.
(b) Deletion of "AT" from the sequence "ATAT" in the CFTR gene.
(c) Deletion of "TTG" from the sequence "TTGTTG" in the FIX gene.
(d) Deletion of "ATAG" from the sequence "ATAGATAG" in the APC gene.
Examples of diseases caused by insertion mutation.
Mutation by Replication Errors
Replication errors are the main source of mutations. It has been estimated that uncorrected
replication errors occur with a frequency of 10-9 - 10-11 for each nucleotide added by DNA
polymerases. Since a cell division requires synthesis of 6 X 109 nucleotides, the mutation
rate is about one per cell division.
A commonly observed replication error is the replication slippage, which occurs at the
repetitive sequences when the new strand mispairs with the template strand. The
microsatellite polymorphism is mainly caused by the replication slippage. If the mutation
occurs in a coding region, it could produce abnormal proteins, leading to diseases. The
Huntington's disease is a well known example.
Exon skipping
Example of exon skipping. Splicing of an intron requires an essential signal:
"GT........AG". If the splice acceptor site AG is mutated (e.g., A to C in this figure), the
splicing machinery will look for the next acceptor site. As a result, the exon between two
introns is also removed.
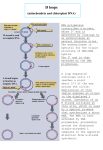
The mutation
caused by
replication
slippage. In this
figure, mispairing
involves only one
repeat. In fact, the
slippage could cause
several repeats to
become
unpaired. (a) Normal
replication. (b)
Backward slippage,
resulting in the
insertion
mutation. (c)
Forward slippage,
resulting in the
deletion mutation.
Mutation Mechanisms
Mutations may be caused by external factors (UV radiation, exposure to chemical agents,
etc.) or spontaneous cellular processes (accidental deamination, replication errors, etc.).
Examples of deamination which involves the removal of an amino group. Accidental
deamination may change the cytosine to uracil, or the methylated cytosine to thymine.
DNA Repair Mechanisms
There are three major DNA repairing mechanisms (damage removal): base excision,
nucleotide excision and mismatch repair.
Proteins involved in the DNA repairing of E. coli.
Tipo
Danno
Proteine di riparazione
Dimero di pirimidina o sito
apurinico
DNA pol. IV e V in E. coli
Pol., , , e negli esseri
umani
Fotoriattivazione
Dimeri di pirimidina
DNA fotoliasi
Rimozione di gruppi metilici
O6-Metilguanina
Metiltransferasi
Riparazione per escissione
delle basi
Base danneggiata
DNA glicosilasi
Riparazione delle basi male
appaiate
Errori di replicazione
MutS,MutL, e MutH in E. coli
MutS, MutL e EXO1 negli
esseri umani
Riparazione per escissione dei
nucleotidi
Dimeri di pirimidina
Grandi addotti sulle basi
UvrA, UvrB, UvrC e UvrD in
E. coli
XPA, XPB, XPC, XPD,
ERCC1/XPF e XPG negli
esseri umani
Riparazione per rottura a
doppio filamento
Rotture a doppio filamento
RecA e REcBCD in E. coli
Complesso MRN, Rad51,
BRCA1, BRCA2, XRCC3,
ecc. negli esseri umani per la
ricombinazione omologa
Proteine Ku, Artemis/DNAPKCS, XRCC4 negli esseri
umani per l’unione non
omologa delle estremità
Aggiramento del danno
Sistemi di riparazione
del DNA
Sintesi translesione del DNA
Inversione del danno
Rimozione del danno
Cytosine is one of four bases in DNA molecules. As shown in the above figure, it may be
mutated to uracil by deamination. Since uracil is not part of DNA, this mutation can easily
be detected and repaired by base excision. Suppose DNA, like RNA, were made up of
uracil, then the cytosine to uracil mutation could be corrected only by mismatch repair
which is very inefficient. This may explain why DNA chooses thymine, instead of uracil,
even though the chemical structure of uracil is simpler than thymine.
Base excision
DNA's bases may be modified by deamination or alkylation. The position of the
modified (damaged) base is called the "abasic site" or "AP site". In E.coli, the DNA
glycosylase can recognize the AP site and remove its base. Then, the AP endonuclease
removes the AP site and neighboring nucleotides. The gap is filled by DNA polymerase I
and DNA ligase.
Nucleotide excision
In E. coli, proteins UvrA, UvrB, and UvrC are involved in removing the damaged
nucleotides (e.g., the dimer induced by UV light). The gap is then filled by DNA
polymerase I and DNA ligase. In yeast, the proteins similar to Uvr's are named RADxx
("RAD" stands for "radiation"), such as RAD3, RAD10. etc.
Modello della sintesi translesione
del DNA
Riparazione del DNA mediante rimozione dei gruppi metilici
Via di riparazione
per escissione
delle basi nelle
cellule dei
mammiferi
Via di riparazione delle
basi male appaiate
nelle cellule dei mammiferi
Via di riparazione per
escissione dei nucleotidi
nei mammiferi
DNA repair by base excision.
DNA repair by nucleotide excision.
Mismatch repair
To repair mismatched
bases, the system has
to know which base is
the correct one. In E.
coli, this is achieved
by a special methylase
called the "Dam
methylase", which can
methylate all adenines
that occur within
(5')GATC
sequences. Immediate
ly after DNA
replication, the
template strand has
been methylated, but
the newly synthesized
strand is not
methylated yet. Thus,
the template strand
and the new strand can
be distinguished.
Mismatch repair.
The repairing process begins with the protein MutS which binds to mismatched base
pairs. Then, MutL is recruited to the complex and activates MutH which binds to GATC
sequences. Activation of MutH cleaves the unmethylated strand at the GATC
site. Subsequently, the segment from the cleavage site to the mismatch is removed by
exonuclease (with assistance from helicase II and SSB proteins). If the cleavage occurs on the
3' side of the mismatch, this step is carried out by exonuclease I (which degrades a single
strand only in the 3' to 5' direction). If the cleavage occurs on the 5' side of the mismatch,
exonuclease VII or RecJ is used to degrade the single stranded DNA. The gap is filled by
DNA polymerase III and DNA ligase.
The distance between the GATC site and the mismatch could be as long as 1,000 base
pairs. Therefore, mismatch repair is very expensive and inefficient.
Mismatch repair in eukaryotes may be similar to that in E. coli. Homologs of MutS and MutL
have been identified in yeast, mammals, and other eukaryotes. MSH1 to MSH5 are
homologous to MutS; MLH1, PMS1 and PMS2 are homologous to MutL. Mutations of
MSH2, PMS1 and PMS2 are related to colon cancer.
In eukaryotes, the mechanism to distinguish the template strand from the new strand is still
unclear.
Origin of CpG (CG) islands
The CG island is a short stretch of DNA in which the frequency of the CG sequence is
higher than other regions. It is also called the CpG island, where "p" simply indicates
that "C" and "G" are connected by a phosphodiester bond.
CpG islands are often located around the promoters of housekeeping genes (which
are essential for general cell functions) or other genes frequently expressed in a
cell. At these locations, the CG sequence is not methylated. By contrast, the CG
sequences in inactive genes are usually methylated to suppress their expression.
The methylated cytosine may be converted to thymine by accidental
deamination. Unlike the cytosine to uracil mutation which is efficiently repaired, the
cytosine to thymine mutation can be corrected only by the mismatch repair which is
very inefficient. Hence, over evolutionary time scales, the methylated CG sequence
will be converted to the TG sequence. This explains the deficiency of the CG
sequence in inactive genes.
Inheritance of the DNA
methylation pattern
Any type of cells have
their own methylation
pattern so that a unique set
of proteins may be
expressed to perform
functions specific for this
cell type. Thus, during
cell division, the
methylation pattern should
also pass over to the
daughter cell. This is
achieved by a specific
enzyme called the
maintenance methylase
which can methylate only
the CG sequence paired
with methylated CG.
Mutation by UV light
UV light may cause two
adjacent pyrimidine residues
(cytosine or thymine) to form
a dimer. In a normal cell, the
dimer can be detected by p53,
which then triggers the
repairing process. However,
if p53 itself is mutated and
become non-functional, the
pyrimidine dimer may lead to
mutation.
Figure. Pyrimidine
dimer induced by
UV light. This
figure uses thymine
as an
example. Cytosine
may form a similar
dimer
A possible mechanism for the
mutation induced by UV light.
The UV light first causes two
adjacent cytosine residues to form a
dimer. During DNA replication, both
strands are used as templates to
synthesize new strands. The cytosine
dimer could cause adenine (instead
of the normal guanine) to be
incorporated into the new
strand. Subsequent DNA replication
will produce CC to TT
mutation. Although the cytosine
dimer may eventually be corrected,
the mutation cannot be detected by
the DNA repair system.
Mutation by
Chemical Agents
The chemical agents which may
cause mutation are called
mutagens. Most of them are
also carcinogens.
•Acridines (e.g., proflavin) are positively
charged molecules. They may be inserted
between two DNA strands, thereby altering
DNA's structure and rigidity. As a result,
DNA replication will not be faithful.
•Alkylating agents are chemicals that add an
alkyl group (CnH2n+1) to another
molecule. Alkylation of a base may change
the normal base pairing. For example, the
alkylating agent EMS converts guanine to 7ethylguanine which pairs with thymine. The
mispairing will lead to mutation. Some
alkylating agents may also cross-link DNA,
resulting in chromosome breaks.
•Nitrous acid is a deaminating agent that
converts cytosine to uracil, adenine to
hypoxanthine, and guanine to xanthine. The
hydrogen-bonding potential of the modified
base is altered, resulting in mispairing.
•Hydroxylamine and free radicals may
modify base structures, resulting in
mispairing.
Mechanism of mutation induced by 5-bromouracil. This
molecule has two tautomeric isoforms. Its keto form (BUk)
pairs with adenine whereas its enol form (BUe) pairs with
guanine. Suppose in the first replication the keto form was
incorporated into a new DNA strand. During the second
replication, if the keto form undergoes a tautomeric shift to the
enol form, it will cause A:T to G:C mutation.
(a) Base structures induced by free radicals. (b) The base change induced by NH2OH.
Riparazione delle rotture a
doppio filamento
Questi meccanismi operano sia nei procarioti che negli eucarioti.
Il meccanismo di unione non omologa è attivo durante tutto il ciclo cellulare.
L’unione omologa ripara le rotture a doppio filamento a livello delle forcelle di
replicazione.
Unione omologa – ricombinazione omologa
• La ricombinazione omologa svolge ruoli essenziali negli
eucarioti:
• Ricombinazione omologa durante la meiosi.
• Ricombinazione omologa nella trasposizione.
• Ricombinazione omologa per il cambiamento del tipo di
accoppiamento nel lievito.
• Ricombinazione omologa per lo scambio di antigeni nei
tripanosomi.
ATM: ataxia-telangectasia mutated.
Unione omologa –
ricombinazione omologa
Proteine coinvolte:
Ser/Thr chinasi nucleare, ATM (trasduttore chiave) aumenta
l’attività di questa chinasi.
ATM fosforila proteine coinvolte nella riparazione del DNA (BRCA1)
e nel controllo del ciclo cellulare (p53; proteina che sopprime i
tumori).
Localizzazione nei siti di rottura di: ATM, proteina Rad52 il
complesso Mre11-Rad50-Nbs1 (MRN) inizia la riparazione (Mre11:
esonucleasi 3’ 5’.
Il DNA a singolo filamento sono riconosciuti da: Rad51.
Altre proteine sono coinvoltenella ricombinazione omologa: Rad54,
Rad55, Rad57, BRCA1 e BRCA2.
Struttura della giunzione di Holliday
(eterodimero)
(nucleasi)
Gli elementi trasponibili del genoma:
trasposoni e retrotrasposoni
Le classi degli elementi trasponibili
Classe
Intermedio di trasposizione
Esempi
Retrotrasposoni LTR
RNA
Lievito: elementi Ty;
Esseri umani: Retrovirus
endogeni umani (HERV);
Topo: particella A
intracisternali (AP).
Retrotrasposoni non LTR
LINE (autonomi)
SINE (non autonomi)
RNA
Esseri umani:
Elementi L1
Elementi Alu
DNA
Batteri:
Sequenze di inserzione
Batteriofago Mu
Trasposoni (batterifago Tn7).
Drosophila:
Elementi P.
Mais:
Elementi Ac e Ds.
Invertebrati e vertebrati:
Superfamiglia Tc1/mariner
Classe I
Classe II
Trasposoni di DNA
LINE: elementi nucleari sparsi lunghi.
SINE: elementi nucleari sparsi corti.
LTR: lunghe ripetizioni terminali