Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Jf. exp. Biol. (1978), 74, 75-8i
Witli 3 figures
Printed in Great Britain
75
REBOUND RESILIENCE OF TENDONS IN THE FEET
OF SHEEP (OVIS ARIES)
BY W. G. CUMING, R. MCN. ALEXANDER AND A. S. JAYES
Department of Pure and Applied Zoology, University of Leeds, Leeds LSz
gJT
(Received 15 August 1977)
SUMMARY
The interosseous muscle of the fore foot of the sheep is multipennate,
with large tendons but extremely short muscle fibres. It is believed to save
energy in locomotion by elastic storage in its tendons and its sheath. Its
rebound resilience (at 5-7 Hz and 20-37 °C) is about 0-62.
INTRODUCTION
A running mammal decelerates and accelerates in each step and its centre of mass
falls and rises (Alexander, 1977). Both the kinetic energy and the potential energy of
a biped are highest between steps, when neither foot is on the ground. They are lowest
in the middle of a step, as the centre of mass passes over the supporting foot. Energy
which would otherwise be lost as heat can be stored at this stage as elastic strain
energy, and re-converted to kinetic and potential energy in an elastic recoil. The same
principle applies to a bouncing ball, which stores elastic strain energy briefly when it
hits the ground. It also applies to a quadruped, which resembles two bipeds running
in tandem.
There is strong evidence that in running men, about half of the power which would
otherwise be needed is saved by elastic storage (Cavagna, Saibene & Margaria, 1964).
There is similar evidence for kangaroos (Alexander & Vernon, 1975) and it seems
reasonable to assume the importance of elastic storage in mammals in general. Rough
calculations indicate that the strain energy must be stored predominantly in tendons:
very little of it can be stored in the contractile tissue of the muscles (Alexander &
Bennet-Clark, 1977).
Suppose an elastic material is deformed and allowed to recoil. The rebound resilience
of the material is the energy recoverable in the recoil, expressed as a fraction of the
work done in the deformation. The efficacy of a material for elastic storage depends
on its having a reasonably high rebound resilience. We have devised a simple
experiment to measure the rebound resilience of a group of tendons which are believed
to be important in locomotion. They are the tendons of the interosseous muscle of
the fore-limb of the sheep, Ovis aries. The functions of the corresponding tendons in
horses were discussed by Camp & Smith (1942).
76
W. G. CUMING, R. McN.
ALEXANDER AND A. S. JAYES
5 cm (a, b)
Carpal bones
Cannon bone
Interosseous t.
1 cm {c-e)
Origin of
/-/—Interosseous sheath \ \^J / interosseous m.
-Deep flexor t.
, cannon bone
-Superficial flexor t.
I
\
Interosseous t.
Digital
extensor t.
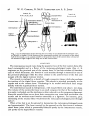
Fig. 1. (a), (b) Dissections of the left fore foot of a sheep in (a) lateral and (6) posterior view.
m, Muscle; t, tendon. (c)-{e) Sections of the interosseous muscle, (c) is a section of the distal
half of the muscle, cut in a plane parallel to the posterior face of the cannon bone, (rf) and (c)
are sections at right angles to the long axis of the cannon bone.
ANATOMY
The interosseous muscle runs along the posterior face of the fore cannon bone (the
fused metacarpals) and is a flexor of the metacarpo-phalangeal joints (Fig. 1). It
originates on the proximal part of the cannon bone. It has four tendons of insertion
each of which bifurcates; one branch of each attaches through a sesamoid to one of
the proximal phalanges while the other crosses to the anterior face of the foot and
merges with the digital extensor tendon.
The muscle is enclosed in a sheath of tough connective tissue which also encloses
the tendons of the digital flexor muscles. This sheath is attached proximally to the
carpals and to the cannon bone. Thickenings in its medial and lateral faces merge
distally with the superficial flexor tendon.
The interosseous muscle is multipennate, with muscle fibres only about 1 mm long.
The volume of the contractile tissue is so small compared to that of the tendons that
it is easily overlooked, and the tendons are often referred to as the suspensory ligament.
Since the muscle fibres are so short their contraction can cause little movement: the
muscle and its tendons must function essentially as a ligament. For instance, 30%
shortening of the muscle fibres would bend the metacarpo-phalangeal joint through
only 2°.
When a fore foot is on the ground in locomotion the metacarpo-phalangeal joints
are hyperextended. The force exerted by the ground on the foot exerts a moment
about these joints which is presumably balanced partly by the interosseous muscle,
and partly by the digital flexor muscles.
Rebound resilience of tendons in the feet of sheep (Ovis aries)
77
Clip
^
_
/
^
Pointer
Fig. 2. The apparatus used in this investigation. Not to scale.
METHODS
The fore foot of a freshly killed sheep was amputated by sawing through the radius
and ulna near their distal ends. The foot was then skinned from its proximal end to
about 4 cm distal to the carpals. The digital flexor muscles and their tendons were
removed from the skinned portion. A hole of diameter 3 mm was bored transversely
through the exposed part of the cannon bone. After skinning and throughout the
experiment the skinned part of the foot was kept moist by applying pads of cotton
wool soaked in 0-94% aqueous sodium chloride.
The soft tissues between the digits were cut and the foot was clamped in the holder
shown in Fig. 2. The posterior surface of the foot is uppermost and the bolt passes
between the digits. The holder was fixed firmly by large G-cramps (not illustrated)
to a concrete bench. A nail was passed through the hole previously bored in the cannon
bone and a mass, which was changed as required, was suspended from it by a chain.
A jubilee clip (as used for fastening hoses in automobile engines) was tightened
round the bone and interosseous tendon immediately proximal to the nail. A cloth
pad moistened with saline was placed under it, to protect the tendon. The clip was
used to hold a light pointer which wrote on a smoked-drum kymograph.
Any vertical displacement of the mass was necessarily accompanied by a change in
length of the interosseous muscle and/or its tendons. The elastic stiffness of the muscle
and tendons interacted with the mass so that the system had a natural frequency of
vibration which was 5-7 Hz in most of our experiments. With the kymograph drum
revolving, the mass was given a small vertical displacement and released. The train
of damped oscillations which followed was recorded on the drum. Particular care was
taken to ensure that the pointer pressed only very lightly against the kymograph drum,
so we are confident that energy losses through friction on the drum were very small
compared to the energy content of the oscillations. Several records were made with
each mass for each specimen. They were subsequently measured to determine
rebound resilience.
Let the amplitudes of successive half cycles of the oscillation be xlt x2, x3,. . ., xn.
The rebound resilience is then {xn+1/xnY (see, for instance, Alexander, 1968). If it is
independent of amplitude, a graph of log x against n will be a straight line: we found
that this was approximately the case. The gradient of such a graph is the logarithm
of the square root of the rebound resilience.
78
W. G. CUMING, R. McN.
ALEXANDER AND A. S. JAYES
The natural frequency of a spring-mass system which is not too heavily damped is
approximately (1/277) (S/m)t, where m is the mass and 5 is the stiffness (i.e. the force
required to effect unit displacement of the mass; see, for instance, Alexander, 1968).
Our records gave frequencies and the applied masses were known, so stiffness could
be calculated. The mass used in the calculation was the suspended mass plus the mass
of the chain, and amounted to 1-94, 5-02 or 10-56 kg. The mass of the preparation
was about 0-16 kg, and was ignored.
The stiffness so calculated was converted to torsional stiffness (i.e. to the moment
required to bend tbe joint through unit angle) by multiplying by the square of the
distance of the nail from the axis of the metacarpo-phalangeal joint. This distance
was measured from X-radiographs.
The stiffness calculated from the natural frequency was a dynamic stiffness. The
static torsional stiffness was also determined for a few specimens, from observations
of the displacement of the pointer caused by hanging weights from the nail.
The static stiffness of the apparatus was investigated by setting it up with a pair of
stout brass bars in place of the specimen. Weights were suspended from the bars at
the same distance from the holders as in the experiments with feet. It was found that
the static torsional stiffness was 300 N m rad"1. The static and dynamic torsional
stiffnesses of our preparations were always less than (and usually much less than)
50 N m rad"1. The elastic compliance of the equipment cannot have affected our
results appreciably.
The masses were suspended on a chain 0-35 m long so the natural frequency of mass
and chain, swinging as a pendulum, was about o-8 Hz. This is much lower than the
frequencies of 5-7 Hz at which the feet vibrated so it is unlikely that our results were
affected appreciably by transfer of energy between the modes of vibration. Nevertheless we took care to displace the mass vertically at the start of each train of oscillations,
trying to avoid setting it swinging as a pendulum.
Our principal experiments, of which the results are shown in Table 1, were
performed precisely as described above. Four additional experiments were performed
in which the same foot was tested both at 37 and 20 °C. For these experiments we used
feet amputated at the carpo-metacarpal joint, obtained from a slaughterhouse and
stored at — 20 CC for up to 15 days before use. These experiments were performed on
a wooden bench which was a little less rigid than the concrete bench used for our
principal experiments.
Temperatures were changed as required, by heating the laboratory.
RESULTS
Table 1 shows the results of our principal experiments. The mean of all the values
of rebound resilience recorded in it is 0-62.
The experiments in which the same feet were tested both at 37 °C and at 20 °C
failed to demonstrate any effect of temperature on rebound resilience. We used a 5 kg
mass and found that the rebound resilience was 0-52-0-71 (mean 0-64) at 37 °C, and
0-65-0-69 (mean 0-67) at 20 °C.
Table 1 and Fig. 3 show that both the dynamic and the static stiffness increase as
the suspended mass is increased. Our values of static stiffness are uniformly lower
Rebound resilience of tendons in the feet of sheep (Ovis aries)
79
Table 1. The results of the principal experiments
(Frequency is the natural frequency of the specimen, mounted in the apparatus shown in
Fig. 2 and with the indicated mass suspended from it. Torsional stiffness is the dynamic
torsional stiffness of the specimen about the metacarpo-phalangeal joints, calculated from the
observed frequencies. Each value of frequency and of rebound resilience is a mean calculated
from measurements of 3-10 trains of oscillations. Specimens A and B were taken from a sheep
of mass 35 kg and the others from young sheep of mass about 20 kg.)
Frequency
Hz
Ternperature
Mass (kg) ...
1-94
A*
B
C
20
7-2
6-2
23
23
23
24
31
6-9
64
69
6-4
69
36
—
6-6
—
20
D
E
F
G
H
Mean
502
Torsional stiffness
(N m red"1)
i-< )4
IO56
5-3
5i
52
55
5'4
5-1
49
—
—
—
—
—
28
46
49
—
—
—
15
13
19
16
502
41
26
24
30
25
23
27
24
—
15
17
18
—
Rebound resilience
10-56
—
—
—
—
37
39
—
—
—
1 94
5-02
10-56
—
0-54
0-67
0-55
o-6i
o-68
0-47
0-63
o-6i
0-59
066
0-57
o-66
0-63
062
059
—
—
—
—
072
0-71
—
—
059
062
0-72
• Specimens.
-
fa)
r
0-4 -
40 Static
-
Z
/
iS
al stiffn
/
0-2
/
L
0
20
c0
1
•
rsi
Ang
5.
Dynamic
1E
J
_
(ft)
1
1
1
1
4
2
Moment (N
1
|
1
6
0
1
0
Tl)
1
1
1
2
4
Moment (N m)
1
1
6
Fig. 3. (a) A graph of angular displacement of the metacarpo-phalangeal joints of specimen F
(Table 1) against the moment applied in static tests. (6) Graphs of torsional stiffness against
applied moment for the same specimen. Both static values calculated from the gradients of
tangents to (a) and dynamic values taken from Table 1 are shown.
than the corresponding values of dynamic stiffness for the same specimen. The
experiments at 37 °C and 20 °C showed no significant effect of temperature on
stiffness.
DISCUSSION
We have to consider to what extent the properties which have been observed are
properties of the tendons, and to what extent they may be properties of the muscle
fibres. The loads which were applied in the experiments extended the metacarpophalangeal joints through 0-2-0-5 rad (Fig. 3 a). We estimate from X-radiographs and
by manipulating dissections that the moment arms about these joints of the tendons
of the interosseous muscle are about 10 mm, so 0-5 rad extension of the joint requires
5 mm extension of the muscle. It seems inconceivable that the muscle fibres, which
80
W. G. CUMING, R. McN.
ALEXANDER AND A. S. JAYES
are only about i mm long, could stretch by more than a small fraction of this: most
of the observed extension must be due to extension of the tendons.
Another rough calculation supports this view. The area of tendon in any crosssection of the muscle is about 20 mm2 and the length of the muscle is about 10 cm.
Hence if only the tendons stretch and if Young's modulus for tendon is io9 N m~a
(Matthews & Ellis, 1968), the tensile stiffness of the muscle is about 2 x io6 N m"1.
If the moment arm about the joint is 10 mm, the torsional stiffness of the joint
should be about 20 N m rad"1, or a little more if the sheath contributes appreciably
to the stiffness. This is in the range of observed values (Fig. 36).
Consider next whether the moments and frequencies involved in the experiments
are appropriate to experiments designed to throw light on locomotion. Jayes &
Alexander (1978) recorded forces exerted by a 45 kg sheep crossing a force platform.
It can be shown from their records and films that in a typical example of trotting at
1-4 m s-1, the force on a fore foot exerted moments up to 12 N m about the metacarpophalangeal joints. In an example of cantering at 2-4 m s"1, moments up to 23 N m
occurred. Most of the experiments reported in Table 1 were on legs from young sheep
of mass about 20 kg so the moments which would have acted about their metacarpophalangeal joints in similar gaits would have been much smaller, probably 4-5 N m
and 8-10 N m. These moments would presumably have been counteracted partly by
the digital flexor muscles and only partly by the interosseous muscle and its sheath.
The moments applied in the experiments ranged from 1-3 to 7 N m, and had to be
counteracted by the interosseous muscle and sheath alone. They were probably
similar in magnitude to the moments which the muscle and sheath have to counteract
in locomotion.
Jayes & Alexander (1978) found that the duration of contact with the ground of the
forefoot of their sheep ranged from o-6 s in a slow walk (at 0-7 m s -1 ) to 0-2 s in
a slow gallop (at 3-4 m s -1 ). It would presumably have been slightly less for smaller
sheep in similar gaits (see Pennycuick, 1975). Jayes & Alexander made no observations
of sheep galloping fast. Alexander, Langman & Jayes (1977) observed contact
durations down to 0-05 s for 20 kg gazelles galloping fast but we do not suppose sheep
could achieve this. The interosseous muscle is stretched and recoils once while the
foot is on the ground so the duration of contact corresponds to half a cycle of oscillation in our experiments. The frequencies of oscillation ranged from 4-6 to 7-2 Hz
so the duration of a half cycle was 0-07-0-11 s. This is possibly a little too short to be
entirely realistic as simulation of locomotion.
The experiments at elevated temperatures were carried out because the temperature
of the muscle in living sheep is presumably well above ordinary room temperatures.
A sheep kept indoors on wire mesh at an ambient temperature of 9 °C was found
to have an interdigital temperature of 37 °C. However, differences of temperature in
the range 20-37 °C na<* n o significant effect on the results of our experiments.
The rebound resiliences of about 0-62 which have been measured in experiments
are far lower than the values for resilin, abductin and feather shafts (over 0-9 in each
case, Jensen & Weis-Fogh, 1962; Alexander, 1966; Pennycuick & Lock, 1976). They
are nevertheless high enough, if they apply to tendons in general, to make substantial
savings of energy possible in locomotion. We are not aware of any previous determinations of the rebound resilience of tendons.
Rebound resilience of tendons in the feet of sheep (Ovis aries)
81
A weakness of this investigation is that the effect of the muscle fibres on our results
is unknown, though it is thought likely to be small since the fibres are so short.
Further experiments should be performed with isolated tendons and might yield
higher values of rebound resilience. A strength of the investigation is that it has been
possible to make the measurements with the metacarpo-phalangeal joints and the skin
covering them intact: any energy losses caused in life by joint friction, skin viscosity
etc. will have occurred also in our experiments.
We are grateful to Mr K. G. Towers for supplying legs and Dr C. G. Scanes for
help in measuring interdigital temperature. This work is part of a programme of
research supported by a grant to R.McN.A. from the Science Research Council.
REFERENCES
ALEXANDER, R. M C N . (1966). Rubber-like properties of the inner hinge-ligament of Pectinidae. J. exp.
Biol. 44, 119—130.
ALEXANDER, R. MCN. (1968). Animal Mechanics. London: Sidgwick & Jackson.
ALEXANDER, R. M C N . (1977). Mechanics and scaling of terrestrial locomotion. In Scale Effects in Animal
Locomotion (ed. T. J. Pedley), 93-110. London: Academic Press.
ALEXANDER, R. M C N . & BENNET-CLARK, H. C. (1977). Storage of elastic strain energy in muscles and
other tissues. Nature, Lond. 265, 114-117.
ALEXANDER, R. MCN., LANGMAN, V. A. & JAYES, A. S. (1977). Fast locomotion of some African
ungulates. J. Zool., Lond. 183, 291-300.
ALEXANDER, R. M C N . & VERNON, A. (1975). The mechanics of hopping by kangaroos (Macropodidae).
J. Zool, Lond. 177, 265-303.
CAMP, C. L. & SMITH, N. (1942). Phylogeny and functions of the digital ligaments of the horse.
Mem. Univ. California 13, 69-124.
CAVAGNA, G. A., SAIBENE, F. & MARGARIA, R. (1964). Mechanical work in running. J. appl. Physiol.
19, 240-256.
JAYES, A. S. & ALEXANDER, R. M C N . (1978). Mechanics of locomotion of dogs (Canis familaris) and
sheep (Ovis aries). (In preparation.)
JENSEN, M. & WEIS-FOGH, T. (1962). Biology and physics of locust flight. V. Strength and elasticity of
locust cuticle. Phil. Trans. R. Soc. Lond. B 245, 137-169.
MATTHEWS, L. S. & ELLIS, D. (1968). Viscoelastic properties of cat tendon: effects of time after death
and preservation by freezing. J. Biomechan. 1, 65-71.
PENNYCUICK, C. J. (1975). On the running of the gnu (Connochaetes taiirinus) and other animals. J. exp.
Biol. 63, 775-799PENNYCUICK, C. J. & LOCK, A. (1976). Elastic energy storage in primary feather shafts. J. exp. Biol.
64, 677-689.