Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
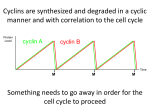
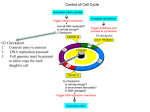
NEW TERMS for Biol 201: CELL CYCLE AND ITS CHECKPOINTS TO ENTER INTO MITOSIS: MPF (mitosis promoting factors): An activity that drives the cell into mitosis and holds it there is not an enzyme or a protein itself, it is just the collection of enzymes and proteins that do drive the cells into mitosis made up of cyclin B and cdc2 (a yeast cdk) (is a heterodimer of cyclin:cdk) MPF also activates the molecules that will get the cell out of mitosis too (APC complex) Entering and leaving mitosis is regulated by phosphorylations of different targets by MPF Cyclin B: a protein that is one of the MPFs (mitosis promoting factors) and that is necessary and sufficient for the cell to enter mitosis. The cell cannot finish mitosis unless cyclin B is degraded (not silenced, but completely degraded from the cell) There is a direct correlation between the amounts of cyclin B in the cell and MPF activity in the cell. When there is more cyclin B, there is more MPF activity; when there is less cyclin B, there is less MPF activity. Cyclin B and MPF peaks also correlated with the entry of the cell into mitosis, and when cyclin B levels and MPF levels drop, the cell leaves mitosis. Cyclin-dependent kinase (cdk).. A kinase that must bind to a cyclin in order to be activated. They are the key kinases that run the cell cycle. Cdc2.. Yeast cdk; homologous with vertebrate cdks. Theses cdks are highly conserved proteins through all species, from yeast to humans. MPF.. cyclin:cdk heterodimer. These heterodimeric kinases drive the cell cycle Wee1.. cdk inhibiting kinase: phosphorylates the cyclin:cdk heterodimer to keep it in an inactive state. Allows us to build up the levels of cyclin:cdk dimers without driving the cell into mitosis just yet. CAK.. Cdk Activating Kinase: phosphorylates cyclin:cdk heterodimer (MPF) after Wee1, adding an activating phosphate to the MPF molecule Cdc25.. phosphatase; reverses wee1 inhibition: removes the inhibitory phosphates that were added on to the MPF molecule by Wee1. == complete activation of the MPF molecule such that now, MPF can drive the cell into mitosis. TO LEAVE MITOSIS FOR ANAPHASE AND TELOPHASE: need protein degradation Ubiquitin.. A small molecule that is bound in a chain to a protein to be targeted for degradation. A molecule that acts like a marker for degradation by the proteasome. Whichever proteins have a chain of ubiquitin on them will be targeted by the proteasome. Ubiquitin ligase.. An enzyme that links ubiquitins to target proteins. This is the enzyme that sticks the ubiquitin molecules onto the proteins that need to be degraded. ONLY PROTEINS THAT CONTAIN A SPECIAL AMINO ACID SEQUENCE CALLED THE “DESTRUCTION BOX” ARE TARGETTED BY UBIQUITIN LIGASE. (cyclin B’s amino acid sequence thus contains a destruction box) Destruction Box (special amino acids in the sequence of certain proteins only)..Recognition site on a protein for ubiquitin ligase, which will then attach a chain of ubiquitins to the protein whose’s sequence contains the destruction box. APC.. Anaphase Promoting Complex; a ubiquitin ligase. Will take the cell out of mitosis by ubiquitinating various proteins that keep the cell in mitosis. Partially activated by MPF phosphorylation during prometaphase. Targetted specifically by Cdc20 and Cdh1for full activation of the APC. Cdc20.. An accessory molecule that targets APC to securin. Complexes to APC during metaphase and then directs APC to ubiqitinate securin for degradation. Leaves after it’s job is done. Cohesin.. a molecular complex that holds mitotic chromosomes together, thereby preventing anaphase. Holds the kinetochores of the sister chromatids together thus preventing anaphase. Is degraded by separase enzyme. Securin.. An molecule that inhibits separase. Binds to separase to prevent separase from degrading the cohesion proteins that keep the kinetochores of the sister chromatids together. Separase.. An enzyme that degrades cohesion and allows the sister chromatids to separate, thus promoting anaphase. Is inactivated by securin until the APC-Cdc20 complex ubiquitates securin. Cdh1.. An accessory molecule that targets APC to mitotic cyclin (B-type cyclins) and cdc20. Held inactive by inhibitory phosphates, but the Cdc14 phosphotase dephosphorylates Cdh1, thus activating it. Active Cdh1 binds to APC, which then targets cyclin B and Cdc20 for degradation, such that there are no more activating kinases left in the cell. Thus, all the proteins that were kept active by cyclin B will slowly become dephosphorylated by various phosphotases. == this ensures mitotic exit. Cdh1 is inactive when it is phosphorylated (it is phosphorylated for most of the cell cycle) Cdc14.. A phosphatase that becomes activated during anaphase and activates cdh1 by removing the inhibitory phosphates on Cdh1. APC bound to Cdc20 == ubiquitinates securin for degradation APC bound to Cdh1 == ubiquitinates mitotic cyclins (such as cyclin B) and Cdc20 HORMONES AND INTRACELLULAR COMMUNICATION The restriction point in mammals and START in yeast are the points in late G1 when a cell becomes committed to continuing through the cell cycle. == the restriction point in mammals and the START point in yeast are the same thing == high conservation between species == once the yeast cell or the mammal cells pass these points, they become commited to division == will divide no matter what (even if you starve a yeast cell after it has passed the START point, it will still undergo division) If a yeast cell is starved it will arrest in G1 until conditions improve. However, if it is starved after it has crossed START, it will continue through the cell cycle until it reaches G1 at which point it will arrest. B-Type cyclins: These are the S-phase and M-phase cyclins that contain a destruction box and so are destroyed late in mitosis. They are the cyclins whose transcription are driven by the G1 cyclins (the early and mid G1 cyclins drive the expression of S-phase and M-phase cyclins. The S-phase and M-phase cyclins are B-type cyclins) All cyclins that contain destruction boxes are B-type cyclins and are targets for degradation by ubiquitin ligase. APC can break all B-type cyclins down: therefore, APC can break down both S-phase and M-phase cyclins. S-phase cyclin: When it is first produced, it is complexed with a cdk to for the SPF (S-phase promoting factor). However, SPF is immediately complexed with a kinase inhibitor called “Sic1” and so SPF is inactive. SPF is only active once Sic1 becomes phosphorylated enough by G1 kinases to be targeted for degradation late in G1. Once Sic1is degraded, a large amount of SPF now becomes active and can now drive the cell into the Sphase. Sic1: The inhibitor that binds and inactivates the S-phase cyclin:cdk heterodimer. Is slowly phosphorylated by G1 cyclins and is degraded late in G1. Its degradation allows SPF to become active and allows the cell to enter into S-phase. ORC (Origin recognition complex): A multimolecular complex that bind to the origins of replication throughout the cell cycle. The origin of replication complexes (ORCs) are licensed in early G1 with the formation of the pre-replication complex. ORCs are fired up during S-phase and DNA replication occurs. ORCs are bound to the origins of replication and they serve as the docking site for many of the pre-replication complex components. When SPF levels are high, the ORCs are phosphorylated and they disassociate from the origin. Once the origins have been replicated, the ORCs right away bind back to the origins (so the origins are more or less never free from the ORCs) Pre-replication complex: The ORC plus a variety of other proteins that bind to it and prepare the origins for DNA replication. Some of these proteins are actual components of the replication machinery (ex: helicase) and other components are inhibitors of DNA synthesis. As long as the components of the pre-replication complex are not phosphorylated by high SPF activity, replication cannot begin. When the S-phase dimers become active, components of the prereplication are phosphorylated. Some of the components of the replication machinery are activated in this fashion (Mcm helicase) while inhibitory molecules are caused to dissociate freeing the origins from inhibition. As long as there are kinases in the cell, you cannot build another prereplication complex. You need to dephosphorylate the components on the complex to actually make another pre-replication complex, and this only happens in early G1. This means that you can only replicate DNA once per cell cycle. ATM: A kinase that is activated by DNA damage or a failure to complete DNA replication. Is a sensor for DNA damage. Activates 2 proteins: p53 and chk kinases. P53: The guardian of the genome. Is a transcription factor that directs transcription of p21. Activated by ATM when DNA is damaged. It directs the transcription of the cdk inhibitor, p21, which then causes G1 arrest. If the DNA damage cannot be fixed, p53 also mediates cell apoptosis. CHK kinases (checkpoint kinases): Activated by ATM when DNA is damaged. It phophorylates cdc25 homologues and inactivates them. This causes G2 and S arrests. CHK kinases inactivate cdc25 homologs (which activate MPF by phosphorylation).