Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
AN ABSTRACT OF THE DISSERTATION OF
Mollie Kim Manier for the degree of Doctor of Philosophy in Zoology presented on
January 28, 2005.
Title: Population Genetics, Ecology and Evolution of a Vertebrate Metacommunity
Abstract approved:
Redacted for Privacy
Population genetic structure is widespread in many organisms and can be found at
small spatial scales. Fine-scale differentiation is the result of ecological and
evolutionary processes working together to produce an overall pattern, but the relative
importance of these factors in population differentiation is poorly understood. The
goals of my research were to describe patterns of population genetic differentiation
and to identify ecological and evolutionary factors important for population
divergence. To this end, I investigated several aspects of genetic differentiation for
three vertebrates in northern California. The focal species were the terrestrial garter
snake (Thamnophis elegans) and the common garter snake (Thamnophis sirtalis) that
occupy a series of ponds, lakes and flooded meadows in northern California. I found
significant genetic differentiation and isolation by distance, as well as correlated
patterns of pairwise divergence in both species. Independent estimates of effective
population size and bi-directional migration rates also uncovered source-sink
dynamics in both species that suggest frequent extinction-recolonization events within
a metapopulation context. The generality of source-sink dynamics for an ecologically
similar species within the same ecosystem was explored using a third species, B.
boreas. I also identified ecological correlates of several population genetic parameters
for all three species. Although Fi's were similar,
B. boreas
had larger effective
population sizes, lower migration rates, lacked source-sink dynamics, and appeared to
be in migration-drift equilibrium, indicative of a temporally stable population
structure. A clustering analysis identified a series of block faults as a common barrier
to dispersal for both garter snakes, and ecological correlates were found to be more
similar among response variables than within species. I then compared degree of
genetic differentiation at quantitative traits with that at neutral markers to infer
strength of selection and adaptive divergence between two ecotypes of 7'.
elegans.
Selection on most traits was relatively weak, but strong diversifying selection was
found for background coloration, total number of ventral scales and number of
infralabials. Overall, my research documented ecological and evolutionary processes
associated with population differentiation in a metacommunity and respresents an
important contribution toward the unification of ecology and evolutionary biology.
©Copyright by Mollie Kim Manier
January 28, 2005
All Rights Reserved
Population Genetics, Ecology and Evolution of a Vertebrate Metacommunity
by
Mollie Kim Manier
A DISSERTATION
submitted to
Oregon State University
in partial fulfillment of
the requirements for the
degree of
Doctor of Philosophy
Presented January 28, 2005
Commencement June 2005
Doctor of Philosophy dissertation of Mollie Kim Manier presented on January 28,
2005.
APPROVED:
Redacted for Privacy
Redacted for Privacy
Chair oJtl
Departmeit of Zoology
Redacted for Privacy
Dean of the G
iate School
I understand that my dissertation will become part of the permanent collection of
Oregon State University libraries. My signature below authorizes release of
my dissertation to any reader upon request.
Redacted for privacy
Mollie Kim Manier, Author
ACKNOWLEDGEMENTS
Many entities should be credited with seeing this project to its completion. Stevan
Arnold, my major advisor, has dedicated tireless hours to the design of this research,
collection of tissue samples, assisting with statistical analysis and editing countless
renditions of proposals, manuscripts and grants, not to mention this thesis. He has
been available and supportive throughout and has been everything I could ask for in an
advisor. I would also like to express gratitude to my committee members, Ross
Kiester, Fred Ramsey, Peter McEvoy, and especially Mike Blouin, who has imparted a
great degree of knowledge and expertise in population genetics methods and theory
over the years.
Funding for this research was provided by an EPA STAR (Science to Achieve
Results) Fellowship (U-91552801-5), NSF GK-12 Teaching Fellowship, NSF DDIG
(Doctoral Dissertation Improvement Grant; DEB-0309017), NSF grant DEB-9903934
(to SJA), and NSF grant DEB-1234567 (to SJA and AMB). I am also most grateful
for teaching opportunities received in the Biology Department from Joe Beatty.
A project of this magnitude could not be successful without a lot of assistance.
Many thanks to the dozens of Eagle Lake Snake Campers who have swelled my
sample sizes each field season. They include Steve Arnold, Jonathan Feder, Daniel
Hanken, Anne Bronikowski, Tim Knight, Karen and Jim Manier, Morgan Manier,
Laura Arnold, Brittany Barker, Al Bennett, Man Bennett, Rudi Berkeihamer, Ariane
Cease, Tim Clarke, Maggie Clarke, Russ Clarke, the Drummond family (Hugh,
Sylvia, Ian and Allen), Suzanne Estes, Shanie Holman, Cindy Houck, Lynne Houck,
Ross Kiester, Juliette Kiester, Lucy Kiester, Robin Lillihie, Minott Kerr, the Leonard
family (Bill, Vicky, Megan and Nick), Kathy Linton, David Scott, Weezie Mead,
Dede Olson, Shannon McDowell, Matt McDowell, Mike Pfrender, Melodie Rudenko,
Sharon Marks, Hart Welsh, M. Layon, K. Lacy, T. & T. Bittner, R. Boleso, and T.
Pappas. Special thanks go to Judith Scan, who flew out from Harvard to assist me for
two weeks in 2001, Tim Knight, and my mom, Karen Manier. Susan Chappell and
Teresa Pustejovsky, field biologists with Lassen National Forest, were also very
helpful with information on toad occurrences and breeding information as well as
access to GIS data.
I am especially grateful for Eric Simandle at University of Nevada, Reno, who
generously shared both his microsatellite primers for Bufo
boreas
for the Pikes Point population. I could not have included B.
as well as his data
boreas
in this dissertation
without his assistance, and he has allowed me to add an extra dimension to the Eagle
Lake landscape that would otherwise be sorely lacking. I will long be in his debt.
I was assisted in the lab by Kirk Wintennute, Kirsten Freed, Ingrid Albrecht
and Melodie Rudenko. Numerous undergraduate workers spent long tedious hours
counting scales, and the datasets were proofed and prepared by Sara Wynveen, Kristen
Rodd and Craig Seylor. Valuable technical and statistical support was provided by
Mike Pfrender, Eric Hoffinan, Catherine Palmer, Adam Jones, Anne Bronikowski,
Mike Westphal, Peter Beerli, Pierre Legendre, Richard Watts, Charles Criscione and
Kirsten Monsen. I am also indebted to the Zoology office staff, especially Sarah Cain,
Traci Durrell-Khalife, Tara Bevandich, Mary Crafts and Virginia Veach; the Cosine
helpdesk, especially Jerod Sapp; Naoki Kitabay and Caprice Rosato in the Central
Services Lab of the Center for Gene Research and Biotechnology; Scott Givan with
the Genetic Analysis Computer; and Joan Rowe at the Nevada Genomics Center.
I would also like to thank Houck/Arnold lab and Blouin lab members for years
of good conversation and camaraderie: Catherine Palmer, Eric Hoffman, Doug
DeGross, Richard Watts, Mike Westphal, Charles Criscione, Leslie Dyal, Erika
Adams, Jerod Sapp, Amy Picard, and Karen Keimnec.
I am forever indebted to my parents, Jim and Karen Manier, for their undying
support in my endeavors. They have provided financial and field assistance as well as
child care both at home and abroad (especially my mom), for which I am eternally
grateful. My brothers, Dan and Greg, have also shown immense support and interest
in my travails, and I am especially grateful to Dan for his assistance with GIS, which I
didn't end up using (thankfully!). I would also like to express a deep gratitude to Tim
Knight for his fantastic intellectual, emotional, co-parental, and culinary support
throughout this process. He's the best ever. And finally, I would like to thank my
children, Morgan and Rowan, for making it all worthwhile.
TABLE OF CONTENTS
Page
Chapter 1.
Chapter 2.
General introduction
Population genetic analysis identifies source-sink dynamics for
two garter snake species (Thamnophis elegans and T. sirtalis)...
Abstract
Introduction
Methods
Results
Discussion
References
Chapter 3.
Ecological correlates of population genetic structure: a
comparative approach using a vertebrate metacommunity
Abstract
Introduction
Methods
Results
Discussion
References
Chapter 4.
Adaptive divergence between ecotypes of the terrestrial garter
snake, Thamnophis elegans
Abstract
Introduction
Methods
Results
Discussion
References
Chapter 5.
Bibliography
General conclusion and future directions
1
6
7
8
13
21
47
55
60
61
63
66
78
98
106
110
111
113
117
124
138
141
145
148
LIST OF FIGURES
Figure
Page
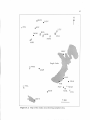
Map of study area showing sampled sites. SVP was a site sampled by
Kephart (1981)
10
2.2
Isolation-by-distance plots for A. T elegans and B. T sirtalis.
24
2.3
Map of the study area showing relative effective population sizes and
directions and rates of migration for T. elegans. Circle size and arrow
width are proportional to effective population size and migration rate,
respectively
38
Map of the study area showing relative effective population sizes and
directions and rates of migration for T. sirtalis. Conventions are as in
Fig. 2.3
45
3.1
Map of the study area showing sampled sites.
65
3.2
Genetic distance (Fs1/(1-Fs1)) as a function of log geographic distance
for B. boreas.
75
Map showing clustering pattern for T. elegans populations. Thickness of
borders indicates to grouping sequence, with thicker lines corresponding
to groups defined earlier in the analysis
86
Map showing clustering pattern for T. sirtalis populations. Conventions
as in Figure 3.3.
88
Map showing clustering pattern for B. boreas populations. Conventions
as in Figure 3.3
89
Scatterplots for pairs of variables used in stepwise regression of effective
population size
91
Scatterplots for pairs of variables used in stepwise regression of migration
rate.
92
Scatterplots for pairs of variables used in stepwise regression of genetic
distance.
93
2.1
2.4
3.3
3.4
3.5
3.6
3.7
3.8
LIST OF FIGURES (Continued)
Figure
4.1
4.2
4.3
4.4
4.5
Page
Map showing locations of meadow (MCY, PAP, NML, MAH) and
lakeshore (P1K, GAL) sites. An additional lakeshore site, WDC, was
used to generate heritabilities for scale counts
118
QST values for males and females for all traits, compared with FST (line)
A. among populations and B. between ecotypes
132
Frequency histograms showing distribution of dorsal stripe color for
males and females in each population. Populations are shown in order
from farthest to closest to Eagle Lake
135
Frequency histograms showing distribution of lateral stripe color for
males and females in each population. Conventions as for Fig. 4.3
136
Frequency histograms showing distribution of background color for
males and females in each population. Conventions as for Fig. 4.3
137
LIST OF TABLES
Table
Page
Names and abbreviations of study sites and their latitude and longitude
in decimal degrees, sample size (N), average observed heterozygosity
(H0), average expected heterozygosity (He) and average number of
alleles per locus (Na) for each species
14
Comparison of the genetic diversity found at all microsatellite loci for
each species averaged over all populations
16
Genetic characteristics of three microsatellite primers that amplify in
both T. elegans and T. sirtalis
18
Pairwise FST values for T. elegans populations below the diagonal.
Associated P-values above the diagonal, obtained after 3000
permutations
26
Pairwise FST values for T. sirtalis populations below the diagonal.
Associated P-values above the diagonal, obtained after 3000
permutations
28
2.6
Estimates of migration rate as measured by
29
2.7
Estimates of migration rate
for T. sirtalis.
2.1
2.2
2.3
2.4
2.5
2.8
4Nem for T. elegans.
(4Nem) and effective population size (4Nu)
41
Comparison of mark-recapture and microsatellite estimates of effective
population size
52
2.9
Comparison of direct and indirect estimates of migration rate.
53
3.1
Names, abbreviations, latitude, longitude, perimeter (km), type (M =
meadow, L = lake, LS = lakeshore), elevation (m), sampling effort and
sample sizes of study sites and populations.
70
Abbreviations and descriptions of variables used in the multiple
regression analysis
72
Geographic distance matrix of pairwise distances between populations,
measured in km.
74
3.2
3.3
LIST OF TABLES (Continued)
Table
3.4
Page
Full models used in each of nine stepwise regression analyses testing for
ecological effects on effective population size, migration rate and genetic
distance for each species.
76
Comparison of the genetic diversity found at all micro satellite loci for B.
boreas averaged over all populations.
79
Study populations of B. boreas and their sample sizes (N), average
observed heterozygosity (H0), average expected heterozygosity (He) and
average number of alleles per locus (Na)
80
Pairwise FST values for B. boreas populations below the diagonal.
Associated P-values above the diagonal, obtained after 3000
permutations
81
Estimates of migration rate (Al) and effective population size (6)) for B.
boreas.
82
3.9
Genetic structure analyses for B. boreas, T. elegans and T sirtalis.
84
3.10
Resuts of nine multiple regression analyses testing for effects of habitat
on effective population size
migration rate (m) and genetic distance
(FST/1-FsT) for each species
94
3.5
3.6
3.7
3.8
(Ne),
4.1
Names, abbreviations, ecotype and sample sizes of study populations.
119
4.2
Tests of sexual dimorphism for scale counts
125
4.3
Sample sizes, means and standard errors of traits for males and females
in each population.
126
Sample sizes, means and standard errors of traits for males and females
in lakeshore and meadow habitats.
128
Tests for differences between ecotypes and among populations within
ecotypes for each trait in males and females
130
Heritabilities of traits and global
ecotypes for males and females
134
4.4
4.5
4.6
QST among
and color scores.
populations and between
POPULATION GENETICS, ECOLOGY AND EVOLUTION OF A VERTEBRATE
METACOMMUNITY
CHAPTER 1
GENERAL INTRODUCTION
Population genetic structure refers to the spatial configuration and connectivity of
local breeding units (Wright 1969). Such structure can be found even at small spatial
scales. Fine-scale differentiation has been found in diverse taxa, from sessile
organisms such as plants (e.g., Vekemans and Hardy 2004 and refs. cited therein) to
marine species with a high potential for dispersal (e.g., Pampoulie et al. 2004, Zardoya
et al. 2004 and refs. cited therein). Population genetic differentiation via random
genetic drift or adaptive divergence in response to local selection and can have a
significant impact on the microevolutionary dynamics and can be an evolutionary
precursor to speciation. Levels of divergence, in turn, are determined by numerous
ecological and evolutionary factors operating on multiple spatial and temporal scales.
Spatial distribution is one ecological factor that can have a significant effect on
population genetic structure. Spatial heterogeneity can be caused by a number of
factors including aggregative social behavior, outbreeding avoidance, limited
dispersal, environmental stochasticity, and habitat heterogeneity. Wright (1922, 1951)
introduced F-statistics to describe differentiation in a system of small, identical
subpopulations and to describe the level of inbreeding within demes. The most
commonly used statistic, FST, describes a population's position along a continuum that
2
ranges from panmixia to total subdivision with no gene flow among subpopulations.
The value of FST can be related to effective population size (Ne) and migration rate (m)
according to the equation, FST
= 11(1
+ 4Nem). This relationship describes the degree
of genetic subdivision due to limited migration according to the island model of spatial
structure.
Population size is another ecological variable that can affect differentiation.
Small populations will diverge more quickly than large populations due to random
genetic drift alone. Effective population size, which describes the rate at which
genetic variability is lost due to genetic drift (Wright 1931), is frequently correlated
with and smaller than census size (Nunney and Elam 1994, Frankham 1995). Any
ecological phenomenon that influences population size, such as resource availability,
competition or predation, can therefore affect levels of population differentiation.
Ecological variables that can decrease Ne relative include nonrandom mating, variation
in reproductive output and factors associated with a decrease in census size such as
competition, predation, and habitat degradation (for review, see Caballero 1994).
Dispersal is a third ecological phenomenon with direct consequences for
population differentiation, in particular for its effect on the first factor discussed,
spatial structure. Much attention has focused on dispersal as a major factor
influencing genetic differentiation of populations, because it has an obvious impact on
levels of interdemic gene flow. Limited dispersal ability leading to isolation by
distance (Wright 1946) is also easy to detect by regressing genetic distance on
geographic distance (Rousset 1997). The isolation-by-distance scatterplot for such a
3
regression typically consists of a cloud of points with a positive slope, but there is
never a perfect relationship between genetic distance and geographic distance. For a
set of population pairs of any given geographic distance, there is some range of
genetic distances between them. Factors other than dispersal must be invoked to
explain this variation. Physical barriers to gene flow or corridors of dispersal can
facilitate (Gamier et al. 2004) or impede (Coulon et al. 2004) genetic differentiation
independent of geographic distance. Migration and consequent gene flow may also be
influenced by habitat characteristics (Lurz et al. 1997, Matter and Roland 2002,
Donahue et al. 2003), conspecific (Negro et al. 1997, Moksnes 2004) and interspecific
interactions (Hakkarainen et al. 2001, Heg et al. 2004), and the spatial distribution of
these ecological attributes on the landscape.
My dissertation is partitioned into three sections that explore the population
genetics, ecology and evolution of a vertebrate metacommunity in Lassen Co.,
California. I use microsatellite markers to investigate the population genetic structure
of two species, the terrestrial garter snake (Thamnophis elegans) and the common
garter snake (T sirtalis), that coexist on a common landscape. I test the hypothesis
that similarities in ecology and evolutionary history between the two species will
produce similar patterns of population differentiation (Chapter 2). I then identify
ecological and evolutionary processes important in shaping the observed patterns of
snake population genetics by using a comparative approach with a third species, the
western toad (Bufo boreas; Chapter 3). Finally, I explore evolutionary processes that
shape adaptive divergence in 7". elegans, by comparing levels of population
4
differentiation at quantitative traits and neutral molecular markers (Chapter 4).
Conclusions of all three data chapters are summarized in Chapter 5, and future
research directions are discussed.
REFERENCES
Caballero, A. 1994. Developments in the prediction of effective population size.
Heredity 73: 657-679.
Coulon, A., J.F. Cosson, J.M. Angibault, B. Cargnelutti, M. Galan, N. Morellet, B.
Petit, S. Aulagnier and A.J.M. Hewison. 2004. Landscape connectivity
influences gene flow in a roe deer population inhabiting a fragmented
landscape: an individual-based approach. Molecular Ecology 13:2841-2850.
Donahue, M.J., M. Holyoak and C. Feng. 2003. Patterns of dispersal and dynamics
among habitat patches varying in quality. American Naturalist 162:302-3 17.
Frankham, R. 1995. Effective population size/adult population size ratios in wildlife:
a review. Genetical Research 66:95-107.
Gamier, S., P. Alibert, P. Audiot, B. Prieur and J.-Y. Rasplus. 2004. Isolation by
distance and sharp discontinuities in gene frequencies: implications for the
phylogeography of an alpine insect species, Cara bus solieri. Molecular
Ecology 13:1883-1897.
Hakkarainen, H., P. Ilmonen and V. Koivunen. 2001. Experimental increase of
predation risk induces breeding dispersal of Tengmalm's owl. Oecologia
126:355-359.
Heg, D., Z. Bachar, L. Brouwer and M. Taborky. 2004. Predation risk is an
ecological constraint for helper dispersal in a cooperatively breeding cichlid.
Proceedings of the Royal Society of London Ser. B. 271:2367-2374.
Lurz, P.W.W., P.J. Garson and L.A. Wauters. 1997. Effects of temporal and spatial
variation in habitat quality on red squirrel dispersal behaviour. Animal
Behaviour 54:427-435.
Matter, S.F. and J. Roland. 2002. An experimental examination of the effects of
habitat quality on the dispersal and local abundance of the butterfly Parnassius
smintheus. Ecological Entomology 27:308-316.
Moksnes, P.-O. 2004. Interference competition for space in nursery habitats:
density-dependent effects on growth and dispersal in juvenile shore crabs
Carcinus maenas. Marine Ecology Progress Series 281:181-191.
Negro, J.J., F. Hiraldo and J.A. Doná.zar. 1997. Causes of natal dispersal in the lesser
kestrel: inbreeding avoidance or resource competition? Journal of Animal
Ecology 66:640-648.
Nunney, L. and D.R. Elam. 1994. Estimating the effective population size of
conserved populations. Conservation Biology 8:175-184.
5
Pampoulie, C., E.S. Gysels, G.E. Maes, B. Hellemans, V. Leentjes, A.G. Jones and
F.A.M. Volckaert. 2004. Evidence for fine-scale genetic structure and
estuarine colonisation in a potential high gene flow marine goby
(Pomatoschistus minutus). Heredity 92:434-445.
Rousset, F. 1997. Genetic differentiation and estimation of gene flow from FStatistics under isolation by distance. Genetics 145:1219-1228.
Vekemans, X. and O.J. Hardy. 2004. New insights from fine-scale spatial genetic
structure analyses in plant populations. Molecular Ecology 13:921-935.
Wright, S. 1922. Coefficients of inbreeding and relationship. American Naturalist 63 :556561.
Wright, S. 1931. Evolution in Mendelian populations. Genetics 16:97-159.
Wright, S. 1943. Isolation by distance. Genetics 28:114-138.
Wright, 5. 1951. The genetical structure of populations. Annals of Eugenics 15:323-354.
Wright, 5. 1969. The theory of gene frequencies, evolution and the genetics of populations.
Vol.2, Chicago: University of Chicago Press.
Zardoya, R., R. Castilho, C. Grande, L. Favre-Krey, S. Caetano, S. Marcato, G. Krey
and T. Patamello. 2004. Differential population structuring of two closely
related fish species, the mackerel (Scomber scombrus) and the chub mackerel
(Scomberfaponicus), in the Mediterranean Sea. Molecular Ecology 13:17851798.
POPULATION GENETIC ANALYSIS IDENTIFIES SOURCE-SINK DYNAMICS
FOR TWO SYMPATRIC GARTER SNAKE SPECIES (THAMNOPHIS
ELEGANS AND T. SIR TALIS)
CHAPTER 2
Mollie K. Manier and Stevan J. Arnold
This thesis chapter has been prepared for submission to the journal Molecular
Ecology.
7
ABSTRACT
Population genetic structure can be shaped by multiple ecological and evolutionary
factors, but the genetic consequences of these factors for multiple species inhabiting
the same environment remain unknown. We used microsatellite markers to examine
the population structures of two coexisting species of garter snake, Thamnop his
elegans and T. sirtalis, to determine if shared landscape and biology imposed similar
population genetic structures. These snakes inhabit a series of ponds, lakes and
flooded meadows in Lassen Co., California and tend to converge on prey type
wherever they coexist. Both garter snakes had comparable effective population sizes
and bi-directional migration rates (estimated using a maximum likelihood method
based on the coalescent) with low but significant levels of genetic differentiation (FST
= 0.025 for T. elegans and 0.035 for T sirtalis). Asymmetrical gene flow revealed
large source populations for both species as well as potential sinks, suggesting
frequent extinction-recolonization and metapopulation dynamics. In addition, we
found a significant correlation between their genetic structures based on both pairwise
FST'S
for shared populations (P = 0.009) and for bi-directional migration rates (P =
0.024). Possible ecological and evolutionary factors influencing similarities and
differences in genetic structure for the two species are discussed. Genetic measures of
effective population size and migration rates obtained in this study are also compared
with estimates obtained from mark-recapture data.
INTRODUCTION
Patchily distributed populations can become genetically distinct over time as a
consequence of random genetic drift and response to locally-varying selection. Gene
flow counters these two processes and acts as a homogenizing force that opposes
differentiation (Wright 1931). The net result of all three processes is some pattern of
population genetic structure. The degree to which a population experiences drift,
selection and gene flow depends on multiple ecological and evolutionary factors.
While similar species inhabiting a common landscape may encounter comparable
factors influencing population genetic differentiation, it is not known whether patterns
of genetic structure tend to evolve in parallel.
Although recent studies have reported population genetic structures for
multiple sympatric species (e.g., McMillen-Jackson and Bert 2003, Michels et al.
2003, Brede and Beebee 2004, Molbo et al. 2004, Zardoya et al. 2004), few studies
have statistically compared genetic structures. Usually, results for each species are
reported separately without statistical comparison. This qualitative approach arises
because multiple species are rarely sampled from the same sites (but see RUber et al.
2001, Brede and Beebee 2004). A general exception comes from studies of symbionts
(Anderson et al. 2004 and references cited therein), but in these cases, the comparison
species are usually so phylogenetically divergent that perceptions of and responses to
a common environment may be very different. Our research seeks to remedy this
situation by comparing two closely related species that coexist on the same landscape.
In this study, we compare patterns of population genetic differentiation at
microsatellite loci for two coexisting garter snake species, the terrestrial garter snake,
Thamnophis elegans, and the common garter snake, T. sirtalis. The study area is located
at and around Eagle Lake in Lassen County, California. The habitat is predominantly arid
sagebrush-yellow pine forest dotted with numerous permanent and semi-permanent lakes
and ponds as well as meadows that flood with snowmelt in the spring. The study system is
comprised of 22 such water bodies that vary in size, permanence and degree of isolation, all
occurring within 1050 km2 and ranging in elevation from 1500 to 2100 m (Fig. 2.1). Both
garter snakes have widespread distributions in the western United States and Canada
(Rossman et al. 1996, Stebbins 2003) and are abundant at the study area. The water
bodies provide habitat for the snakes' primary prey: amphibians, small fish and
leeches (Kephart 1982, Kephart and Arnold 1982). Garter snakes in temperate climates
hibernate during winter and emerge to mate in the immediate vicinity of those hibemacula
in the spring (Aleksiuk and Gregory 1974, Moore and Lindzey 1992, Whittier and Tokarz
1992). Because garter snake hibemacula are found at many of our study sites (Kephart
1981), we expected garter snake populations to be genetically structured about the lakes,
ponds and meadows that comprise our study system
Populations of T elegans in the study area are more common and tend to be larger
than those ofT sirtalis. Of the 22 water bodies included in this study, T elegans are found
at 20, and T sirtalis are found at 13. Overall, T elegans outnumbers I'. sirtalis at most sites
in the study area. Half of the study sites support both species, and only two (Feather Lake
10
BLW
0JCK
STF
LTC
.
BUL
GOR
Q
SUM
CLG
ASH
RKYI
Eagle Lake
PVM
jk)
AMP
GAL
. DNS
P1K
o FEA
0
PAP.
MAH
.
RON
NML
COL
SvP
Figure 2.1. Map of study area showing sampled sites. SVP was a site
sampled by Kephart (1981).
11
and Gordon Lake) are dominated almost exclusively by T sirtalis. We therefore expect
larger effective population sizes for T elegans, and a higher degree of population
differentiation for T sirtalis. Because T sirtalis occupies fewer sites, we also expect to find
a greater degree of isolation by distance in this species.
The diets of T elegans and T sirtalis at the study area tend to converge at sites
where they coexist in our study system (Kephart 1982, Kephart and Arnold 1982).
Specifically, both species capitalize on the explosive breeding events of\anurans, whose
larvae and metamorphs are a primary prey source when abundant. Both species will also
prey on fish (Rhinichthys osculus, Richardsonius egregius, Gila bicolor and Castomus
tahoensis) and leeches (Erpobdella spp.), although T elegans does so to a much greater
degree. Thamnophis elegans are better at controlling their buoyancy in water and can
therefore dive to actively hunt fish, whereas 7'. sirtalis can consume fish only when
they are easily caught in drying pools (Kephart 1981). This difference in ability to
catch fish may explain why T sirtalis are rarely found along the shoreline of Eagle
Lake, where fish are abundant but amphibian breeding is inconsistent from year to
year.
Recent phylogenetic reconstructions of garter snake relationships based on
mitochondrial sequence data suggest that T elegans and T sirtalis diverged from a
common ancestor in the Pliocene. Assuming a rate of sequence divergence of 1.3%
per million years based on agamid lizards (Macey et at. 1998), the two species
diverged around 6.9 million years ago (de Queiroz et al. 2002). The evolutionary
history of T. elegans and T. sirtalis at the study area begins after the Pleistocene, when
12
much of the currently suitable habitat in western North America was covered by
glaciers (Barnosky et al. 1987, Janzen et al. 2002). Glacial recession at the end of the
Pleistocene around 10 000 years ago allowed recolonization of parts of the western
United States by Thamnophis sirtalis from multiple refugia. Lassen Co. populations
of T sirtalis probably originated from the Great Basin or southern California (Janzen
et al. 2002). Overall, populations of T. elegans in western North America are much
more divergent (0.3-7%; Bronikowski and Arnold 2001) than those of 7'. sirtalis (0.3
to 0.6%; Bronikowski and Arnold 2001, Janzen et al. 2002), suggesting that T. sirtalis
is a relative newcomer to western North America.
The goal of this study was to use microsatellite markers to examine genetic
differentiation in two closely related species inhabiting a common landscape. We
investigated patterns of isolation-by-distance and estimated measures of genetic
differentiation, including bi-directional migration rates and effective population sizes.
We then estimated the correlation between species using two different measures of
genetic structure. Previous research in the study system estimated effective population
sizes for two 7'
elegans populations
and two T
sirtalis
populations based on mark-
recapture data (Kephart 1981). Furthermore, only six dispersal events out of over 800
recaptures were documented over distances less than 5 km, leading to the conclusion
that dispersal events were relatively rare. We compared these mark-recapture
estimates for effective population size and migration rate from Kephart (1981) to those
based on microsatellite data in an effort to compare direct and indirect methods of
estimating population genetic parameters.
13
METHODS
Sampling
We collected tissue samples from T. elegans and T. sirtalis in and around the Eagle
Lake basin in Lassen Co., California (Fig. 2.1). A total of 858 T. elegans from 20
populations and 433 T sirtalis from 13 populations were sampled (Table 2.1).
Overall, 22 sites were included in the study, 12 of which support both species. For
each snake, the tail tip or a 2x4-mm piece of ventral scale was taken and stored in
Drierite®, an anhydrous calcium sulfate desiccant.
Obtaining microsatellite data
A total of 11 primer sets for microsatellite markers were obtained from the literature
or developed de novo (Table 2.2). An enriched library was prepared according to the
method of Hamilton et al. (1999), and positive clones were screened based on the
method of Hoffman et al. (2003). In summary, whole genomic DNA was digested
with MspI, size-selected, ligated to oligo linkers and amplified with linker primers.
Four biotinylated oligo probes (dGACA4, dGATA4, dGGAT4 and dGGGA4) were
hybridized to linker DNA and selected using streptavidin magnetic particles
(Promega). Enriched DNA was then amplified using linker primers and purified using
a PCR purification kit (Qiagen). Linkers were removed and DNA was ligated into
pBluescript vector and transformed into Epicurian Coli XL1-Blue Supercompetent
14
Table 2.1. Names and abbreviations of study sites and their latitude and longitude in
decimal degrees, sample size (N), average observed heterozygosity (H0), average
expected heterozygosity (He) and average number of alleles per locus (Na) for each
species. Observed and expected heterozygosities were calculated without TS042 and
TEO5 1 B for T elegans and T. sirtalis, respectively. Study sites indicated with an
asterisk are informal names, not official geographic place names. SVP represents a
study site from Kephart (1981) that was not sampled for this study.
T. sirtalis
T. elegans
Site
AntelopeMountainPond*
Ashurst Lake
Blue Water
Bullard Lake
Cleghorn Reservoir
Colman Lake
Deans Meadow
Feather Lake
Gallatin Shoreline*
Gordon Lake
Jacks Lake
LittleCleghornReservoir
Mahogany Lake
McCoy Flat Reservoir
Nameless Meadow*
Papoose Meadows
Pikes Point
Pine Valley Meadow*
Rocky Point
RoneyCorral
Camp Stanford
Small Vernal Pools*
Summit Lake
Lat.
40.614
40.750
40.834
40.775
40.777
40.516
40.557
40.542
40.562
40.768
40.810
40.787
40.534
40.453
40.524
40.528
40.557
40.619
40.684
40.511
40.803
40.507
40.766
Abbrev.
Long.
-120.923
AMP
-120.965 ASH
-120.919
BLW
-120.901
BTJL
-120.804
CLG
-120.714
COL
-120.719
DNS
-121.018
FEA
-120.760
GAL
-120.882
GOR
-121.025
JKS
-120.794
LTC
-120.732 MAH
-120.940 MCY
-120.743 NML
-120.757
PAP
-120.784
P1K
-120.969
PVM
-120.757
RKY
-120.857
RON
-120.932
STF
-120.73 1
SVP
-120.839
SUM
N
30
30
27
67
44
24
26
H0
He
0.50 0.53
0.55 0.58
0.51 0.56
0.51 0.52
0.48 0.53
0.47 0.56
0.50 0.55
56
0.47 0.51
27
0.50
0.50
0.52
0.47
0.54
Na
N
5
19
H0
0.61
5
38
6
6
35
5
He
Na
5
0.58
0.64
0.59
0.55
0.57
7
27 0.59 0.57
27 0.47 0.52
24 0.53 0.63
7
42
0.51
0.56
7
26
0.53
0.60
7
29
0.49 0.56
7
83
0.57
0.61
9
7
5
5
5
7
5
0.54
0.59
0.53
5
0.51
0.53
4
6
24
45
0.54
0.52 0.54
0.52 0.51
0.47 0.58
0.45 0.50
0.54 0.54
4
6
29
32
0.47 0.60
0.50 0.59
6
7
27
0.50 0.52
5
22
0.58
5
18
91
16
29
140
48
70
19
0.51
5
5
5
4
5
5
0.61
16
Cells (Stratagene). Positive colonies were picked into sterile water, boiled and
sequenced at the Nevada Genomics Center at the University of Nevada, Reno.
Primers were designed using Oligo v. 6.0 (Rychlik 1998) and optimized with an MJ
Research Peltier gradient thermocycler.
Table 2.2. Comparison of the genetic diversity found at all microsatellite loci for
each species averaged over all populations. Total number of alleles, size range in
base pairs, observed (H0) and expected (He) heterozygosities and average number of
alleles per population (Na) are given. The numbers in parentheses represent numbers
of populations sampled.
Allele
Pops.
size
Total
out of
range
Locus
alleles
HWE
Reference
(bp)
T. elegans (20)
Ns.t10
12
Prosseretal. (1999)
120-144 0.53 0.58 5
0
Nsp2
6
139-160 0.59 0.65 4
0
Prosser et al. (1999)
Nsj.t3
19
129-193 0.84 0.88 11
0
Prosser et al. (1999)
Ns7
2
170-176 0.06 0.09 2
Prosser et al. (1999)
0
Nst8
5
129-161 0.41 0.41
3
0
Prosser et al. (1999)
TEO51B
4
96-104 0.11 0.14 2
this study
1
TSO1O
15
108-146 0.78 0.83 9
this study
0
TS042
19
148-194 0.34 0.79 9
17
this study
Ts2
16
118-149 0.63 0.69 8
0
McCracken et al. (1999)
Ts3
4
92-129 0.48 0.49 2
McCracken et al. (1999)
0
T. sirtalis (13)
H0
Ns10
He
Na
13
141-168
0.86
0.92
Nst2
1
138
---
---
Nsp3
20
0.69
0.73
0.70
0.30
0.38
11
9
2
145-195
174-200
120-128
145-198
108-124
0.74
Nspi
0.81
0.71
0.39
0.86
5
1
3
3
0
9
0
---
---
2
16
0
0
Nsj.t8
TSO1O
24
4
TS042
Ts3
3Ts
1
191
2
33
86-92
338-416
TEO51B
0.30 0.29
0.16 0.23
8
0
Prosseretal.(1999)
Prosser et al. (1999)
11
0
Prosseretal. (1999)
Prosser et al. (1999)
Prosser et al. (1999)
this study
this study
this study
McCracken et al. (1999)
Gamer et al. (2002)
17
To conduct preliminary analyses of microsatellite polymorphism, 100 ng of
DNA from four individuals of each species were amplified in 25 j.iL volumes
containing 10 mM Tris-HC1 (pH 9.0), 50 mM KCI, 0.1% Triton X-100, 0.2 mM each
of dNTPs, 1.5 mM MgCl2, 0.48 p.M forward (labeled with fluorescent ABI dye) and
reverse primer, and 0.3 U Taq DNA polymerase. PCR profiles were 94 °C for 5 mm
followed by 36 cycles of 94 °C for 45 sec, the optimized annealing temperature for 45
sec and 72 °C for 1 mm 30 sec, ending with 72 °C for 10 mi
PCR products were
separated using an ABI 3100 capillary electrophoresis genetic analyzer and data were
visualized using Genotyper 3.7 (ABI Prism). Table 2.3 shows the primer sequences,
repeat sequences and optimal annealing temperatures of the three microsatellite
markers cloned for this study.
DNA extraction, PCR, genotyping
Whole genomic DNA was extracted using sodium dodecyl sulphate-proteinase K
digestion followed by a standard phenol-chloroform extraction, NaC1 purification and
isopropanol precipitation. For all species, 5-100 ng DNA was PCR amplified in a
12.5 p.L reaction with the above reagent concentrations. PCR profiles consisted of 94
°C for 2 mm followed by 36 cycles of 94 °C for 30 sec, appropriate annealing
temperature for 30 sec and 72 °C for 30 sec, ending with 72 °C for 2 mm. PCR
products were genotyped and analyzed as above.
Data analysis
Table 2.3. Genetic characteristics of three microsatellite primer sets that amplify in both T elegans and 7'. sirtalis.
F and R indicate forward and reverse primers, respectively. Tm is the optimum annealing temperature.
Locus
Repeat motif
TEO51B (TTCC)3(TTCA)2(TTCC)3
Primer sequences (5'-3')
F GATTCAAGGCAGTGAACATACC
R ACCACTGTCCCAAACCTACCTC
Species of origin
7'. elegans
Tm (°C)
63
ISO 10
(ATGG)3,(ATGA)6
F TGACTCAGATGCCCTCAGTCTA
R CGGACCAACCAGGAACAGAAAT
T. sirtalis
60
TS042
(GA(CA)4)4
F TCAGGATACGGCAACCAGGCTT
7'. sirtalis
68
R GCTCCCCCCATCACTCAG
19
Genetic diversity. Exact tests for departure from Hardy-Weinberg equilibrium were
performed for each locus separately, and significance was evaluated using the Markov
chain method (Guo and Thompson 1992; Markov chain parameters: 5000
dememorizations; 500 000 steps per chain). Tests for linkage disequilibrium were
performed for each population and globally for each species using a likelihood-ratio
test with level of significance determined by permutation (Slatkin and Excoffier 1996;
Markov chain parameters: 5000 dememorizations, 1000 batches, 5000 iterations per
batch). Levels of statistical significance were adjusted according to a sequential
Bonferroni correction for multiple comparisons (Rice 1989). Genetic variability
within each population was quantified by counting the number of alleles and
determining observed and expected heterozygosities. Number of alleles per locus in
each population and over all populations as well as measures of linkage disequilibrium
were calculated in GENEPOP (Raymond and Rousset 1995). All other analyses were
perfomed in ARLEQUIN v. 2.000 (Schneider et al. 2000).
Genetic dfjerentiation, effective population size and gene flow. Overall and
population pairwise estimates of FST were obtained using a hierarchical analysis of
molecular variance, AMOVA (Excoffier et al. 1992) in ARLEQUIN v. 2.000. This
analysis makes the same assumptions as other methods for estimating FST under
Wright's island model, namely an infninite number of populations with equal sizes
that are constant over time, equal migration rates that are very low, no mutation and no
selection (Wright 1931). Significance was assessed after 16 000 permutations for
20
global estimates and 3000 permutations for pairwise estimates. P-values were
adjusted with the sequential Bonferroni correction.
In order to visualize the pattern of isolation by distance, we regressed genetic
distance, defined as FST/( 1 -FST), on the logarithm of geographic distance, as suggested
by Rousset (1997). We then evaluated the relative roles of gene flow and random
genetic drift using the pattern revealed by the scatter plot (Hutchison and Templeton
1999). The Pearson product moment correlation coefficients between the genetic and
geographic distance matrices were assessed using Mantel tests (Mantel 1967, Mantel
and Valand 1970, Manly 1997), implemented in ARLEQUIN v. 2.000. P-values were
obtained through 10 000 permutations.
MIGRATE v. 1.7.6.1 (Beerli and Felsenstein 2001) was used to calculate
effective population size as a function of mutation rate (0 = 4Np) as well as effective
numbers of migrants (4Nem), where Ne is effective population size, 1u is mutation rate
and m is the rate of migration into the population. This analysis used the stepwise
mutation model (Ohta and Kimura 1973), which assumes that mutations occur in a
stepwise fashion, with the addition or deletion of one repeat unit at a time, and that
loci are neutral and unlinked. Assumptions of the maximum likelihood approximation
using the coalescent approach include diploid individuals reproducing according to a
diffusion equation approximation of a Wright-Fisher model with constant population
sizes, and constant migration and mutation rates (mutation-migration equilibrium;
Beerli 1998, Beerli and Felsenstein 1999).
FST
estimates were used as starting values
for the initial analysis. For all other analyses, ending parameters of the previous run
21
were used as starting values for the next run until results equilibrated at approximately
the same values. Ten short chains with 10 000 sampled genealogies each and two
long chains with 100 000 sampled genealogies each were run for each analysis. One
of every 20 constructed genealogies was sampled, and multiple long chains were
combined for estimates. We used adaptive heating with temperature specifications of
1.0, 1.2, 1.5 and 3.0. Heating allows chains to be run at different temperatures, the
highest of which explores the most genealogy space. Chains can swap based on an
acceptance-rejection step so that colder chains explore peaks while hotter chains
sample more widely. The temperature difference between chains can be adjusted
based on rate of swapping. This method is based on the analysis of Geyer and
Thompson (1991) and is called MC3 or MCMCMC (Markov coupled Markov chain
Monte Carlo).
The correlation between the genetic structures of T.
elegans
and T sirtalis was
assessed for sites in common using Mantel tests. Pairwise genetic distance matrices,
using FSTI(1-FST), were compared in Arlequin 2.000, and the asymmetric matrices of
bi-directional migration rate were compared in CADM (Legendre 2001). Significance
of all Mantel test were assessed over 10 000 permutations. A correlation coefficient
for effective population sizes between species was obtained in SAS (v. 9.2; SAS
Institute 2002).
RESULTS
22
Tests of disequilibrium
Most microsatellite loci were in Hardy-Weinberg and linkage equilibrium in all
populations. For 1'. elegans, the TS042 locus had a significant heterozygote deficit in
17 out of 19 genotyped populations after sequential Bonferroni correction and was
excluded from further analysis. An additional locus was also out of HWE at one
population. For T. sirtalis, the TEO5 lB locus had a significant heterozygote deficit in
9 out of 12 populations genotyped and was excluded from further analysis. An
additional locus was also out of HWE at one population. Linkage disequilibrium was
found in the Feather Lake T sirtalis population between Nsj.t3 and 3Ts.
Allelic variation
Average observed and expected heterozygosities for all populations are shown in
Table 2.1 and for all loci in Table 2.2. Overall observed and expected heterozygosities
were calculated excluding both TS042 for 7'. elegans and TEO5 lB for T sirtalis. The
total number of alleles at a locus varied from two to 19 for T elegans and one to 33
for 7'. sirtalis (Table 2.2). The average number of alleles per locus within a population
ranged from four to six for 7'. elegans and five to nine for 7'. sirtalis (Table 2.1), while
the average number of alleles per population for a locus ranged from two to 11 for T
elegans and two to 16 for T. sirtalis (Table 2.2). Both T elegans and T sirtalis
populations (12 7'. elegans and six 7'. sirtalis) had low incidences of private alleles,
with no more than three in a population (7'. sirtalis at Pine Valley Meadow)
23
Population structure
Global estimates of FST were relatively low but highly significant for both species.
Thamnophis sirtalis had an FST of 0.035 (P < 0.0000 1), and that ofT. elegans was
slightly lower at 0.025 (P < 0.00001). Both garter snakes had approximately the same
total variance in allele size (T. elegans: 24.49, T sirtalis: 21.12). Pairwise estimates
of FST are shown in Table 2.4 for T. elegans and Table 2.5 for T. sirtalis. Thirty-two
out of 190 T. elegans population pairs were found to be significantly differentiated
after sequential Bonferroni correction. Pikes Point had the highest number of
significant comparisons (nine), while populations at Ashurst Lake, Nameless Meadow,
and Rocky Point showed no significant differentiation from other populations. Rocky
Point was remarkably undifferentiated; its most significant comparison was with Pikes
Point with a P-value of 0.20. Among a total of 78 T sirtalis population comparisons,
25 were statistically significant. Colman Lake had the most significant comparisons
(seven), while Roney Corral had none.
Isolation by distance
A significant positive relationship was found between genetic and geographic distance
for both T. elegans (slope
0.015) and T. sirtalis (slope = 0.033; Fig. 2.2). Mantel
tests for the geographic and genetic distance matrices showed that T. elegans and T.
sirtalis had significant Pearson correlation coefficients (r = 0.200, P = 0.0055 for T
elegans; r = 0.4 16, P < 0.0001 for T. sirtalis). The slopes of the regressions for both
species were significantly different (P = 0.04 1).
24
Figure 2.2. Isolation-by-distance plots for A. T elegans and B. T. sirtalis. Circled
points refer to data causing deviations from a pattern of migration-drift equilibrium:
solid circles - restricted gene flow relative to geographic distance, dotted circle high
gene flow relative to geographic distance.
25
0.1
A.
.
0.06
.
0.O5j
0.04
%
0.
0
.
S
55
0.5
S
S
...
S
2
1.5
1
S
In(geographic distance)
$. 3..
S
S Sq
:
4
0.12
1
B.
S
0.1
5
S
S
S
S
0.08
S
5
S
5
S
S
S
s5
0.06
S
S
S
0.04
S
S
S
S
S
S
0.02
S
S
S
S
00
S
S
S
.
0.5
1
1.5
2
In(geographic distance)
2.5
S
S.
3
3.5
41
Table 2.4. Pairwise FST values for T. elegans populations below the diagonal. Associated P-values above the
diagonal, obtained after 3000 permutations. Bold values are significant at the 0.00026 after sequential Bonferroni
correction.
PVM
PVM
AMP
ASH
BLW
BUL
CLG
COL
DNS
GAL
AMP
ASH
BLW
BUL
CLG
COL
DNS
GAL
JKS
LTC
0.0198
0.0552
0.0010
0.0030
0.0327
0.0000
0.0000
0.0000
0.0000
0.0691
0.5349
0.6717
0.9114
0.9293
0.0132
0.0007
0.0073
0.2413
0.0979
0.1785
0.1392
0.9779
0.0374
0.0043
0.0020
0.0152
0.0321
0.1732
0.3874
0.1038
0.0466
0.0023
0.2565
0.0665
0.8549
0.0046
0.0003
0.0007
0.0793
0.0089
0.0169
0.0013
0.0003
0.0314
0.0741
0.4020
0.0033
0.0066
0.0046
0.0003
0.0003
0.0040
0.0017
0.0000
0.013
0.010
-0.002
0.029
-0.006
0.007
0.014
-0.008
0.007
0.005
0.009
-0.008
-0.009
0.000
-0.005
0.040
0.057
0.030
0.053
0.034
0.021
0.014
0.027
0.022
0.046
0.033
0.020
0.034
0.034
0.022
0.025
0.029
0.017
0.029
0.031
0.037
0.005
0.024
0.004
0.009
0.016
0.033
0.048
0.029
LTC
MAH
MCY
0.014
0.019
0.028
0.025
0.032
0.018
0.063
0.061
0.057
0.063
0.028
0.036
0.030
0.030
0.030
0.031
0.017
0.015
0.015
0.043
0.041
0.024
0.001
-0.012
0.008
0.012
-0.006
0.041
0.040
0.059
0.030
0.017
NIML
0.023
0.019
0.010
0.014
0.018
0.017
0.010
0.006
0.009
0.031
0.029
PAP
0.044
0.037
0.037
0.036
0.025
0.074
0.074
0.018
0.048
0.060
0.049
0.035
0.038
0.031
0.025
0.037
0.043
0.058
0.033
P1K
0.035
0.029
RKY
-0.031
-0.045
-0.033
-0.025
-0.034
-0.048
-0.016
-0.033
-0.010
-0.003
-0.043
RON
0.019
0.003
-0.004
0.014
0.023
0.008
0.032
0.047
0.031
0.029
0.026
STF
0.015
-0.009
0.004
-0.004
-0.004
-0.007
0.036
0.041
0.026
0.017
0.020
SUM
0.015
0.006
0.003
0.024
0.014
0.007
0.064
0.071
0.039
0.040
0.020
JKS
0.002
0.0003
0.057
t'J
Table 2.4. (continued)
PVM
AMP
ASH
BLW
BUlL
CLG
COL
DNS
GAL
JKS
LTC
MAH
MCY
NML
PAP
P1K
RKY
RON
STF
SUM
MAH
0.0000
0.0003
0.0017
0.0036
0.0000
0.0000
0.0370
0.0377
0.0040
0.0003
0.0030
0.054
-0.002
0.002
0.031
-0.010
0.038
0.038
0.057
MCY
0.0126
0.3584
0.8737
0.1984
0.0922
0.6714
0.0116
0.0119
0.0003
0.0172
0.1825
0.0003
0.029
0.059
0.085
0.007
-0.006
-0.003
0.005
NML
0.0007
0.0261
0.1045
0.0678
0.0089
0.0165
0.1431
0.2007
0.0754
0.0030
0.0324
0.5425
0.0122
0.001
0.037
-0.021
0.010
0.022
0.034
PAP
0.0000
0.0000
0.0003
0.0000
0.0000
0.0000
0.0010
0.0033
0.0000
0.0000
0.0003
0.2003
0.0003
0.3405
0.029
0.002
0.047
0.047
0.065
P1K
0.0000
0.0013
0.0007
0.0000
0.0000
0.0000
0.0000
0.0000
0.0056
0.0000
0.0003
0.0003
0.0000
0.0003
0.0000
0.007
0.057
0.032
0.060
RKY
0.9990
0.9984
0.9993
0.9574
0.9984
0.9997
0.8519
0.9914
0.8245
0.4813
0.9964
0.9078
0.2341
0.9501
0.3018
0.2026
-0.003
-0.059
-0.013
RON
0.0149
0.3574
0.6503
0.1273
0.0112
0.1415
0.0202
0.0013
0.0017
0.0149
0.0840
0.0007
0.6063
0.1365
0.0000
0.0000
0.5226
0.017
0.006
STF
0.0036
0.9613
0.2103
0.5974
0.7395
0.9590
0.0013
0.0003
0.0003
0.0222
0.0559
0.0000
0.4979
0.0056
0.0000
0.0003
0.9997
0.0327
0.009
SUM
0.0139
0.1679
0.2668
0.0205
0.0222
0.1154
0.0000
0.0000
0.0007
0.0003
0.0668
0.0003
0.1967
0.0010
0.0000
0.0000
0.7623
0.1970
0.0754
Table 2.5. Pairwise FST values for T sirtalis populations below the diagonal. Associated P-values above the diagonal,
obtained after 3000 permutations. Bold values are significant at the 0.0006 level after sequential Bonferroni correction.
PVS
AMP
ASH
BUL
DNS
COL
FEA
GOR MAR NML RON STF SUM
PVS
0.9762
AMP
-0.014
ASH
BUL
COL
0.046
0.028
0.018
0.027
0.053
DNS
0.023
0.072
0.090
FEA
0.004
GOR
0.024
MAH
NML
0.046
0.044
RON
STF
SUM
0.0000
0.0060
0.0000
0.0043
0.2307
0.0010
0.0003
0.0003
0.9448
0.0119
0.1422
0.0096
0.0222
0.0007
0.0003
0.3845
0.0671
0.0073
0.0013
0.2083
0.3035
0.3008
0.0000
0.0000
0.0000
0.0000
0.0013
0.0013
0.0000
0.0043
0.0040
0.0265
0.0000
0.0003
0.0169
0.8519
0.0000
0.0000
0.0268
0.1111
0.2112
0.0109
0.0013
0.0000
0.7937
0.0040
0.0013
0.0000
0.0000
0.0344
0.0010
0.0060
0.0000
0.0565
0.0003
0.0013
0.0506
0.0020
0.0030
0.4777
0.0403
0.1547
0.0007
0.0003
0.0093
0.0569
0.3266
0.0175
0.0026
0.0000
0.0023
0.0013
0.0000
0.0000
0.1597
0.1888
0.079
0.060
0.036
0.003
0.045
0.049
0.085
0.045
0.024
0.050
0.028
0.021
0.031
-0.007
0.062
0.056
0.0188
0.046
0.034
-0.008
0.046
0.0470
0.052
0.098
0.068
0.082
0.043
0.083
-0.007
0.011
0.033
0.020
0.057
0.023
0.019
0.006
0.023
0.010
0.068
0.008
0.004
0.020
0.005
0.072
0.065
0.055
0.0453
0.0509
0.0568
0.0293
0.0007
0.0342
0.0569
0.0230
0.0155
0.0130
0.0035
0.0576 0.0840
0.0524 0.0869
0.0659
0.0115
0.0103
0.4850
0.0003
29
Table 2.6. Estimates of migration rate as measured by 4Nem for T elegans. The
direction of gene flow is from populations given as column labels to populations given
as row labels. Effective population size as measured by 4Nu is shown on the diagonal
(in bold). U95C1 shows upper 95% confidence limit, and L95C1 shows lower 95%
confidence limit.
30
PVM
AMP
ASH
BLW
BUL
CLG
COL
DNS
PVM
0.230
U95C1 0.218
L95C1 0.244
1.602
1.293
1.956
1.798
1.470
5.394
4.810
6.025
3.026
2.593
3.504
1.068
2.171
1.709
1.389
2.074
1.361
1.549
1.246
1.898
AMP
2.007
U95C1 1.581
L95C1 2.504
0.107
0.096
0.118
1.402
1.052
1.823
0.880
0.609
0.880
0.609
1.221
2.282
1.826
2.809
1.221
0.852
0.586
1.189
0.385
0.217
0.623
ASH
3.801
U95C1 3.233
L95C1 4.431
0.441
0.268
0.678
0.104
0.094
0.114
0.932
0.666
1.260
2.599
2.135
3.126
1.814
1.432
2.260
0.640
0.424
0.919
0.441
0.268
0.678
BLW
2.075
U95C1 1.604
L95C1 2.631
0.856
0.568
1.228
0.594
0.360
0.914
0.108
0.097
0.121
2.732
2.186
3.363
1.448
1.062
1.920
0.724
0.462
1.070
0.494
0.284
0.787
BUL
5.225
U95C1 4.650
L95C1 5.845
0.889
0.664
1.158
1.759
1.435
2.129
0.764
0.558
1.016
0.226
0.214
0.240
2.146
1.786
2.553
1.244
0.975
1.559
0.764
0.558
1.016
CLG
3.217
U9SCI 2.706
L95C1 3.788
0.993
0.722
1.325
0.591
0.389
0.854
1.514
1.172
1.915
3.619
3.075
4.223
0.153
0.142
0.165
1.182
0.884
0.804
0.563
1.541
1.105
COL
2.325
U95C1 1.832
L95C1 2.900
0.223
0.096
0.431
0.444
0.249
0.720
0.382
0.205
0.641
2.930
2.372
3.571
1.274
0.919
1.711
0.106
0.095
0.119
0.255
0.116
0.474
DNS
2.070
U95C1 1.625
L95C1 2.589
0.612
0.386
0.913
0.962
0.670
1.329
0.759
0.504
1.090
1.720
1.318
2.197
0.671
0.432
0.983
0.408
0.230
0.661
0.113
0.101
0.126
GAL
3.426
U95C1 2.963
L95C1 3.935
1.308
1.030
1.631
1.273
0.663
0.472
0.901
3.461
2.995
3.972
2.277
1.904
2.696
0.538
0.368
0.754
1.309
1.031
1.633
JKS
2.580
U95C1 2.111
L95C1 3.114
0.531
1.112
1.795
0.335
0.791
0.815
1.473
0.936
0.666
1.271
1.409
2.245
1.263
0.945
1.646
0.202
0.092
0.376
0.783
0.539
1.092
LTC
2.087
U95C1 1.654
L95C1 2.588
0.325
0.174
0.545
0.054
0.009
0.167
0.217
0.099
0.403
2.437
1.967
2.975
0.704
0.467
1.010
0.217
0.099
0.403
0.190
0.081
0.367
0.999
1.592
0.820
31
Table 2.6. (continue
GAL
3.026
JKS
LTC
MAH
1.175
1.601
6.501
U95C1 2.594
L95C1 3.504
0.914
1.482
1.293
AMP
1.429
U95C1 1.075
L95C1 1.854
ASH
1.447
U95C1 1.108
PVS
L95C1 1.848
BLW
2.467
U95C1 1.949
L95C1 3.068
NML
1.955
5.856
7.191
MCY
0.445
0.293
0.643
1.481
PAP
11.465
10.602
12.374
0.385
0.217
0.623
0.275
0.138
0.482
3.107
2.569
3.716
1.017
0.724
1.381
1.402
1.052
1.823
5.472
4.746
6.267
0.859
0.605
1.176
0.790
0.515
1.150
0.417
0.249
0.647
0.329
0.165
0.577
2.893
2.402
3.447
3.061
2.481
3.726
0.123
0.044
0.263
0.856
0.568
1.228
0.662
0.443
0.944
0.560
0.334
0.869
6.081
5.355
6.869
5.432
4.644
6.303
2.332
1.511
5.562
4.969
6.202
4.754
4.127
5.442
0.498
0.335
0.705
0.473
0.295
0.712
2.168
1.806
2.576
0.662
0.446
0.939
12.623
11.717
13.575
7.403
6.613
8.254
2.882
2.460
3.349
2.461
2.018
2.964
1.175
0.914
P1K
3.275
2.824
3.771
1.757
1.360
2.224
1.498
1.152
1.905
1.811
1.373
BUL
1.883
U95C1 1.547
L95C1 2.265
CLG
2.388
U95C1 1.952
L95C1 2.885
1.212
1.855
1.301
0.987
1.675
1.599
1.291
1.953
0.378
0.222
0.595
COL
2.230
U95C1 1.747
L95C1 2.794
0.892
0.601
1.264
0.289
0.139
0.521
3.986
3.327
4.727
0.159
0.057
0.342
0.542
0.323
0.841
5.987
5.171
6.884
1.338
0.973
1.784
DNS
2.680
U95C1 2.169
L95C1 3.266
0.117
0.036
0.271
0.029
0.002
0.128
3.761
3.149
4.447
0.379
0.208
0.624
0.787
0.526
1.122
6.239
5.440
7.112
0.990
1.767
GAL
0.176
U95C1 0.165
L95C1 0.187
1.543
1.239
1.892
1.004
0.764
1.290
3.675
3.195
4.202
0.556
0.382
0.775
0.861
0.640
1.128
6.509
5.862
7.202
2.242
JKS
U95C1
L95C1
LTC
U95C1
L95C1
0.107
0.097
0.118
0.514
0.316
0.782
0.505
0.315
0.760
0.073
0.065
0.083
2.956
2.452
3.524
0.455
0.276
0.698
0.135
0.049
0.291
0.429
0.256
0.667
0.352
0.194
0.579
6.597
5.829
7.430
3.630
3.049
4.280
0.884
0.623
1.210
1.768
1.386
2.216
0.406
0.234
0.648
1.543
1.176
1.979
1.341
1.871
2.658
0.921
0.645
1.266
32
Table 2.6. (continued)
RKY
RON
STF
SUM
PVS
U95C1
L95C1
0.445
0.293
0.643
1.656
1.342
2.016
2.030
1.680
2.426
1.460
1.166
1.799
AMP
U95C1
0.385
0.217
0.623
0.550
0.343
0.827
1.292
0.957
1.698
1.017
0.724
1.381
0.368
0.212
0.586
0.809
0.563
1.117
0.785
0.543
1.089
0.638
0.423
0.915
0.691
0.436
1.031
0.362
0.188
0.620
1.416
1.033
1.882
0.296
0.142
0.534
0.338
0.208
0.514
0.817
0.603
1.077
2.116
1.758
2.519
1.991
1.645
2.383
0.828
0.583
1.133
1.230
0.925
1.595
2.128
1.718
2.599
0.782
0.545
1.080
0.382
0.205
0.641
0.510
0.299
0.802
2.040
1.580
2.581
0.637
0.397
0.958
0.175
0.070
0.354
1.079
0.767
1.465
2.391
1.910
2.946
1.136
0.816
1.531
L95C1
0.556
0.382
0.775
0.555
0.382
0.775
2.403
2.019
2.833
0.681
0.487
0.921
JKS
U95C1
L95C1
0.429
0.256
0.667
0.253
0.127
0.443
1.213
0.901
1.589
0.455
0.276
0.698
LTC
U95C1
L95C1
0.217
0.099
0.403
1.327
1.570
1.200
0.298
0.155
0.510
L95C1
ASH
U95C1
L95C1
BLW
U95C1
L95C1
BUL
U95C1
L95C1
CLG
U95C1
L95C1
COL
U95C1
L95C1
DNS
U95C1
L95C1
GAL
U95C1
0.989
1.734
2.010
33
Table 2.6.
MAH
U95C1
L95C1
MCY
(continued)
PVM
AMP
ASH
BLW
BUL
CLG
COL
DNS
5.344
4.797
5.932
1.975
1.650
1.897
1.579
2.255
1.003
0.777
1.270
4.155
3.674
4.675
3.449
3.013
3.925
1.803
1.493
2.153
1.709
1.408
2.050
1.842
1.397
2.372
0.569
0.340
0.884
0.100
0.025
0.260
0.600
0.363
0.923
2.340
0.687
1.408
0.502
0.289
0.801
0.368
0.191
0.63 1
0.301
0.145
0.543
2.828
2.366
3.347
0.553
0.363
0.798
0.553
0.363
0.798
0.442
0.276
0.665
2.033
1.646
2.478
0.774
0.545
1.059
0.685
0.471
0.956
0.464
0.293
0.692
2.041
1.746
2.367
1.869
1.587
2.182
1.181
U95C1
L95C1
7.831
7.239
8.455
0.960
1.433
5.681
5.178
6.215
3.198
2.825
3.603
1.414
1.171
1.688
2.544
2.213
2.907
P1K
U95C1
L95C1
2.427
2.040
2.860
0.880
0.656
0.557
0.383
0.777
2.858
2.436
3.326
1.151
0.892
1.457
0.989
0.750
1.151
2.014
1.664
2.411
1.273
0.342
0.210
0.519
RKY
1.718
1.288
0.168
0.060
0.361
0.740
0.472
2.234
0.134
0.042
0.312
1.093
0.874
0.580
1.254
0.842
0.553
1.216
0.403
0.216
0.677
0.403
0.216
0.677
1.602
1.256
2.007
0.229
0.115
0.401
0.732
0.507
1.016
0.458
0.285
0.689
1.946
1.561
2.389
0.618
0.413
0.881
0.275
0.147
0.461
0.526
0.340
0.772
2.534
0.830
1.419
0.524
3.734
0.961
0.983
0.721
U95C1
L95C1
2.100
0.593
1.102
0.341
3.202
0.704
0.723
0.502
3.023
1.122
1.793
0.763
4.322
1.273
1.298
0.995
SUM
U95C1
L95C1
2.758
2.280
3.298
0.781
0.541
1.084
0.781
0.541
1.084
0.268
0.139
0.460
2.538
2.081
3.057
0.879
0.622
1.198
0.488
0.304
0.734
0.488
0.304
0.734
U95C1
L95C1
NML
U95C1
L95C1
PAP
U95C1
L95C1
RON
U95C1
L95C1
STF
1.005
34
Table 2.6.
(continued)
GAL
JKS
LTC
MAH
MCY
NML
PAP
P1K
3.652
3.203
4.142
1.412
1.140
1.724
0.721
0.532
0.950
0.290
0.277
0.304
0.862
0.654
1.723
1.421
1.111
2.066
15.053
14.120
16.025
2.525
2.155
2.935
0.402
0.215
0.674
0.569
0.340
0.884
0.469
0.264
0.759
2.110
0.074
0.064
0.085
0.536
0.314
0.843
3.584
2.948
4.307
1.105
0.770
1.526
1.348
1.038
1.716
0.464
0.293
0.692
0.265
0.142
0.445
3.935
3.385
4.542
0.111
0.040
0.237
0.110
0.100
0.121
5.858
5.181
6.592
0.663
0.453
0.929
L95C1
5.299
4.815
5.815
2.090
1.792
2.421
1.094
0.882
1.337
9.997
9.325
10.700
0.861
0.674
1.078
3.160
2.789
3.562
0.463
0.448
0.479
5.042
4.570
5.546
P1K
U95C1
L95C1
2.194
1.827
2.606
1.276
1.002
1.597
0.377
0.238
0.563
4.963
4.401
5.572
0.198
0.103
0.339
1.276
1.002
1.597
6.616
5.963
7.315
0.133
0.125
0.143
RKY
U95C1
1.311
0.271
0.942
1.767
0.124
0.503
0.202
0.080
0.409
1.816
1.373
2.344
0.168
0.060
0.361
0.538
0.316
0.846
1.917
1.462
0.874
0.580
2.459
1.255
0.916
0.660
1.229
0.618
0.413
0.881
2.751
2.286
3.275
0.504
U95C1
L95C1
0.755
0.525
1.042
0.744
0.481
0.303
0.717
5.083
4.443
5.780
0.687
0.469
0.963
STF
U95C1
L95C1
2.489
2.060
2.975
1.179
0.892
1.522
0.524
4.958
4.341
5.631
0.393
0.238
0.604
1.572
1.236
1.964
7.228
6.477
8.034
2.009
SUM
U95C1
L95C1
1.784
1.405
2.225
0.390
0.229
0.614
0.415
0.248
0.644
2.172
0.220
0.106
0.396
1.098
0.808
1.451
5.855
5.145
6.628
0.634
0.420
0.910
MAB
U95C1
L95C1
MCY
U95C1
L95C1
NIML
U95C1
L95C1
PAP
U95C1
L95C1
RON
0.341
0.763
1.631
2.674
1.751
2.655
0.321
1.626
2.448
35
Table 2.6.
(continued)
RKY
RON
STF
SUM
0.737
0.546
0.969
1.33 1
1.068
1.635
2.728
2.342
3.153
1.834
1.522
2.187
0.301
0.145
0.543
0.335
0.168
0.587
0.971
0.659
1.369
0.469
0.264
0.759
0.133
0.053
0.269
0.287
0.158
0.473
2.210
2.672
0.818
0.582
1.110
L95C1
0.418
0.293
0.575
2.324
2.008
2.671
3.405
3.020
3.822
2.238
1.928
2.578
P1K
U95C1
L95C1
0.198
0.103
0.339
0.665
0.473
0.903
0.899
0.672
1.171
0.521
0.354
0.735
RKY
0.070
0.060
0.081
0.403
0.216
0.677
1.176
0.828
1.610
0.538
0.316
0.846
0.298
0.164
0.490
0.086
0.078
0.096
0.870
0.622
1.176
0.572
0.376
0.827
0.393
0.238
0.604
1.202
0.911
1.548
0.152
0.141
0.163
1.397
1.082
1.768
0.317
0.175
0.522
0.415
0.248
0.644
0.781
0.541
1.084
0.109
0.099
0.119
MAR
U95C1
L95C1
MCY
U95C1
L95C1
NML
U95C1
L95C1
PAP
U95C1
U95C1
L95C1
RON
U95C1
L95C1
STF
U95C1
L95C1
SUM
U95C1
L95C1
1.805
36
The pattern produced by the regression of genetic distance on geographic distance was
similar for both species. The scatterplots were wedge-like, with the variance in
genetic distance increasing with geographic distance. For T. elegans, the points most
responsible for this effect had relatively high genetic distances in relation to
geographic distance and were all associated with Pikes Point. The two most deviant
points described distances from Pikes Point to Deans Meadow and Colman Lake (Fig.
2.2A). The most deviant point for T. sirtalis was associated with Deans Meadow and
Nameless Meadow, which were relatively more differentiated given the geographic
distance. A number of points for T. elegans also had relatively low genetic distances
given the geographic distances. These points were largely due to distances from
Rocky Point, which had many FST'S of 0, suggesting recent colonization of that site
(Table 2.4).
Both species showed patterns of isolation-by-distance that indicate lack of
regional migration-drift equilibrium (case IV; Hutchison and Templeton 1999).
Specifically, the relative importance of gene flow and drift varies depending on
geographic distance. Smaller geographic distances were associated with smaller
genetic distances that had less scatter, a pattern consistent with the homogenizing
effect of gene flow over short distances. Larger geographic distances were associated
genetic distances that had more scatter, indicating little correlation between the two
parameters. Such a pattern is consistent with more extreme genetic isolation, where
drift is driving genetic differentiation among populations irrespective of geographic
distance, and gene flow is minimal. For T. elegans, the distance at which the
37
dominant evolutionary force shifts from gene flow to genetic drift is approximately 5.5
km, while for T. sirtalis, this distance is around 4 km.
Effective population size and migration rate.
Estimates of effective population size for T elegans ranged from a (9of 0.070 to 0.463
with an average of 0.149 (Table 2.6). Assuming a typical vertebrate microsatellite
mutation rate of 1 0 per locus per generation (Dallas 1992, Edwards et al. 1992,
Weber and Wong 1993, Banchs et al. 1994, Ellegren 1995), this value for (9
translates to an average Ne of approximately 375. Papoose Meadows had by far the
largest Ne (1160) with Mahogany Lake, Pine Valley Meadow and Bullard Lake as
runners up with effective sizes of 725, 575 and 565, respectively (Fig. 2.3). All of
these populations were also the largest sources of migrants, even after taking their
large effective sizes into account. Papoose Meadows was the most important source
population for the entire study area with a combined 4Nem of 128.6, which translates
to a migration rate of 0.097 (Figure 2.3). In other words, 9.7% of individuals at all
other sites originated from the Papoose Meadows population. The smallest
populations were Little Cleghom Reservoir (N = 185), McCoy Flat Reservoir
185) and Rocky Point
(Ne
(Ne =
= 175). Estimates of 4Nem for paired T. elegans populations
were low, averaging 1.40, which translates to an Nem of 0.35 and a migration rate of
0.00 12 migrants per generation. Papoose Meadows and Mahogany Lake had
relatively low rates of immigration, while both Pine Valley Meadow and Bullard Lake
had high rates of immigration, although they were somewhat lower than emigration
Figure 2.3. Map of the study area showing relative effective population sizes and
directions and rates of migration for T. elegans. Circle size and arrow width are
proportional to effective population size and migration rate, respectively.
'1
N
BLW
0
JCK
STF.
LTC
..
'BUL
ASH
CLG
... ..
SUM
a,
RKY:
0
A
PVM
o
0
MAH
PAPNC
5km
40
rates. Populations with the highest immigration rates from all other populations were
Ashurst Lake (m = 0.026), Colman Lake (m = 0.025), Antelope Mountain Pond (m =
0.025) and Blue Water (m = 0.025). Rocky Point had a high input and low output of
migrants combined with the smallest Ne (175). Pikes Point, another lakeshore
population, had approximately equal rates of immigration and emigration.
Effective population sizes for T sirtalis were comparable to but slightly
smaller than those for 7'.
with 6) ranging from 0.073 to 0.324 and averaging
elegans,
0.130. Again, assuming a mutation rate of 10, the average Ne was approximately 325
(Table 2.9). Pine Valley Meadow had by far the largest Ne (810), followed by Gordon
Lake (405; Fig. 2.4). The smallest population was Antelope Mountain Pond with 180
effective individuals. Average
slightly higher than for 7'.
4Nem between population pairs was also low but
elegans
at 2.14, which corresponds to an Nem of 0.54 and a
migration rate of 0.0017. Pine Valley Meadow was the only major source of migrants
in the study area, with a total 4Nem out of the population of 84.3 (m = 0.076). Bullard
Lake, Feather Lake and Summit Lake were all primary recipients of migrants, most of
which came from Pine Valley Meadow (Fig. 2.4). However, Bullard Lake also
received a substantial proportion of migrants from Colman Lake (m = 0.006) and
Gordon Lake (m = 0.004), both of whose rates of emigration were highest for migrants
to Bullard Lake. Colman Lake had approximately equal rates of immigration and
emigration.
Overall, the population genetic structures of the two species at 11 shared sites
were significantly correlated. Mantel tests comparing symmetric matrices of pairwise
41
Table 2.7. Estimates of migration rate (4Nem) and effective
population size (4Nu) for T sirtalis. Conventions are as in Table
2.6.
PVS
AMP ASH
BUL
COL
DNS
FEA
PVS
U95C1
L95C1
0.324
0.309
0.340
1.940
1.644
2.270
5.298
4.797
5.832
3.814
3.392
4.270
3.009
2.636
3.415
2.756
2.400
3.146
4.236
AMP
U95C1
4.981
4.300
5.731
0.073
0.063
0.084
1.518
1.157
1.948
0.453
0.270
0.703
1.362
1.022
1.771
0.533
0.332
0.801
0.560
0.353
0.836
7.382
6.689
8.122
0.850
0.629
1.116
0.141
0.130
0.153
1.915
1.573
1.842
1.508
1.641
1.326
2.303
2.224
2.003
0.900
0.672
1.174
8.853
7.866
9.920
1.918
1.479
2.436
5.015
4.282
5.828
0.127
0.115
0.141
7.939
7.006
8.951
2.354
1.864
2.924
2.259
5.080
4.521
5.683
0.811
0.600
1.066
1.484
2.122
1.769
2.520
0.104
0.094
0.115
1.395
1.112
1.721
0.795
0.587
1.048
6.344
5.606
7.145
0.532
0.339
0.786
1.451
1.114
1.850
1.163
1.526
1.179
1.934
0.100
0.089
0.112
0.819
0.570
1.129
6.380
5.613
7.215
0.801
0.550
1.118
1.995
1.582
2.475
0.961
0.684
1.305
1.476
1.125
1.894
0.100
0.089
0.113
L95C1
ASH
U95C1
L95C1
BUL
U95C1
L95C1
COL
U95C1
L95C1
DNS
U95C1
L95C1
FEA
U95C1
L95C1
1.191
1.821
0.864
1.523
2.276
1.831
2.787
3.791
4.715
1.759
1.339
42
Table 2.7.
(continued)
GOR MAH NML RON
PVS
U95C1
L95C1
4.542
4.080
5.038
2.256
AMP
U95C1
1.891
1.484
L95C1
2.365
ASH
U95C1
3.610
L95C1
1.935
2.986
2.614
2.610
3.391
0.505
0.310
0.768
0.876
0.610
0.904
0.676
4.134
2.297
1.920
2.720
BUL
4.528
U95C1 3.833
L95C1 5.304
1.206
0.866
1.626
1.701
1.290
2.191
COL
U95C1
1.688
1.375
2.045
0.828
0.616
1.085
2.634
2.169
L95C1 3.160
L95C1
3.131
DNS
U95C1
FEA
U95C1
L95C1
2.599
2.122
3.142
1.211
STF
SUM
2.983
2.612
3.389
3.035
2.660
3.443
2.085
1.777
2.427
0.824
0.567
1.150
1.038
0.745
1.398
1.225
1.552
1.246
1.904
1.788
1.458
0.904
1.614
2.164
1.301
1.023
1.625
1.805
2.853
2.013
1.562
2.542
0.997
0.690
1.384
1.486
1.194
1.822
0.484
0.326
0.686
0.777
0.571
1.026
0.432
0.284
0.624
1.597
1.242
2.014
1.614
1.256
2.034
0.532
0.340
0.787
1.065
0.781
1.411
1.376
1.048
1.766
1.479
1.128
1.897
1.317
0.987
1.714
2.466
2.003
2.997
1.558
1.197
1.986
1.165
1.178
2.290
0.856
1.540
43
Table 2.7.
(continued)
PVS
AMP ASH
BUL
COL
DNS
FEA
7.997
7.227
8.821
1.017
0.759
1.329
4.436
3.870
5.055
3.468
2.970
4.018
1.653
1.317
2.042
2.129
1.744
2.567
1.687
1.346
2.081
6.874
6.102
7.708
0.609
0.401
0.880
2.489
2.037
3.004
1.145
0.848
1.505
0.759
0.522
1.059
1.122
0.828
1.478
0.930
0.665
6.910
6.108
7.781
1.363
1.025
1.768
2.806
2.307
3.372
0.788
0.539
1.104
1.566
1.201
1.998
1.047
0.755
1.406
2.986
2.471
3.568
8.929
8.027
9.896
0.354
0.200
0.574
2.228
1.795
1.975
1.568
0.304
0.163
1.730
1.351
2.727
2.447
0.835
0.582
1.154
0.511
2.174
STF
U95C1
L95C1
7.660
6.783
8.611
1.299
0.959
1.712
2.449
1.969
3.002
3.593
3.004
4.255
1.190
0.866
1.587
1.044
0.743
1.418
1.385
1.032
1.810
SUM
U95C1
L95C1
6.942
6.065
7.900
0.537
0.320
0.834
1.927
1.482
2.452
3.059
2.489
3.709
0.912
0.617
1.287
0.473
0.272
0.755
0.538
0.321
0.799
GOR
U95C1
L95C1
MAll
U95C1
L95C1
NML
U95C1
L95C1
RON
U95C1
L95C1
1.258
44
Table 2.7. (continued)
GOR MAH NML RON
GOR
U95C1
0.162
L95C1
0.177
STF
SUM
1.713
1.370
1.280
0.988
1.626
1.777
1.428
2.179
1.409
1.100
1.771
3.570
3.064
4.127
1.952
1.554
2.412
0.100
0.090
1.875
1.487
0.877
0.620
0.113
2.326
0.731
0.500
1.025
1.195
0.341
0.192
0.552
1.908
1.502
2.381
1.128
0.824
1.500
0.127
1.624
1.253
2.062
0.786
0.537
1.102
0.840
0.582
1.166
2.508
2.045
3.035
1.339
1.009
1.733
1.012
0.730
1.359
0.113
1.191
0.881
1.565
0.658
0.437
0.934
STF
3.845
U95C1 3.234
L95C1 4.528
1.100
0.789
1.483
1.470
1.106
1.907
1.984
1.553
2.487
0.124
1.271
0.112
0.935
1.679
SUM
2.934
U95C1 2.377
L95C1 3.571
1.263
0.911
1.696
1.232
0.884
1.660
1.863
1.428
1.200
0.858
1.623
MAIl
U95C1
L95C1
NIML
U95C1
L95C1
RON
U95C1
L95C1
0.149
0.113
0.143
0.102
0.127
2.380
0.139
2.109
0.091
0.080
0.105
45
Figure 2.4. Map of the study area showing relative effective population sizes and
directions and rates of migration for T. sirtalis. Conventions are as in Fig. 2.3.
46
STF
ASH
PVM
RON
NML
5km
000L
47
FST
estimates revealed a correlation of 0.412 (P = 0.009). The same test comparing
asymmetrical matrices of pairwise bi-directional migration rates (m) revealed a
slightly lower but still significant correlation (r 0.275; P = 0.024). Effective
population sizes
(Ne),
however, were insignificantly correlated for both species (r =
0.397; P = 0.227).
DISCUSSION
Predictions based on the relative abundances of each species and the spatial
structuring of their populations were all supported by the microsatellite data. Effective
population sizes were slightly larger on average for T. elegans (mean Ne = 375), which
is more abundant locally, than for T sirtalis (mean Ne = 325), though this difference is
not statistically significant (results not shown). More population differentiation was
found for T. sirtalis
(FST
= 0.035), which has fewer populations distributed over the
same area, than T. elegans
(FST = 0.025). Thamnophis sirtalis also exhibited a
stronger pattern of isolation by distance (r = 0.4 164; P < 0.0001) than T elegans (r =
0.1995; P = 0.0055), although their average migration rates were comparable (m =
0.0012 for T. elegans; m = 0.0017 for T. sirtalis). Overall patterns of population
genetic structure for T. elegans and T. sirtalis estimated by two methods were found to
be significantly correlated.
We found a moderate degree of population differentiation for the two garter
snakes at a small spatial scale, even though they are locally abundant, have widespread
distributions and have the capacity to travel long distances. Several other studies have
also found significant genetic differentiation in snakes over short geographic
distances. Prior et al. (1997) obtained comparable levels of population differentiation
at distances less than 50 km using RAPD markers in the black rat snake (Elaphe
obsoleta; FST
= 0.006
- 0.056), and Gibbs et al. (1997) found even higher levels of
differentiation at micro satellite loci between populations of eastern massasauga
rattlesnakes (Sistrurus catenatus). The latter populations were between two and 500
km apart, with FST values ranging from 0.085 to 0.261. Finally, Prosser et al. (1999)
studied microsatellite variation among three populations of the northern water snake
(Nerodia s. sipedon), each within two km of the others, and found a small but
significant global FST (0.006, P = 0.009). Two other studies on genetic differentiation
among Lake Erie T. sirtalis populations (Lawson and King 1996; Bittner and King
2003) found comparable levels of global FST for allozymes (0.01 to 0.08) and
microsatellites (0.03 to 0.04) across populations separated by a maximum of 108 km,
although significance levels of FST were not reported. Previous studies in high latitude
populations of T. sirtalis have documented seasonal migrations away from den sites at
up to 18 km (Gregory and Stewart 1975, Gregory 1977), so it is surprising that both T
elegans and T. sirtalis populations are genetically structured on such a small spatial
scale.
49
Estimates of effective population size and bi-directional migration rates
between population pairs identified important source populations for both species. In
these populations, total emigration exceeded immigration, while other populations
received more total migrants than they exported. These two kinds of populations meet
Pullium's (1998) criteria for source and sink populations, respectively. Papoose
Meadows for T.
elegans
and Pine Valley Meadow for 7'. sirtalis had much larger
effective sizes and emigration rates than the other populations. In both cases, they are
approximately three times the average size of the other populations and contribute
around five times the migrants. The observed source-sink patterns, in conjunction
with a regional lack of migration-drift equilibrium for both species, suggest frequent
extinction-recolonization events indicative of metapopulation dynamics (Hanski and
Simberloff 1997, Stacey et al. 1997). This conclusion is also supported by field
observations of massive snake die-offs and population crashes following drought years
in the study system (Arnold, unpubl. data). Further research is needed to determine
the relative frequency of extinction events by investigating stability of effective
population sizes over time (Hoffthan et al. 2004). To our knowledge, this is the first
study to document source-sink metapopulation dynamics in a natural system using
molecular markers. The ability to identify sources using neutral markers has
important implications for conservation biology. Maximum likelihood methods based
on the coalescent allow bi-directional estimation of migration rates and subsequent
identification of likely source populations that may be prioritized for conservation.
50
Sink populations, those with negative population growth, are more difficult to
identify using these methods. The net removal of individuals from a population may
be due to emigration or mortality. Migration data alone are not sufficient to determine
if a population's size is decreasing due to a high mortality rate and would go extinct in
the absence of immigration. Nevertheless, potential candidates for extinction can be
identified as populations with small effective sizes and rates of immigration that
exceed emigration. Such populations can then be targeted for studies on direct
measures of population growth using mark-recapture techniques to verify the
molecular results. In this study, we found one potential sink in the Rocky Point T.
elegans
population. It had the smallest effective size, and its total immigration far
exceeded total emigration. Unusually low pairwise Fi values between Rocky Point
and other populations support the hypothesis that a high proportion of the population
are immigrants or recent descendants of immigrants from throughout the study area.
Because T
elegans
and 1'. sirtalis tend to converge on amphibian prey
wherever they coexist (Kephart 1982, Kephart and Arnold 1982), it is not surprising
that they have different source localities. Although effective population sizes for the
two species were uncorrelated (P = 0.227), Kephart's (1982) observation that
coexistence is more likely at sites with more species of breeding amphibians suggests
that these two species are competitors. Why are their population genetic patterns so
similar? Both species exhibit metapopulation dynamics dominated by a single
massive population with similar rates of emigration. A logical next step is to ask what
ecological or evolutionary factors might be causing the observed similarities (and
51
differences) in these two species. Their similar environment and community
interactions could play a significant role, though evolutionary constraints may
uniquely limit each species' ecological response.
Although we attempted to sample every population of significance in the study
area, some important populations were undoubtedly excluded. In particular,
populations along the perimeter of the study area may receive migrants from
unsampled sites. The effect of missing populations on estimates of population
structure are discussed by Beerli (2004). In a two-population model, estimates of
effective population size as measured by (9 (4Nu), will be biased upward by "ghost"
populations, but estimates of M(m/p) are robust to the presence of unsampled
populations. Values for
4Nem ((9M), therefore, would also be biased upward. Sample
size does not significantly affect these results, and the influence of ghost populations
decreases as more populations are sampled (Bittner and King 2003, Beerli 2004).
Because we sampled 20 populations ofT.
elegans
and 13 of T. sirtalis, the adverse
effects of missing populations should be minimal.
Direct measures of migration and effective population size for T.
elegans
and
T sirtalis at the study area using mark-recapture methods produced estimates (Kephart
1981) comparable to those found in this study. In Kephart's study, variance effective
size was estimated as a function of census size and variance in reproductive success
using the equation,
N
2N-2
1+ (Vk 1k)
52
(Wright 1931, Crow and Kimura 1971) where N represents population census size,
Vk
is the variance in reproductive success, and k is the average number of gametes each
individual contributes to the next generation (assumed to equal 2; Crow and Norton
1955). Reproductive success in garter snakes was assessed by palping pregnant
females to count embryos, thereby obtaining an estimate of litter size (Fitch 1987, Fan
and Gregory 1991). Sufficient data were available to estimate effective population
size for T. elegans populations at Mahogany Lake and Pikes Point and T. sirtalis
populations at Mahogany Lake and Roney Corral (Table 2.10). Microsatellite
estimates of effective population size were obtained from estimates of e, assuming a
Table 2.8. Comparison of mark-recapture and microsatellite
estimates of effective population size.
Population
mark-recapture microsatellite
T. elegans Mahogany Lake
235
724
Pikes Point
204
333
T sirtalis Mahogany Lake
148
251
Roney Corral
511
283
mutation rate of 10. Three of the four comparisons showed underestimation of
effective population size from mark-recapture methods, with only the T sirtalis Roney
Corral population having a larger estimate from mark-recapture, which was almost
twice the value of the microsatellite estimate. The greatest difference was for the T.
elegans Mahogany Lake population, for which the microsatellite estimate of effective
size exceeded the mark-recapture estimate by a factor of three. Overall, however, the
two methods produced roughly comparable estimates of effective population size.
53
Kephart (1981) documented six dispersal events from 823 recapture events at
21 study sites (Table 2.11), 12 of which were included in this study, all between sites
less than 3 km apart. Four of these cases involved movement by T. elegans
individuals, and two involved T. sirtalis. Of the T elegans dispersal events, one was
from a series of five vernal ponds not included in the present study (SVP, see Table
2.1) to Colman Lake (1.7 km), one was from SVP to Mahogany Lake (3.0 km), one
was from Mahogany Lake to Nameless Meadow (1.4 km), and one was from
Nameless Meadow to Mahogany Lake (1.4 km). Of the T. sirtalis dispersal events,
both were between Colman Lake and SVP (1.7 km), one in each direction. From these
data, Kephart (1981) concluded that
Table 2.9. Comparison of mark-recapture and microsatellite estimates of migration
rate. Values in italics are the average migration rates, m (microsatellite), into the
receiving population estimated from microsatellite data (this study). Census sizes
for the recipient population and m (mark-recapture) are from Kephart (1981).
m
Census m (markFrom
To
size
recapture) (microsat)
0.001
27
0.037
1'. elegans SVP
Colman Lake
0.001
371
0.003
SVP
Mahogany Lake
0.004
Mahogany Lake Nameless Mdw
111
0.009
0.001
371
0.003
Nameless Mdw Mahogany Lake
T. sirtalis
74
0.014
0.001
SVP
Colman Lake
0.111
Colman Lake
SVP
9
interdemic migration was very low. We calculated those migration rates as the
proportion of migrants in a population, using the number of individuals captured at
each site as an estimate of population size. Indirect estimates of migration rate were
calculated from estimates of e. Because SVP were not included as a population in the
current study, the indirect estimate of migration from this site was replaced by the
54
average migration rate into the receiving population. Migration rates obtained by
direct observation of dispersal events always overestimated those obtained using
maximum likelihood of molecular marker data. Direct estimates of migration were
from a factor of three to an order of magnitude higher than indirect estimates. This
discrepancy is probably because observed dispersals only represent movement per se
and not gene flow. Of the individuals that disperse from their natal populations, only a
fraction of those will succeed in contributing gametes to subsequent generations of the
new population, thus allowing detection of those successful dispersals using molecular
markers. Nevertheless, Kephart' s (1981) conclusion that migration events are rare is
supported by the molecular data.
For study sites supporting populations of both T elegans and T sirtalis, there
was a weak but significant correlation between genetic structure of the two species.
Such a relationship may be a function of a common environment, ecological
similarity, evolutionary history, or some combination of these factors. The
microgeographic scale of the study area points to a greater role for landscape and
ecological factors rather than evolutionary processes in shaping the spatial partitioning
of genetic variation in this system. Interestingly, garter snake populations were
significantly structured around hibemacula associated with water bodies, even though
water per se is not required for thermoregulation, courtship, parturition or hibernation.
The observed pattern appears to be a direct effect of the patchy distribution of their
main prey items (amphibians, fish and leeches) for which water is an obvious
necessity. The shared diet of both species and the patchy distribution of their prey
55
may therefore be central to the population genetic dynamics of the garter snake
community.
In contrast to the Lassen Co. study system, sympatric populations of T. elegans
and T. sirtalis in coastal California and Washington have been shown to exploit
terrestrial prey species (Arnold 1992). While Lassen Co. garter snakes converge on
prey types where they coexist, T elegans in both coastal systems specialize on slugs,
whereas T. sirtalis consumes amphibians. Since slugs are distributed homogeneously
across the landscape, as opposed to the discrete ponds inhabited by amphibians, one
can hypothesize that T. sirtalis populations in the coastal systems will be more
spatially structured than the sympatric T. elegans populations. Such a comparative
approach to studies of systems with divergent species dynamics promises to provide
further insight into how ecological processes shape community genetics.
REFERENCES
Aleksiuk, M. and P.T. Gregory. 1974. Regulation of seasonal mating behavior in
Thamnophis sirtalis parietalis. Copeia 1974:681-689.
Anderson, B., I. Olivieri, M. Lourmas and B.A. Stewart. 2004. Comparative
population genetic structures and local adaptation of two mutualists. Evolution
58:1730-1747.
Arnold, S.J. 1992. Behavioural variation in natural populations. VI. Prey responses
by two species of garter snakes in three regions of sympatry. Animal
Behaviour 44:705-7 19.
Banchs, I., A. Bosh, J. Guimera, C. Lazaro, A. Puig and X. Estivill. 1994. New
alleles at microsatellite loci in CEPH families mainly arise from somatic
mutations in the lymphoblastoid cell line. Human Mutation 3:365-372.
Barnosky, C.W., P.M. Anderson and P.J. Bartlein. 1987. The northwestern U.S.
during deglaciation; vegetational history and paleoclimatic implications. In
Geology of North America. North America and Adjacent Oceans During the
p
56
Last Deglaciation (Eds. W.F. Ruddiman and H.E. Wright, Jr.), Vol. K-3, pp.
289-321. Geological Society of America, Boulder.
Beerli, P. 1998. Estimation of migration rates and population sizes in geographically
structured populations. In: Advances in molecular ecology (Ed. G. Carvalho),
pp. 39-53. NATO-ASI workshop series. lOS Press, Amsterdam.
Beerli, P. 2004. Effect of unsampled populations on the estimation of population
sizes and migration rates between sampled populations. Molecular Ecology
13:827-836.
Beerli, P. and J. Felsenstein. 1999. Maximum likelihood estimation of migration
rates and population numbers of two populations using a coalescent approach.
Genetics 152:763-773.
Beerli, P. and J. Felsenstein. 2001. Maximum likelihood estimation of a migration
matrix and effective population sizes in n subpopulations by using a coalescent
approach. Proceedings of the National Academy of Sciences 98:4563-4568.
Bittner, T.D. and R.B. King. 2003. Gene flow and melanism in garter snakes
revisited: a comparison of molecular markers and island vs. coalescent
models. Biological Journal of the Linnean Society 79:389-399.
Brede, E.G. and T.J.C. Beebee. 2004. Contrasting population structures in two
sympatric anurans: implications for species conservation. Heredity 92:110117.
Bronikowski, A.M. and S.J. Arnold. 2001. Cytochrome b phylogeny does not match
subspecific classification in the western terrestrial garter snake, Thamnophis
elegans. Copeia 2001 :508-513.
Crow, J.F. and M. Kimura. 1971. The effective number of a population with
overlapping generations: a correction and further discussion. American
Journal of Human Genetics 24:1-10.
Crow, J.F. and N.E. Norton. 1955. Measurement of gene frequency drift in small
populations. Evolution 9:202-214.
Dallas, J.F. 1992. Estimation of microsatellite mutation rates in recombinant inbred
strains of mouse. Mammalian Genome 5:32-38.
de Queiroz, A., R. Lawson and J.A. Lemos-Espinal. 2002. Phylogenetic relationships
of North American garter snakes (Thamnophis) based on four mitochondrial
genes: how much DNA sequence is enough? Molecular Phylogenetics and
Evolution 22:315-329.
Edwards, A., H.A. Hammond, L. Jin, C.T. Caskey and R. Chakraborty. 1992.
Genetic variation at five trimeric and tetrameric tandem repeat loci in four
human population groups. Genomics 12:241-253.
Ellegren, H. 1995. Mutation rates at porcine microsatellite loci. Mammalian Genome
6:376-377.
Excoffier, L., P.E. Smouse and J.M. Quattro. 1992. Analysis of molecular variance
inferred from metric distances among DNA haplotypes: application to human
mitochondrial DNA restriction data. Genetics 131:479-491.
Farr, D.R. and P.T. Gregory. 1991. Sources of variation in estimating litter
characteristics of snakes. Journal of Herpetology 25:261-267.
-
57
Fitch, H.S. 1987. Collcting and life-history techniquest. In Snakes: Ecology and
Evolutionary Biology (Eds. R.A. Seigel, J.T. Collins and S.S. Novak), pp. 143164. McGraw-Hill, New York.
Garner, T.W.J., P.T. Gregory, G.F. McCracken, G.M. Burghardt, B.F. Koop, S.E.
McLain and R.J. Nelson. 2002. Geographic variation of multiple paternity in
the common garter snake (Thamnophis sirtalis). Copeia 2002:15-23.
Geyer, C. and E.A. Thompson. 1994. Annealing Markov chain Monte Carlo with
Applications to Ancestral Inference. Technical report, University of
Minnesota Nr. 589 R(1).
Gibbs, H.L., K.A. Prior, P.W. Weatherhead and G. Johnson. 1997. Genetic structure
of populations of the threatened eastern massasauga rattlesnake, Sistrurus c.
catenatus: evidence from microsatellite DNA markers. Molecular Ecology
6:1123-1132.
Gregory, P.T. and K.W. Stewart. 1975. Long-distance dispersal and feeding strategy
of the red-sided garter snake (Thamnophis sirtalis parietalis) in the Interlake of
Manitoba. Canadian Journal of Zoology 53:238-245.
Gregory, P.T. 1977. Life-history parameters of the red-sided garter snake
(Thamnophis sirtalis parietalis) in an extreme enviromnent, the Interlake
region of Manitoba. Publications de Zoologie, National Museum of Canada
13: 1-44.
Guo, S.W. and E.A. Thompson. 1992. Performing the exact test for Hardy-Weinberg
proportions for multiple alleles. Biometrics 48:36 1-372.
Hamilton, M.B., E.L. Pincus, A. DiFiore and R.C. Fleischer. 1999. Universal linker
and ligation procedures for construction of genomic DNA libraries enriched
for microsatellites. Biotechniques 27:500-507.
Hanski, I. and D. Simberloff. 1997. The metapopulation approach, its hisotry,
conceptual domain, and application to conservation. In Metapopulation
Biology (Eds. l.A. Hanski and M.E. Gilpin), pp. 5-26. San Diego: Academic
Press.
Hoffman, E.A., W.R. Ardren and M.S. Blouin. 2003. Nine polymorphic
microsatellite loci for the northern leopard frog (Rana pipiens). Molecular
Ecology Notes 3:115-116.
Hoffman, E.A., F.W. Schueler and M.S. Blouin. 2004. Effective population sizes and
temporal stability of genetic structure in Rana pipiens, the northern leopard
frog. Evolution 5 8:2536-2545.
Hutchison, D.W. and A.R. Templeton. 1999. Correlation of pairwise genetic and
geographic distance measures: inferring the relative influences of gene flow
and drift on the distribution of genetic variability. Evolution 53:1898-1914.
Janzen, F.J., J.G. Krenz, T.S. Haselkorn, E.D. Brodie, Jr. and E.D. Brodie, III. 2002.
Molecular phylogeography of common garter snakes (Thamnophis sirtalis) in
western North America: implications for regional historical forces. Molecular
Ecology 11:1739-1751.
Kephart, D.G. 1981. Population ecology and population structure of Thamnophis
elegans and Thamnophis sirtalis. Ph.D. dissertation, Dept. of Biology,
University of Chicago.
Kephart, D.G. 1982. Microgeographic variation in the diets of garter snakes.
Oecologia 52:287-291.
Kephart, D.G. and S.J. Arnold. 1982. Garter snake diets in a fluctuating environment:
a seven-year study. Ecology 63:1232-1236.
Lawson, R. and R.B. King. 1996. Gene flow and melanism in Lake Erie garter snake
populations. Biological Journal of the Linnean Society 59:1-19.
Legendre, P. 2001. Congruence among distance matrices: Program CADM user's
guide. Département de sciences biologiques, Université de Montréal. 7 pages.
Macey, J.R., Schulte, J.A., II., N. B. Ananjeva, A. Larson, N. Rastegar-Pouyani, S.J.
Shanimakov and T.J. Papenfuss. 1998. Phylogenetic relationships among
agamid lizards of the Laudakia caucasia species group: testing hypotheses of
biogeographic fragmentation and an area cladogram for the Iranian Plateau.
Molecular Phylogenetics and Evolution 10:11 8-131.
Manly, B.F. 1997. Randomization and Monte Carlo methods in biology. 2' ed.
Chapman and Hall, New York.
Mantel, N. 1967. The detection of disease clustering and generalized regression
approach. Cancer Research 27:209-220.
Mantel, N. and R.S. Valand. 1970. A technique of nonparametric multivariate
analysis. Biometrics 26:547-558.
McCracken, G.F., G.M. Burghardt and S.E. Houts. 1999. Microsatellite markers and
multiple paternity in the garter snake Thamnophis elegans. Molecular Ecology
8:1475-1479.
McMillen-Jackson, A.L. and T.M. Bert. 2003. Disparate patterns of population
genetic structure and population history in two sympatric penaeid shrimp
species (Farfantepenaeus aztecus and Litopenaeus setferus) in the eastern
United States. Molecular Ecology 12:2895-2905.
Michels, E., E. Audenaert, R. Ortells, L. De Meester. 2003. Population genetic
structure of three pond-inhabiting Daphnia species on a regional scale
(Flanders, Belgium). Freshwater Biology 48:1825-1839.
Molbo, D., C.A. Machado, E.A. Herre, L. Keller. 2004. Inbreeding and population
structure in two pairs of cryptic fig wasp species. Molecular Ecology 13:16 131623.
Moore, M. and J. Lindzey. 1992. The physiological basis of sexual behavior in male
reptiles. In C. Gans and D. Crews (eds.), Biology of the Reptilia, vol. 18, pp.
70-113. University of Chicago Press, Chicago, Illinois.
Ohta, T. and M. Kimura. 1973. A model of mutation appropriate to estimate the
number of electrophoretically detectable alleles in a finite population.
Genetical Research 22:20 1-204.
Prior, K.A., H.L. Gibbs and P.J. Weatherhead. 1997. Population genetic structure in
the black rat snake: implications for management. Conservation Biology
11:1147-1158.
w
Prosser, M.R., H.L. Gibbs, P.J. Weatherhead. 1999. Microgeographic population
genetic structure in the northern water snake, Nerodia siedon sipedon
detected using microsatellite DNA loci. Molecular Ecology 8:329-333.
Pullium, H.R. 1988. Sources, sinks, and population regulation. American Naturalist
132:653-661.
Raymond, M. and F. Rousset. 1995. GENEPOP Version 1.2: population genetics
software for exact tests and ecumenicism. Journal of Heredity 86:248-249.
Rice, W.R. 1989. Analysing tables of statistical tests. Evolution 43:223-225.
Rossman, D.A., N.B. Ford and R.A. Seigel. 1996. The garter snakes: evolution and
ecology. Norman, Oklahoma: University of Oklahoma Press.
Rousset, F. 1997. Genetic differentiation and estimation of gene flow from FStatistics under isolation by distance. Genetics 145:1219-1228.
Rüber, L., A. Meyer, C. Sturmbauer and E. Verheyen. 2001. Population structure in
two sympatric species of the Lake Tanganyika cichlid tribe Eretmodini:
evidence for introgression. Molecular Ecology 10:1207-1225.
Rychlik, W. 1998. Oligo Primer Analysis Software v. 6 for Mac. Molecular Biology
Insights, Inc. Cascade, CO.
SAS Institute. 2002. SAS/STAT software user's guide. Re!. 9.1. SAS Institute, Inc.,
Cary, NC.
Schneider, S., D. Roessli and L. Excoffier. 2000. Ar!equin ver 2.000: a software for
population genetics data analysis. Genetics and Biometry Laboratory,
University of Geneva, Switzerland.
Slatkin, M. and L. Excoffier. 1996. Testing for linkage disequilibrium in genotypic
data using the EM algorithm. Heredity 76:377-383.
Stacey, P.B., V.A. Johnson and M.L. Taper. 1997. Migration within
metapopulations: the impact upon local population dynamics. In
Metapopulation Biology (Eds. l.A. Hanski and M.E. Gilpin), pp. 267-292. San
Diego: Academic Press.
Stebbins, R.C. 2003. Western amphibians and reptiles. Houghton Mifflin Co., New
York.
Weber, J.L. and C. Wong. 1993. Mutation of human short tandem repeats. Human
Molecular Genetics 2:1123-1128.
Whittier, J.M. and R.R. Tokarz. 1992. Physiological regulation of sexual behavior in
female reptiles. In C. Gans and D. Crews (eds.), Biology of the Reptilia, vol.
18, pp. 24-69. University of Chicago Press, Chicago, Illinois.
Wright, S. 1931. Evolution in Mendelian populations. Genetics 16:97-159.
Zardoya, R., R. Castilho, C. Grande, L. Favre-Krey, S. Caetano, S. Marcato, G. Krey
and T. Patamello. 2004. Differential population structuring of two closely
related fish species, the mackerel (Scomber scombrus) and the chub mackerel
(Scomberjaponicus), in the Mediterranean Sea. Molecular Ecology 13:17851798.
ECOLOGICAL CORRELATES OF POPULATION GENETIC STRUCTURE: A
COMPARATIVE APPROACH USING A VERTEBRATE METACOMMUNITY
CHAPTER 3
Mollie K. Manier and Stevan J. Arnold
This thesis chapter has been prepared for submission to the journal American
Naturalist.
61
ABSTRACT
Determining the ecological and evolutionary causes of population genetic
differentiation is important for understanding microevolutionary processes. This goal
can be achieved through comparative studies of different species with predator-prey or
competitive interactions across multiple populations in a common landscape. Our
study species were two coexisting garter snakes (Thamnophis elegans and T sirtalis)
and one of their prey species, the western toad (Bufo boreas). We know from previous
research that populations of both garter snakes species exhibit source-sink dynamics,
and we use a comparison with the western toad to evaluate the generality of such
population structure within the study system. We found substantially different
population dynamics for B. boreas that were characterized by much larger effective
population sizes, lower migration rates and migration-drift equilibrium, suggesting a
stable population structure. We also identified a series of block faults that separated
genetically differentiatied population clusters, serving as a barrier to dispersal.
Relative to their representation, biotic interactions and nearest neighbor characteristics
were less important than habitat descriptors such as perimeter of water bodies and
elevation in explaining differences in population genetic parameters (effective
population size, migration rate, genetic distance). Presence of other species did not
influence effective population size of any species. Instead, effective size was
accounted for by habitat or nearest neighbor variables. Overall, migration rate was
determined by habitat features and biotic interactions. In particular, migration
occurred from sites with higher densities to sites with lower densities of both the focal
62
and interacting species. Genetic distance was explained largely by geographic
distance.
63
INTRODUCTION
Population genetic differentiation by random genetic drift or response to divergent
local selection can have a significant impact on the microevolutionary dynamics of a
region and can be an evolutionary precursor to speciation. Identifying ecological and
evolutionary factors that control local population differentiation is critical to
understanding fine-scale microevolutionary processes.
Exploration of how ecological factors influences population genetic
differentiation has focused on the effects of spatial structure. These studies generally
use spatial analyses of isolation by distance and autocorrelation but ignore landscape
features (e.g., Arnaud et al. 2001, Gamier et al. 2004, Schweiger et al. 2004, Van
Rossum et al. 2004, Whiteley et al. 2004). A few authors have incorporated dispersal
variables into the calculation of geographic distance. A resulting isolation by distance
analysis then evaluates the relationship between genetic distance and a more
ecologically relevant measure of geographic distance (Keyghobadi et al. 1999, Castric
et al. 2001, Roach et al. 2001, Coulon Ct al. 2004, Eriksson et al. 2004). Over the last
15 years, a few studies have included specific landscape features such as altitude
(Castric et al. 2001), climate (Nevo et al. 1998), vegetation characteristics, land use
(Reh and Seitz 1990), and general habitat descriptors (Scribner and Chesser 1993,
Scribner et al. 2001). A few studies have also studied the effect of population biology
(e.g., persistence) on genetic structure (Scribner and Chesser 1993, Roach et al. 2001).
64
The present study contributes to the field of ecological genetics by putting
more emphasis on the ecological determinants of differentiation. We use a
comparative approach to investigate the association of habitat features and abundance
of coexisting species with population genetic parameters for three vertebrates
inhabiting a common landscape. Previous research has identified metapopulation
dynamics for two of these species, and a third is used as an additional comparison to
examine the generality of source-sink metapopulation dynamics in the region. The
focal species were the terrestrial garter snake (Thamnophis elegans), the common
garter snake (Thamnophis sirtalis), and the western toad (Bufo boreas). These three
species occupy a series of ponds, lakes and flooded meadows in Lassen County,
California. Previous research revealed significant population genetic differentiation
and isolation by distance for both garter snakes in this study system (Chapter 2).
Genetic distances between population pairs at sites that supported both snake species
were also found to be weakly correlated. Similarities in genetic structure between
these two species may be a function of a correlated response to the same landscape,
their phylogenetic relatedness, or some interaction thereof. If the shared landscape
played a larger role in the observed pattern, one would expect an unrelated species
coexisting on the landscape to have a similar pattern of genetic differentiation and
isolation by distance. Alternatively, if the garter snake population genetic structures
were more a result of common evolutionary processes, one would expect an unrelated
species to have an independent pattern of genetic differentiation. Consequently, a
secondary goal of the study was to determine if correlated patterns of genetic
65
N
BLW
JCK
UGV
STF
LTC
MOS
BUL
0
p rGOR
CLG
SUM
ASH
RKY
\
k\ç'\
/
Eagle Lake
I
PVM
I?
AMP
DNS
oFEA
PAP
RON
MAH
NML
MCY
5
Figure 3.1. Map of the study area showing sampled sites.
differentiation for both species were indicative of correlated responses to the same
ecological variables or more a function of their shared phylogeny. To address this
question, the western toad, Bufo boreas was included as a third species for
comparison. This species was chosen because of its ecological similarity to the garter
snakes. Bufo. boreas uses the same ponds, lakes and flooded meadows occupied by
both garter snakes for breeding habitat and its tadpoles and metamorphs are eaten by
both snake species (Kephart 1982, Kephart and Arnold 1982).
We looked for physical barriers to dispersal using a clustering analysis that
detected genetically similar groups of geographically proximate populations. We also
used stepwise regression to identify ecological factors that influence genetic
differentiation as expressed by effective population size, migration rate and genetic
distance for each species. Explanatory variables used in the analyses described habitat
features, biotic interactions and spatial structure.
METHODS
Genetic analysis for Bufo boreas
Genetic analysis of microsatellite variation for T. elegans and T sirtalis populations
has been reported in Chapter 2. Analysis of B. boreas microsatellite data followed the
same procedures, with the few exceptions outlined below.
67
We collected a total of 150 tissue samples from six adult B.
boreas
populations
within a 1000 square kilometer area in and around the Eagle Lake basin in Lassen Co.,
California. For each toad, the tip a hind toe was clipped and stored in Drierite®, an
anhydrous calcium sulfate desiccant. Additionally, 77 B.
boreas
tadpoles and newly
metamorphosed toads were collected and analyzed. These samples were excluded
from further analysis, because they showed some significant deviations from Hardy-
Weinberg equilibrium in the direction of excess homozygosity. Simandle (2005)
provided all genotypic data for B.
boreas
at Pikes Point and shared 16 microsatellite
primer sequences used for the remaining populations. Procedures used for DNA
extraction, amplification and genotyping are as described in Chapter 2. PCR profiles
consisted of 94 °C for 2 mm followed by 36 cycles of 94 °C for 30 sec, 55 °C for 30
sec and 72 °C for 30 sec, ending with 72 °C for 2 mm.
All statistical analyses for B.
boreas
genetic data were as described in Chapter
2. They included exact tests for departure from Hardy-Weinberg equilibrium (Guo
and Thompson 1992; Markov chain parameters: 5000 dememorizations; 500 000
steps per chain) calculated in ARLEQUIN v. 2.000 (Schneider et al. 2000) and tests for
linkage disequilibrium (Slatkin and Excoffier 1996; Markov chain parameters: 5000
dememorizations, 1000 batches, 5000 iterations per batch), performed in GENEPOP
(Raymond and Rousset 1995). Significance levels were adjusted for multiple
comparisons (Rice 1989). Number of alleles and observed and expected
heterozygosities in each population and over all populations were calculated in
GENEPOP (Raymond and Rousset 1995).
Overall and population pairwise estimates of FST were calculated using AMOVA
(Excoffier et al. 1992) in ARLEQUIN v. 2.000. Significance was assessed after 16 000
permutations for global estimates and 3000 permutations for pairwise estimates, with
P-values adjusted with the sequential Bonferroni correction. We regressed genetic
distance
(FSTI(l-FST);
Rousset 1997) on log geographic distance to look for patterns of
migration-drift equilibrium (Hutchison and Templeton 1999). A Mantel test (Mantel
1967, Mantel and Valand 1970, Manly 1997), implemented in ARLEQUIN v. 2.000 was
used to estimate the correlation between genetic distance and log geographic distance
(significance over 10 000 permutations).
Estimates of effective population size and migration rate were obtained with
MIGRATE v. 1.7.6.1 (Beerli and Felsenstein 2001). Assumptions, protocols and
parameters used for the analysis are as outlined in Chapter 2. Effective population
size was expressed as & = 4Nu, where N is effective population size and 1u is
mutation rate. Assuming a mutation rate typical for vertebrates of 1 0 per locus per
generation (Dallas 1992, Edwards et al. 1992, Weber and Wong 1993, Banchs et al.
1994,
Ellegren
1995), Ne
as obtained from 9. Estimates of migration were expressed
as 4Nem, where m is migration rate, and Ne refers to the receiving population. Using
Ne
estimates from the previous step, m from each population into all other populations
was calculated.
Barriers to gene flow
A total of 24 water bodies that support varying combinations of T. elegans, T sirtalis
and B. boreas were analyzed for spatial clustering at microsatellite loci for each
species (Table 3.1). Clusters and hence potential barriers to dispersal were identified
by partitioning the genetic variation among populations using AMOVA. The program
SAMOVA 1.0 (Dupanloup et al. 2002) was used to obtain a starting configuration of
groups of populations. For each species, SAMOVA used decimal degree latitude and
longitude for each population (obtained using Maptech Terrain Navigator v. 3.02) to
create a user-defined number of maximally differentiated groups of geographically
proximate populations. Because the missing value threshold for rejecting a locus was
set relatively low (5%) in SAMOVA, we tested the proposed groups again using
ARLEQUIN v. 2.000 (rejection threshold of 50%). In this way, we used SAMOVA to
generate hypotheses about population clusters and tested their veracity in ARLEQUIN.
For each hypothesis, we confirmed that the proposed groupings were spatially realistic
and made necessary adjustments accordingly. The F-statistic analogs,
IST,
cDsc and
(Dc1 were estimated for each hypothesis, and their significance levels were determined
with 16 000 pennutations (Excoffier et al. 1992). DST represents the correlation
between randomly drawn pairs of alleles within populations relative to the whole array
of populations.
CT
is the correlation between randomly drawn pairs of alleles within
a group of populations relative to the whole array of populations, and Dsc is the
correlation between randomly drawn pairs of alleles within populations relative to a
group of populations (Excoffier et al. 1992).
70
Table 3.1. Names, abbreviations, latitude, longitude, perimeter (km), type (M =
meadow, L = lake, LS = lakeshore), elevation (m), sampling effort and sample sizes of
study sites and populations. Sampling effort is the number of collector trips to a
population from 1999 to 2004. N is the number of individuals caught from 1999 to
2004. d and n indicate sampling during the day for snakes and at night for toads. N is
the number of individuals used in the micro satellite analysis. For populations where N
exceeds N, tissue samples collected in previous years were used in the genetic
analysis. B = B. boreas, E = T. elegans and S = T. sirtalis.
Type
N
Code
Lat.
Long.
Antelope Mountain Pond*
Ashurst Lake
Blue Water
Bullard Lake
Cleghorn Reservoir
Colman Lake
Deans Meadow
Feather Lake
Gallatin Shoreline*
Gordon Lake
Jacks Lake
Little Cleghorn Reservoir
Mahogany Lake
McCoy Flat Reservoir
Mosquito Flat
Nameless Meadow*
AMP
ASH
BLW
40.614
40.750
40.834
40.775
40.777
40.516
40.557
40.542
40.562
40.768
40.810
40.787
40.534
40.453
40.771
40.524
40.528
40.557
40.619
40.684
40.511
40.803
40.766
40.831
-120.923
1920
0.52
M
15
0
-120.965
-120.919
-120.901
-120.804
-120.714
-120.719
-121.018
-120.760
-120.882
-121.025
-120.794
-120.732
-120.940
-120.943
-120.743
1950
3.5
M
7
1770
0.425
L
9d,4n
lid, IOn
1860
3.3
1880
3.6
1965
2.2
1980
0.95
1740
5.3
1575
2.3
1850
3.1
1690
2
1880
3
2065
1700
1890
1.7
18.9
1915
1.5
1645
5
1555
3.2
1730
9.1
1580
4.1
M
M
L
L
L
LS
M
L
M
L
M
M
M
M
LS
M
LS
1825
1.2
L
1870
1890
0.458
2.5
L
M
1920
0.2
M
Papoose Meadows
Pikes Point
Pine Valley Meadow*
Rocky Point
RoneyCorral
Camp Stanford
Summit Lake
Upper Gooch Valley Pond*
MAll
MCY
MOS
NML
PAP
P1K
PVM
RKY
RON
STF
SUM
UGV
-120.757
-120.784
-120.969
-120.757
-120.857
-120.932
-120.839
-120.874
1.6
E
B
Site
BUL
CLG
COL
DNS
FEA
GAL
GOR
JKS
LTC
Perim.
(km)
Samp.
Effort
Elev.
(m)
S
N
N
N
N
32
30
20
19
---
30
30
39
38
18
18
25
27
2
13
0
67
1
28
0
18
1
15
0
27
4
0
9
0
28
67
44
24
26
--56
--27
35
15
15
0
-------------------
40
18
8
39
0
167
91
94
25
0
20
16
3
2d, 2n
24
----24
9
I
33
0
---
95
--29
30
45
7
---
306
140
16
53d, 4n
32
32
108
48
0
9
0
72
70
83
21
0
13
19
1
28d,8n
8d,8n
28
----28
29
15
31
32
13
0
---
24
45
27
25
15
26
47
27
27
22
7d, 8n
33
33
0
---
0
0
N
34
49
32
3
113
2
35
4
35
27
29
25
27
24
0
43
42
1
26
29
83
--
72
Ecological analysis. Multiple linear regression was used to identify ecological
variables that best predicted population differentiation
(FST),
effective population size
Table 3.2. Abbreviations and descriptions of variables used in the multiple
regression analysis. All census sizes are adjusted according to sampling effort.
Abbreviation
PERIM
TYPE
ELEV
BUFO
ELEG
SIRT
SNAKES
NN-DIST
NN-PERIM
NN-TYPE
NN-ELEV
NN-BUFO
NN-ELEG
NN-SIRT
NN-SNAKES
PERIMDIFF
TYPEDIFF
ELEVDIFF
BUFODIFF
ELEGDIFF
SJIRTDIFF
SNAKEDIFF
DIST
Variable description
Site perimeter (km)
Site type (1 = meadow, 2 = lake, 3 = lakeshore)
Elevation (m)
Census size of B. boreas
Census size of T. elegans
Census size of T. sirtalis
Summed census size of T. elegans and T sirtalis
Distance to nearest neighbor (km)
Nearest-neighbor site perimeter (km)
Nearest neighbor site type (1 = meadow, 2 = lake, 3 = lakeshore)
Nearest neighbor elevation (m)
Nearest neighbor B. boreas census size
Nearest neighbor T. elegans census size
Nearest neighbor T sirtalis census size
Nearest neighbor summed census size of T. elegans and 7'. sirtalis
Difference in perimeter between two sites (kin)
Difference in type between two sites
Difference in elevation between two sites (m)
Difference in B. boreas census sizes between two sites
Difference in T. elegans census sizes between two sites
Difference in 7'. sirtalis census sizes between two sites
Difference in summed 7'. elegans and T. sirtalis census sizes
between two sites
Distance between sites (km)
(Ne) and one-way migration rate (m). These values for B. boreas were obtained from
this study as described above, and those for T. elegans and 7'. sirtalis are reported in
f.
73
Chapter 2. Independent variables used in the analyses varied according to the
dependent variable and species and are described in Table 3.2. The independent
variables described ecological characteristics that centered around a core theme
consisting of site perimeter (km), site type, census size of a focal and/or interacting
species, and the logarithm of intersite or nearest neighbor distance. Variations on
these basic parameters focused on their values for a site's nearest neighbor and the
absolute or actual difference between those values for two sites. Site perimeter was
defined as the circumference of a pond or meadow or the length of Eagle Lake
shoreline and was measured in km. Perimeter was chosen as a biologically relevant
measure of size, because B.
boreas breeds
in shallow water, and the garter snakes forage
primarily near the water's edge. Site perimeter, elevation (m; Table 3.1) and pairwise
distances between study sites (km; Table 3.3) were estimated using Maptech Terrain
Navigator v. 3.02. Site type was characterized as meadow, lake or lakeshore based on
average depth and resulting degree of annual change in water level. Thus, some
shallow water lakes were classified as meadows. Numeric values for site type were
substituted to represent increasing order of depth and permanence, according to the
following: meadow = 1, lake =2 and lakeshore =3. Lakeshore type corresponds to
shorelines of Eagle Lake, a consistently deep and permanent body of water. Census
size was measured as the total number of individuals caught over a six-year period
between 1999 and 2004, adjusted by sampling effort, defined as the total number of
collectors summed over all sampling trips. The final value represented number of
individuals caught at each site per person-sampling event. Census sizes normalized in
Table 3.3. Geographic distance matrix of pairwise distances between populations, measured in km.
ANT ANS ASH BLW BUL CLG COL DNS FEA GAL GOR JKS LTC MAH
3.9
ANS
ASH
14.5
15.3
BLW 24.2 24.4 10.1
18.3 17.9
BUL
6.1
6.8
22.4 20.7 13.9 11.6
CLG
8.2
COL 24.4 20.8 33.5 39.4 32.8
30
22.2 18.5 29.8 35.1 28.6 25.4 4.6
DNS
FEA
9.5
11.4 23.5 33.7 27.7 31.9 25.6 25.4
GAL
18.8
15
27.1
33.2 26.4 24.1
6.4
3.5
22
GOR 18.5 17.4 7.2
8
1.7
6.8
31.4 27.2 27.6 25.1
21.7 23.4 8.5
JKS
11.2
9.4
19
41.9 38.2 29.8 35.5 12.9
LTC
23.7 22.5 14.9 11.8
9.1
1.4
30.8 26.3 33.2 25.1
7.7 19.6
MAR 22.1 18.6 31.2 36.9 30.3 27.7 2.5
2.8 24.2
3.9
28.9 39.5 28.6
MAR 24.9 24.5 11.8
3.8
6.7
8.5
37.5 33.1 34.4 31.4
7.1
12.9 8.4
35.2
MCY 18.6
18
33.1
42.4 22.3 37.8 20.5 22.1 11.9 19.5 35.3 40.3 39.1
19.9
MOS
17
17.4
3
7.4
3.6
11.8 34.3 30.4 26.2 27.9
5.2
8.2 12.6 31.8
NIML 21.8
18.3 31.2 37.6 30.9 28.5
2.6
4.2 23.4 4.4
29.6 39.7 29.5
1.4
PAP
20.6 17.1 30.2 36.6
27.9
4
30
4.6 22.2 3.8
28.7 38.7 28.9
2.2
P1K
17.1
13.4 26.3 32.9 26.1 24.5
7.5
5.5
20
2.1
24.8 34.7 25.6
5.1
RKY
19.3
16.1
15.7
11
19.1
18.9 21.5
14.5 27.2 13.6
14.1 26.6 11.8
16.9
RON
15.3 12.8
28
36.3 29.6 29.9 12.2 12.8 14.1
9.9
28.6 36.2 31.1
10.9
STF
20.5 20.9 6.5
3.7
4
11.1
36.8 32.7
30
30.5
5.7
8
11.7
34.4
SUM
19.7 18.2 10.7 10.2
29.6 25.3 29.1 23.6
5.3
3.2
3.7
16.5 4.4
27.3
MAR MCY
42.4
8.9
35.9
35.1
31.4
19.1
35.6
5.8
7.8
35.3
18.5
17.7
17.6
31.1
9.6
39
35.8
75
Table 3.3. (continued)
MOS NML PAP P1K RKY RON STF
NML
PAP
P1K
32.2
31.2
27.2
RKY
RON
STF
SUM
18.4
29.8
3.7
8.9
1.3
5.1
17.8
9.8
34.9
28
4
17.3
8.8
34
27.3
14.3
8
21
30
23.6
19.8
11.4
20.6
28.3
9
this way were used instead of effective population sizes, because census size is a more
ecologically-relevant factor, whereas effective size reflects historical processes that
have occurred on an evolutionary time scale.
The interacting species variable differed according to focal species. For T
elegans and T sirtalis, the interacting species was its competitor, T. sirtalis or T.
elegans, respectively. Bufo boreas was not included as an interacting prey species,
because both garter snakes consume other types of prey as well. For B. boreas, the
interacting species were both garter snake predators, and their census sizes were
summed. Study sites whose nearest neighbor was not sampled for genetic variation
were excluded from the analysis (Feather Lake, Jacks Lake and McCoy Flat
Reservoir). Table 3.4 shows the full model for each analysis. A stepwise regression
was used to determine the final model.
For multiple regression analyses with pairwise genetic distance or migration
rate as the dependent variable, the response is a matrix of distances or dissimilarities.
Elements of the genetic distance matrix, representing a population pair, were
Table 3.4. Full models used in each of nine stepwise regression analyses testing for ecological effects on effective population size,
migration rate and genetic distance for each species. Variables in bold were selected by stepwise regression.
PERIM
TYPE
ELEV
NN-DIST ELEG
SIRT
NN-PERIM NN-TYPE
ElegNe
SirtNe
PERIM
TYPE
ELEV
NN-DIST SIRT
ELEG
NN-PERIM NN-TYPE
BufoNe PERIM
TYPE
ELEV
NN-DIST BUFO
SNAKES
NN-PERIM NN-TYPE
X9
NN-ELEV
NN-ELEV
NN-ELEV
Y
Elegm
Sirtm
Bufom
Y
X1O
Xli
NN-SIRT
NN-ELEG
NN-SNAKES
NN-ELEG
NN-SIRT
NN-BUFO
Xi
X2
PERIMDIFF TYPEDIFF
X3
X4
X5
X6
PERIMDIFF TYPEDIFF
DIST
DIST
ELEYDIFF DIST
ELEGDIFF SIRTDIFF
SIRTDIFF ELEGDIFF
BUFODIFF SNAKEDIFF
Xl
X3
X5
ELEGDIFF
SIRTDJFF
X6
SIRTDIFF
BUFODIFF
SNAKEDIFF
PERIMDIFF
TYPEDIFF
X2
ElegDist PERIMDIFF TYPEDIFF
SirtDist
PERIMDIFF TYPEDIFF
BufoDist PERIMDIFF TYPEDIFF
ELEVDIFF
ELEVDIFF
X4
ELEVDIFF DIST
ELEVDJFF DIST
ELEVDIFF DIST
ELEGDIFF
77
expressed as F51/(1-
FST),
as suggested by Rousset (1997). Migration rate, m,
represents the proportion of one population that originated from a different population,
and is therefore actually a measure of similarity. This value was converted to a
measure of dissimilarity, the proportion of residents, p = 1
1995). For both FST/(l-
FST)
m (Sokal and Rohlf
andp, the higher the value of any element in the matrix,
the higher the degree of dissimilarity between the two populations represented by that
element.
The regression model for each of these response matrices describes ecological
dissimilarities between the two sites by asking if differences in certain features were
important in predicting population genetic differentiation or migration rate. The
explanatory variables were therefore also matrices that described differences between
population pairs for site perimeter, type, elevation, census size of both the focal and
interactive species, as well as pairwise log geographic distance. Explanatory matrix
variables of the genetic distance response matrix represented absolute differences
between population pairs, because FST/(
distance between two populations.
In
1- Fs1)
characterizes the overall genetic
contrast, explanatory matrix variables of the p
response matrix contained information on the directionality of the difference. In other
words, instead of describing the overall difference between population A and
population B (an absolute value), they show which one is larger (a positive or negative
value). The matrices for the p analysis are thus asymmetrical full matrices of bidirectional dissimilarities, whereas those for the genetic distance analysis are
triangular or symmetric. Significance of the coefficients and associated R2 were
determined using a permutation method specific for distance matrices (Legendre et al.
1994). Multiple regression analyses using effective population size as the response
variable were performed in SAS (v. 9.2; SAS Institute 2002), and those using
migration or genetic distance matrices were implemented using Permute! (v. 3.4 alpha
9; Legendre et al. 1994).
RESULTS
Population genetics of B. boreas
Only one of 16 B. boreas loci (1281) deviated significantly from Hardy-Weinberg
equilibrium (HWE) in one population (Mosquito Flat) after sequential Bonferroni
correction (Table 3.5). All loci were in linkage equilibrium at all populations.
Average observed and expected heterozygosities for all B. boreas loci are shown in
Table 3.5 and for all populations in Table 3.6. The total number of alleles overall
found at a locus varied from six to 27, while number of alleles per population at a
single locus ranged from seven to ten. Population averages for number of alleles over
all loci ranged from four to 16. All B. boreas populations had at least one private
allele, with 11 in the Upper Gooch Valley pond population. Of those, seven were
from the T29 locus.
Genetic differentiation of B. boreas populations was low but highly significant,
with a global FST of 0.024 (P < 0.00001). Pairwise estimates and their significance
79
levels are shown in Table 3.7. After sequential Bonferroni correction, five out of 15
population pairs for B. boreas were found to be significantly differentiated.
Populations at Pikes Point and Roney Corral were genetically dissimilar from those at
Blue Water and Upper Gooch Valley pond. Pikes Point was
Table 3.5. Comparison of the genetic diversity found at all microsatellite
loci for B. boreas averaged over all populations. For each locus, total
number of alleles, allele size range, observed heterozygosity (H0), expected
heterozygosity (He), average number of alleles per population (Na) and
number of populations deviating from Hardy-Weinberg equilibrium are
shown.
Locus
B34-2
B4-2
B45
F16
F36
F4
117
1201
T233
T281
T29
T292
T293
T297
T86
T87b
Total
alleles
9
7
12
19
15
14
9
7
6
14
15
14
9
15
7
27
Allele size
range (bp)
179-215
220-244
140-175
173-237
210-257
173-229
119-145
123-167
208-226
136-182
88-101
97-158
145-182
106-129
150-172
133-254
H0
0.78
0.61
0.75
0.80
0.75
0.79
0.48
0.35
0.40
0.62
0.50
0.81
0.76
0.74
0.75
0.84
He
0.81
Na
0.76
0.84
0.89
0.90
0.85
0.56
0.45
0.57
0.87
0.54
0.87
0.82
0.83
0.79
0.92
5
Pops. out of
HWE
6
4
4
0
0
0
0
0
0
0
0
0
11
1
8
10
6
0
0
0
0
0
16
0
7
9
12
10
11
11
7
also differentiated from Mosquito Flat. We found strong evidence for a pattern of
isolation by distance among B. boreas populations. The regression ofFST/(1-FST) on
log geographic distance shows a pattern congruent with a hypothesis of migration-drift
equilibrium (Hutchison and Templeton 1999; Fig. 3.2), with a slope of 0.028. There
was also a significant correlation between the geographic and genetic distance
matrices according to a Mantel test (r
= 0.678; P = 0.0 16).
Estimates of effective population size for B.
boreas
were relatively high with an
average 0 of 0.497 (Table 3.8). Assuming a microsatellite mutation rate of 10,
which is typical for vertebrates (Dallas
1992,
Edwards et al.
1992,
Weber and Wong
Table 3.6. Study populations of B. boreas and their
sample sizes (N), average observed heterozygosity
(H0), average expected heterozygosity (He) and
average number of alleles per locus (Na).
Population
N
H0
He
Na
BLW
0.79
8
18
0.66
0.74
9
MOS
24
0.65
1993,
1240
P1K
32
RON
0.72
0.76
9
28
0.63
0.73
8
STF
15
0.67
0.80
7
UGV
33
0.70
0.77
10
Banchs et al.
1994,
Ellegren
1995),
this value of 0 translates to approximately
individuals. Upper Gooch Valley Pond had the largest Ne with
1765
individuals.
Pikes Point also had a large Ne of approximately 1560, while the smallest Ne was
estimated for Camp Stanford
(765).
The average 4Nem between populations was 3.12,
which, taking recipient Ne'S into account, converts to a migration rate m of 0.0007
migrants per generation. Upper Gooch Valley Pond was also the largest source of
migrants, with a combined 4Nem to all populations of 20.7 and an m of 0.005. In other
words, 0.5% of individuals in other populations originated from Upper Gooch Valley
Pond. Relative to its Ne, the Camp Stanford population is the smallest source of
81
Table 3.7. Pairwise Fi values for B.
boreas populations below the
diagonal. Associated P-values above the diagonal, obtained after
3000 permutations. Bold values are significant at the 0.003 3 level
after sequential Bonferroni correction.
BLW
BLW
UGV
MOS
P1K
RON
STF
0.06
0.007
-0.003
0.028
0.039
0.013
UGV
0.153
0.018
0.054
0.037
0.018
MOS
0.571
0.013
0.033
0.022
0.006
P1K
0.002
0.000
0.001
0.002
0.004
RON
0.002
0.000
0.014
0.297
STF
0.159
0.037
0.262
0.317
0.022
0.024
1
S
0.05
0.04
0.40
S
0.60
0.80
1.00
1.20
1.40
Ln (geographical distance) (km)
Fig. 3.2. Genetic distance
for B. boreas.
(FST/(l -FsT))
as a function of log geographic distance
1.60
migrants (m = 0.002). Blue Water receives migrants at the highest rate (0.005), most
of which come from Upper Gooch Valley Pond. Both Blue Water and Camp Stanford
receive more migrants than they contribute, while Upper Gooch Valley pond, Pikes
Table 3.8. Estimates of migration rate (M) and effective population
size () for B. boreas. Gene flow occurs in the direction from
populations on top of matrix to populations on side of matrix.
Effective population size is shown on the diagonal (in bold). U95C1
shows upper 95% confidence limit, and L95C1 shows lower 95%
confidence limit.
BLW
BLW
UGV
MOS
P1K
RON
STF
0.395
7.336
6.631
8.102
5.004
4.415
5.634
3.515
3.030
4.053
2.646
2.221
3.120
3.008
2.559
3.520
0.706
0.665
0.750
4.452
4.033
4.897
3.750
3.356
4.164
4.269
4.716
2.281
1.983
2.605
4.588
4.147
5.054
0.477
0.445
0.513
2.753
2.417
3.117
4.355
3.939
4.798
2.032
1.749
2.343
1.523
3.212
2.879
3.570
2.815
2.496
3.158
0.623
0.588
0.661
3.791
3.427
4.183
0.722
0.570
0.898
1.699
1.463
1.961
3.379
3.039
3.745
2.896
2.581
3.234
4.249
3.854
4.665
0.475
0.445
0.507
1.740
1.500
2.002
2.837
2.203
1.868
2.578
2.495
2.131
1.739
1.444
2.974
2.073
3.826
3.385
4.307
0.306
0.277
0.339
U95C1 0.356
L95C1 0.440
UGV
2.695
U95C1 2.361
L95C1 3.054
MOS
U95C1
1.907
1.628
L95C1
2.210
1.290
P1K
U95C1
L95C1
RON
U95C1
L95C1
STF
1.081
U95C1 2.456
L95C1 3.257
3.851
Point and Roney Corral all appear to act as sources of migrants for the region. The
rates of migration into and out of Mosquito Flat are approximately equal.
Population structure
Results of tests of population structure for each species are shown in Table 3.9.
Global estimates of (DST (number of groups = 1) for T. elegans and T sirtalis are
FST
values as reported in Chapter 2. The partitioning of T. elegans into two, three, four
and five groups first involved the separation of Colman Lake and Deans Meadow (P
0.011), followed by Little Cleghorn Reservoir (P = 0.003), McCoy Flat Reservoir (P =
0.004) and Blue Water (P = 0.015; Fig. 3.3). The maximum number of significantly
differentiated T. sirtalis populations was six, with Colman Lake, Deans Meadow and
Mahogany Lake clustering first (P = 0.006). This smaller group then split into two
groups, consisting of Deans and Mahogany separate from Colman (P = 0.0 19). Next,
Ashurst split from the other populations to make four groups (P
0.007), followed by
Summit (P = 0.018) and then Bullard and Gordon (P = 0.013; Fig. 3.4). Further
structuring of B. boreas populations into population clusters was not statistically
significant, but separation of northern populations (Blue Water, Man Pond, Mosquito
Flat, Camp Stanford) from southern populations (Pikes Point, Roney Corral) was only
slightly non-significant (P = 0.066; Fig. 3.5). The isolation-by-distance scatterplot for
B. boreas (Fig. 3.2) shows two clusters of points at smaller and larger geographic
distances. These clusters correspond with distances within and between the groups of
populations identified by SAMOVA. Points associated with smaller distances
correspond to distances between populations within groups, and points
associated with larger distances correspond to among-group distances.
Table 3.9. Genetic structure analyses for B. boreas, T. elegans and T. sirtalis. Best results for each pre-defined number
of groups shown. Only number of groups which gave significant or almost significant 'DCT are shown.
Number of
groups
B. boreas
Structure tested
Total
Var.
I
(BLW, MAR, MOS, P1K, RON, STF)
58.43
ST
2
(BLW, MAR, MOS, STF) (P1K, RON)
58.96
ST
SE
P-value
= 0.024
<0.00001
<0.00001
= 0.035
CT°°28
<0.00001
<0.00001
0.0019
<0.00001
<0.00001
0.066
(1)-statistic
1)sc=O.007
T. elegans
1
(ANT, ANS, ASH, BLW, BUL, CLG, COL, DNS, GAL, JCK,
LTC, MAlI, MCY, NML, PAP, P1K, RKY, RON, STF, SUM)
24.49
1)ST =
0.025
<0.00001
<0.00001
2
(ANT, ANS, ASH, BLW, BUL, CLG, GAL, JCK, LTC, MAH,
MCY, NML, PAP, P1K, RKY, RON, STF, SUM) (COL, DNS)
25.51
ST =
0.040
<0.00001
<0.00001
=
1)sc 0.023
<0.0000 1
1)cr°.°l8
<0.00001
0.0008
0.039
<0.0000 1
<0.0000 1
= 0.022
<0.0000 1
<0.0000 1
0.0004
0.003
<0.0000 1
<0.00001
0.0005
<0.00001
<0.00001
0.004
<0.0000 1
<0.0000 1
<0.0000 1
<0.0000 1
0.0009
0.015
3
(ANT, ANS, ASH, BLW, BUL, CLG, GAL, JCK, MAlI, MCY,
NML, PAP, P1K, RKY, RON, STF, SUM) (COL, DNS) (LTC)
25.47
ST =
I
0.0 17
=
0.037
ST
=
'1)sc 0.022
CT =
4
(ANT, ANS, ASH, BLW, BUL, CLG, GAL, JCK, MAll, NML,
PAP, P1K, RKY, RON, STF, SUM) (COL, DNS) (LTC) (MCY)
25.43
CT°.°i5
5
(ANT, ANS, ASH, BUL, CLG, GAL, JCK, MAH, NML, PAP,
P1K, RKY, RON, STF, SUM) (COL, DNS) (LTC) (MCY) (BLW)
25.33
0.03 3
=
1)sc 0.022
ST =
CT°.°11
0.011
00
Table 3.9. (continue
Number
of groups
Structure tested
Total
Var.
F-statistic
SE
P-value
T. sirtalis
I
(ANT, ANS, ASH, BUL, COL, DNS, FEA, GOR, MAH,
NML, RON, STF, SUM)
21.12
'ST = 0.035
<0.00001
<0.00001
2
(ANT, ANS, ASH, BUL, PEA, GOR,
NML, RON, STF, SUM) (COL, DNS, MAll)
21.44
3
(ANT, ANS, ASH, BUL, DNS, FEA, GOR,
NML, RON, STF, SUM) (DNS, MAH) (COL)
21.44
Dsi = 0.054
1sc 0.025
= 0.029
DCT
DST = 0.049
= 0.027
Isc
<0.00001
<0.00001
0.0007
<0.00001
<0.00001
0.0011
<0.00001
<0.00001
0.0006
<0.00001
<0.00001
0.0011
<0.00001
<0.00001
0.0009
<0.00001
<0.00001
0.006
<0.00001
<0.00001
0.019
<0.00001
<0.00001
0.007
<0.00001
<0.00001
0.018
<0.00001
<0.00001
0.013
cb1=O.O23
4 (ANT, ANS, ASH, BUL, DNS, FEA, GOR,
NML, RON, STF, SUM) (DNS, MAH) (COL) (ASH)
21.40
5
(ANT, ANS, ASH, BUL, DNS, FEA, GOR,
NML, RON, STF, SUM) (DNS, MAH) (COL) (ASH) (SUM)
21.30
(ANT, ANS, ASH, FEA, NML, RON, STF, SUM)
(DNS, MAH) (COL) (ASH) (SUM) (BUL, GOR)
21.23
6
= 0.048
Dsc = 0.023
Dci = 0.026
= 0.044
'DST
DST
bsc 0.024
CTO.O2O
1ST = 0.041
=
Dsc 0.020
1CT°.°22
Fig. 3.3. Map showing clustering pattern for T elegans populations. Thickness of
borders indicates to grouping sequence, with thicker lines corresponding to groups
defined earlier in the analysis.
87
:JCK5TF
CLG
0
SUM
ASH
RKY
n.
i
Eagle Lake
PVM
.
AMP
GAL \
,DNS
PIK
MAH
PAP .
.
S
RON
NMI
5km
OL
a
V
0
BULOO
I
GOR
I
SUM
I1M
I
Eagle Lake
PVM
AMP
DNS
°FEA
NML
S
'RON
.
COL
5
km
Fig. 3.4. Map showing clustering pattern for T. sirtalis populations.
Conventions as in Figure 3.3.
I
\
Eagle Lake
IK
5h
Fig. 3.5. Map showing clustering pattern for B.
Conventions as in Figure 3.3.
boreas
populations.
)
Ecological correlates
Scatterp lots for variables used in the regression analyses of effective population size,
migration rate and genetic distance are shown in Figures 3.6-8, and the results of the
stepwise regression are shown in Table 3.10. SNAKES was never used in the same
model as ELEG and SIRT, and SNAKEDIFF was never used in the same analysis as
ELEGDIFF and STRTDIFF. A total of nine analyses were constructed with tests of
ecological effects on three dependent variables for all three species. Analyses with Ne
as a response variable identified ecological factors that were influential after taking
census size into account. No ecological variables were significant determinants Of Ne
in T. elegans after census size was accounted for
(F1,20
= 20.73, P
0.0002). Census
size accounted for only 52% of the variance in Ne. For T. sirtalis, Ne was influenced
by elevation and nearest-neighbor elevation (F3,20
24.78, P < 0.000 1), such that sites
with higher elevations had higher effective sizes, and sites whose nearest neighbor had
higher elevations and lower effective sizes. For B. boreas, Ne was much larger for
populations whose nearest neighboring site was a deeper water body (F2,20 =31.92, P
<0.0001) which, combined with census size, explained 78% of the variation in Ne.
Stepwise regression of distance matrices implemented in PERMUTE! did not
give coefficients for selected matrix variables. However, an analysis with the same
variables performed in SAS generally gave almost identical results, so coefficients and
their standard errors given in Table 3.9 are from the SAS analyses. For T. elegans,
91
'II
:::E
-
ri
t
r,I
U__
-
U
-
_5i IIdJ1U -1='
I
JII!IrUI
_
I11riy1U
.
II
i
UUr1
.iq
______
Fig. 3.6. Scatterplots for pairs of variables used in stepwise regression of effective
population size.
migration rate from one population to another was most influenced by differences in
T elegans census size, T sirtalis census size and perimeter, after taking geographic
distance into account. These four variables explained only 30% of the variation in
migration rate. Migration among T elegans populations is more likely to occur
toward smaller sites with fewer T elegans and more T sirtalis. Previous research has
92
..
DJST
PIIfVJLI
k I
P4i
.
0
j
TYPE
DIFF
-. ._.
-
!
-- . -
-
.
- -
..c:
:
**e-*
..:..
I
.
I
s**
III
-
i
.:...:.
.-.z.I..
.
i
.
%
I
.
.
:
:
'.
!91'
.
.
1
.
Sq
1
......
.
!
SNAKE
$1
I
I
I
p.
.
I'.
Fig. 3.7. Scatterplots for pairs of variables used in stepwise regression of migration
rate.
93
DIST
'
I
3
I
PERIM.
::?
.
DIFF
.
i..
:
.
..
..:
S
.
.
I
-
TYPE
DIFF
ELEV
::4;':
'.
t
:
BUFO
DIFF
i
.. .
.
j.L
Z,ts.
._...
.
!.
.
z
t4- ip1
E±I
_'
!
'
DIFF
I
I
I
:iI.
_
.
':
SNAKE
DIFF
,i:.,'P4t:
Fig. 3.8. Scatterplots for pairs oi variables used in stepwise regression of genetic
distance.
Table 3.10. Resuts of nine multiple regression analyses testing for effects of habitat on effective population size (Ne),
migration rate (m) and genetic distance (FST/J-FST) for each species, showing parameter estimates (standard error), Pvalues and R2 contribution for each explanatory variable. P-values and R2 for the whole model are shown in the
columns on the far right. Stepwise regression of migration and genetic distance matrices does not give estimates for
the intercept.
Y
Species
Intercept
Xl
X2
X3
P-value
R2
T
elegans
ELEG
0.0002 0.5218
Ne
49.059 (74.729)
87.251 (19.164)
0.5 194
0.0002; 0.5218
T. sirtalis
-488.28 (325.38)
0.1518
SIRT
19.915 (2.4304)
<0.0001; 0.6709
ELEV
0.62 12 (0.1720)
0.0022; 0.1091
-431.26 (151.24)
0.0106
BUFO
148.22 (24.101)
<0.0001; 0.5426
NN-TYPE
394.68 (89.530)
0.0003; 0.2375
B. boreas
NN-ELEV
-0.2964 (0.1687)
0.0969; 0.0338
<0.0001
0.8139
<0.000 1
0.7800
Table 3.10.
(continued)
Y Species
m T elegans
Xl
ELEGDJFF
2.6094e-4 (2.160e-5)
0.014; 0.2421
T. sirtalis
SIRTDIFF
6.644e-5 (5.88e-6)
0.001; 0.4487
B. boreas
SNAKEDIFF
-2.462e-5 (7.1 7e-6)
0.003; 0.145
DIST
-8.51e-6 (3.23e-6)
0.011; 0.0969
X2
SIRTDIFF
-2.398e-5 (2.33 le-5)
0.001; 0.03
PERIMDIFF
2.698e-4 (5.146e-5)
0.001; 0.1015
X3
PERIMDIFF
2.879e-5 (9.39e-6)
0.003; 0.0176
ELEVDIFF
3.Ole-6 (6.387e-7)
0.001; 0.1077
X4
DIST
-7.83e-6 (4.73e-6)
0.009; 0.014
TYPEDIIFF
3.425e-4 (8.972e-5)
0.001; 0.2067
P-value
R2
0.001
0.304
0.001
0.449
0.001
0.658
Table 3.10. (continued)
Y
Species
FST/(1-FsT)
T.
elegans
Xl
DIST
4.809e-5 (1 .324e-4)
0.001; 0.0579
T sirtalis
DIST
0.0013 (2.58 le-4)
0.001; 0.1583
B. boreas
DIST
8.859e-4 (2.39 le-4)
0.003; 0.4639
X2
ELEVDJFF
4.1 08e-5 (1.1 69e-5)
0.002; 0.0555
X3
ELEGDIFF
-0.0040 (0.0013)
0.001; 0.1391
PERIMDIFF
-0.0068 (0.0024)
0.009; 0.0552
P-value
R2
0.001
0.1134
0.001
0.3526
0.007
0.4639
97
identified Papoose Meadows as the major source of migrants in the study area, a site
that has a very large perimeter, a large T. elegans population and very few T. sirtalis.
Migration by T. sirtalis was more frequent into populations with smaller census sizes.
When geographic distance was included in the model, 46% of the variation in
migration rate was explained. Rates of B. boreas migration were found to be
influenced by differences in snake census size, site perimeter, elevation and site type,
which together with geographic distance explain 66% of the variation.
Genetic distance between T. elegans populations was greater for populations
with larger differences in elevation, but the model chosen by the stepwise multiple
regression procedure explained only 11% of the variation. Genetic distance of T
sirtalis populations was greater at sites with larger differences in T. elegans census
size and more similar site perimeter sizes, after accounting for geographic distance, in
a model that explained 35% of the variation. Bufo boreas genetic distance was related
to geographic distance alone, which explained 46% of the variation. No other
variables were selected by the stepwise multiple regression procedure.
Relative to their representation, biotic interactions and nearest neighbor
characteristics were less important than habitat descriptors such as perimeter and
elevation. Regression models with effective population size as a response variable
always included census size. Interspecific interactions did not influence effective
population size for any species, and
Ne
was explained by one to three variables that
accounted for 71% of the variation, on average. These variables largely described
habitat of the site itself or its nearest neighbor. Overall, migration rate was determined
by one to five variables of habitat features and both intra- and interspecific
interactions. In particular, migration occurred from sites with higher densities to sites
with lower densities of both the focal and interacting species, and the selected models
explained, on average, 47% of the variation in migration rate. Genetic distance was
explained by one to three variables, which also accounted for the smallest percentage
of variation in genetic distance (31% on average).
DISCUSSION
Spatial genetic structure
Overall, spatial structuring of garter snake populations was characterized by one or
two genetically distinct populations. Identification of a group that included Colman
Lake and Deans Meadow is characteristic of both species. A series of block faults lies
between these populations and Nameless and Papoose Meadows to the west. The
resulting 300 m escarpment appears to be a significant barrier to dispersal for both
species. In the northeast portion of the study area, the T elegans population at Little
Cleghorn Reservoir is genetically differentiated from most other populations, a
surprising result considering that it is only 1.4 km away from Cleghorn Reservoir.
Including Cleghorn Reservoir in a group with Little Cleghom Reservoir within a
three-group structure lowers the (DCT to 0.0075 (P = 0.057) from the
DCT
of 0.017 (P =
0.003; Table 3.8). The pairwise estimate of DsT for Cleghom Reservoir and Little
I1*]
Cleghorn is 0.018 (P = 0.0741), not significant, but nevertheless large considering
their close proximity. The general results of the spatial clustering analysis were
similar for both species, consistent with previous research that found correlated
genetic structures for T. elegans and T. sirtalis in the study area (Chapter 2). Tn
contrast, clustering of B. boreas populations, though not statistically significant, is
different from that of the garter snakes in identifying a disjunction between the
southern and northern populations.
Previous research investigating patterns of isolation by distance for T. elegans
and T sirtalis found evidence for lack of migration-drift equilibrium (Chapter 2).
Specifically, the pattern of the regression of genetic distance on geographic distance
showed restricted gene flow between sites separated by moderate distances (Chapter 2,
Fig. 2.2). When the data were re-examined after removing Colman Lake and Deans
Meadow, the regression became much more linear. Restricted gene flow with these
two sites seems to explain much of migration-drift disequilibrium observed for these
two species. The same study also found a correlation between pairwise FST'S ofT
elegans
and T
sirtalis.
This result does not appear to be due entirely to the Colman
Lake and Deans Meadow sites, although their exclusion from the analysis produces a
correlation that is slightly nonsignificant (P = 0.057).
Ecological correlates ofpopulation genetic parameters
Stepwise multiple regression analysis identified habitat, biotic and nearest neighbor as
features that were statistically important in explaining effective population size,
migration rate and genetic distance in T. elegans, T sirtalis and B. boreas. Models for
T elegans explained much less of the variation in population genetic parameters than
for the other species, suggesting that different factors are important for genetic
differentiation in that species than for T sirtalis and B. boreas. One possible
explanation is that evolutionary forces such as selection play a larger role in driving
divergence of T. elegans populations. This hypothesis is supported by previous work
that has identified genetically-based life-history and color pattern differences between
T. elegans populations on the shore of Eagle Lake (e.g., Gallatin Shoreline, Pikes
Point) and those at meadows (e.g., Papoose Meadows, Mahogany Lake).
For analyses of T elegans and T. sirtalis, variables selected to explain
migration rates were strongly influenced by the primary source population. Migration
occurred most often into populations with lower census sizes of the focal species,
which is an obvious result considering that the largest source of migrants for both
species also has the largest census (and effective) population sizes. Nevertheless, the
results may indicate that garter snakes were more likely to migrate toward sites with
lower snake density, perhaps because competition is less intense. Bufo boreas
migrants also tend to migrate away from sites with higher snake density, in this case
probably because predation is less intense. Overall, there were no trends in ecological
variable selection that were common to any species pair, providing little support for
the notion that a common landscape exerts similar ecological effects on genetic
responses.
101
Models with FST as the response variable differed markedly from those with m
as the response.
FST
for all species was explained primarily by geographic distance,
while m generally was associated with more variables and a higher R2. Analyses of Ne
and m also selected very different explanatory models, indicating that analyses using
Fi alone may be losing important information gained by obtaining independent
estimates Ne and m.
Some debate has recently arisen regarding the amount of variation that can be
explained by factors of interest in studies of ecological and evolutionary processes
(Møller and Jennions 2002, Peek et al. 2003). The R2 values obtained in this study are
consistent with the findings of Peek et al. (2003), who maintain that the amount of
explained variance can be relatively high, up to 50%. The regression models from this
study explained between 11% and 81% of the variance in the population genetic
parameters, with an average of 50%. In contrast, a metaanalysis by Møller and
Jennions (2002) concluded that only 2.5% 4.5% of variance is in ecological and
evolutionary studies is explained by factors of interest. Although higher R2 values can
result from the inclusion of more explanatory variables, we saw no relationship
between number of variables selected by the stepwise regression procedure and the
amount of variation they explained (r = 0.128, P = 0.743).
B. boreas population genetics
We found low but significant genetic structuring of B. boreas populations, with an FST
of 0.024. This value was similar to those previously reported for T elegans (0.025)
102
and T. sirtalis (0.03 5; Chapter 2), implying a similar overall level of genetic
differentiation. Populations of B. boreas, however, had effective sizes that were
substantially larger than those for the garter snakes (average Ne = 1240 for B. boreas,
375 for T elegans, 325 for T. sirtalis), while bi-directional migration rates were
approximately half those ofT. elegans and T. sirtalis (average m = 0.0007 for B.
boreas, 0.0012 for T. elegans and 0.0017 for T sirtalis). Thus, comparable FST values
among all species masked differences in Ne and m.
Genetic structuring observed for B. boreas within the study area is consistent
with the repeated observation for many anurans of fidelity to a breeding site and low
juvenile dispersal, despite a high capacity for vagility in both juveniles and adults.
Subadult leopard frogs (Rana pipiens) can travel up to five km (Dole 1971), and
juvenile natterjack toads (Bufo calamita) have been found to disperse from their natal
ponds up to two km within a few weeks of metamorphosis (Sinsch 1997). A high
capacity for mobility has also been documented in other toad species (B. woodhousii
fowleri, John-Alder and Morin 1990; B. bufo, Goater et al. 1993). In our study
system, adult toads were frequently seen crossing roads at night, far from a water
source. Despite their capacity for long-distance movement, many amphibians have
breeding site philopatry that leads to genetic structuring around ponds. Berven and
Grudzien (1990) found 100% fidelity in adult wood frogs (Rana sylvatica) to a
breeding pond, although 18% of juveniles dispersed away from their natal pond.
Berven (1982) found comparable levels of juvenile dispersal between ponds separated
by 50 m, and Jameson (1956) documented homing behavior in the Pacific tree frog
103
(Hyla regilla) from a distance of 1000 yards. These studies suggest that site fidelity
may be a critical factor driving fine-scale genetic differentiation in pond-breeding
anurans.
Although measures of FST observed in this study are considerably smaller than
those obtained from studies on comparable systems, our results support a general
observation of population differentiation over short geographic distances in anurans
(e.g., Hitchings and Beebee 1997, Brede and Beebee 2004). Studies using
microsatellite markers obtained estimates of population differentiation that were
comparable to those from this study or higher. For example, Rowe et al. (2000)
examined three groups of B. calamita populations, each with among-site distances of
less than 30 km. Overall FST from microsatellite data ranged from 0.060 to 0.224, all
with significant P-values. Monsen and Blouin (2003) surveyed microsatellite loci
from populations of the montane frog (Rana cascadae) in the Pacific Northwest and
found FST across distances less than 100 km to range from 0.04 to 0.28. Furthermore,
Monsen and Blouin (2004) found a sharp decrease in gene flow over distances greater
than 10km.
Bufo boreas effective population sizes were high relative to effective sizes for
both garter snakes but only moderately large when compared with other anurans. In
our study system, B. boreas Ne averaged 1240. Bufo marinus populations in Hawaii
(N = 390) and Australia (N = 346) had estimates lower by a factor of three, based on
means and variances of FST (Easteal 1985). A recent study in northern leopard frogs
(Rana pipiens) found slightly smaller estimates that are roughly comparable to ours
104
(average
Ne
= 851) using a moment estimator (Waples 1989), which was deemed most
accurate of the three temporal methods used (Hoffman et al. 2004). Estimation of
effective number of breeders (Merrell 1968, Berven and Grudzien 1990, Seppa and
Laurila 1999) tended to be an order of magnitude lower, averaging less than 100, in
other anurans. On the other hand, dirt frogs (Eleutherodactylus spp.) in Central
America had very high effective sizes (31 000 and 10 000), estimated using sequence
data at a nuclear locus (Crawford 2003). These estimates were also based on genetic
diversity and, like those obtained for R. pipiens, may actually represent effective sizes
at a wider regional scale, perhaps over the entire species.
Levels of within-population genetic variation expressed as expected
heterozygosity were higher for B. boreas (average H = 0.77) relative to both T.
elegans (0.54) and 7'. sirtalis (0.59). Because heterozygosity can be lost due to
stochastic fluctuations in population size or extinction-recolonization dynamics
(Newman and Squire 2001), the relatively higher heterozygosity for B. boreas
populations suggests that they may be more stable regionally than the garter snake
populations. This hypothesis is supported by the much higher effective sizes and
lower migration rates found for B. boreas, which indicate that populations are
relatively closed with very little gene flow. Conversely, previous research has found
that migration among the garter snake populations is dominated by a single source
population whose average output of migrants is approximately five times that of the
other populations (Chapter 2), suggesting extinction-recolonization dynamics within a
source-sink metapopulation. In contrast, the largest source of B. boreas migrants is
105
Upper Gooch Valley Pond, with only slightly larger-than-average output. Other
studies have found stable genetic structure in anurans (Hoffman et al. 2004, Rowe and
Beebee 2004), though population sizes can fluctuate widely (Berven and Grudzien
1990). These results are consistent with the general observation that source-sink
dynamics in anurans is rare (Breven and Grudzien 1990).
General conclusions
In this study, we identified ecological factors related to population genetic
differentiation for three vertebrate species that coexist on the same landscape.
Overall, B. boreas population dynamics appear to be more stable than garter snake
dynamics, with larger effective sizes, lower migration and migration-drift equilibrium.
An escarpment was identified as a significant barrier to dispersal for both garter
snakes, but toad populations appeared to cluster based on isolation by distance alone.
We found no common trends in ecological correlates of population genetic parameters
among the three study species, but certain response variables produced similar
explanatory models. Effective population size was largely explained by census size,
migration rate was strongly associated with competitive and predator-prey
interactions, and genetic distance was most correlated with geographic distance. The
similarities and differences we observed in the population structure suggest that each
species responded to the shared landscape differently.
106
REFERENCES
Arnaud, J.-F., L. Madec, A. Guiller and A. Bellido. 2001. Spatial analysis of
allozyme and microsatellite DNA polymorphisms in the land snail Helix
aspersa (Gastropoda: Helicidae). Molecular Ecology 10:1563-1576.
Banchs, I., A. Bosh, J. Guimera, C. Lazaro, A. Puig and X. Estivill. 1994. New
alleles at microsatellite loci in CEPH families mainly arise from somatic
mutations in the lymphoblastoid cell line. Human Mutation 3:365-372.
Beerli, P. and J. Felsenstein. 2001. Maximum likelihood estimation of a migration
matrix and effective population sizes in n subpopulations by using a coalescent
approach. Proceedings of the National Academy of Sciences 98:4563-4568.
Berven, K.A. 1982. The genetic basis of altitudinal variation in the wood frog Rana
sylvatica. I. An experimental study of life history traits. Evolution 36:962983.
Berven, K.A. and T.A. Grudzien. 1990. Dispersal in the wood frog (Rana sylvatica):
implications for genetic population structure. Evolution 44:2047-2056.
Brede, E.G. and T.J.C. Beebee. 2004. Contrasting population structures in two
sympatric anurans: implications for species conservation. Heredity 92:110117.
Castric, V., F. Bonney and L. Bematchez. 2001. Landscape structure and hierarchical
genetic diversity in the brook charr, Salvelinusfontinalis. Evolution 55:10161028.
Coulon, A., J.F. Cosson, J.M. Angibault, B. Cargnelutti, M. Galan, N. Morellet, E.
Petit, S. Aulagnier and A.J.M. Hewison. 2004. Landscape connectivity
influences gene flow in a roe deer population inhabiting a fragmented
landscape: an individual-based approach. Molecular Ecology 13:2841-2850.
Crawford, A.J. 2003. Huge populations and old species of Costa Rican and
Panamanian dirt frogs inferred from mitochondrial and nuclear gene
sequences. Molecular Ecology 12:2525-2540.
Dallas, J.F. 1992. Estimation of microsatellite mutation rates in recombinant inbred
strains of mouse. Mammalian Genome 5:32-38.
Dole, J.W. 1971. Dispersal of recently metamorphosed leopard frogs, Ranapipiens.
Copeia 1971:221-228.
Dupanloup, I., S. Schneider, A. Langaney and L. Excoffier. 2002. A simulated
annealing approach to define the genetic structure of populations. Molecular
Ecology 11:2571-2581.
Edwards, A., H.A. Hammond, L. Jin, C.T. Caskey and R. Chakraborty. 1992.
Genetic variation at five trimeric and tetrameric tandem repeat loci in four
human population groups. Genomics 12:241-253.
Easteal, S. 1985. The ecological genetics of introduced populations of the giant toad
Bufo marinus. H. Effective population size. Genetics 110:107-122.
Ellegren, H. 1995. Mutation rates at porcine microsatellite loci. Mammalian Genome
6:376-377.
107
Eriksson, J., G. Hohmann, C. Boesch and L. Vigilant. 2004. Rivers influence the
population genetic structure of bonobos (Pan paniscus). Molecular Ecology
13:3425-3435.
Excoffier, L., P.E. Smouse and J.M. Quattro. 1992. Analysis of molecular variance
inferred from metric distances among DNA haplotypes: application to human
mitochondrial DNA restriction data. Genetics 131:479-491.
Gamier, S., P. Alibert, P. Audiot, B. Prieur and J.-Y. Rasplus. 2004. Isolation by
distance and sharp discontinuities in gene frequencies: implications for the
phylogeography of an alpine insect species, Cara bus solieri. Molecular
Ecology 13:1883-1897.
Goater, C.P., R.D. Semlitsch and M.V. Bemasconi. 1993. Effects of body size and
parasite infection on the locomotory performance ofjuvenile toads, Bufo bufo.
Oikos 66:129-136.
Guo, S.W. and E.A. Thompson. 1992. Performing the exact test for Hardy-Weinberg
proportions for multiple alleles. Biometrics 48:36 1-372.
Hitchings, S.P. and T.J.C. Beebee. 1997. Genetic substructuring as a result of barriers
to gene flow in urban Rana temporaria (common frog) populations:
implications for biodiversity conservation. Heredity 79:117-127.
Hoffman, E.A., F.W. Schueler and M.S. Blouin. 2004. Effective population sizes and
temporal stability of genetic structure in Rana pzpiens, the northern leopard
frog. Evolution 58:2536-2545.
Hutchison, D.W. and A.R. Templeton. 1999. Correlation of pairwise genetic and
geographic distance measures: inferring the relative influences of gene flow
and drift on the distribution of genetic variability. Evolution 53:1898-1914.
Jameson, D.L. 1956. Growth, dispersal and survival of the Pacific tree frog. Copeia
1956:25-29.
John-Alder, H.B. and P.J. Morin. 1990. Effects of larval density on jumping ability
and stamina in the newly metamorphosed Bufo woodhousiifowleri. Copeia
1990:856-860.
Keyghobadi, N., J. Roland and C. Strobeck. 1999. Influence of landscape on the
population genetic structure of the alpine butterfly Parnassius smintheus
(Papilionidae). Molecular Ecology 8:1481-1495.
Legendre, P., F.-J. Lapointe and P. Casgrain. 1994. Modeling brain evolution from
behavior: a permutational regression approach. Evolution 48:1487-1499.
Manly, B.F. 1997. Randomization and Monte Carlo methods in biology. 2' ed.
Chapman and Hall, New York.
Mantel, N. 1967. The detection of disease clustering and generalized regression
approach. Cancer Research 27:209-220.
Mantel, N. and R.S. Valand. 1970. A technique of nonparametric multivariate
analysis. Biometrics 26:547-558.
Merrell, D.J. 1968. A comparison of the estimated size and the "effective size" of
breeding populations of the leopard frog, Ranapipiens. Evolution 22:274-283.
Møller, A.P. and M.D. Jennions. 2002. How much variance can be explained by
ecologists and evolutionary biologists? Oecologia 132:492-500.
Monsen, K.J. and M.S. Blouin. 2003. Genetic structure in a montane ranid frog:
restricted gene flow and nuclear-mitochondrial discordance. Molecular
Ecology 12:3275-3286.
Monsen, K.J. and M.S. Blouin. 2004. Extreme isolation by distance in a montane
frog Rana cascadae. Conservation Genetics 5:827-835.
Nevo, E., B. Baum, A. Beiles and D.A. Johnson. 1998. Ecological correlates of
RAPD DNA diversity of wild barley, Hordeum spontaneum, in the Fertile
Crescent. Genetic Resources and Crop Evolution 45:151-159.
Newman, R.A. and T. Squire. 2001. Microsatellite variation and fine-scale
population structure in the wood frog (Rana sylvatica). Molecular Ecology
10:1087-1100.
Peek, M.S., A.J. Leffler, S.D. Flint and R.J. Ryel. 2003. How much variance is
explained by ecologists? Additional perspectives. Oecologia 137:161-170.
Raymond, M. and F. Rousset. 1995. GENEPOP Version 1.2: population genetics
software for exact tests and ecumenicism. Journal of Heredity 86:248-249.
Reh, W. and A. Seitz. The influence of land use on the genetic structure of
populations of the common frog Rana temporaria. Biological Conservation
54:239-249.
Rice, W.R. 1989. Analysing tables of statistical tests. Evolution 43 :223-225.
Roach, J.L., P. Stapp, B. Van Home and M.F. Antolin. 2001. Genetic structure of a
metapopulation of black-tailed prairie dogs. Journal of Mammalogy 82:946959.
Rousset, F. 1997. Genetic differentiation and estimation of gene flow from Fstatistics under isolation by distance. Genetics 145:1219-1228.
Rowe, G., T.J.C. Beebee and T. Burke. 2000. A microsatellite analysis of natterjack
toad, Bufo calamita metapopulations. Oikos 88:641-651.
SAS Institute. 2002. SAS/STAT software user's guide. Rd. 9.1. SAS Institute, Inc.,
Cary, NC.
Schneider, S., D. Roessli and L. Excoffier. 2000. Arlequin ver 2.000: a software for
population genetics data analysis. Genetics and Biometry Laboratory,
University of Geneva, Switzerland.
Schweiger, 0., M. Frenzel and W. Durka. 2004. Spatial genetic structure in a
metapopulation of the land snail Cepaea nemoralis (Gastropoda: Helicidae).
Molecular Ecology 13:3645-3655.
Scribner, K.T., J.W. Amtzen, N. Cruddace, R.S. Oldham and T. Burke. 2001.
Environmental correlates of toad abundance and population genetic diversity.
Biological Conservation 98:201-210.
Scribner, K.T. and R.K. Chesser. 1993. Environmental and demographic correlates of
spatial and seasonal genetic structure in the eastern cottontail (Sylvilagus
floridanus). Journal of Mammalogy 74:1026-1044.
Seppa, P. and A. Laurila. 1999. Genetic structure of island populations of the anurans
Rana temporaria and Bufo bufo. Heredity 82:309-317.
Sinsch, U. 1997. Postmetamorphic dispersal and recruitment of first breeders in a
Bufo calamita metapopulation. Oecologia 112:42-47.
109
Simandle, E. 2005. Phylogeography and metapopulation structure of the Bufo boreas
group in the Great Basin. Ph.D. dissertation; Ecology, Evolution and
Conservation Biology Program; University of Nevada, Reno.
Slatkin, M. and L. Excoffier. 1996. Testing for linkage disequilibrium in genotypic
data using the EM algorithm. Heredity 76:377-3 83.
Sokal, R. and F.J. Rohlf. 1995. Biometry, 31( ed. New York, New York: W.H.
Freeman.
Van Rossum, F., I. Bonnin, S. Fénart, M. Pauwels, D. Petit and P. Saumitou-Laprade.
2004. Spatial genetic structure within a metallicolous population of
Arabidopsis halleri, a clonal, self-incompatible and heavy-metal-tolerant
species. Molecular Ecology 13:2959-2967.
Waples, R. S. 1989. A generalized approach for estimating effective population size
from temporal changes in allele frequency. Genetics 121:379-391.
Weber, J.L. and C. Wong. 1993. Mutation of human short tandem repeats. Human
Molecular Genetics 2:1123-1128.
Whiteley, A.R., P. Spruell and F.W. Allendorf. 2004. Ecological and life history
characteristics predict population genetic divergence of two salmonids in the
same landscape. Molecular Ecology 13:3675-3688.
110
ADAPTIVE DIVERGENCE BETWEEN ECOTYPES OF THE TERRESTRIAL
GARTER SNAKE, THAMNOPHIS ELEGANS
CHAPTER 4
Mollie K. Manier and Stevan J. Arnold
This thesis chapter has been prepared for submission to the journal Evolution.
111
ABSTRACT
Genetic differentiation was estimated for two types of quantitative traits and compared
with estimates obtained using neutral microsatellite markers for six populations of the
common garter snake, Thamnophis elegans. The study populations are part of a larger
metapopulation with source-sink dynamics and exhibit marked differences in life-
history variation and color pattern. These ecotypes have been shown to be genetically
based and are thought to have evolved in response to differences in food availability
and avian predation. The goal of this study was to compare estimates of QST for color
and scalation traits with FST for neutral loci in order to test for selection on the
phenotypic traits. We found phenotypic differences between ecotypes in both
scalation and coloration, and QST values for most traits were substantially higher than
FST.
We found no correlation between pairwise FST and QST estimates. Patterns of
Qsi among populations were similar to those between ecotypes. QST values for color
variables were higher between ecotypes than among populations only for females. Of
the scale counts, number of ventral scales on the body and tail and number of
infrabials were most differentiated. Background coloration showed more amongpopulation divergence than any other phenotypic trait. Scale counts were
proportionally more differentiated than stripe color variables. Stronger selection was
observed for females than males in traits associated with locomotion and crypticity,
perhaps due to the locomotory constraints imposed by relatively larger bodies and
pregnancy. Overall, we found strong genetic differentiation in phenotypic traits
112
relative to neutral expectations, indicating local adaptation in the face of moderate
levels of gene flow.
113
INTRODUCTION
Adaptive divergence is in local populations is often a function of two opposing forces:
selection and gene flow. Whereas selection can eliminate low-fitness alleles from a
population and magnify local adaptation, gene flow can homogenize population
differences and reduce local adaptation. In this way, gene flow can effectively swamp
out locally advantageous alleles, preventing a population from reaching an adaptive
optimum. Standing levels of phenotypic divergence in natural populations may be the
net product of these processes, but differentiation can also occur in the absence of
selection as a result of random genetic drift. Determining the relative influence of drift
and selection in population divergence is key to understanding local dynamics of adaptive
evolution.
Phenotypic differentiation among populations is exceedingly common in a wide
variety of taxa, but to relate this differentiation to local adaptation requires distinguishing
between the effects of selection and genetic drift. The statistical comparison of
population differentiation at quantitative traits (QsT) and neutral molecular markers
(FST) provides a test for selection acting on the traits (Lande 1992, Spitze 1993).
While a global correspondence between Qsi and FST has been a matter of recent
controversy (Merilä and Cmokrak 2001, Cmokrak and Merilä 2002, Latta and McKay
2002, McKay and Latta 2002), the comparison of genetic variation at quantitative traits with
that at neutral markers is a powerful tool for identifying the relative roles of selection and
drift in phenotypic differentiation (Lande 1992).
114
A growing volume of literature explores the relationship between neutral marker
and quantitative trait divergence. Much of the early research in this area was done in plant
and invertebrate systems (McKay and Latta 2002, Merilä and Cmokrak 2001), but a recent
influx of studies compare estimates of FST and QST in vertebrate systems, particularly in
anurans (Palo et al. 2003, Cano et al. 2004). Recent studies have also tended to use a priori
expections of the FST-QST relationship to predict modes of selection acting on quantitative
traits (Prout and Barker 1993, Spitze 1993, Bonnin et al. 1996, Yang et al. 1996, Palo et al.
2003, Cano et al. 2004). A few studies have also tested specific hypotheses of local
selection (Baker 1992, Waldmann and Andersson 1998, Gomez-Mestre and Tejedo 2004).
The present report provides additional insight into microevolutionary processes driving
population divergence in vertebrates by comparing quantitative and neutral divergence
among closely proximate garter snake populations with known life-history differences.
In this study, we investigate quantitative trait and microsatellite genetic structure
among natural populations of the terrestrial garter snake (Thamnophis elegans) in Lassen
Co., California Previous research has documented ecotypic variation in life history, growth
and survival between populations along the shoreline of Eagle Lake and those inhabiting
lakes and flooded meadows at higher elevations (Bronikowski and Arnold 1999). Theory
on the evolution of life history strategies predicts that growth rate is genetically
coupled with other life-history characters as a coevolving unit (Charlesworth 1990,
Steams 1992). Common garden experiments have shown that growth rate differences
among T. elegans ecotypes have a genetic component (Bronikowski 1998). The
nighttime temperatures at the meadow sites are five degrees colder than those at the
115
lakeshore sites. Neonates who were raised experiencing their native nighttime
temperature grew faster than those who were raised with the nighttime temperature of
the other ecotype, even if that temperature was higher than the native temperature.
This intrinsic growth rate difference is part of syndrome of differences between the
two ecotypes in life history traits. Lakeshore populations experience faster growth
rates, earlier age at first reproduction, higher fecundity, and lower adult survival than
meadow populations.
We focus on differences in coloration that appear to increase crypticity in
lakeshore and meadow environments. Lakeshore snakes tend to have dull yellow or
tan stripes on a gray background color, whereas meadow snakes have bright yellow or
orange stripes on a black background. In the lakeshore habitat, snakes forage along
the rocky shoreline of Eagle Lake for small fish and leeches (Kephart 1982, Kephart
and Arnold 1982). The snakes are exposed to predators as they commute between the
water and rock refuges 5-40m from the shoreline. The muted colors of the lakeshore
snakes tend to match the rocky substrate of the lakeshore. In contrast, meadow sites
are relatively shallow water bodies where amphibians and leeches are the principle
food source. Meadow habitats are dominated by aquatic rushes and other emergent
vegetation that carpet the shallow lake bottoms. The meadow coloration pattern
(yellow or orange stripes on a black background) closely resembles dead rushes that
litter the flooded meadows. When chased in shallow water, snakes move quickly,
change direction sharply, then lie motionless at the bottom. This behavior is an
effective escape tactic against visual-based predators such as humans. The difference
116
in coloration between lakeshore and meadow ecotypes may therefore be a result of
differential selection for crypticity.
We also expected to see adaptive differences between T. elegans ecotypes in
vertebral numbers. Kelley et al. (1997) and Arnold and Phillips (1999) argued that
selection arising from differences in vegetation density between coastal California and
the Eagle Lake shoreline is responsible for higher body and tail vertebral counts in
lakeshore T. elegans. Snakes use substrate irregularities as points against which their
vertebral column can push and propel themselves forward (Gray 1946, 1953; Gans 1986;
Jayne 1988; Gasc et al. 1989). In habitats with a higher density of these "push points," T
elegans populations have fewer vertebrae, whereas habitat with fewer push points support
garter snakes with more vertebrae (Kelley et al. 1997). Push points are more abundant in
habitat with dense vegetation, as in meadows, and less abundant in rocky habitat, as at
lakeshore sites. Kelley et al. (1997) also examined locomotory performance of T
elegans with different vertebral counts at varying push-point densities and found that
snakes with more vertebrae moved slower than snakes with fewer vertebrae over all
push point densities. Because lakeshore habitats provide fewer push points than
meadow habitats, we expected to see more body and tail vertebrae (ventral and
subcaudal scale counts) in lakeshore than in meadow T. elegans. In contrast, we did
not expect to see differentiation or differential selection on other scalation traits.
Coloration and scale counts have been shown to be under selection in these and
other populations of garter snakes (Arnold 1988, Arnold and Bennett 1988; Bittner and
King 2003). Tn this report we use estimates of neutral divergence among populations at
117
microsatellite loci to determine whether the quantitative traits have experienced diversifying
selection, especially among ecotypes. We can reject neutrality as an explanation of
population differentiation in quantitative traits if QST FST. QST > Fsr suggests diversifying
selection, while QST <FST indicates stabilizing selection (Lande 1992, Spitze 1993). We
expect to find QST estimates that far exceed FST in both scale counts and coloration.
Because we expect differentiation to coincide with ecotypic differences, QST should be
higher between ecotypes than among populations within ecotypes.
METHODS
Population dferentiation in scale counts
Snakes were collected by hand from six localities at and around Eagle Lake in Lassen
County, California (Fig. 4.1, Table 4.1). The following six scale counts were made
on 2251 preserved and live specimens: number of ventral scales on the body (VENT);
number of subcaudal scales (SUB); total number of ventral scales (TOTAL); total
number of total number of infralabial (ILAB), supralabial (SLAB), postocular (POST)
scales on the left and right sides; and number of dorsal scale rows at midbody (MID),
as described by Arnold and Phillips (1999). VENT and SUB correspond, respectively,
to the numbers of body and tail vertebrae (Alexander and Gans 1966, Voris 1975).
Missing values comprised less than 4% of the dataset, three-fourths of which were
N
Eagle
WDC
2
(AL
PIK
0 MAH
PAP.
.
NML
MCY
5km
Fig. 4.1. Map showing locations of meadow (MCY, PAP, NML, MAH) and lakeshore (P1K, GAL)
sites. An additional lakeshore site, WDC, was used to generate heritabilities for scale counts.
00
119
Table 4.1. Names, abbreviations, ecotype and sample sizes of study populations for
scalation and color variables (for males and females) and microsatellite markers.
* Study site name is informal only, not an official geographic place name.
Population name
Gallatin Shoreline*
Mahogany Lake
McCoy Flat Res.
Nameless Meadow*
Papoose Meadows
Pikes Point
Abbreviation
GAL
MAH
MCY
NML
PAP
P1K
Scalation
Color
Ecotype
M
F
M
F Microsat
lakeshore 387 406 363 382
56
meadow 56 100 48
65
91
meadow 72 116 23 27
16
meadow 41 109 57
55
29
meadow 62 111 36
56
140
lakeshore 346 445 77 79
48
attributed to missing tail tips (SUB). Scale counts do not change during the ontogeny
of an individual. Sex was determined by eversion of hemipenes.
Inheritance
of scale
counts
Estimates of heritability for the scale count traits were taken from Arnold and Phillips
(1999). Those estimates are based on mother-offspring regressions using a sample of 102
mothers and 911 offspring from the Pikes Point and Wildcat Point (WDC; see Fig. 4.1)
populations. We used the average of male and female heritabilities, estimated from the
inland genetic and phenotypic variances given in Tables 3 and 4 of Arnold and Phillips
(1999).
Population differentiation in color
Coloration traits were scored on 1268 live T elegans from six populations
(Mahogany, McCoy-North, Nameless, Papoose, Pikes, Gallatin). Population samples
120
ranged from 50-745 individuals. Three aspects of coloration were scored by matching
each individual to color standards: darkness of the unstretched dorsal area between the
dorsal and lateral stripes at midbody (Background), colors of the dorsal (Dorsal) and
lateral (Lateral) stripes at midbody. In all three cases, to make a match, individual
snakes were held directly over a series of color standards in diffuse natural light. A
Kodak gray scale was used as a standard for Background. Five adjacent categories of
gray on this scale (arbitrarily denoted 3-7) were represented in the total sample.
Pantone color swatches were used as a standard for Dorsal and Lateral. Although
snakes were compared to over 36 swatches, matches were made to only 10 Pantone
colors (127, 128, 134, 135, 136, 137, 141, 148, 155, 162). Because these are arbitrary
color codes, they were translated to two different, three-element vector systems (red,
green, blue or RGB and hue, saturation, lightness or HSL) using Pantone related
software (Colorist) and Powerpoint. All analyses of Dorsal and Lateral were
conducted using the RGB and HSL vectors rather than the Pantone categories. Both
vector systems used because they provided slightly different, complimentary
perspectives on coloration. The sexes were separately analyzed because of a slight
sexual difference in coloration. Variable abbreviations represented all combinations of
stripe and color component; for example, saturation of the dorsal stripe was DORSAT
(Table 4.2). Background color (BKGRND) was quantified as various degrees of
darkness, with higher numbers corresponding to a darker background.
The RGB classification method uses the relative intensities of different types
of wavelengths to describe the net color in terms of short (red), medium (green) and
121
long (blue) wavelengths. Many vertebrates (e.g., humans; Hurvich 1981) with visual
systems have three types of receptors for detecting wavelength classes (Levine and
MacNichol 1979, Lythgoe 1979, Jacobs 1981). Although birds have four receptor
types (Chen and Goldsmith 1986, Jane and Bowmaker 1988), the RGB system should
better approximate the visual perception of avian predators. HSL, on the other hand,
is derived from Munsell codes (Munsell Color Company 1976), which classifies color
into groups based on human perception. We therefore expect greater selection to be
acting on RGB values than on HSL.
Inheritance of coloration scores
Heritabilities of the three coloration traits were assessed using a sample of 325
individuals representing 35 sibships from the Pikes and Wildcat populations. All
coloration scores were made on neonates, no more than one month after birth.
Coloration traits appeared to be fully expressed at birth, and no obvious ontogenetic
trends were observed when individuals were reared to maturity in the laboratory.
Heritabilities (h2) were estimated by treating the sibships as unrelated sets of fullsibs
using software (H2B00T; Phillips 1998) available at a website maintained by P.C.
Phillips (http://www.uoregon.edulpphillsofiware.html). Standard errors of
heritability were estimated and tests of the hypothesis that h2
0 were conducted
using 1000 bootstrap samples in H2BOOT. Fullsib anova (Falconer and McKay 1996)
rather than mother-offspring regression was used to estimate heritabilities because
coloration scores for mothers were missing for over a third of the sib ships.
122
Heritabilities were separately estimated for males (184 individuals in 35 sibships) and
females (141 individuals in 32 sibships), and the average of the separate sex estimates
was used in QST analyses.
Microsatellites
Tissue samples were collected between 1999 and 2004, as described in Chapter 2. Briefly,
tail tip or piece of ventral scale was clipped and stored in Drierite®, an anhydrous calcium
sulfate desiccant. The time lag between collection of scale count data and microsatellite
data should not affect the results, because heritability estimates are not expected to change
on the scale of the time difference between the two datasets (Jones et al. 2004). Genetic
variation at neutral loci was assessed for nine microsatellite markers (see Table 2.2; Chapter
2). Whole genomic DNA was extracted using sodium dodecyl sulphate-proteinase K
digestion followed by a standard phenol-chloroform extraction, NaC1 purification and
isopropanol precipitation. DNA was PCR amplified in a 12.5 j.tL reaction with 10 mM
Tris-HC1 (pH 9.0), 50mM KC1, 0.1% Triton X-100, 0.2 mM each of dNTPs, 1.5 mM
MgCl2, 0.48 !tM forward (labeled with fluorescent ABI dye) and reverse primer, and
0.3 U Taq DNA polymerase. PCR profiles consisted of 94 °C for 2 mm followed by
36 cycles of 94 °C for 30 sec, appropriate annealing temperature for 30 sec and 72 °C
for 30 sec, ending with 72 °C for 2 mm. PCR products were separated using an ABI
3100 capillary electrophoresis genetic analyzer and data were visualized using
Genotyper 3.7 (ABI Prism).
123
Data analysis
Quantitative traits. Differences between males and females at scale counts and color scores
were calculated using ANOVA in SAS (v. 9.1, SAS Institute 2002). All populations were
pooled to assess sexual dimorphism over the entire study area.
For all traits and within sexes, phenotypic variance was partitioned within and
among populations, as well as within and between ecotypes (lakeshore and meadow), using
a nested analysis of variance (GLM in SAS). We tested for ecotypic differences using the
mean square among populations within ecotypes to compute the F-ratio. Tests for
population differences within ecotypes were conducted using the error variance @ooled
within population variance).
We calculated QST using the formula QST = (r2Gs/(
(I2GB
+2 02(jw), where CGB is the
among-population component of genetic variance, and 02GW is the within-population
component of genetic variance (Wright 1945, 1969 ; Lande 1992; Spitze 1993). The
among-population component of phenotypic variance was equated with O2GB. The withinpopulation component of genetic variance was computed by multiplying the withinpopulation component of phenotypic variance by the heritability of the trait in question.
Molecular traits. Analysis of genetic data were as described in Chapter 2. These analyses
included exact tests for departure from Hardy-Weinberg equilibrium (Guo and
Thompson 1992; Markov chain parameters: 5000 dememorizations; 500 000 steps per
chain) calculated in ARLEQUIN v. 2.000 (Schneider et al. 2000) and tests for linkage
disequilibrium (Slatkin and Excoffier 1996; Markov chain parameters: 5000
dememorizations, 1000 batches, 5000 iterations per batch), performed in GENEPOP
124
(Raymond and Rousset 1995). Significance levels were adjusted for multiple
comparisons (Rice 1989). Number of alleles and observed and expected
heterozygosities in each population and over all populations were calculated in
GENEPOP (Raymond and Rousset 1995).
Overall and population pairwise estimates of FST were calculated using AMOVA
(Excoffier et al. 1992) in ARLEQUIN
V.
2.000. Significance was assessed after 16 000
permutations for global estimates and 3000 permutations for pairwise estimates, with
P-values adjusted with the sequential Bonferroni correction.
Statistical comparisons. A Mantel test (Mantel 1967, Mantel and Valand 1970, Manly
1997), implemented in ARLEQUIN v. 2.000 was used to assess the correlation between
pairwise estimates ofFST and
QST for
each trait (significance over 10 000
permutations).
RESULTS
After sequential Bonferroni correction for multiple comparisons (Rice 1989), we found
sexual dimorphism for two scale counts (VENT and SUBC) and four color scores
(DORGREEN, DORHUE, LATGREEN and LATHUE; Table 4.2). Consequently, we
analyzed males and females separately. Sample sizes, means and standard deviations for
each trait are given for males and females within populations (Table 4,3) and ecotypes
(Table 4.4).
125
Table 4.2. Tests of sexual dimorphism for scale counts
and color scores. N indicates numbers of males and
females.
N
Scale count
VENT
SUBC
TOTAL
MID
SLAB
ILAB
POST
Color score
DORRED
DORGREEN
DORBLUE
DORHUE
DORSAT
DORLT
LATRED
LATGREEN
LATBLUE
LATHUE
LATSAT
LATLT
BKGRND
2162
Adjusted means
males females
2234
2238
2235
171.07
86.00
255.29
20.14
16.05
20.15
6.04
1254
1254
1254
1254
1254
1254
1124
1124
1124
1124
1124
1124
1267
243.47
192.50
110.04
24.84
204.31
166.14
243.82
202.80
123.04
26.43
201.71
172.44
5.14
1869
1848
2231
167.63
80.32
246.07
20.16
16.01
20.17
6.11
243.56
189.39
109.32
23.91
204.61
165.82
243.91
198.54
122.20
25.09
201.82
172.06
5.13
P
<0.0001
<0.0001
<0.0001
0.5846
0.053
0.3928
0.0206
0.7552
<0.0001
0.4923
<0.0001
0.7194
0.5373
0.7741
<0.0001
0.4398
<0.0001
0.9246
0.4287
0.7941
Table 4.3. Sample sizes, means and standard deviations of phenotypic traits for males and females in each population.
Gallatin
Trait
Scale counts
VENT
SUBC
TOTAL
MID
SLAB
ILAB
POST
Color scores
DORRED
DORGREEN
DORBLUE
DORHUE
DORSAT
DORLT
LATRED
LATGREEN
LATBLUE
LATHUE
LATSAT
LATLT
BKGRND
N
Males
Mean
367 170.3
307 84.1
304 254.5
384 20.5
386 16.0
386 20.3
386
6.2
356 243.5
356 195.9
356 114.7
356 25.2
356 203.4
356 168.4
305 243.4
305 201.9
305
125.2
305
25.8
200.1
305
305
363
173.3
4.8
Mahogany
Females
Mean SD
N
Males
Mean SD
170.1
4.4
55
167.1
6.4
97
163.7
84.0
8.4
48
10.7 48
0.9 56
0.4 55
0.6 55
0.6 56
78.8
245.3
19.9
16.0
10.6
77
76.1
15.0
238.9
0.4
0.5
0.4
77
97
100
99
99
48
12.2 48
19.7 48
245.1
1.8
65
245.1
1.5
23
185.9
100.6
23.9
210.6
162.4
14.7
65
182.6
12.7
13.8
65
97.8
2.8
4.8
65
23.3
65
211.2
6.9
65
161.1
2.3
65
245.4
13.8
65
11.0
65
4.0
8.6
65
65
5.1
65
0.4
65
SD
N
4.9
8.6
0.8
0.3
0.7
0.7
373
317
315
401
405
405
405
5.2
375
243.3
13.8
192.0
17.8
375
375
3.7
375
24.1
15.7
375
202.9
8.8
375
167.8
5.3
324
324
243.2
324
3.8 324
19.3 324
8.2 324
0.7 382
125.4
11.5
11.9
18.0
McCoy
254.0
20.4
16.0
20.2
6.2
113.7
198.7
24.8
198.7
173.2
4.7
5.3
2.9
16.0
48
19.8
6.0
48
9.7 48
5.4 48 245.5
12.6 48 203.2
21.8 48 116.7
3.5 48 27.3
21.2 48 208.6
9.7 48 170.3
0.6 48
6.2
1.0
Females
N Mean SD
Males
Mean SD
N
168.6
7.9
115
82.9
251.4
9.5
12.5
19.8
1.2
15.9
0.5
19.8
5.9
0.9
0.7
99
78.9
98 241.5
114 19.9
114 16.0
115
19.9
114
5.9
0.6
0.8
0.8
2.6
19.0
27
2.2
23
244.0
191.9
12.8
23
103.8
15.6
23
4.4
23
25.7
206.9
23
163.5
7.5
23
243.1
2.8
197.5
2.0
2.9
6.6
3.9
9.4
23
8.7
116.2
14.5
23
25.3
2.0
23
217.3
125.8
31.5
207.9
169.8
6.2
15.9
23
199.5
8.8
6.0
0.4
23
173.7
4.7
23
6.5
0.5
19.7
16.0
20.0
6.1
N
5.6
72
8.1
68
10.6 68
1.1
72
0.4 72
0.9 72
0.6 71
8.3
8.7
3.2
27
27
27
27
27
27
27
27
27
27
27
27
Females
Mean SD
163.1
6.4
8.2
9.8
1.2
244.4
184.8
15.6
98.5
13.4
24.0
3.2
209.3
6.0
161.1
6.6
244.9
207.7
119.6
28.4
206.3
171.4
2.9
6.3
12.1
9.1
4.1
9.8
4.2
0.4
Table 4.3.
(continued)
Nameless
Pikes
Papoose
N
Males
Mean
SD
N
VENT
SUBC
TOTAL
40
168.8
5.6
105
164.0
5.4
62
169.7
5.0
111
165.8
26
78.1
12.9
46
75.7
7.8
55
80.9
9.4
93
26
245.2
16.3
46
238.4
11.0
55
250.7
10.5
MID
41
19.5
0.9
109
19.7
1.1
62
19.4
0.9
SLAB
39
15.9
0.4
108
16.0
0.2
62
16.0
0.4
110
ILAB
39
19.9
0.4
107
20.0
1.0
62
19.9
0.7
110
POST
39
6.0
0.4
107
6.1
0.2
61
6.0
0.4
57
244.5
1.6
55
245.1
1.4
35
243.6
57
179.4
11.6
55
181.7
11.7
35
57
94.0
10.6
55
97.5
13.2
57
22.9
2.0
55
23.0
57
210.2
3.7
55
57
159.0
5.4
56
245.8
56
Females
Mean SD
Males
Females
Mean
SD
N
Males
Mean
SD
N
5.5
336
173.7
5.2
429
168.9
4.6
72.8
11.5
330
90.9
5.6
403
80.8
4.3
93
239.2
12.3
323
264.4
7.8
394
249.8
7.0
110
19.8
1.2
343
20.0
1.0
442
20.3
1.0
15.9
0.4
345
16.0
0.6
442
16.1
0.8
19.8
0.8
343
20.2
0.8
441
20.4
0.9
109
6.1
0.5
345
5.9
0.7
443
6.0
0.7
3.2
57
245.2
2.1
77
241.4
6.2
79
241.1
6.0
188.3
17.6
57
188.3
13.6
77
192.6
9.4
79
190.4
11.2
35
100.6
14.7
57
104.6
16.6
77
112.3
14.6
79
113.2
19.2
1.4
35
24.9
3.8
57
23.9
3.2
77
24.8
2.0
79
24.2
2.2
211.3
2.0
35
206.7
9.3
57
210.2
6.1
77
198.4
17.9
79
196.9
18.3
55
160.9
6.8
35
161.7
7.1
57
164.3
8.2
77
166.2
7.5
79
166.5
9.4
2.2
52
246.2
2.7
33
244.4
3.2
52
245.4
3.3
70
242.7
5.6
69
242.6
5.6
201.7
12.2
52
194.9
7.3
33
207.4
15.1
52
200.8
11.5
70
200.2
8.5
69
196.4
10.0
56
115.6
9.0
52
111.4
9.7
33
120.5
13.4
52
119.4
14.4
70
124.3
19.1
69
124.1
20.2
56
26.8
3.7
52
24.8
0.9
33
28.6
4.4
52
26.0
3.3
70
25.5
3.2
69
24.4
2.8
LATSAT
LATLT
56
209.6
8.3
52
211.8
9.4
33
203.9
12.7
52
207.4
14.1
70
197.4
22.0
69
197.7
20.5
56
169.8
4.0
52
167.9
4.5
33
171.6
5.8
52
171.5
6.1
70
172.5
8.1
69
172.5
BKGRND
57
6.0
0.3
55
6.0
0.4
35
6.1
0.4
56
6.1
0.5
77
4.6
0.5
79
4.5
Trait
N Mean SD
N
Females
Mean
SD
Scale counts
Color scores
DORRED
DORGREEN
DORDLUE
DORHUE
DORSAT
DORLT
LATRED
LATGREEN
LATBLUE
LATHUE
9.3 0.6
Table 4.4. Sample sizes, means and standard deviations of traits for males and females in lakeshore and meadow habitats.
Difference between means for the two ecotypes is expressed in units of average phenotypic standard deviation.
Lakeshore
Meadow
Ecotypic
Males
Females
Males
Females
difference in SD
Trait
N Mean SD
N Mean SD
N Mean SD
N Mean SD
Males
Fem
Scale counts
VENT 703 171.9 5.3
802 169.5 4.6
229 168.6 6.5
428 164.2 5.9
0.56
1.02
SUBC 637 87.6 7.9
720 82.2
6.6
197 80.7 10.4
315 76.0 9.5
0.76
0.77
TOTAL 627 259.6 10.9
709 251.7 9.1
197 248.9 13.4
314 239.7 11.0
0.88
1.19
MID 727 20.3
1.0
843 20.4
1.0
231
19.7
1.1
430 19.8
1.1
0.60
0.56
SLAB 731 16.0
0.5
847 16.1
0.6
228 15.9
0.4
432 16.0 0.4
0.22
0.15
ILAB 729 20.2
0.7
846 20.3
0.8
228 19.8
0.7
431
19.9
0.9
0.58
0.50
POST 731
6.1
0.7
848
6.1
0.7
227 5.9
0.5
429 6.0
0.6
0.23
0.15
Color scores
DORRED 433 243.1 5.5
163 242.9 5.5
454 244.4 2.3
204 245.0 1.8
0.33
0.59
DORGREEN 433 195.3 13.2
163 191.7 12.0
454 185.0 15.6
204 184.3 13.3
0.72
0.59
DORBLUE 433 114.3 17.3
163 113.6 19.6
454 98.7 13.6
204 99.8 14.4
1.01
0.82
DORHUE 433 25.1
3.5
163 24.1
2.8
454 24.0
3.2
204 23.5
2.5
0.33
0.24
DORSAT 433 202.5 16.2
163 201.9 16.6
454 209.1 6.5
204 210.7 4.4
0.59
0.84
DORLT 433 168.0 8.6
163 167.6 9.7
454 161.2 6.7
204 162.0 7.2
0.88
0.66
LATRED 375 243.3 5.3
160 243.1 5.4
393 245.0 2.7
196 245.5 3.3
0.43
0.56
LATGREEN 375 201.6 11.3
160 198.3 12.2
393 205.6 13.8
196 199.1 10.7
0.32
0.07
LATBLUE 375 125.0 18.2
160 125.1 21.5
393 118.4 11.1
196 116.2 13.0
0.45
0.52
LATHUE 375 25.8 3.7
160 24.7
3.4
393 28.0
4.2
196 25.8
2.8
0.55
0.33
LATSAT 375 199.6 19.9
160 198.5 21.1
393 206.7 10.1
196 208.6 13.2
0.47
0.59
LATLT 375 173.1 8.2
160 173.1 9.7
393 170.9 5.0
196 170.0 5.6
0.34
0.41
BKGRND 440 4.8
0.7
4.7
0.6
163
461
6.1
0.4
203
6.1
0.4
2.45
2.59
129
Many traits showed significant differences among populations within ecotypes, and
a few showed significant differences between the two ecotypes (Table 4.5). More traits
showed ecotypic differences in females than in males. After Bonferroni correction, MID,
LATRED, LATSAT and BKGRND showed significant divergence between ecotypes.
VENT, SUBC, ILAB, and several aspects of stripe coloration were significantly different
between ecotypes before, but not after, Bonferroni correction in females.
Mahogany Lake was out of Hardy-Weinberg equilibrium at one microsatellite
locus, and we found no evidence for linkage disequilibrium (Chapter 2). FST was 0.026 (P
> 0.00001) among populations and 0.014 (P = 0.065) among ecotypes.
Heritability estimates of color traits ranged from 0.072 (LATRED) to 0.654
(DORI{UE), with an average of 0.363 (Table 4.6). BKGRND heritability was also high
(0.640). For male color scores, among-population QST averaged 0.193 and ranged from
0.038 (DORRED) to 0.560 (BKGRND), while the average for females was 0.182, ranging
Values of QST for scale counts and color scores were generally much higher than FST both
among populations and between ecotypes (Figure 4.2). For male scale counts, among-
population QST ranged from 0.058 (SLAB) to 0.402 (POST) and averaged 0.243. Amongpopulation QST for female scale counts was slightly lower on average (0.217) and varied
from 0.036 (SLAB) to 0.327 (ILAB; Table 4.6). Values for between-ecotype QST for males
averaged 0.239 and ranged from 0.142 (POST) to 0.479 (ILAB), and females averaged
0.255, varying from 0.065 (SLAB) to 0.410 (ILAB). Overall, QST was highest for ILAB
and TOTAL but lowest for SLAB and MID. QST among populations and between ecotypes
were statistically equivalent for both sexes (P = 0.3 54 for males, P = 0.113 for females for
130
Table 4.5. Tests for differences between ecotypes and among populations within ecotypes
for each trait in males and females. Mean squares are given for ecotype and population
within ecotype. Asterisks indicate significance levels for differences among populations
within ecotypes. P-values are for differences among ecotypes; those in bold are significant
after sequential Bonferroni correction.
Trait
Scale counts
Ecotype
Males
Population
within Ecotype
VENT
SUBC
TOTAL
MID
1947.86
551.96****
3.53
0.1335
8004.92
188.45****
42.48
0.0029
7343.25
1976.08****
3.72
0.1261
8669.75
888.63****
9.76
0.0354
17530.50
4250.43****
4.12
0.1121
31411.93
898.07****
34.98
0.0041
62.89
16.28****
3.86
0.1208
98.78
0.96
102.83
0.0005
SLAB
1.66
0.09
18.17
0.013
1.74
0.28**
6.12
0.0687
ILAB
27.70
0.35
78.40
0.0009
47.38
2.63
18.04
0.0132
2.54
5.17****
0.49
0.5221
2.86
2.80****
1.02
0.3695
310.20
79.46**
3.90
0.1194
817.24
81.75**
10.00
0.0341
5696.46
1004.57***
5.67
0.0759
4963.28
403.55*
12.30
0.0247
17427.11
640.72*
27.20
0.0064
20378.51
481.14
42.35
0.0029
37.75
41.58**
0.91
0.3946
36.49
7.47
4.89
0.0916
5533.40
513.52*
10.78
0.0304
12058.82
623.06*
19.35
0.0117
2916.60
197.17*
14.79
0.0184
2977.28
137.94
21.58
0.0097
POST
Color scores
DORRED
DORGREEN
DORBLUE
DORHUE
DORSAT
DORLT
LATRED
LATGREEN
LATBLUE
Females
F-stat
P-value
Ecotype
Population
within Ecotype
F-stat
P-value
225.99
41.87
5.40
0.0808
652.45
13.10
49.80
0.0021
3531.40
1145.26****
3.08
0.1539
694.10
890.10****
0.78
0.4271
2285.43
511.05
4.47
0.1019
6493.07
538.76
12.05
0.0255
LATHTJE
718.56
103.83****
6.92
0.0581
228.34
69.44****
3.29
0.1440
LATSAT
LATLT
3879.18
614.51
6.31
0.0659
10254.48
210.61
48.69
0.0022
203.74
80.41
2.53
0.1867
731.02
108.44
6.74
0.0603
BKGRND
205.68
2.06***
99.61
0.0006
240.30
1.47**
163.19
0.0002
* P <0.05, ** P <0.01,
P <0.001,
< 0.0001
CD
rI)
CD
CD
H
CD
-t
0
CD
-t
-t
OC#D
CD
CD
CD
QcdD
I4)
CIDO
OH
0
>1j
p
(0
(0
o
p
Q,
p
p
p
00
BKGRND
C'
(0
o
p
p
0'
-
p
00
p
1
(jJ
133
one-tailed paired t-tests). from 0.012 (DORHUE) to 0.608 (BKGRND). Among-ecotype
QST for
males averaged 0.204 and varied from 0.004 (DORGREEN) to 0.648 (BKGRND),
and females averaged 0.228, ranging from -0.002 (LATGREEN) to 0.689 (BKGRND).
Overall, BKGRND was by far the most divergent trait, with a stronger difference between
ecotypes than among populations for both sexes. Background color also differed between
the two ecotypes by over two standard deviations (Table 4.2). DORBLUE and LATRED
were highly differentiated, with QST'S comparable to the highest estimates for scale counts.
Color scores on the dorsal and lateral stripe were approximately equally divergent (P =
0.467), as were RGB and HSL scores (P= 0.351). Color divergence was greater between
ecotypes than among populations for females (P = 0.008) but not males (P
0.298).
Mantel tests of pairwise FST and QST matrices showed no evidence of correlated
patterns of population differentiation. F51 and QST were not significantly correlated for any
trait, and Pearson correlation coefficients varied from -0.23 7 to 0.3 02 for males and -0.283
to 0.290 for females. Phenotypic variance was significantly partitioned among ecotypes
relative to variance among populations within each ecotype for more traits in females than
males (Table 4.6). After Bonferroni correction, MID, LATRED, LATSAT and BKGRND
showed significantly more divergence between ecotypes than among populations within
ecotype. VENT, TOTAL, DORBLUE and DORLT had suggestive but inconclusive
results.
In meadow populations the dorsal stripe tends to be orange, but in lakeshore
populations it is light brown or even pink (Fig. 4.3). Lateral stripe color shifts from tarmish
light orange
134
Table 4.6. Heritabilities of traits and global Qsi among populations
and between ecotypes for males and females.
Trait
Scale counts
VENT
SUBC
TOTAL
MID
SLAB
ILAB
POST
Color scores
DORRED
DORGREEN
DORBLUE
DORHTJE
DORSAT
DORLT
LATRED
LATGREEN
LATBLUE
LAT}{UE
LATSAT
LATLT
BKGRND
Heritability
Amongpopulation Qst
males females
Betweenecotype Qst
males females
0.475
0.465
0.47
0.42
0.06
0.09
0.06
0.178
0.300
0.356
0.198
0.058
0.320
0.402
0.279
0.235
0.350
0.104
0.036
0.327
0.189
0.152
0.258
0.313
0.183
0.147
0.479
0.142
0.368
0.264
0.447
0.164
0.065
0.410
0.069
0.476
0.474
0.255
0.654
0.375
0.298
0.072
0.402
0.245
0.462
0.102
0.266
0.640
0.038
0.178
0.365
0.042
0.101
0.265
0.268
0.128
0.109
0.165
0.239
0.055
0.560
0.091
0.125
0.280
0.012
0.171
0.185
0.304
0.055
0.112
0.081
0.276
0.063
0.608
0.031
0.004
0.466
0.035
0.123
0.363
0.300
0.057
0.137
0.144
0.274
0.073
0.648
0.095
0.157
0.360
0.017
0.207
0.242
0.463
-0.002
0.176
0.047
0.405
0.108
0.689
in meadow snakes to light brown in lakeshore snakes (Fig. 4.4). Lakeshore populations
exhibit wider variation in dorsal and lateral stripe than meadow populations, with lower
frequencies of meadow-type color patterns. Background coloration shifts from
predominantly dark shades at meadow sites to lighter grays at lakeshore sites (Fig. 4.5).
135
MALES
FEMALES
McCOY
::
0.4
0.4
-
0.2
0.2
_____
08
:
MAHOGANY
06
0.4
McCOY
::
- _____
-nfl
MAHOGANY
0.8
0.6
0.4
-
0.2
0.2
____ -._____
NAMELESS
0.8
0.6
04
0.6
T
0.4
-
02
0.2
PAPOOSE
0.8
06
PAPOOSE
0.8
0.6
-
04
02
NAMELESS
0.8
0.4
flfl
0.2
Iii
GALLATThT
0.8
0.8
fi
GALLATIN
0.2
flfl
PIKES
0.8
06
06
0.4
0.4
0.2
flfl
fl
DORSAL STRIPE COLOR
PIKES
0.8
0.2
[p fl
DORSAL STRIPE COLOR
Fig. 4.3. Frequency histograms showing distribution of dorsal stripe
color for males and females in each population. Populations are shown
in order from farthest to closest to Eagle Lake.
136
FEMALES
08
McCOY
:
0.6
MALES
McCOY
0.8
:
0.6
-
04
0.2
0.2
lHn
08
-
-
MAHOGANY
flH
MAHOGANY
0.8
06
0.6
0.4
-
0.4
0.2
fl
I
fl
NAMELESS
0.8
0.2
NAMELESS
8
0.6
0.4
0.2
2
__ _n
-II
PAPOOSE
0.8
=
0.6
04
PAPOOSE
0.8
0.6
-
0.2-
-n n
0.4
-
0.2
:fl
n
n ii
GALLATIN
0.8
06
GALLATIN
0.8
0.6
::
PIKES
0.8
06
-
0.6
0.4
0.2:
PIKES
0.8
0.4
fl
fl
LATERAL STRIPE COLOR
-
0.2
LATERAL STRIPE COLOR
Fig. 4.4. Frequency histograms showing distribution of lateral strip
color for males and females in each population. Conventions as for
Fig. 4.3.
137
FEMALES
0.8
McCOY
MALES
0.8
0.6
0.6
:.:
::
MAHOGANY
LI.
08
AMELES1
I AHOGAN
LI
08
0.6
0.6
04
0.4
0.2
0.2
0.8
PAPOOSE
0.8
0.6
0.6
0.4
0.4
_.
0.2
08
-GALLATIN
McCOY
.JAMELESS
PAPOOSE
_.
0.2
0.8
02J1
0.8
PIKES
0.8
0.6
0.6
0.4
0.4
0.2
0.2
BACKGROUND
PIKES
BACKGROUND
Fig. 4.5. Frequency histograms showing distribution of
background color for males and females in each
population. Conventions as for Fig. 4.3.
138
There are very low frequencies of snakes with meadow-like background color at lakeshore
sites, consistent with the high QST values for BKGRNI) we observed.
DISCUSSION
Coloration and scale counts showed strong differentiation between lakeshore and meadow
sites as well as weaker differentiation among populations within ecotypes. This
differentiation cannot be attributed to drift. We found strong diversifying selection on both
scale counts and color scores. Only three out of 80 Q5T estimates were lower than F51, and
almost 75% were greater than FST by a factor of four or more. QST for background color
was consistently high for both sexes among populations and between ecotypes (range of
0.560 to 0.689). There was significant diversifying selection on numbers of body vertebrae
(0.152 to 0.368), but a less marked difference with tail vertebrae (0.235 to 0.300). Higher
numbers of vertebrae in lakeshore populations may be a result of differential selection on
locomotory performance in a lakeshore versus meadow habitat. A difference in push-point
density between lakeshore and meadow habitats may explain the observed differentiation in
ventral and subcaudal scale counts.
Between-ecotype QST values for females for body and total vertebral counts are
higher than males, suggesting that the two sexes are experiencing different intensities of
selection. Garter snakes exhibit sexual dimorphism in size such that females are larger
(Rossman et al 1996). Garter snakes also bear live young so females become even larger
139
with pregnancy, decreasing their speed. Because locomotor performance has important
fitness consequences, any functional advantage gained by females is likely to be under
strong selection. Interestingly, we also observed stronger selection for females on color
traits, despite a lack of overall of sexual dimorphism in color pattern. None of the four color
scores that showed significant differences between the sexes had
QST'S
greater than 0.2.
Again, larger-bodied females may rely more on crypticity than males, especially when
gravid, because their burst speed may be compromised. Both speed and crypticity are
important anti-predator mechanisms for garter snakes, and local adaptation in these traits at
the small geographic scale of this study point to strongly divergent selection on females in
different habitats. Further work is needed to determine if sexual dimorphism in selection for
crypticity or locomotor performance is common in snakes with female-biased size
dimorphism.
The high QST estimates obtained for background color indicates that it is under
strong selection, probably due to its role in substrate matching and increasing crypticity.
Darker background coloration in meadow-type T
elegans,
in conjunction with the yellow
dorsal stripe, results in a snake that is cryptic in dark water with bright yellow vegetation.
Lakeshore snakes probably match their environment with more subdued colors, including a
lighter background. There was no discemable difference between the RGB and HSL color
scoring methods in revealing differentiation among populations or between ecotypes. Both
scoring schemes revealed population differences that could not be explained by random
drift.
140
Meadow and lakeshore populations are four to 20 km apart, yet we found
differentiation and evidence for diversifying selection on quantitative traits. Previous work
showed that Papoose Meadows was the major source of migrants in the region and
contributes approximately six effective migrants per generation to each lakeshore site
(Chapter 2). Our results indicate that diversifying selection on quantitative traits can
overcome gene flow of this magnitude and produce local adaptation on a small spatial scale.
Concluding remarks
Our results are consistent with the picture of ecotypic differentiation documented in
this system by Bronikowski and Arnold (1999). Differentiation in coloration and
scalation traits coincided with the ecotypic differences in life history and established
that those differences could not be attributed to drift. Departure from neutral
differentiation at these traits was stronger for females than for males, perhaps
reflecting locomotory constraints caused by relatively larger bodies and pregnancy.
Other studies have found much stronger divergence for quantitative traits than
expected under neutrality (e.g., Porter and Geiger 1995, Pfrender et al. 2000, Wong et
al. 2003), as well as a nonsignificant relationship between pairwise FST and QST (Palo
et al. 2003). Approaches to
FST-QST
comparisons have generally taken one of two
approaches. The first approach asks whether neutral markers can act as surrogates for
adaptive quantiative traits in natural populations that might be targets for conservation
management. The overall conclusion here is that genetic structuring among populations at
quantitative traits is usually greater than that observed with neutral markers (Pfrender et al.
141
2000, Reed and Frankham 2001, McKay and Latta 2002, Merilä and Cmokrak 2001) and
may be uncorrelated (Reed and Frankham 2001, McKay and Latta 2002). Though
convenient for conservation purposes, FST cannot be a proxy for QST, because neutral
markers will usually underestimate genetic variation available for evolutionary adaptation.
The other approach is to infer selection on quantitative traits by comparing their
differentiation with that at neutral markers. This approach has enormous potential for
strengthening our understanding of microevolutionary processes in diverse systems.
REFERENCES
Alexander, A.A. and C. Gans. 1966. The pattern of dermal-vertebral correlation in snakes
and amphisbaenians. Zoologische Mededelingen 41:171-190.
Arnold, S.J. 1988. Quantitative genetics and selection in natural populations:
microevolution of vertebral numbers in the garter snake Thamnophis elegans.
Second International Conference on Quantitative Genetics. Sinauer, Sunderland,
Massachussetts.
Arnold, S.J. and A.F. Bennet. 1988. Behavioural variation in natural populations. V.
Morphological correlates of locomotion in the garter snake (Thamnophis radix).
Biological Journal of the Linnean Society 34:175-190.
Arnold, S.J. and P.C. Phillips. 1999. Hierarchical comparison of genetic variancecovariance matrices. II. Coastal-inland divergence in the garter snake, Thamnophis
elegans. Evolution 53:1516-1527.
Baker, A.J. 1992. Genetic and morphometric divergence in ancestral European and
descendent New Zealand populations of chaffiches (Fringilla coelebs). Evolution
46: 1784-1800.
Bittner, T.D. and R.B. King. 2003. Gene flow and melanism in garter snakes
revisited: a comparison of molecular markers and island vs. coalescent
models. Biological Journal of the Linnean Society 79:389-399.
Bonnin, I., J.-M. Prosperi and I. Olivieri. 1996. Genetic markers and quantitative genetic
variation in Medicago truncatula (Leguminosa): a comparative analysis of
population structure. Genetics 143:1795-1805.
Bronikowski, A.M. and S.J. Arnold. 1999. The evolutionary ecology of life history
variation in the garter snake Thamnophis elegans. Ecology 80:23 14-2325.
142
Cano, J.M., A. Laurila, J. Palo and J. Merilä. 2004. Population differentiation in G matrix
stmcture due to natural selection in Rana temporaria. Evolution 58:2013-2020.
Charlesworth, B. 1990. Natural selection and life history patterns. In Genetc Effects on
Aging II (D.E. Harrison, ed..), pp. 21-40. Caidwell, NJ: Telford Press.
Chen, D.-M. and T.M. Goldsmith. 1986. Four spectral classes of cone in the retinas of
birds. Journal of Comparative Physiology 1 59A:473 -479.
Excoffier, L., P.E. Smouse and J.M. Quattro. 1992. Analysis of molecular variance
inferred from metric distances among DNA haplotypes: application to human
mitochondrial DNA restriction data. Genetics 131:479-491.
Falconer, D.S. and T.F.C. MacKay. 1996. Introduction to quantitative genetics. 4th ed.
Harlow, U.K.: Prentice Hall.
Gans, C. 1986. Locomotion of limbless vertbrates: pattern and evolution. Herpetologica
42:33-46.
Gasc, J.-P., D. Cattaert, C. Chasserat and F. Clarac. 1989. Propulsive action of a snake
pushing against a single site: its combined analysis. Journal of Morphology
201:315-329.
Gomez-Mestre, I. and M. Tejedo. 2004. Contrasting patterns of quantitative and neutral
genetic variation in locally adapted populations of the natterjack toad, Bufo
calamita. Evolution 58:5343-5352.
Gray, J. 1946. The mechanism of locomotion in snakes. Journal of Experimental Biology
23: 101-1 19.
Gray, J. 1953. Undulatory propulsion. Quarterly Journal of Microscopical Science 94:551578.
Guo, S.W. and E.A. Thompson. 1992. Performing the exact test for Hardy-Weinberg
proportions for multiple alleles. Biometrics 48:36 1-372.
Hurvich, L.M. 1981. Color vision. Sunderland, Mass: Sinauer Associates, Inc.
Jacobs, G.H. 1981. Comparative color vision. New York: Academic Press.
Jane, S.D. and J.K. Bowmaker. 1988. Tetrachromatic colour vision in the duck (Anas
platyrhynchos L.): microspectrophotometry of visual pigments and oil droplets.
Journal of Comparative Physiology 162A:225-235.
Jayne, B.C. 1988. Muscular mechanisms of snake locomotion: an electromyographic
study of lateral undulation of the Florida Banded Water Snake (Nerodiafasciata)
and the Yellow Rat Snake (Elaphe obsoleta). Journal of Morphology 197:159-181.
Jones, A.G., S.J. Arnold and R. Btirger. 2004. Evolution and stability of the G-matrix on a
landscape with a moving optimum. Evolution 58:1639-1654.
Kelley, K.C., S.J. Arnold and J. Gladstone. 1997. The effects of substrate and vertebral
number on locomotion in the garter snake, Thamnophis elegans. Functional
Ecology 11:189-198.
King, R.B. 1993. Color-pattern variation in lake Erie water snakes: prediction and
measurement of natural selection. Evolution 47:1819-1833.
Lande, R. 1992. Neutral theory of quantitative genetic variance in an island model with
local extinction and colonization. Evolution 46:381-389.
143
Levine, J.S. and E.F. MacNichol. 1979. Visual pigments in teleost fishes: effects of
habitat, microhabitat, and behavior on visual system evolution. Sensory Processes
3:95-131.
Lythgoe, J.N. 1979. The ecology of vision. Oxford: Oxford University Press.
Manly, B.F. 1997. Randomization and Monte Carlo methods in biology. 21 ed.
New York: Chapman and Hall.
Mantel, N. 1967. The detection of disease clustering and generalized regression
approach. Cancer Research 27:209-220.
Mantel, N. and R.S. Valand. 1970. A technique of nonparametric multivariate
analysis. Biometrics 26:547-558.
McKay, J.K. and R.G. Latta. 2002. Adaptive population divergence: markers, QTL and
traits. Trends in Ecology and Evolution 17:285-291.
Merilä, 3. and P. Cmokrak. 2001. Comparison of genetic differentiation at marker loci and
quantitative traits. Journal of Evolutionary Biology 14:892-903.
Munsell Color Company. 1976. Munsell Book of Color, Glossy Finish Collection (2
volumes). Baltimore: Munsell/Macbeth/Koilmorgen Corp.
Palo, J.U., RB. O'Hara, A.T. Laugen, A. Laurila, C.R. Primmer and 3. Merilä. 2003.
Latitudinal divergence of common frog (Rana temporaria) life history traits by
natural selection: evidence from a comparison of molecular and quantitative
genetic data. Molecular Ecology 12:1963-1978.
Pfrender, M.E., K. Sptize, J. Hicks, K. Morgan, L. Latta, M. Lynch.. 2000. Lack of
concordance between genetic diversity estimates at the molecular and
quantitative trait levels. Conservation Genetics 1:263-269.
Phillips, P.C. 1998. H2boot: bootstrap estimates and tests of quantitative genetic
data. Univ. of Texas at Arlington. Software available at
www.uta.edu/biology/phillips/software.
Porter, A.H. and H. Geiger. 1995. Limitations to the inference of gene flow at
regional geographic scales: an example from the Pieris napi group
(Lepidoptera: Pieridae) in Europe. Biological Journal of the Linnean Society
54:329-348.
Prout, T. and J.S.F. Barker. 1993. F statistics in Drosophila buzzatii: selection,
population size and inbreeding. Genetics 134:369-375.
Raymond, M. and F. Rousset. 1995. GENEPOP Version 1.2: population genetics
software for exact tests and ecumenicism. Journal of Heredity 86:248-249.
Reed, D.H. and R. Frankham. 2001. How closely correlated are molecular and
quantitative measures of genetic variation? a meta-analysis. Evolution
55:1095-1103.
Rice, W.R. 1989. Analysing tables of statistical tests. Evolution 43 :223-225.
Rossman, D.A., N.B. Ford and R.A. Seigel. 1996. The garter snakes: evolution and
ecology. Norman, Oklahoma: University of Oklahoma Press.
SAS Institute. 2002. SAS/STAT software user's guide. Rel. 9.1. SAS Institute, Inc.,
Cary, NC.
144
Schneider, S., D. Roessli and L. Excoffier. 2000. Arlequin ver 2.000: a software for
population genetics data analysis. Genetics and Biometry Laboratory,
University of Geneva, Switzerland.
Slatkin, M. and L. Excoffier. 1996. Testing for linkage disequilibrium in genotypic
data using the EM algorithm. Heredity 76:377-383.
Spitze, K. 1993. Population structure in Daphnia obtusa: quantitative genetic and
allozyme variation. Genetics 135:367-374.
Steams, S.C. 1992. The Evolution of Life Histories. Oxford: Oxford University
Press.
Voris, H.K. 1975. Dermal scale-vertebra relationships in sea snakes (Hydrophidae).
Copeia 1975:746-755.
Waldmann, P. and S. Andersson. 1998. Comparison of quantitative genetic variation and
allozyme diversity within and between populations of Scabiosa canescens and S.
columbaria. Heredity 8 1:79-86.
Wright, S. 1943. An analysis of local variability of flower color in Linanthuspariyae.
Genetics 28:139-156.
Wright, S. 1969. The theory of gene frequencies, evolution and the genetics of populations.
Vol.2, Chicago: University of Chicago Press.
Wong, A., M.L. Smith and M.R. Forbes. 2003. Differentiation between subpopulations of
a polychromatic damselfly with respect to morph frequencies, but not neutral
genetic markers. Molecular Ecology 12:3505-35 13.
Yang, R.-C., F.C. Yeh and A.D. Yanchuk. 1996. A comparison of isozyme and
quantitative genetic variation in Pinus contorta ssp. latifolia by FST. Genetics
142:1045-1052.
145
MICROEVOLUTIONARY PAUERNS AND PROCESSES OF POPULATION
DIVERGENCE WITHiN A VERTEBRATE METACOMMUNITY
CHAPTER 5
GENERAL CONCLUSIONS AND FUTURE DIRECTIONS
Population divergence was documented in a natural system of coexisting vertebrates. First,
I examined the population structures of two coexisting species of garter snake,
Thamnophis elegans and T. sirtalis to determine if a shared landscape and common
biological history imposed similar population genetic structures. Similar patterns of
population differentiation were uncovered for both species, including source-sink
metapopulation dynamics and lack of migration-drift equilibrium (Chapter 2). I then
compared a third species, the western toad (Bufo boreas) to identify ecological and
evolutionary processes important in shaping the observed pattern of snake population
genetics. I found that B. boreas differend substantially in its population dynamics
from the garter snakes. The toad populations were characterized by much larger
effective population sizes, lower migration rates and migration-drift equilibrium
suggesting a stable population structure. A barrier to dispersal was also identified for
both garter snakes, and ecological variables associated with genetic differentiation
were identified (Chapter 3). Finally, I estimated genetic differentiation for two types
of quantitative traits and, using a null hypothesis of neutrality as measured by
differentiation at molecular markers, tested for the role of selection between two
different habitats for T. elegans in the vicinity of Eagle Lake. I found substantial
146
deviations from neutrality, indicating local adaptation for many of the traits examined,
most notably for traits associated with locomotory performance and crypticity.
Females were more divergent between habitats than males, perhaps as a result of
locomotory constraints imposed by large body size and gravidity (Chapter 4).
These results point to several different arenas of future research. The
similarities in genetic structure found for T elegans and T. sirtalis in the Eagle Lake
basin may be due to similarities in diet. This hypothesis could be tested by repeating
the analysis in a different system. Coastal regions in California and Washington also
support sympatric populations of both species, but there, T elegans prefers slugs, and
T. sirtalis retains an anuran diet. If similarities in diet are responsible for the
correlated genetic structure of Lassen Co. populations, coastal populations should
have uncorrelated genetic structures.
Further research is also needed to determine if the barrier to dispersal common
to both garter snakes also impedes gene flow in B. boreas. Breeding populations of
toads occur at Colman Lake and Deans Meadow. Further sampling will be necessary
to determine if any breeding populations east in that region are significantly
differentiated from those in the rest of the study area.
Complementary research is needed to quantify the selection strengths of
lakeshore and meadow habitats to further test the case for local adaptation in T
elegans. Potential projects could investigate performance of meadow-lake hybrids or
reciprocal exchanges in both habitats. Use of models mimicking color pattern
147
variation in both habitats could also be used to quantify predation on different color
morphs.
Finally, similar studies are needed that use multiple species in a community
format to ask questions regarding how ecological and evolutionary processes shape
microgeographic patterns in nature.
148
BIBLIOGRAPHY
Aleksiuk, M. and P.T. Gregory. 1974. Regulation of seasonal mating behavior in
Thamnophis sirtalis parietalis. Copeia 1974:681-689.
Alexander, A.A. and C. Gans. 1966. The pattern of dermal-vertebral correlation in snakes
and amphisbaenians. Zoologische Mededelingen 41:171-190.
Anderson, B., I. Olivieri, M. Lourmas and B.A. Stewart. 2004. Comparative
population genetic structures and local adaptation of two mutualists. Evolution
58:1730-1747.
Arnaud, J.-F., L. Madec, A. Guiller and A. Bellido. 2001. Spatial analysis of
allozyme and microsatellite DNA polymorphisms in the land snail Helix
aspersa (Gastropoda: Helicidae). Molecular Ecology 10:1563-1576.
Arnold, S.J. 1988. Quantitative genetics and selection in natural populations:
microevolution of vertebral numbers in the garter snake Thamnophis elegans.
Second International Conference on Quantitative Genetics. Sinauer, Sunderland,
Massachussetts.
Arnold, S.J. 1992. Behavioural variation in natural populations. VI. Prey responses
by two species of garter snakes in three regions of sympatry. Animal
Behaviour 44:705-719.
Arnold, S.J. and A.F. Bennet. 1988. Behavioural variation in natural populations. V.
Morphological correlates of locomotion in the garter snake (Thamnophis radix).
Biological Journal of the Linnean Society 34:175-190.
Arnold, S.J. and P.C. Phillips. 1999. Hierarchical comparison of genetic variancecovariance matrices. II. Coastal-inland divergence in the garter snake, Thamnophis
elegans. Evolution 53:15 16-1527.
Baker, A.J. 1992. Genetic and morphometric divergence in ancestral European and
descendent New Zealand populations of chaffiches (Fringilla coelebs). Evolution
46:1784-1800.
Banchs, I., A. Bosh, J. Guimera, C. Lazaro, A. Puig and X. Estivill. 1994. New
alleles at microsatellite loci in CEPH families mainly arise from somatic
mutations in the lymphoblastoid cell line. Human Mutation 3:365-372.
Barnosky, C.W., P.M. Anderson and P.J. Bartlein. 1987. The northwestern U.S.
during deglaciation; vegetational history and paleoclimatic implications. In
Geology of North America. North America and Adjacent Oceans During the
Last Deglaciation (Eds. W.F. Ruddiman and H.E. Wright, Jr.), Vol. K-3, pp.
289-321. Geological Society of America, Boulder.
Beerli, P. 1998. Estimation of migration rates and population sizes in geographically
structured populations. In: Advances in molecular ecology (Ed. G. Carvaiho),
pp. 39-53. NATO-ASI workshop series. lOS Press, Amsterdam.
Beerli, P. 2004. Effect of unsampled populations on the estimation of population
sizes and migration rates between sampled populations. Molecular Ecology
13:827-836.
149
Beerli, P. and J. Felsenstein. 1999. Maximum likelihood estimation of migration
rates and population numbers of two populations using a coalescent approach.
Genetics 152:763-773.
Beerli, P. and J. Felsenstein. 2001. Maximum likelihood estimation of a migration
matrix and effective population sizes in n subpopulations by using a coalescent
approach. Proceedings of the National Academy of Sciences 98:4563-4568.
Berven, K.A. 1982. The genetic basis of altitudinal variation in the wood frog Rana
sylvatica. I. An experimental study of life history traits. Evolution 36:962983.
Berven, K.A. and T.A. Grudzien. 1990. Dispersal in the wood frog (Rana sylvatica):
implications for genetic population structure. Evolution 44:2047-2056.
Bittner, T.D. and R.B. King. 2003. Gene flow and melanism in garter snakes
revisited: a comparison of molecular markers and island vs. coalescent
models. Biological Journal of the Linnean Society 79:389-399.
Bonnin, I., J.-M. Prosperi and I. Olivieri. 1996. Genetic markers and quantitative genetic
variation in Medicago truncatula (Leguminosa): a comparative analysis of
population structure. Genetics 143:1795-1805.
Brede, E.G. and T.J.C. Beebee. 2004. Contrasting population structures in two
sympatric anurans: implications for species conservation. Heredity 92:110117.
Bronikowski, A.M. and S.J. Arnold. 1999. The evolutionary ecology of life history
variation in the garter snake Thamnophis elegans. Ecology 80:2314-2325.
Bronikowski, A.M. and S.J. Arnold. 2001. Cytochrome b phylogeny does not match
subspecific classification in the western terrestrial garter snake, Thamnophis
elegans. Copeia 2001 :508-513.
Caballero, A. 1994. Developments in the prediction of effective population size.
Heredity 73: 657-679.
Cano, J.M., A. Laurila, J. Palo and J. Merilä. 2004. Population differentiation in G matrix
structure due to natural selection in Rana temporaria. Evolution 58:2013-2020.
Castric, V., F. Bormey and L. Bernatchez. 2001. Landscape structure and hierarchical
genetic diversity in the brook charr, Salvelinusfontinalis. Evolution 55:10161028.
Charlesworth, B. 1990. Natural selection and life history patterns. In Genetc Effects on
Aging II (D.E. Harrison, ed..), pp. 2 1-40. Caldwell, NJ: Telford Press.
Chen, D.-M. and T.M. Goldsmith. 1986. Four spectral classes of cone in the retinas of
birds. Journal of Comparative Physiology 159A:473-479.
Coulon, A., J.F. Cosson, J.M. Angibault, B. Cargnelutti, M. Galan, N. Morellet, E.
Petit, S. Aulagnier and A.J.M. Hewison. 2004. Landscape connectivity
influences gene flow in a roe deer population inhabiting a fragmented
landscape: an individual-based approach. Molecular Ecology 13:2841-2850.
Crawford, A.J. 2003. Huge populations and old species of Costa Rican and
Panamanian dirt frogs inferred from mitochondrial and nuclear gene
sequences. Molecular Ecology 12:2525-2540.
150
Crow, J.F. and M. Kirnura. 1971. The effective number of a population with
overlapping generations: a correction and further discussion. American
Journal of Human Genetics 24:1-10.
Crow, J.F. and N.E. Norton. 1955. Measurement of gene frequency drift in small
populations. Evolution 9:202-2 14.
Dallas, J.F. 1992. Estimation of microsatellite mutation rates in recombinant inbred
strains of mouse. Mammalian Genome 5:32-38.
de Queiroz, A., R. Lawson and J.A. Lemos-Espinal. 2002. Phylogenetic relationships
of North American garter snakes (Thamnophis) based on four mitochondrial
genes: how much DNA sequence is enough? Molecular Phylo genetics and
Evolution 22:315-329.
Dole, J.W. 1971. Dispersal of recently metamorphosed leopard frogs, Ranapipiens.
Copeia 1971:221-228.
Donahue, M.J., M. Holyoak and C. Feng. 2003. Patterns of dispersal and dynamics
among habitat patches varying in quality. American Naturalist 162:302-3 17.
Dupanloup, I., S. Schneider, A. Langaney and L. Excoffier. 2002. A simulated
annealing approach to define the genetic structure of populations. Molecular
Ecology 11:2571-2581.
Easteal, S. 1985. The ecological genetics of introduced populations of the giant toad
Bufo marinus. II. Effective population size. Genetics 110:107-122.
Edwards, A., H.A. Hammond, L. Jin, C.T. Caskey and R. Chakraborty. 1992.
Genetic variation at five trimeric and tetrameric tandem repeat loci in four
human population groups. Genomics 12:241-253.
Ellegren, H. 1995. Mutation rates at porcine microsatellite loci. Mammalian Genome
6:376-377.
Eriksson, J., G. Hohmann, C. Boesch and L. Vigilant. 2004. Rivers influence the
population genetic structure of bonobos (Pan paniscus). Molecular Ecology
13:3425-3435.
Excoffier, L., P.E. Smouse and J.M. Quattro. 1992. Analysis of molecular variance
inferred from metric distances among DNA haplotypes: application to human
mitochondrial DNA restriction data. Genetics 131:479-491.
Falconer, D.S. and T.F.C. MacKay. 1996. Introduction to quantitative genetics. 4th ed.
Harlow, U.K.: Prentice Hall.
Farr, D.R. and P.T. Gregory. 1991. Sources of variation in estimating litter
characteristics of snakes. Journal of Herpetology 25 :261-267.
Fitch, H.S. 1987. Collcting and life-history techniquest. In Snakes: Ecology and
Evolutionary Biology (Eds. R.A. Seigel, J.T. Collins and S.S. Novak), pp. 143164. McGraw-Hill, New York.
Frankham, R. 1995. Effective population size/adult population size ratios in wildlife:
a review. Genetical Research 66:95-107.
Gans, C. 1986. Locomotion of limbless vertbrates: pattern and evolution. Herpetologica
42:33-46.
Gamier, S., P. Alibert, P. Audiot, B. Prieur and J.-Y. Rasplus. 2004. Isolation by
distance and sharp discontinuities in gene frequencies: implications for the
151
phylogeography of an alpine insect species, Cara bus solieri. Molecular
Ecology 13:1883-1897.
Gasc, J.-P., D. Cattaert, C. Chasserat and F. Clarac. 1989. Propulsive action of a snake
pushing against a single site: its combined analysis. Journal of Morphology
201:315-329.
Geyer, C. and E.A. Thompson. 1994. Annealing Markov chain Monte Carlo with
Applications to Ancestral Inference. Technical report, University of
Minnesota Nr. 589 R(1).
Gibbs, H.L., K.A. Prior, P.W. Weatherhead and G. Johnson. 1997. Genetic structure
of populations of the threatened eastern massasauga rattlesnake, Sistrurus c.
catenatus: evidence from microsatellite DNA markers. Molecular Ecology
6:1123-1132.
Goater, C.P., R.D. Semlitsch and M.V. Bernasconi. 1993. Effects of body size and
parasite infection on the locomotory performance ofjuvenile toads, Bufo bufo.
Oikos 66:129-136.
Gomez-Mestre, I. and M. Tejedo. 2004. Contrasting patterns of quantitative and neutral
genetic variation in locally adapted populations of the natterjack toad, Bufo
calamita. Evolution 58:5343-5352.
Gray, J. 1946. The mechanism of locomotion in snakes. Journal of Experimental Biology
23: 101-1 19.
Gray, J. 1953. Undulatory propulsion. Quarterly Journal of Microscopical Science 94:55 1578.
Gregory, P.T. and K.W. Stewart. 1975. Long-distance dispersal and feeding strategy
of the red-sided garter snake (Thamnophis sirtalis parietalis) in the Interlake of
Manitoba. Canadian Journal of Zoology 53:238-245.
Gregory, P.T. 1977. Life-history parameters of the red-sided garter snake
(Thamnophis sirtalis parietalis) in an extreme environment, the Interlake
region of Manitoba. Publications de Zoologie, National Museum of Canada
13:1-44.
Guo, S.W. and E.A. Thompson. 1992. Performing the exact test for Hardy-Weinberg
proportions for multiple alleles. Biometrics 48:36 1-372.
Hamilton, M.B., E.L. Pincus, A. DiFiore and R.C. Fleischer. 1999. Universal linker
and ligation procedures for construction of genomic DNA libraries enriched
for micro satellites. Biotechniques 27:500-507.
Hakkarainen, H., P. Ilmonen and V. Koivunen. 2001. Experimental increase of
predation risk induces breeding dispersal of Tengmalm's owl. Oecologia
126:355-359.
Heg, D., Z. Bachar, L. Brouwer and M. Taborky. 2004. Predation risk is an
ecological constraint for helper dispersal in a cooperatively breeding cichlid.
Proceedings of the Royal Society of London Ser. B. 271:2367-2374.
Hanski, I. and D. Simberloff. 1997. The metapopulation approach, its hisotry,
conceptual domain, and application to conservation. In Metapopulation
Biology (Eds. l.A. Hanski and M.E. Gilpin), pp. 5-26. San Diego: Academic
Press.
152
Hitchings, S.P. and T.J.C. Beebee. 1997. Genetic substructuring as a result of barriers
to gene flow in urban Rana temporaria (common frog) populations:
implications for biodiversity conservation. Heredity 79:117-127.
Hoffman, E.A., W.R. Ardren and M.S. Blouin. 2003. Nine polymorphic
microsatellite loci for the northern leopard frog (Ranapipiens). Molecular
Ecology Notes 3:115-116.
Hoffman, E.A., F.W. Schueler and M.S. Blouin. 2004. Effective population sizes and
temporal stability of genetic structure in Rana pipiens, the northern leopard
frog. Evolution 58:2536-2545.
Hurvich, L.M. 1981. Color vision. Sunderland, Mass: Sinauer Associates, Inc.
Hutchison, D.W. and A.R. Templeton. 1999. Correlation of pairwise genetic and
geographic distance measures: inferring the relative influences of gene flow
and drift on the distribution of genetic variability. Evolution 53:1898-1914.
Jacobs, G.H. 1981. Comparative color vision. New York: Academic Press.
Jameson, D.L. 1956. Growth, dispersal and survival of the Pacific tree frog. Copeia
1956:25-29.
Jane, S.D. and J.K. Bowmaker. 1988. Tetrachromatic colour vision in the duck (Anas
platyrhynchos L.): microspectrophotometry of visual pigments and oil droplets.
Journal of Comparative Physiology 1 62A:225-235.
Janzen, F.J., J.G. Krenz, T.S. Haselkorn, E.D. Brodie, Jr. and E.D. Brodie, III. 2002.
Molecular phylogeography of common garter snakes (Thamnophis sirtalis) in
western North America: implications for regional historical forces. Molecular
Ecology 11:1739-1751.
Jayne, B.C. 1988. Muscular mechanisms of snake locomotion: an electromyographic
study of lateral undulation of the Florida Banded Water Snake (Nerodiafasciata)
and the Yellow Rat Snake (Elaphe obsoleta). Journal of Morphology 197:159-181.
John-Alder, H.B. and P.J. Morin. 1990. Effects of larval density on jumping ability
and stamina in the newly metamorphosed Bufo woodhousiifowleri. Copeia
1990:856-860.
Jones, A.G., S.J. Arnold and R. Btrger. 2004. Evolution and stability of the G-matrix on a
landscape with a moving optimum. Evolution 58:1639-1654.
Kelley, K.C., S.J. Arnold and J. Gladstone. 1997. The effects of substrate and vertebral
number on locomotion in the garter snake, Thamnophis elegans. Functional
Ecology 11:189-198.
Kephart, D.G. 1981. Population ecology and population structure of Thamnophis
elegans and Thamnophis sirtalis. Ph.D. dissertation, Dept. of Biology,
University of Chicago.
Kephart, D.G. 1982. Microgeographic variation in the diets of garter snakes.
Oecologia 52:287-291.
Kephart, D.G. and S.J. Arnold. 1982. Garter snake diets in a fluctuating environment:
a seven-year study. Ecology 63:1232-1236.
Keyghobadi, N., J. Roland and C. Strobeck. 1999. Influence of landscape on the
population genetic structure of the alpine butterfly Parnassius smintheus
(Papilionidae). Molecular Ecology 8:1481-1495.
153
King, R.B. 1993. Color-pattern variation in lake Erie water snakes: prediction and
measurement of natural selection. Evolution 47:18 19-1833.
Lande, R. 1992. Neutral theory of quantitative genetic variance in an island model with
local extinction and colonization. Evolution 46:38 1-389.
Levine, J.S. and E.F. MacNichol. 1979. Visual pigments in teleost fishes: effects of
habitat, microhabitat, and behavior on visual system evolution. Sensory Processes
3:95-13 1.
Lawson, R. and R.B. King. 1996. Gene flow and melanism in Lake Erie garter snake
populations. Biological Journal of the Linnean Society 59:1-19.
Legendre, P. 2001. Congruence among distance matrices: Program CADM user's
guide. Département de sciences biologiques, Université de Montréal. 7 pages.
Legendre, P., F.-J. Lapointe and P. Casgrain. 1994. Modeling brain evolution from
behavior: a permutational regression approach. Evolution 48:1487-1499.
Lurz, P.W.W., P.J. Garson and L.A. Wauters. 1997. Effects of temporal and spatial
variation in habitat quality on red squirrel dispersal behaviour. Animal
Behaviour 54:427-435.
Lythgoe, J.N. 1979. The ecology of vision. Oxford: Oxford University Press.
Macey, J.R., Schulte, J.A., II., N. B. Ananjeva, A. Larson, N. Rastegar-Pouyani, S.J.
Shammakov and T.J. Papenfuss. 1998. Phylogenetic relationships among
agamid lizards of the Laudakia caucasia species group: testing hypotheses of
biogeographic fragmentation and an area cladogram for the Iranian Plateau.
Molecular Phylogenetics and Evolution 10:118-13 1.
Manly, B.F. 1997. Randomization and Monte Carlo methods in biology. 2nd ed.
Chapman and Hall, New York.
Mantel, N. 1967. The detection of disease clustering and generalized regression
approach. Cancer Research 27:209-220.
Mantel, N. and R.S. Valand. 1970. A technique of nonparametric multivariate
analysis. Biometrics 26:547-558.
Matter, S.F. and J. Roland. 2002. An experimental examination of the effects of
habitat quality on the dispersal and local abundance of the butterfly Parnassius
smintheus. Ecological Entomology 27:308-316.
McCracken, G.F., G.M. Burghardt and S.E. Houts. 1999. Microsatellite markers and
multiple paternity in the garter snake Thamnophis elegans. Molecular Ecology
8: 1475-1479.
McKay, J.K. and R.G. Latta. 2002. Adaptive population divergence: markers, QTL and
traits. Trends in Ecology and Evolution 17:285-291.
McMillen-Jackson, A.L. and T.M. Bert. 2003. Disparate patterns of population
genetic structure and population history in two sympatric penaeid shrimp
species (Farfantepenaeus aztecus and Litopenaeus setferus) in the eastern
United States. Molecular Ecology 12:2895-2905.
Merilä, J. and P. Crnokrak. 2001. Comparison of genetic differentiation at marker loci and
quantitative traits. Journal of Evolutionary Biology 14:892-903.
Merrell, D.J. 1968. A comparison of the estimated size and the "effective size" of
breeding populations of the leopard frog, Ranapipiens. Evolution 22:274-283.
154
Michels, E., E. Audenaert, R. Ortells, L. Dc Meester. 2003. Population genetic
structure of three pond-inhabiting Daphnia species on a regional scale
(Flanders, Belgium). Freshwater Biology 48:1825-1839.
Moksnes, P.-O. 2004. Interference competition for space in nursery habitats:
density-dependent effects on growth and dispersal in juvenile shore crabs
Carcinus maenas. Marine Ecology Progress Series 281:181-191.
Molbo, D., C.A. Machado, E.A. Herre, L. Keller. 2004. Inbreeding and population
structure in two pairs of cryptic fig wasp species. Molecular Ecology 13:16131623.
Møller, A.P. and M.D. Jennions. 2002. How much variance can be explained by
ecologists and evolutionary biologists? Oecologia 132:492-500.
Monsen, K.J. and M.S. Blouin. 2003. Genetic structure in a montane ranid frog:
restricted gene flow and nuclear-mitochondrial discordance. Molecular
Ecology 12:3275-3286.
Monsen, K.J. and M.S. Blouin. 2004. Extreme isolation by distance in a montane
frog Rana cascadae. Conservation Genetics 5:827-835.
Moore, M. and J. Lindzey. 1992. The physiological basis of sexual behavior in male
reptiles. In C. Gans and D. Crews (eds.), Biology of the Reptilia, vol. 18, pp.
70-113. University of Chicago Press, Chicago, Illinois.
Munsell Color Company. 1976. Munsell Book of Color, Glossy Finish Collection (2
volumes). Baltimore: Munsell/Macbeth/Kolimorgen Corp.
Negro, J.J., F. Hiraldo and J.A. Doná.zar. 1997. Causes of natal dispersal in the lesser
kestrel: inbreeding avoidance or resource competition? Journal of Animal
Ecology 66:640-648.
Nevo, B., B. Baum, A. Beiles and D.A. Johnson. 1998. Ecological correlates of
RAPD DNA diversity of wild barley, Hordeum spontaneum, in the Fertile
Crescent. Genetic Resources and Crop Evolution 45:151-159.
Newman, R.A. and T. Squire. 2001. Microsatellite variation and fine-scale
population structure in the wood frog (Rana sylvatica). Molecular Ecology
10:1087-1100.
Nunney, L. and D.R. Elam. 1994. Estimating the effective population size of
conserved populations. Conservation Biology 8:175-184.
Ohta, T. and M. Kimura. 1973. A model of mutation appropriate to estimate the
number of electrophoretically detectable alleles in a finite population.
Genetical Research 22:201-204.
Palo, J.U., R.B. O'Hara, A.T. Laugen, A. Laurila, C.R. Primmer and J. Merilä. 2003.
Latitudinal divergence of common frog (Rana temporaria) life history traits by
natural selection: evidence from a comparison of molecular and quantitative
genetic data. Molecular Ecology 12:1963-1978.
Pampoulie, C., E.S. Gysels, G.E. Maes, B. Hellemans, V. Leentjes, A.G. Jones and
F.A.M. Volckaert. 2004. Evidence for fine-scale genetic structure and
estuarine colonisation in a potential high gene flow marine goby
(Pomatoschistus minutus). Heredity 92:434-445.
155
Peek, M.S., A.J. Leffler, S.D. Flint and R.J. Rye!. 2003. How much variance is
exp!ained by ecologists? Additional perspectives. Oecologia 137:161-170.
Pfrender, M.E., K. Sptize, J. Hicks, K. Morgan, L. Latta, M. Lynch.. 2000. Lack of
concordance between genetic diversity estimates at the molecular and
quantitative trait levels. Conservation Genetics 1:263-269.
Phillips, P.C. 1998. H2boot: bootstrap estimates and tests of quantitative genetic
data. Univ. of Texas at Arlington. Software available at
www.uta.edu/biology/phillips/software.
Porter, A.H. and H. Geiger. 1995. Limitations to the inference of gene flow at
regional geographic sca!es: an example from the Pieris napi group
(Lepidoptera: Pieridae) in Europe. Biological Journal of the Linnean Society
54:329-348.
Prior, K.A., H.L. Gibbs and P.J. Weatherhead. 1997. Population genetic structure in
the black rat snake: implications for management. Conservation Biology
11:1147-1158.
Prosser, M.R., H.L. Gibbs, P.J. Weatherhead. 1999. Microgeographic population
genetic structure in the northern water snake, Nerodia sipedon sipedon
detected using microsatellite DNA loci. Molecular Ecology 8:329-3 33.
Prout, T. and J.S.F. Barker. 1993. F statistics in Drosophila buzzatii: selection,
population size and inbreeding. Genetics 134:369-375.
Pullium, H.R. 1988. Sources, sinks, and population regulation. American Naturalist
132:653-661.
Raymond, M. and F. Rousset. 1995. GENEPOP Version 1.2: population genetics
software for exact tests and ecumenicism. Journal of Heredity 86:248-249.
Reed, D.H. and R. Frankham. 2001. How closely correlated are molecular and
quantitative measures of genetic variation? a meta-analysis. Evolution
55:1095-1103.
Reh, W. and A. Seitz. The influence of land use on the genetic structure of
populations of the common frog Rana temporaria. Biological Conservation
54:239-249.
Rice, W.R. 1989. Analysing tables of statistical tests. Evolution 43:223-225.
Roach, J.L., P. Stapp, B. Van Home and M.F. Antolin. 2001. Genetic structure of a
metapopulation of black-tailed prairie dogs. Journal of Mammalogy 82:946959.
Rossman, D.A., N.B. Ford and R.A. Seigel. 1996. The garter snakes: evolution and
ecology. Norman, Oklahoma: University of Oklahoma Press.
Rousset, F. 1997. Genetic differentiation and estimation of gene flow from FStatistics under isolation by distance. Genetics 145:1219-1228.
Rowe, G., T.J.C. Beebee and T. Burke. 2000. A microsatellite analysis of natterjack
toad, Bufo calamita metapopulations. Oikos 88:641-651.
Rtiber, L., A. Meyer, C. Sturmbauer and E. Verheyen. 2001. Population structure in
two sympatric species of the Lake Tanganyika cichlid tribe Eretmodini:
evidence for introgression. Molecular Ecology 10:1207-1225.
156
Rychlik, W. 1998. Oligo Primer Analysis Software v. 6 for Mac. Molecular Biology
Insights, Inc. Cascade, Co.
SAS Institute. 2002. SAS/STAT software user's guide. Rel. 9.1. SAS Institute, Inc.,
Cary, NC.
Schneider, S., D. Roessli and L. Excoffier. 2000. Arlequin ver 2.000: a software for
population genetics data analysis. Genetics and Biometry Laboratory,
University of Geneva, Switzerland.
Schweiger, 0., M. Frenzel and W. Durka. 2004. Spatial genetic structure in a
metapopulation of the land snail Cepaea nemoralis (Gastropoda: Helicidae).
Molecular Ecology 13:3645-3655.
Scribner, K.T., J.W. Arntzen, N. Cruddace, R.S. Oldham and T. Burke. 2001.
Environmental correlates of toad abundance and population genetic diversity.
Biological Conservation 98:201-210.
Scribner, K.T. and R.K. Chesser. 1993. Environmental and demographic correlates of
spatial and seasonal genetic structure in the eastern cottontail (Sylvilagus
floridanus). Journal of Mammalogy 74:1026-1044.
Seppa, P. and A. Laurila. 1999. Genetic structure of island populations of the anurans
Rana temporaria and Bufo bufo. Heredity 82:309-3 17.
Sinsch, U. 1997. Postmetamorphic dispersal and recruitment of first breeders in a
Bufo calamita metapopulation. Oecologia 112:42-47.
Simandle, E. 2005. Phylogeography and metapopulation structure of the Bufo boreas
group in the Great Basin. Ph.D. dissertation; Ecology, Evolution and
Conservation Biology Program; University of Nevada, Reno.
Slatkin, M. and L. Excoffier. 1996. Testing for linkage disequilibrium in genotypic
data using the EM algorithm. Heredit1 76:377-383.
Sokal, R. and F.J. Rohlf. 1995. Biometry, 3' ed. New York, New York: W.H.
Freeman.
Spitze, K. 1993. Population structure in Daphnia obtusa: quantitative genetic and
allozyme variation. Genetics 135 :367-374.
Stacey, P.B., V.A. Johnson and M.L. Taper. 1997. Migration within
metapopulations: the impact upon local population dynamics. In
Metapopulation Biology (Eds. l.A. Hanski and M.E. Gilpin), pp. 267-292. San
Diego: Academic Press.
Steams, S.C. 1992. The Evolution of Life Histories. Oxford: Oxford University
Press.
Stebbins, R.C. 2003. Western amphibians and reptiles. Houghton Mifflin Co., New
York.
Van Rossum, F., I. Bonnin, S. Fénart, M. Pauwels, D. Petit and P. Saumitou-Laprade.
2004. Spatial genetic structure within a metallicolous population of
Arabidopsis halleri, a clonal, self-incompatible and heavy-metal-tolerant
species. Molecular Ecology 13:2959-2967.
Vekemans, X. and O.J. Hardy. 2004. New insights from fine-scale spatial genetic
structure analyses in plant populations. Molecular Ecology 13:921-935.
157
Voris, H.K. 1975. Dermal scale-vertebra relationships in sea snakes (Hydrophidae).
Copeia 1975:746-755.
Waldmann, P. and S. Andersson. 1998. Comparison of quantitative genetic variation and
allozyme diversity within and between populations of Scabiosa canescens and S.
columbaria. Heredity 81:79-86.
Waples, R.S. 1989. A generalized approach for estimating effective population size
from temporal changes in allele frequency. Genetics 121:379-391.
Weber, J.L. and C. Wong. 1993. Mutation of human short tandem repeats. Human
Molecular Genetics 2:1123-1128.
Whiteley, A.R., P. Spruell and F.W. Allendorf. 2004. Ecological and life history
characteristics predict population genetic divergence of two salmonids in the
same landscape. Molecular Ecology 13:3675-3688.
Whittier, J.M. and R.R. Tokarz. 1992. Physiological regulation of sexual behavior in
female reptiles. In C. Gans and D. Crews (eds.), Biology of the Reptilia, vol.
18, pp. 24-69. University of Chicago Press, Chicago, Illinois.
Wong, A., M.L. Smith and M.R. Forbes. 2003. Differentiation between subpopulations of
a polychromatic daniseifly with respect to morph frequencies, but not neutral
genetic markers. Molecular Ecology 12:3505-3513.
Wright, 5. 1922. Coefficients of inbreeding and relationship. American Naturalist 63:556561.
Wright, S. 1931. Evolution in Mendelian populations. Genetics 16:97-159.
Wright, S. 1943a.. Isolation by distance. Genetics 28:114-138.
Wright, S. 1943b. An analysis of local variability of flower color inLinanthuspariyae.
Genetics 28:139-156.
Wright, S. 1951. The genetical structure of populations. Annals of Eugenics 15:323-354.
Wright, S. 1969. The theory of gene frequencies, evolution and the genetics of populations.
Vol.2, Chicago: University of Chicago Press.
Yang, R.-C., F.C. Yeh and A.D. Yanchuk. 1996. A comparison of isozyme and
quantitative genetic variation in Pinus contorta ssp. latifolia by Fs1. Genetics
142:1045-1052.
Zardoya, R., R. Castilho, C. Grande, L. Favre-Krey, S. Caetano, S. Marcato, G. Krey
and T. Patamello. 2004. Differential population structuring of two closely
related fish species, the mackerel (Scomber scombrus) and the chub mackerel
(Scomberjaponicus), in the Mediterranean Sea. Molecular Ecology 13:17851798.