Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Heart failure wikipedia , lookup
Cardiac contractility modulation wikipedia , lookup
Coronary artery disease wikipedia , lookup
Electrocardiography wikipedia , lookup
Cardiothoracic surgery wikipedia , lookup
Arrhythmogenic right ventricular dysplasia wikipedia , lookup
Cardiac surgery wikipedia , lookup
Hypertrophic cardiomyopathy wikipedia , lookup
Myocardial infarction wikipedia , lookup
Cardiac arrest wikipedia , lookup
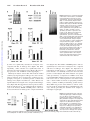
Upregulation of Heat Shock Transcription Factor 1 Plays a Critical Role in Adaptive Cardiac Hypertrophy Masaya Sakamoto,* Tohru Minamino, Haruhiro Toko,* Yosuke Kayama,* Yunzeng Zou, Masanori Sano, Eiichi Takaki, Teruhiko Aoyagi, Katsuyoshi Tojo, Naoko Tajima, Akira Nakai, Hiroyuki Aburatani, Issei Komuro Downloaded from http://circres.ahajournals.org/ by guest on April 29, 2017 Abstract—Exercise-induced cardiac hypertrophy has been reported to have better prognosis than pressure overloadinduced cardiac hypertrophy. Cardiac hypertrophy induced by exercise was associated with less cardiac fibrosis and better systolic function, suggesting that the adaptive mechanisms may exist in exercise-induced hypertrophy. Here, we showed a critical role of heat shock transcription factor 1 (HSF1), an important transcription factor for heat shock proteins, in the adaptive mechanism of cardiac hypertrophy. We examined expression of 8800 genes in the heart of exercise-induced hypertrophy model using DNA chip technique and compared with pressure overload–induced hypertrophy. Expression of HSF1 and its target molecule heat shock proteins was significantly upregulated in the heart by exercise but not by chronic pressure overload. Constitutive activation of HSF1 in the heart significantly ameliorated death of cardiomyocytes and cardiac fibrosis and thereby prevented cardiac dysfunction as well as hypertrophy induced by chronic pressure overload. Conversely, decreased activity of HSF1 in the heart promoted cardiac dysfunction in response to exercise, a load that normally leads to adaptive hypertrophy with preserved systolic function. Likewise, cardiac function was significantly impaired from the early phase of pressure overload, when HSF1 activation was inhibited. These results suggest that HSF1 plays a critical role in the transition between adaptive and maladaptive hypertrophy. (Circ Res. 2006;99:1411-1418.) Key Words: pressure overload 䡲 exercise 䡲 heart failure C ardiac hypertrophy is an adaptive response to increased wall stress. At the beginning, cardiac hypertrophy has beneficial effects to maintain cardiac output by reducing wall stress; however, long-term stresses induce systolic dysfunction, leading to heart failure.1 Clinical studies have demonstrated that cardiac hypertrophy is not only a cause of congestive heart failure but also an independent risk factor for myocardial infarction, arrhythmia, and sudden death.2 Therefore, it is very important to elucidate the as yet uncharacterized mechanisms underlying the transition from cardiac hypertrophy to heart failure and to develop potential therapeutic strategies for this condition. A metaanalysis of the hearts of athletes has demonstrated that the endurance-trained heart shows an increase in both left ventricular (LV) diameter and LV wall thickness without impairment of systolic and diastolic function.3 Because exercise-induced cardiac hypertrophy, the so called “sports heart,” has been reported not to progress to heart failure,4,5 it is thought to be “adaptive hypertrophy.” On the other hand, pressure overload initially induces adaptive hypertrophy, but it leads to “maladaptive hypertrophy” in the chronic phase, resulting in heart failure.6 Although there have been reports demonstrating different expression patterns of some genes between these 2 forms of cardiac hypertrophy,7,8 the precise mechanism of different prognosis remains unclear. In the present study, we examined expression of 8800 genes in the hearts of 2 hypertrophy rat models using DNA chip. In the DNA chip analysis, we found that the heat shock protein (Hsp) genes such as Hsp70 and Hsp27 were markedly upregulated in the heart of the exercise model but not in that of the pressure-overload model. Heat shock transcription factor 1 (HSF1), which regulates Hsps gene expression,9 –11 was activated only in the heart of the exercise model. Overexpression of activated HSF1 in the heart prevented cardiomyocyte death and cardiac fibrosis in response to sustained pressure overload and thereby preserved cardiac Original received March 31, 2006; resubmission received September 19, 2006; revised resubmission received October 28, 2006; accepted October 31, 2006. From the Department of Cardiovascular Science and Medicine (M. Sakamoto, T.M., H.T., Y.K., Y.Z., M. Sano, I.K.), Chiba University Graduate School of Medicine; Department of Biochemistry and Molecular Biology (E.T., A.N.), Yamaguchi University Graduate School of Medicine, Ube; Japan Red Cross Medical Center (T.A.), Tokyo; Division of Diabetes, Metabolism and Endocrinology (M. Sakamoto, K.T., N.T.), Department of Internal Medicine, Jikei University School of Medicine, Tokyo; and Genome Science Division (H.A.), Research Center for Advanced Science and Technology, University of Tokyo, Japan. *These authors contributed equally to this work. Correspondence to Issei Komuro, MD, PhD, Department of Cardiovascular Science and Medicine, Chiba University Graduate School of Medicine, 1-8-1 Inohana, Chuo-ku, Chiba 260-8670, Japan. E-mail [email protected] © 2006 American Heart Association, Inc. Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000252345.80198.97 1411 1412 Circulation Research December 8/22, 2006 function. Conversely, decreased expression of HSF1 in the heart impaired the adaptive response to exercise or acute pressure overload and thus caused cardiac dysfunction, suggesting a protective role of HSF1 in cardiac physiology. Materials and Methods Rat Models Downloaded from http://circres.ahajournals.org/ by guest on April 29, 2017 All protocols were approved by the Institutional Animal Care and Use Committee of Chiba University. Male Wistar rats obtained from Nippon Bio-Supply Center (Tokyo, Japan) were divided into 3 groups: sham-operated model, exercise-induced hypertrophy model, and pressure overload–induced hypertrophy model (n⫽8, each group). Five-week-old male rats were individually housed and voluntary exercised in a specially manufactured cage equipped with a controlled running wheel and a distance counter as described previously.12 In this cage, there were 2 rooms (a room for wheel running and another for rest), and therefore rats could run or rest ad libitum. Pressure overload was produced by constriction of abdominal aorta as described previously.13 Briefly, 8-week-old male rats were anesthetized with an intraperitoneal injection of a cocktail of ketamine HCl (100 mg/kg) and xylazine (5 mg/kg). The abdominal aorta was constricted above the right renal artery by 4-0 silk suture tied around both the aorta and a blunted 22-gauge needle, which was then pulled out. lary profiles and nuclei (n⫽100, each group).18 To determine the degree of collagen fiber accumulation, we selected 5 fields at random and calculated the ratio of Masson-stained fibrosis area to total myocardium area with the software “NIH Image” (Bethesda, Md) for image analysis as described previously.19 Cardiomyocyte death was detected in situ by terminal deoxyribonucleotide transferase–mediated dUTP nick-end labeling (TUNEL) method using Cardio TACS (Trevigen Inc, Gaithersburg, Md), in paraffin-embedded heart tissue sections. Ribonuclease Protection Assay and Northern and Western Blot Analysis Total RNA (20 g) was separated on a 1.0% agarose/formaldehyde gel and was hybridized with the cDNA fragments of Hsp70, Hsp27, and HSF1 genes. Ribonuclease protection assay (RiboQuant, Pharmingen, Franklin Lakes, NJ) was performed according to the instructions of the manufacturer. Western blotting was performed as previously described.15 Antibody for inducible Hsp70 was purchased from Stressgen Biotechnologies (Victoria, Canada) and antibody for HSF1 was from Santa Cruz Biotechnology (Santa Cruz, Calif). Gel Mobility-Shift Assay DNA-binding activity of HSF1 was examined using a selfcomplementary oligonucleotide containing the heat shock element (5⬘-CTAGAAGCTTCTAGAAGCTTCTAG-3⬘) as a probe.15 DNA Chip Analysis ECG Analysis Total RNA (5 g) was extracted from LV of each model by the lithium/urea method and was used to synthesize biotin-labeled cRNA, which was then hybridized to high-density oligonucleotide array (GeneChip U34A array; Affymetrix, Santa Clara, Calif) according to the previously published protocol.14 Arrays contain probe sets for approximately 8800 genes and expressed-sequence tags, which were selected from Build no. 34 of the UniGene Database (created from GenBank 107/dbEST, November 18, 1998). The GeneChip 3.3 software (Affymetrix) was used to calculate the average difference for each probe on the array, which was shown as an intensity value of gene expression defined by Affymetrix using their algorithm. The average difference has been shown to quantitatively reflect the abundance of particular mRNA molecule in a population.14 Transthoracic echocardiograph was performed with the HP Sonos 4500 (Hewlett-Packard Co, Palo Alto, CA) with a 10-MHz imaging transducer, and the progress of cardiac hypertrophy was evaluated as described previously.20 HSF1 Transgenic Mice Construction of active form of human HSF1 transgene and generation of the hHSF1 transgenic mice have been previously described.15 Eight-week-old male transgenic and their wild-type littermates were used. The transgenic mice were apparently healthy, and there were no significant differences in the body weight (BW), heart weight to BW ratio (HW/BW), or hemodynamic parameters, such as blood pressure (BP) and heart rate (HR), between the transgenic mice and wild-type littermates. Strong pressure overload was imposed on the heart for 5 weeks by surgical constriction of transverse aorta (TAC) as described previously.16 HSF1-Deficient Mice The generation of HFS-1– deficient mice has been described previously.17 The HFS-1– deficient heterozygote mice were apparently healthy. There were no significant differences in echocardiogram parameters such as posterior wall thickness (PWTd) and fractional shortening between the mutant mice and wild-type littermates. The mice underwent 4 weeks of exercise (wheel running) or 1 week of TAC as described above. Histological Analysis For histological analysis, hearts were fixed by perfusion with 10% formalin. Fixed hearts were embedded in paraffin and sectioned at 4-m thickness. The myocyte cross-sectional diameter was measured in the sections stained with hematoxylin and eosin, and suitable cross-sections were defined as having nearly circular capil- Hemodynamic Measurements In Vivo To measure hemodynamic effects of aortic constriction and exercise, the right carotid artery was cannulated with a polyethylene catheter (MILLAR, Houston, Tex). The transducer (model MP 5100, Baxter, Deerfield, Ill) was connected to Mac Laboratory system (model MacLab4/s, AD Instruments, Castle Hill, Australia), and BP, HR, and LV end-diastolic pressure were measured. Statistical Analysis Data were shown as mean⫾SEM. Multiple group comparison was performed by 1-way ANOVA, followed by the Bonferroni procedure for comparison of means. Comparison between 2 groups were analyzed by the 2-tailed Student’s t test or 2-way ANOVA. Values of P⬍0.05 were considered statistically significant. Results Physiological Analysis After 8 weeks of voluntary exercise or 5 weeks of pressure overload, physiological analyses were performed in 13-weekold rats. Echocardiogram revealed that PWTd and LV enddiastolic dimension were larger in both groups compared with those in the sham-operated group (Table I in the online data supplement, available at http://circres.ahajournals.org). Sustained pressure overload developed cardiac hypertrophy more profoundly than exercise (supplemental Table I). Fractional shortening was decreased in the pressure-overload group but not in the exercise group (supplemental Table I). In hemodynamic parameters, BP measured in the right carotid artery and LV end-diastolic pressure were much higher in the pressureoverload group than the sham-operated group (supplemental Table II). On the other hand, BP was significantly lower and HR was less in the exercise model compared with the pressure-overload model and the sham-operated model (sup- Sakamoto et al HSF1 in Cardiac Hypertrophy 1413 Figure 1. Histological analysis of rat models. A through D, High-magnification views of hematoxylin and eosin–stained LV of sham-operated rats (A), pressureoverloaded rats (B), and exercised rats (C). Scale bar⫽10 m. D, Both pressure overload (PO) and exercise (EX) induced an increase in diameter of cardiomyocytes. *P⬍0.01 vs sham (n⫽8). E through G, High-magnification views of Massonstained LV of sham-operated rats (E), pressure-overloaded rats (F), and exercised rats (G). H, An increase in perivascular and interstitial fibrosis was observed only in the pressure-overload model. *P⬍0.01 vs sham (n⫽8). Downloaded from http://circres.ahajournals.org/ by guest on April 29, 2017 plemental Table II). In both cardiac hypertrophy models, LV weight to BW ratio was increased (supplemental Table II). The increase was more prominent in the pressure-overload model than in the exercise model. Histological analysis demonstrated that myocyte diameter was larger in both hypertrophy models compared with the sham-operated model (Figure 1A through 1D). In contrast, Masson staining revealed that cardiac fibrosis was significantly increased in only the pressure-overload model but not in the exercise model (Figure 1E through 1H). These results indicate that chronic pressure overload but not exercise induces cardiac fibrosis, thereby promoting maladaptive cardiac hypertrophy. Expression Level of Hsp70 Protein and Activity of HSF1 Were Elevated in Exercise-Induced Hypertrophy To determine the molecular difference between exercise- and chronic pressure overload–induced hypertrophy, we performed DNA chip analysis. Expression levels of ⬇100 genes were differently elevated in each model compared with the sham-operated model. mRNA levels of fetal genes such as atrial natriuretic peptide, brain natriuretic peptide, and skeletal ␣-actin were elevated ⬇2- to 3-fold in the pressureoverload model but not in the exercise model (Table 1). It is important to note that expression level of the cardioprotective TABLE 1. DNA Chip Analysis Sham Pressure Overload Exercise ANP 807 2992 710 BNP 776 1552 656 Natriuretic factor Organism defense Hsp70 35 29 104 Hsp27 934 1315 2150 Muscle protein Smooth muscle MHC Skeletal ␣-actin 21 69 40 1985 4640 1393 Genes increased in rats with pathological (pressure overload) and physiological (exercise) cardiac hypertrophy compared with a sham-operated rat (Sham). ANP indicates atrial natriuretic peptide; BNP, brain natriuretic peptide; MHC, myosin heavy chain. gene Hsp70 and Hsp27 was upregulated ⬇2- to 3-fold in the exercise model but not in the pressure-overload model (Table 1). We performed Northern blot analysis and Western blot analysis to confirm the results of the DNA chip analysis. Northern blot analysis revealed that mRNA levels of Hsp70 and Hsp27 were significantly increased in only the exercise model (Figure 2A and data not shown). HSF1, a key molecule regulating gene expression of Hsps, was also upregulated only in the heart of the exercise model (Figure 2A). Protein levels of inducible form of Hsp70 and HSF1 were significantly higher in the exercise model than those in the pressureoverload model and the sham-operated model (Figure 2B). We next examined the activity of HSF1 by gel mobilityshift assay using a HSF1-binding DNA sequence. Shifted band was more obvious in cardiac extracts prepared from the exercise model than in those from the pressureoverload model and the sham-operated model (Figure 2C), suggesting that HSF1 is activated in exercise-induced hypertrophy. Protective Role of HSF1 in Sustained Pressure Overload–Induced Hypertrophy To elucidate the role of HSF1 in cardiac hypertrophy, we established the cardiac hypertrophy model by TAC in transgenic mice that express constitutively active HSF115 and analyzed them 5 weeks after the operation. We also performed the same operation in their wild-type littermates as controls. Expression of activated HSF1 was detected in the heart of transgenic mice but not in wild-type mice (Figure 3A). Inducible Hsp70 expression was markedly increased in the heart of transgenic mice compared with wild-type mice (Figure 3A). Pressure studies revealed that there was no difference in BP and HR between transgenic and wild-type mice before the operation (data not shown). After the operation, BP was significantly increased in both transgenic mice and wild-type mice to the same extent (Table 2). LV end-diastolic pressure was significantly elevated in wild-type mice, which was significantly ameliorated in transgenic mice (Table 2). HW/BW was significantly increased in wild-type mice, but the increases in HW/BW were significantly less in transgenic mice than in wild-type mice (Table 2). Echocardiographic analysis demonstrated that an increase in PWTd 1414 Circulation Research December 8/22, 2006 Downloaded from http://circres.ahajournals.org/ by guest on April 29, 2017 Figure 2. Expression of Hsp70 and HSF1 in the heart of rat models. A, Total RNA was prepared from the heart of shamoperated rats, pressure-overloaded rats (PO), or exercised rats (EX) and examined for Hsp70 and HSF1 expression by Northern blot analysis. Ethidium bromide staining of 18S was shown as a loading control. Expression levels relative to sham-operated rats were plotted in the graph. *P⬍0.05, **P⬍0.01 vs sham; †P⬍0.05, ††P⬍0.01 vs pressure overload (n⫽4). B, Whole-cell lysates were prepared from the heart of shamoperated rats, pressure overloaded rats (PO) or exercised rats (EX) and examined for inducible Hsp70 and HSF1 expression by Western blot analysis. Expression levels relative to sham-operated rats were plotted in the graph. *P⬍0.05 vs sham; †P⬍0.05 vs PO (n⫽4). C, Nuclear extracts from the hearts were subjected to the gel mobility shift assay. Heat shock response element was used as a DNA probe. More prominent shifted band was observed in the exercise model than the pressure-overload and sham-operated models. Same experiments were performed at least 3 times and similar results were obtained. by TAC was significantly attenuated in transgenic mice compared with that in wild-type mice (Figure 3B). More importantly, cardiac function was significantly better in transgenic mice than in wild-type mice (Figure 3B), suggesting a protective role of HSF1 in cardiac hypertrophy. Histological analysis showed that TAC induced marked cardiomyocyte hypertrophy in wild-type mice after 5 weeks, whereas transgenic mice developed less cardiomyocyte hypertrophy in response to sustained pressure overload than wild-type mice (Figure 4A). Moreover, cardiac fibrosis was significantly less in transgenic mice compared with wild-type mice (Figure 4B). Because TAC has been reported to induce death of cardiomyocytes,21 we examined death of cardiomyocytes in the heart by TUNEL. TUNEL-positive cells were increased in the heart of wild-type mice compared with sham mice (Figure 4C). The number of TUNEL-positive cells was significantly less in the heart of transgenic mice compared with that of wild-type mice (Figure 4C). Expression of transforming growth factor-, a cytokine that is crucial for tissue fibrosis,22 was markedly increased in the heart after pressure overload (Figure 4D). This induction was significantly prevented by overexpression of HSF1. It has been reported that HSF1 and Hsps suppress activation of transcription factors such as activator protein-1 and nuclear factor B,23–25 both of which are known to upregulate proinflammatory cytokines including transforming growth factor-. Thus, HSF1 may prevent cardiac fibrosis as well as cardiomyocyte death by antagonizing these proinflammatory pathways, thereby preventing the transition from adaptive to maladaptive hypertrophy. Figure 3. Sustained pressure overload– induced hypertrophy in HSF1 transgenic mice. A, Whole-cell lysates were prepared from the hearts of wild-type mice (WT) and HSF1 transgenic mice (TG) and examined for HSF1 and inducible Hsp70 expression by Western blot analysis. Arrow indicates a constitutively active form of HSF1. B, Pressure overload was imposed on wild-type mice (WT-TAC) and HSF1 transgenic mice (TG-TAC) by 5 weeks of TAC. Sham operation was also imposed on wild-type mice (Sham). Cardiac hypertrophy (PWTd) and systolic function (percentage of fractional shortening [%FS]) were examined 5 weeks after TAC by echocardiogram. *P⬍0.05 vs sham, †P⬍0.05 vs WT-TAC (n⫽5). Sakamoto et al HSF1 in Cardiac Hypertrophy 1415 HSF1 Deficiency Impairs the Adaptive Response to Exercise or Pressure Overload hypertrophy, we produced the TAC model in HSF1⫹/⫺ mice and their wild-type littermates and analyzed cardiac function 1 week later. In this adaptive phase, TAC increased expression of inducible Hsp70 in the heart of wild-type mice (Figure 6A). This induction was markedly attenuated in the heart of HSF1⫹/⫺ mice (Figure 6A). Consistent with the results of our rat model, expression of inducible Hsp70 was downregulated in the maladaptive phase (supplemental Figure). Both HSF1⫹/⫺ mice and wild-type mice developed cardiac hypertrophy to the degree similar to that of 4 weeks of exercise (Figures 5C, 5D, 6B, and 6C). Cardiac function was preserved in wild-type mice, but it was significantly impaired in HSF1⫹/⫺ mice (Figure 6C), further supporting a notion that HSF1 has an important role in adaptive hypertrophy. We next determined whether HSF1 has a protective role in exercise-induced hypertrophy. We produced the exerciseinduced cardiac hypertrophy model in HSF1-deficient heterozygote (HSF1⫹/⫺) mice17 and compared them with their wild-type littermates. Western blot analysis demonstrated that expression of HSF1 was less in the heart of HSF1⫹/⫺ mice than wild-type mice (Figure 5A). Expression of inducible Hsp70 was increased in the heart of wild-type mice by exercise, but this increase was attenuated in the heart of HSF1⫹/⫺ mice (Figure 5B). There were no differences in HW/BW and wall thickness (PWTd) between wild-type mice and HSF1⫹/⫺ mice before exercise (Figure 5C and 5D). After 4 weeks of exercise, both mice developed cardiac hypertrophy to a similar extent (Figure 5C and 5D). In contrast to the preserved function of wild-type mice, cardiac function of HSF1⫹/⫺ mice was significantly reduced after exercise (Figure 5D), suggesting a critical role of HSF1 in the adaptive mechanism of cardiac hypertrophy induced by exercise. We noted that pressure overload by TAC induced adaptive hypertrophy until 1 to 2 weeks after operation to preserve cardiac function; however, this adaptive mechanism could not protect the hypertrophied heart against sustained pressure overload, resulting in systolic dysfunction at 4 to 5 weeks (Figure 3B). To further test the role of HSF1 in adaptive In the present study, we performed DNA chip analysis to elucidate the molecular difference between exercise-induced and sustained pressure overload–induced cardiac hypertrophy. We found that ⬇100 genes were differently elevated in each hypertrophy model compared with the sham-operated model. Among them, expression levels of Hsp70 and Hsp27 were elevated in only the heart of the exercise model, which was confirmed by Northern and Western blot analysis. Hsps such as Hsp70 and Hsp27 are ubiquitously expressed and its expression is enhanced by various stresses such as heat shock,26 reactive oxygen species,26 heavy metals,27 and inflammation.28 It has been reported that stimuli that result in the transient induction of Hsps in the heart protect, to some degree, against detrimental effects of ischemia/reperfusion injury.29,30 Constitutive expression of Hsps in transgenic mice efficiently recovers cardiac function after ischemia and reperfusion,31–33 whereas absence of Hsps leads to cardiac dysfunction and impairs stress response.34 Several studies have reported that expression of Hsps is rapidly induced in the heart in response to acute pressure overload, as well as volume overload.35 However, there has been no report concerning expression of Hsps in the chronic stage of cardiac TABLE 2. Hemodynamic Parameters and Heart Weight Sham WT-TAC 102.3⫾7.3 153.0⫾5.6† 145.5⫾6.3† 622⫾32 602⫾15 588⫾18 LVEDP, mm Hg 7.8⫾1.8 16.6⫾2.2† 12.0⫾0.8†‡ HW, mg/BW (g) 4.72⫾0.16 6.74⫾0.29† 5.79⫾0.40*‡ BP, mm Hg HR, bpm TG-TAC Sham indicates sham-operated mice; WT-TAC and TG-TAC, wild-type and transgenic mice, respectively, receiving pressure overload by 5 weeks of TAC; LVEDP, LV end-diastolic pressure. *P⬍0.05, †P⬍0.01 vs Sham; ‡P⬍0.05 vs WT-TAC (n⫽7). Discussion Downloaded from http://circres.ahajournals.org/ by guest on April 29, 2017 Figure 4. Histological analysis of the heart after 5 weeks of TAC. A through C, Pressure overload was imposed on wildtype mice (WT-TAC) and HSF1 transgenic mice (TG-TAC) by 5 weeks of TAC. Sham operation was also imposed on wild-type mice (Sham). The transverse diameter of cardiomyocytes (A), the extent of perivascular and interstitial fibrosis (B), and the number of TUNELpositive cells/10 000 cardiomyocytes (C) were estimated as described in Materials and Methods. *P⬍0.05 vs sham, †P⬍0.05 vs WT-TAC (n⫽5). D, Total RNA was extracted from the hearts of sham, WT-TAC, or TG-TAC and examined for expression of transforming growth factor (TGF)- by ribonuclease protection assay. Expression levels relative to sham-operated mice were plotted in the graph. *P⬍0.05 vs sham, †P⬍0.05 vs WT-TAC (n⫽4). 1416 Circulation Research December 8/22, 2006 Figure 5. Exercise-induced hypertrophy in HSF1-deficient mice. A, Western blot analysis for HSF1 in the heart of HSF1deficient heterozygote mice (KO) and their wild-type littermates (WT). B, HSF1deficient heterozygote mice (KO-EX) and their littermate wild-type mice (WT-EX) were exercised for 4 weeks. Sham operation was also imposed on both types of mice (KO-Sham and WT-Sham). Wholecell lysates were then prepared from the hearts and examined for inducible Hsp70 expression by Western blot analysis. C, HW/BW was measured before and after exercise. *P⬍0.05 vs before exercise (n⫽3– 4). D, Hypertrophy (PWTd) and cardiac function (percentage of fractional shortening [%FS]) were examined before and after exercise by echocardiogram. *P⬍0.01 vs before exercise, †P⬍0.01 vs wild-type mice after exercise (n⫽4). Downloaded from http://circres.ahajournals.org/ by guest on April 29, 2017 hypertrophy. In this study, Hsp70 and Hsp27 were upregulated in the heart after exercise but not in the heart after long-term pressure overload. These results suggest that exercise persistently upregulates expression of Hsps, thereby inhibiting cardiac dysfunction associated with hypertrophy, whereas pressure overload transiently induces expression of Hsps and thus fails to prevent impaired function in the chronic phase. The transcription of Hsps genes is mainly controlled by HSF1.9 –11 Impaired activation of HSF1 in the aged heart results in diminished induction of Hsps by stressful stimuli such as heat shock or ischemia. This impairment is associated with reduced protective effects of mild heat shock or ischemia against subsequent severe ischemic stresses in the aged heart.36 HSF1 deficiency has been reported to decrease cardiac expression of Hsps,37 further supporting a critical role of HSF1 in the regulation of Hsps expression. Although pressure overload and ischemia/reperfusion injury have been reported to rapidly induce HSF1 activation and Hsps expression in the heart,30,38 we found that HSF1 was activated in only the heart of the exercise-induced hypertrophy model but not in pressure overload–induced hypertrophy model at the chronic stage, suggesting that exercise promotes sustained activation of HSF1, which results in constitutive expression of Hsps in the heart. It remains unclear how HSF1 is downregulated under sustained pressure overload. Extracellular signal-regulated protein kinase (ERK) is known to increase HSF1 activity.39 It is also reported that ERK is activated by pressure overload and that its activation is involved in the adaptive mechanism of hypertrophy.40 Moreover, we observed that ERK was activated in exerciseinduced hypertrophy but not in the chronic phase of pressure overload–induced hypertrophy (M. Sakamoto and I. Komuro, unpublished data, 2005), suggesting that the ERK signaling pathway may participate in the activation of HSF1 in exercise-induced hypertrophy and that its downregulation may cause the maladaptive response to chronic pressure overload by inactivating HSF1. Constitutive activation of HSF1 prevented cardiac dysfunction as well as hypertrophy under chronic pressure overload. Decreased activity of HSF1 did not exacerbate hypertrophy but impaired systolic function in response to exercise or acute pressure overload, a load that usually results in adaptive hypertrophy. Thus, HSF1 activation may play an essential role in inhibiting the transition from adaptive to maladaptive hypertrophy rather than in regulating the size of cardiomyocytes. The different manifestations of hypertrophy between the exercise model and the long-term pressureoverload model might be attributable to the differences in the continuity and degree of stimulus. In a recent report, Perrino Figure 6. Acute pressure overload–induced hypertrophy in HSF1-deficient mice. A, Pressure overload was imposed on wild-type mice (WT-TAC) and HSF1-deficient mice (KO-TAC) by 1 week of TAC. Sham operation was also imposed on wild-type mice (Sham). Whole-cell lysates were prepared from the hearts of wild-type mice and HSF1-deficient mice and examined for inducible Hsp70 expression by Western blot analysis. B, HW/BW was measured 1 week after pressure overload. *P⬍0.01 vs sham (n⫽5–7). C, Hypertrophy (PWTd) and cardiac function (percentage of fractional shortening [%FS]) were examined 1 week after pressure overload by echocardiogram. *P⬍0.01 vs sham, †P⬍0.05 vs WT-TAC (n⫽4 to 7). Sakamoto et al Downloaded from http://circres.ahajournals.org/ by guest on April 29, 2017 et al41 applied intermittent pressure overload to the hearts of mice and tested the roles of duration and nature of the stress on the development of cardiac failure. Despite a mild hypertrophic response, hearts exposed to intermittent pressure overload displayed various pathological features including diastolic dysfunction. Thus, the nature of the stress on the heart, rather than its duration, is the key determinant of the maladaptive phenotype. To test the role of the degree of stimulus, we compared the effect of 1 week of TAC on cardiac function with that of 4 weeks of exercise for the following reasons. First, pressure overload by TAC induces adaptive hypertrophy until 1 to 2 weeks after operation to preserve cardiac function. Second, 4 weeks of exercise did not induce cardiac dysfunction in wild-type mice. Third, 4 weeks of exercise induces cardiac hypertrophy to the degree similar to that of 1 week of TAC. Finally, expression of inducible Hsp 70 is upregulated in the heart of wild-type mice in both conditions. We produced these 2 different models for adaptive hypertrophy in HSF1⫹/⫺ mice and their wild-type littermates. Cardiac function of HSF1⫹/⫺ mice was significantly impaired in both models compared with their wild-type littermates. Moreover, expression of inducible Hsp 70 was markedly reduced in the heart of HSF1⫹/⫺ mice. Although the nature of 2 different stimuli may lead to the differential molecular response, these results suggest a critical role of HSF1 in adaptive hypertrophy. Downregulation of HSF1 activity in the chronic phase may be a key factor to promote the transition from adaptive to maladaptive hypertrophy. Alternatively, the different manifestations of hypertrophy (adaptive or maladaptive) may be partly attributed to the difference in HSF1 activity between the exercise model and the chronic pressure-overload model because reducing its activity caused maladaptive hypertrophy in both conditions. Sustained pressure overload induced cardiomyocyte death and fibrosis in the heart, both of which have been reported to cause heart failure.42 In the heart of HSF1 transgenic mice, the number of TUNEL-positive cardiomyocytes and the extent of fibrosis were significantly less than those of wild-type mice. These protective effects may be attributable to the functions of Hsps in protein folding and degradation. In addition to such well-known functions, accumulating evidence indicates that different Hsps directly act on the cell death machinery and inhibit the signaling pathway for cell death at various points.43 For example, Hsp27 binds to cytochrome c and prevent it binding to Apaf-1,44 whereas Hsp70 prevents Apaf-1 from recruiting pro– caspase-9,45 thereby inhibiting apoptotic cell death. Consequently, induction of HSF1 activity rather than individual Hsps may be more effective to protect the heart from severe stresses and will be an attractive therapeutic target in cardiovascular pathophysiology. Sources of Funding This work was supported by a grant-in-aid for scientific research from the Ministry of Education, Science, Sports, and Culture; health and labor sciences research grants from the Ministry of Health, Labor and Welfare; and a grant for research on life science from Uehara Memorial Foundation, Japan. HSF1 in Cardiac Hypertrophy 1417 Disclosures None. References 1. Katz AM. Cardiomyopathy of overload. A major determinant of prognosis in congestive heart failure. N Engl J Med. 1990;322:100 –110. 2. Levy D, Garrison RJ, Savage DD, Kannel WB, Castelli WP. Prognostic implications of echocardiographically determined left ventricular mass in the Framingham Heart Study. N Engl J Med. 1990;322:1561–1566. 3. Pluim BM, Zwinderman AH, van der Laarse A, van der Wall EE. The athlete’s heart. A meta-analysis of cardiac structure and function. Circulation. 2000;101:336 –344. 4. Adams TD, Yanowitz FG, Fisher AG, Ridges JD, Lovell K, Pryor TA. Noninvasive evaluation of exercise training in college-age men. Circulation. 1981;64:958 –965. 5. Pelliccia A, Maron BJ. Outer limits of the athlete’s heart, the effect of gender, and relevance to the differential diagnosis with primary cardiac diseases. Cardiol Clin. 1997;15:381–396. 6. Panidis IP, Kotler MN, Ren JF, Mintz GS, Ross J, Kalman P. Development and regression of left ventricular hypertrophy. J Am Coll Cardiol. 1984;3:1309 –1320. 7. Iemitsu M, Miyauchi T, Maeda S, Sakai S, Kobayashi T, Fujii N, Miyazaki H, Matsuda M, Yamaguchi I. Physiological and pathological cardiac hypertrophy induce different molecular phenotypes in the rat. Am J Physiol Regul Integr Comp Physiol. 2001;281:R2029 –R2036. 8. Kinugawa K, Yonekura K, Ribeiro RC, Eto Y, Aoyagi T, Baxter JD, Camacho SA, Bristow MR, Long CS, Simpson PC. Regulation of thyroid hormone receptor isoforms in physiological and pathological cardiac hypertrophy. Circ Res. 2001;89:591–598. 9. Abravaya K, Myers MP, Murphy SP, Morimoto RI. The human heat shock protein hsp70 interacts with HSF, the transcription factor that regulates heat shock gene expression. Genes Dev. 1992;6:1153–1164. 10. Morimoto RI. Cells in stress: transcriptional activation of heat shock genes. Science. 1993;259:1409 –1410. 11. Xiao H, Lis JT. Germline transformation used to define key features of heat-shock response elements. Science. 1988;239:1139 –1142. 12. Eto Y, Yonekura K, Sonoda M, Arai N, Sata M, Sugiura S, Takenaka K, Gualberto A, Hixon ML, Wagner MW, Aoyagi T. Calcineurin is activated in rat hearts with physiological left ventricular hypertrophy induced by voluntary exercise training. Circulation. 2000;101:2134 –2137. 13. Komuro I, Kurabayashi M, Takaku F, Yazaki Y. Expression of cellular oncogenes in the myocardium during the developmental stage and pressure-overloaded hypertrophy of the rat heart. Circ Res. 1988;62: 1075–1079. 14. Ishii M, Hashimoto S, Tsutsumi S, Wada Y, Matsushima K, Kodama T, Aburatani H. Direct comparison of GeneChip and SAGE on the quantitative accuracy in transcript profiling analysis. Genomics. 2000;68: 136 –143. 15. Nakai A, Suzuki M, Tanabe M. Arrest of spermatogenesis in mice expressing an active heat shock transcription factor 1. EMBO J. 2000; 19:1545–1554. 16. Harada K, Komuro I, Zou Y, Kudoh S, Kijima K, Matsubara H, Sugaya T, Murakami K, Yazaki Y. Acute pressure overload could induce hypertrophic responses in the heart of angiotensin II type 1a knockout mice. Circ Res. 1998;82:779 –785. 17. Inouye S, Izu H, Takaki E, Suzuki H, Shirai M, Yokota Y, Ichikawa H, Fujimoto M, Nakai A. Impaired IgG production in mice deficient for heat shock transcription factor 1. J Biol Chem. 2004;279:38701–38709. 18. Harada K, Komuro I, Shiojima I, Hayashi D, Kudoh S, Mizuno T, Kijima K, Matsubara H, Sugaya T, Murakami K, Yazaki Y. Pressure overload induces cardiac hypertrophy in angiotensin II type 1A receptor knockout mice. Circulation. 1998;97:1952–1959. 19. Shimoyama M, Hayashi D, Takimoto E, Zou Y, Oka T, Uozumi H, Kudoh S, Shibasaki F, Yazaki Y, Nagai R, Komuro I. Calcineurin plays a critical role in pressure overload-induced cardiac hypertrophy. Circulation. 1999;100:2449 –2454. 20. Kojima M, Shiojima I, Yamazaki T, Komuro I, Zou Z, Wang Y, Mizuno T, Ueki K, Tobe K, Kadowaki T. Angiotensin II receptor antagonist TCV-116 induces regression of hypertensive left ventricular hypertrophy in vivo and inhibits the intracellular signaling pathway of stretchmediated cardiomyocyte hypertrophy in vitro. Circulation. 1994;89: 2204 –2211. 1418 Circulation Research December 8/22, 2006 Downloaded from http://circres.ahajournals.org/ by guest on April 29, 2017 21. Teiger E, Than VD, Richard L, Wisnewsky C, Tea BS, Gaboury L, Tremblay J, Schwartz K, Hamet P. Apoptosis in pressure overloadinduced heart hypertrophy in the rat. J Clin Invest. 1996;97:2891–2897. 22. Rosenkranz S. TGF-beta1 and angiotensin networking in cardiac remodeling. Cardiovasc Res. 2004;63:423– 432. 23. Chan JY, Ou CC, Wang LL, Chan SH. Heat shock protein 70 confers cardiovascular protection during endotoxemia via inhibition of nuclear factor-kappaB activation and inducible nitric oxide synthase expression in the rostral ventrolateral medulla. Circulation. 2004;110:3560 –3566. 24. Chen Y, Currie RW. Small interfering RNA knocks down heat shock factor-1 (HSF-1) and exacerbates pro-inflammatory activation of NF-kappaB and AP-1 in vascular smooth muscle cells. Cardiovasc Res. 2006;69:66 –75. 25. Knowlton AA. NFkappaB, heat shock proteins, HSF-1, and inflammation. Cardiovasc Res. 2006;69:7– 8. 26. Benjamin IJ, McMillan DR. Stress (heat shock) proteins: molecular chaperones in cardiovascular biology and disease. Circ Res. 1998;83: 117–132. 27. Li GC, Laszlo A. Amino acid analogs while inducing heat shock proteins sensitize CHO cells to thermal damage. J Cell Physiol. 1985;122:91–97. 28. Pockley AG. Heat shock proteins, inflammation, and cardiovascular disease. Circulation. 2002;105:1012–1017. 29. Hutter MM, Sievers RE, Barbosa V, Wolfe CL. Heat-shock protein induction in rat hearts. A direct correlation between the amount of heat-shock protein induced and the degree of myocardial protection. Circulation. 1994;89:355–360. 30. Andres J, Sharma HS, Knoll R, Stahl J, Sassen LM, Verdouw PD, Schaper W. Expression of heat shock proteins in the normal and stunned porcine myocardium. Cardiovasc Res. 1993;27:1421–1429. 31. Suzuki K, Sawa Y, Kaneda Y, Ichikawa H, Shirakura R, Matsuda H. In vivo gene transfection with heat shock protein 70 enhances myocardial tolerance to ischemia-reperfusion injury in rat. J Clin Invest. 1997;99: 1645–1650. 32. Hollander JM, Martin JL, Belke DD, Scott BT, Swanson E, Krishnamoorthy V, Dillmann WH. Overexpression of wild-type heat shock protein 27 and a nonphosphorylatable heat shock protein 27 mutant protects against ischemia/ reperfusion injury in a transgenic mouse model. Circulation. 2004;110: 3544–3552. 33. Latchman DS. Heat shock proteins and cardiac protection. Cardiovasc Res. 2001;51:637– 646. 34. Kim YK, Suarez J, Hu Y, McDonough PM, Boer C, Dix DJ, Dillmann WH. Deletion of the inducible 70-kDa heat shock protein genes in mice 35. 36. 37. 38. 39. 40. 41. 42. 43. 44. 45. impairs cardiac contractile function and calcium handling associated with hypertrophy. Circulation. 2006;113:2589 –2597. Chang J, Wasser JS, Cornelussen RN, Knowlton AA. Activation of heat-shock factor by stretch-activated channels in rat hearts. Circulation. 2001;104:209 –214. Locke M, Tanguay RM. Diminished heat shock response in the aged myocardium. Cell Stress Chaperones. 1996;1:251–260. Yan LJ, Christians ES, Liu L, Xiao X, Sohal RS, Benjamin IJ. Mouse heat shock transcription factor 1 deficiency alters cardiac redox homeostasis and increases mitochondrial oxidative damage. EMBO J. 2002;21: 5164 –5172. Nishizawa J, Nakai A, Higashi T, Tanabe M, Nomoto S, Matsuda K, Ban T, Nagata K. Reperfusion causes significant activation of heat shock transcription factor 1 in ischemic rat heart. Circulation. 1996;94: 2185–2192. Hung JJ, Cheng TJ, Lai YK, Chang MD. Differential activation of p38 mitogen-activated protein kinase and extracellular signal-regulated protein kinases confers cadmium-induced HSP70 expression in 9L rat brain tumor cells. J Biol Chem. 1998;273:31924 –31931. Bueno OF, De Windt LJ, Tymitz KM, Witt SA, Kimball TR, Klevitsky R, Hewett TE, Jones SP, Lefer DJ, Peng CF, Kitsis RN, Molkentin JD. The MEK1-ERK1/2 signaling pathway promotes compensated cardiac hypertrophy in transgenic mice. EMBO J. 2000;19:6341– 6350. Perrino C, Naga Prasad SV, Mao L, Noma T, Yan Z, Kim HS, Smithies O, Rockman HA. Intermittent pressure overload triggers hypertrophyindependent cardiac dysfunction and vascular rarefaction. J Clin Invest. 2006;116:1547–1560. Nadal-Ginard B, Kajstura J, Leri A, Anversa P. Myocyte death, growth, and regeneration in cardiac hypertrophy and failure. Circ Res. 2003;92: 139 –150. Sreedhar AS, Csermely P. Heat shock proteins in the regulation of apoptosis: new strategies in tumor therapy: a comprehensive review. Pharmacol Ther. 2004;101:227–257. Bruey JM, Ducasse C, Bonniaud P, Ravagnan L, Susin SA, Diaz-Latoud C, Gurbuxani S, Arrigo AP, Kroemer G, Solary E, Garrido C. Hsp27 negatively regulates cell death by interacting with cytochrome c. Nat Cell Biol. 2000;2:645– 652. Beere HM, Wolf BB, Cain K, Mosser DD, Mahboubi A, Kuwana T, Tailor P, Morimoto RI, Cohen GM, Green DR. Heat-shock protein 70 inhibits apoptosis by preventing recruitment of procaspase-9 to the Apaf-1 apoptosome. Nat Cell Biol. 2000;2:469 – 475. Downloaded from http://circres.ahajournals.org/ by guest on April 29, 2017 Upregulation of Heat Shock Transcription Factor 1 Plays a Critical Role in Adaptive Cardiac Hypertrophy Masaya Sakamoto, Tohru Minamino, Haruhiro Toko, Yosuke Kayama, Yunzeng Zou, Masanori Sano, Eiichi Takaki, Teruhiko Aoyagi, Katsuyoshi Tojo, Naoko Tajima, Akira Nakai, Hiroyuki Aburatani and Issei Komuro Circ Res. 2006;99:1411-1418; originally published online November 9, 2006; doi: 10.1161/01.RES.0000252345.80198.97 Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Copyright © 2006 American Heart Association, Inc. All rights reserved. Print ISSN: 0009-7330. Online ISSN: 1524-4571 The online version of this article, along with updated information and services, is located on the World Wide Web at: http://circres.ahajournals.org/content/99/12/1411 Data Supplement (unedited) at: http://circres.ahajournals.org/content/suppl/2006/11/09/01.RES.0000252345.80198.97.DC1 Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in the Permissions and Rights Question and Answer document. Reprints: Information about reprints can be found online at: http://www.lww.com/reprints Subscriptions: Information about subscribing to Circulation Research is online at: http://circres.ahajournals.org//subscriptions/ Online Supplement Sakamoto et al. HSF1 in cardiac hypertrophy Supplemental Table 1 Echocardiographic analysis Sham PO EX PWTd (mm) 1.54±0.06 2.19±0.13**† 1.76±0.02* LVDd (mm) 6.6±0.1 7.61±0.16* 7.14±0.02* FS (%) 49.43±1.10 41.20±2.33**† 53.42±0.94 Sham, sham-operated rat; PO, pressure-overloaded rat; EX, exercised rat; PWTd, posterior wall thickness at diastole; LVDd, left ventricular dimension at diastole; FS, fractional shortening. *p<0.05, **p<0.01 vs. Sham; †p<0.01 vs. EX (n=8). Online Supplement Sakamoto et al. HSF1 in cardiac hypertrophy Supplemental Table 2 Hemodynamic parameters and heart weight Sham PO EX BP (mmHg) 99.3±7.2 144.6±9.0**† 84.4±4.3* HR (bpm) 318±19 323±15† 250±11** LVEDP (mmHg) 0.85±0.05 4.77±0.18* 1.12±0.10 LVW (mg)/BW(g) 1.83±0.05 2.77±0.19 **† 2.17±0.10* Sham, sham-operated rat; PO, pressure-overloaded rats; EX, exercised rats; BP, blood pressure; HR, heart rate; LVEDP, left ventricular end-diastolic pressure; LVW, left ventricular weight; BW, body weight. *p<0.05, **p<0.01 vs. Sham; †p<0.05 vs. EX (n=8). Online Supplement Sakamoto et al. HSF1 in cardiac hypertrophy Supplemental Figure Expression of inducible Hsp70 after pressure overload. Pressure overload was imposed on wild-type mice. Whole cell lysates were extracted from the hearts at the indicated time points and examined for expression of inducible Hsp70 by Western blot analysis. Supplementary figure Supplemental Figure TAC Hsp70 Actin 0 3 7 14 28 (days)