Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
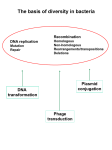
DSB Double-Strand Breaks causate da radiazioni stress ossidativo farmaci DSB e CROMATINA • Higher-order chromatin packaging is a barrier to the detection and repair of DNA damage • DSBs induce a local decrease in the density of the chromatin fibre, in addition to altering the position of nucleosomes • DSBs also elicit post-translational modifications on the protruding histone tails Chromating remodelling and DSBs RSC remodels the DSB chromatin The PIKKs Mec1 and Tel1 phosphorylate H2A(X), and RSC accumulates in the regions flanking the DSB SWI/SNF is recruited and remodels the donor template chromatin RSC complex RSC (remodels the structure of chromatin) ATP-dependent chromatin-remodelling RSC can mediate nucleosome sliding, alter histoneDNA contacts and remove histones from DNA. The chromatin-remodelling activity of RSC is important for transcriptional regulation of genes that are involved in stress responses and cell-cycle progression Chromating remodelling and DSBs Phosphorylated H2A recruits cohesin, which helps to bridge interactions between sister chromatids The HDAC complex Sin3–Rpd3 removes acetylation from H4. The protein kinase CK2 is also recruited and this phosphorylates H4 The INO80 complex enters the region of the DSB and removes some nucleosomes. MODIFICAZIONE ISTONI • Eukaryotes have several histone variants, which, as a result of their altered amino-acid composition, can affect both the structure of individual nucleosomes and the ability of nucleosomes to form higher order chromatin structure • The earliest and most robust modification induced by DSB is phosphorylation of the histone H2A variant H2AX on its extended C-terminal tail. • Within seconds, phosphorylated H2AX (known as γH2AX) spreads over a region spanning thousands to millions of bases surrounding a DSB H2AX extended Ctail PIKKs = phosphatidylinositol-3OH-kinase -like kinases DNA-damage sensor proteins Ku70–Ku80, MRN, RPA TOPBP1 DDR signal spreading DDR proteins initially accumulate at DSB sites and then spread at distance via a positive feedback loop involving MDC1, which binds gH2AX, the MRN complex, and ATM kinase, which phosphorylates additional H2AX molecules further away from the break site. Temporal regulation of DDR protein accumulation at DNA breaks Proteine piattaforma Mre, Rad50 SSB Nbs1, a subunit of a complex that recognizes DNA DSBs HRR kinase signaling ATM P The MDC1 TQXF motifs are ATM targets required for 53BP1 IRIF. (A) Domain architecture of MDC1, with ATM consensus sites (dots). Specialized binding modules for recognition of post-translational modifications (PTMs) at DNA breaks . Domain architecture of RNF8 Ubiquitin ligase activity Forkhead associated(FHA) domain bind phosphothreonine-bearing epitopes interaction with ATM-phosphorylated MDC1. • FHA-(R42A) and • Forkhead domain RING finger (C406s) mutants. Orchestration of the DNA-Damage Response by the RNF8 Ubiquitin Ligase (Nadine Science Feb2008) • Cells respond to DSBs by recruiting the DNAdamage mediator protein MDC1, the p53-binding protein 1 (53BP1), and the breast cancer susceptibility protein BRCA1 to sites of damaged DNA. Orchestration of the DNA-Damage Response by the RNF8 Ubiquitin Ligase (Nadine Science Feb2008) • Cells respond to DSBs by recruiting the DNAdamage mediator protein MDC1, the p53-binding protein 1 (53BP1) to sites of damaged DNA. • 53BP1 is an established player- important role in modulating chromatin structure surrounding the break site- in the cellular response to DNA damage and is a canonical component of ionizingradiation induced foci (IRIF) Ranking by z score of 500 siRNAs giving the least 53BP1 foci from a siRNA screen Orchestration of the DNA-Damage Response by the RNF8 Ubiquitin Ligase (Nadine Science Feb2008) • Cells respond to DSBs by recruiting the DNAdamage mediator protein MDC1, the p53-binding protein 1 (53BP1), and the breast cancer susceptibility protein BRCA1 to sites of damaged DNA. Irradiated (10 Gy) HeLa cells transfected with the indicated siRNAs were stained with antibodies to gH2AX, BRCA1 Orchestration of the DNA-Damage Response by the RNF8 Ubiquitin Ligase (Nadine Science Feb2008) • Cells respond to DSBs by recruiting the DNA-damage mediator protein MDC1, the p53-binding protein 1 (53BP1), and the breast cancer susceptibility protein BRCA1 to sites of damaged DNA. • The ubiquitin ligase RNF8 mediates ubiquitin conjugation and 53BP1 and BRCA1 focal accumulation at sites of DNA lesions. MDC1 recruits RNF8 through phosphodependent interactions between the RNF8 forkhead-associated domain and motifs in MDC1 that are phosphorylated by the DNA-damage activated protein kinase ataxia telangiectasia mutated (ATM). Depletion of the E2 enzyme UBC13 impairs 53BP1 recruitment to sites of damage, which suggests that it cooperates with RNF8. RNF8 promotes the G2/M DNA damage checkpoint and resistance to ionizing radiation. the DNA-damage response is orchestrated by ATM-dependent phosphorylation of MDC1 and RNF8-mediated ubiquitination. Mutazioni in BRCA 1 o 2 inattivazione meccanismo HRR predisposizione allo sviluppo di Carcinoma mammario ereditario, con insorgenza precoce tumore seno e ovaie BRCA 1: 50% mutazioni tumore mammario famigliare BRCA 2: 35% mutazioni tumore mammario famigliare Eredità di un allele mutante predisposizione al tumore, che insorge solo quando la seconda copia del gene è persa o mutata (perdita di eterozigosità) BRCA 1 • impairment of homologous repair in Brca1deficient mouse embryonic stem cells • increase in the frequency of NHEJ in Brca1deficient cells Riparazione per ricombinazione omologa (HRR) Ripara le DSBs (Double-Strand Breaks) causate da radiazioni, stress ossidativo, farmaci Replicazione e trascrizione vengono bloccate nel sito della DSB e le estremità esposte sono soggette a degradazione con perdita di materiale genetico importanza HRR HRR utilizza come stampo il cromatidio fratello protezione dagli errori HRR avviene in tarda fase S o in G2, quando i cromatidi fratelli sono vicini RPA (ss DNA replication binding protein) la polimerizzazione di RAD51 sul 3’ libero è BRCA1/2-dipendente RAD51 (aiutata dall’elicasi RAD54) cerca la sequenza omologa sul cromatidio fratello e invade la doppia elica; le regioni 3’ a singola elica si appaiano con quelle complementari Sensor MRN La DNA polimerasi allunga l’estremità 3’ libera (in cui si trova la nucleoproteina RAD51) utilizzando come stampo il cromatidio omologo non danneggiato L’altra regione 3’ a singola elica attorno alla zona danneggiata si appaia all’elica ormai corretta e gli eventuali gap sono riempiti da polimerasi e ligasi BRCA1/BRCA2 PALB2 and BRCA1 are co-mediators that directly and indirectly, respectively, interact with BRCA2 and assist in localizing it, with RAD51 bound, to the sites of DNA damage Human BRCA2 is a mediator that interacts directly with approximately eight RAD51 molecules and transports them to the site of ss-DNA bound by RPA Determination of regions required for the BRCA1-PALB2 interaction. (A) Graphical projection of association between PALB2 (residues 9 – 42) and BRCA1 (residues 1393–1424) coiledcoil domains. Positions of the heptad repeat (positions a to g) were predicted by the Coil program Boxed residues were experimentally demonstrated to be responsible for the heterooligomeric interaction between PALB2 and BRCA1. BRCA 1 • The BRCA1 tumor suppressor exists as a heterodimeric complex with BARD1, and this complex is thought to mediate many of the functions ascribed to BRCA1, including its role in tumor suppression. • The two proteins share a common structural organization : an N-terminal RING domain and two C-terminal BRCT motifs • • the BRCA1/BARD1 heterodimer enhances chromosome stability by promoting homology-directed repair (HDR) of double strand DNA breaks. BRCA 1 - Gene oncosoppressore, localizzato sul cromosoma 17 - Espressione nel nucleo di molti tipi cellulari N-ter: dominio RING FINGER, coordina il legame con 2 ioni zinco contatta DNA C-ter: dominio BRCT induce / reprime geni; interazione con RNA pol II BRCA 1 Ubc13 and Rnf8are recruited to sites of DNA damage through DNA-damage-induced phosphorylation of a chromatin-associated protein BRCA 1 • An E3 ubiquitin ligase mediates the transfer of activated ubiquitin from an E2 ubiquitinconjugating enzyme to its substrate lysine residues. • BRCA1 has the ability to direct the synthesis of specific polyubiquitin chain linkages, depending on the E2 bound to its RING. •Lega p53 attivando p21 •Attiva TNFa Regolazione delle proliferazione cellulare •Lega Rad50/51 e BRCA2 Riparazione HRR •Inibisce la trasduzione del segnale da parte del recettore per l’estrogeno Reprime la proliferazione dell’epitelio mammario che è estrogeno-dipendente Mutazioni in BRCA1 possono determinare un’incontrollata proliferazione, in particolare delle cellule che rispondono all’estrogeno tumore BRCA1 regulates human mammary stem/progenitor cell fate PNAS February 5, 2008 BRCA1 is implicated in multiple cellular functions Red, ubiquitylation; Blue, transcription; Green, cell cyclecheckpoint control; Light blue, DNA repair; Gray, chromatin modifications. Riparazione per ricombinazione non omologa (NHEJ) E’ il meccanismo prevalente per la riparazione delle DSBs, soprattutto negli organismi con elevato numero di sequenze ripetute identiche (l’HRR rischierebbe di appaiare sequenze ripetute di cromosomi diversi) Può causare l’introduzione di errori nel codice genetico (perdita o aggiunta di nucleotidi) La via nota è quella Ku-dipendente, ma il NHEJ può avvenire, con meno efficienza, anche in estratti cellulari privi di Ku e DNA-PK esistenza di vie Ku-indipendenti non note • Mammals have evolved at least two genetically discrete ways to mediate DNA DSB repair: homologous recombination (HR) and non-homologous end joining (NHEJ). • In mammalian cells, most DSBs are preferentially repaired by NHEJ. • NHEJ consists of at least two sub-pathways— • the main Ku heterodimer-dependent or “classic” NHEJ (CNHEJ) pathway and • an “alternative” NHEJ (A-NHEJ) pathway, which usually generates microhomology-mediated signatures at repair junctions. L’eterodimero Ku 70/80 lega le DSBs, in tal modo proteggendo le estremità libere dalla degradazione finchè la giunzione non è completa Ku recluta DNA-PK DNA-PK ha siti di legame sia per le DSBs sia per il DNA a doppia elica adiacente ad esse e promuove l’allineamento delle regioni che hanno microomologie Autophosphorylation of the DNAPKcatalytic subunit (DNA-PKcs) after recruitment of DNA-PKcs to cornerstone of the NHEJ pathway Ku translocates inwards DNA-PKcs interacts with the nuclease Artemis synaptic complex Dati sperimentali: Davydov et al, 2003 Analisi su omogenati estratti dal cervello di 39 pazienti affetti da Alzheimer (AD) e 7 soggetti sani di pari età (C) EMSA: electrophoretic mobility shift assay Utilizza una sonda radioattiva contenente la sequenza di legame per la proteina di interesse (in questo caso Ku) e incubata con estratti nucleari Principio: in un gel di PAA i complessi DNA-Proteine migrano più lentamente rispetto a DNA libero Capacità di legame di Ku al DNA Supershift: per confermare che la proteina che si è legata è Ku-80, si aggiunge nell’EMSA un anticorpo specifico. L’anticorpo andrà a legare la proteina Ku e se questa è legata al DNA marcato, il legame dell’anticorpo causerà un ulteriore ritardo nella corsa eleettrofeoretica Controllo sano Paziente Alzheimer Sonda – Ku - anticorpo Sonda - Ku Sonda libera Alterazione di una delle proteine coinvolte nel meccanismo di NHEJ può determinare invecchiamento precoce Dati sperimentali: Vogel et al., 1999 Topi privi di entrambe le copie del gene per Ku-86 (Ku 86-/-) Vita media Ku 86 -/-: 3814 wks Vita media controlli: 9717 wks Età massima raggiunta da un topo Ku 86 -/-: 87 wks Età massima raggiunta da un topo di controllo: 127 wks Topi Ku 86-/-: insorgenza precoce di tumori, atrofia della pelle e dei follicoli, chiusura delle epifisi, curvatura della spina dorsale, assottigliamento della pelle Topi DNA-PK-/-: non vanno incontro ad invecchiamento precoce, forse perché l’attività fosforilante di DNA-PK può essere svolta da altre chinasi presenti nella cellula Alterazioni nel NHEJ possono determinare invecchiamento precoce anche nell’uomo: •Sindrome di Werner: vita media di 50 anni, alopecia, osteoporosi, arteriosclerosi, cataratte, atrofia della pelle,… E’ causata da mutazioni nella proteina WRN, una DNA elicasi/nucleasi. WRN ha un ruolo sia nel HRR sia nel NHEJ (interagisce con Ku) IPOTESI: nei soggetti sani, quando la ricombinazione omologa non è possibile, WRN lega Ku e facilita la riparazione della DSB mediante NHEJ Nei pazienti affetti, invece, la ricombinazione omologa diventa meno efficiente e anche l’alternativa offerta dal NHEJ non è più favorita dal legame tra WRN e Ku