Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Recent Developments
in TEXTAL
Phenix Workshop
Berkeley
Sept. 2006
Thomas R. Ioerger
Texas A&M University
NCS Identification via Pattern Recognition
• Pai, R., Sacchettini, J.C. and Ioerger, T.R. (2006).
Identifying non-crystallographic symmetry in protein
electron-density maps: a feature-based approach. Acta
Crystallographica, D62(9):1012-1021.
• The Problem:
– Symmetry averaging can greatly improve phases.
– Typical methods for finding NCS require ≥ 3 heavy
atoms, and are sensitive to errors in coordinates.
– Despite noise and breaks from symmetry, similar
patterns of density exist over large regions of real
space (even if imperfectly phased).
– How to efficiently identify these similarities and derive
symmetry operators?
Our Approach to NCS
• Step 1: calculate backbone using CAPRA
– Putative C-alpha atoms become centers of regions for initial
matching
• Step 2: Calculate local features for each CA based on
pattern of surround CA’s and density; select subset of
candidates that are likely to be similar
– Example features: #CAs, center of mass, moments of inertia,
std.dev., skewness, kurtosis…
• Step 3: Calculate local density correlation between each
pair of CA’s (over 5A spheres), with rotation-optimization
• Step 4: Cluster pairs of matching regions with similar
rotation matrices
– How can you tell if two local transformations are
related (from same pair of domains)?
– Each can transform the coordinates of the other.
Definition 1: similar rotation matrices. Given RUV and RPQ as
rotation matrices that optimally superpose regions U and V
and regions P and Q, respectively, and u, v, p and q as the
coordinates of the centers of regions U, V, P and Q, respectively,
then RUV is similar to RPQ if q RUV p ≤ 2 A° and
u RPQ v ≤ 2 A°.
P
V
U
Q
• Step 5: Extend regions to molecular boundaries
(excluding non-symmetric deviations)
• - caveat: doesn’t work for proper symmetry
(can’t identify unique boundaries)
• Step 6: Organize and output N-1 operators
• (Step 7): Run DM to do symmetry-averaging
Results on Experimental Maps
Protein
Native
reso (Å)
# NCS
subunits
in ASU
Map
corr.
# NCS
subunits
found
RMS of
superposition
NCSaveraged
map corr.
1a7a
2.8
2
0.845
2
0.670
0.859
1bkj
1.8
2
0.443
2
0.819
0.600
1l1e
2.8
2
0.505
2
0.739
2gmf
2.35
2
2
0.857
1f61
1.8
2
2
0.655
1nye
3
8
0.506
7
0.713, 0.757,
0.771, 0.819,
0.844, 0.917
1kwa
1.93
2
0.475
2
1.43
1l8w
2.3
4
0.454
4
0.82, 0.858,
1.09
1p32
2.25
3
3
0.801, 0.883
1nf2
3
3
0.313
3
0.954, 0.979
1ytt
1.8
2
0.667
2
0.780
0.531
0.692
2a2u
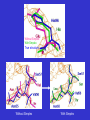
1a7a
1p32
One subunit (identified by algorithm) superposed on the other
subunits using symmetry operators (also identified by algorithm)
Availability
• Pattern Recognition Algorithm for NCS (by Reetal Pai,
PhD student in Ioerger lab)
– Initial implementation in C and csh scripts
– User input: structure factors (.mtz), expected # copies
– Runs CAPRA, extracts features, matches regions…
– Automatically runs DM to improve phases via averaging
– Output:
• NCS operators
• masks for each region
• C-alpha chains for each region
• NCS-averaged structure factors (.mtz)
• Web server: textal.tamu.edu/NCS
– Users can upload reflection file; results emailed back
Port to Python
• Command line
# first source phenix_setup and ccp4_setup
>textal.find_ncs prot.mtz <N> <FP> <PHIB> <FOM>
...
Outputs: prot_ncs_ops.dat, prot_ncs_avg.mtz
prot_mask_1.xplor, prot_mask_2.xplor...
prot_region_1.pdb, prog_region_2.pdb...
• Script-level API
from textal.find_ncs import find_ncs
from textal.io.reflection_file import reflection_file
ref = reflection_file("mbp.mtz")
obj = find_ncs(reflections=ref,copies=2,
amplitude='FP',phases='PHIB',FOM='FOM')
obj.find_ncs()
(rot_mat,trans_vec) = obj.get_operators(0)
model1 = obj.get_subunit(0) # type pdb_extended
mask1 = obj.get_mask(0)
# type emap
Improving Sequence Alignment with Simplex
• Romo, T.R., Sacchettini, J.C. and Ioerger, T.R. (2006).
Improving Amino Acid Identification, Fit, and C-alpha
Prediction using the Simplex Method in Automated ModelBuilding. Acta Crystallographica, accepted.
• The Problem:
– Most model-building programs build backbone first,
then try to recognize side-chains (using probabilities,
free atoms, features…)
– Identification of amino acids is sensitive to errors in
predicted Ca coordinates (often up to 1Å rms)
– Even if sequence alignment is used to correct
mistakes, initial side-chains must be sufficiently
accurate
Our Approach: Simplex Optimization
• Simplex is a classic optimization algorithm
– High radius of convergence
– Does not require explicit computation of derivatives
• Simplex can be applied to refine individual residues
as rigid bodies (translation+rotation)
– Several programs do local real-space rigid-body
refinement of individual side-chains to improve fit.
– Typically, applied after aa identity has been determined
• We apply Simplex in Textal (LOOKUP) during
residue selection, to help pick the template from our
database that matches the local density pattern
best, allowing the Ca atom to shift up to 2Å
Effect of Errors in Ca Coordinates
Artificially-introduced
Ca errors, starting
from perfect Ca’s from refined model
70
Percent
Strict amino
Amino Acid acid
Identity identity
60
50
40
Original LOOKUP
Simplex LOOKUP
30
20
10
0
0
0.2
0.4
0.6
0.8
Ca-RMSD (Å)
1
1.2
1.4
Accuracy of amino acids output by LOOKUP for
CzrA (without sequence alignment)
1.6
Procedure
• Step 1: Given a Ca, extract density-based
features and retrieve K=400 most similar
regions from database
• Step 2: Re-rank by local density correlation (5Å)
– Original method:
• try to find optimal rotation only
worst score
– New method:
mean of
• Generate initial Simplex: N+1 perturbations of
rest
configuration vector (6-DOF)
new
• Evaluate density correlation coefficient of each
• Pick the lowest, and ‘reflect’ over average of
remaining configuration vectors
6D config. space
Vector representing original position (3 coords) and orientation (3 angles) of side-chain
Results on Experimental Maps
Protein
Reso.
Mean phase error
Map corr.
CzrA
2.3 Å
18.1
0.95
If5a
2.1 Å
36.8
0.91
MVK
2.4 Å
42.8
0.84
ICL
3.0 Å
44.1
0.81
PcaA
2.8 Å
54.2
0.73
Percent identity of model compared to true (refined) structure:
without Simplex
with Simplex
no alignment
with alignment
no alignment
with alignment
CzrA
40.0
94.4
47.8
93.3
If5a
30.2
92.2
38.8
93.0
MVK
18.1
40.1
30.8
77.6
ICL
23.5
55.3
26.0
76.4
PcaA
15.6
38.7
19.3
47.4
average:
25.5
64.1
32.5
77.5
Without Simplex
With Simplex
True structure
Without Simplex
With Simplex
TEXTAL for Molecular Replacement
• Motivation:
– Why not exploit the MR
search model if available?
– No excuse for mistakes in
connectivity or aa identities
• Steps toward larger goal of
Model Completion
• Idea:
– Rotate search model into
density (MR solution)
– Replace amino acid identities
with new sequence
– Run LOOKUP to build sidechains into new density
• Issues:
– Backbones sometimes diverge (e.g. in loops)
– Phase improvement: How to identify and edit-out
incorrect parts of the model built?
– Avoiding model bias
• Our Approach:
–
–
–
–
Use CAPRA to generate backbone for new density
Match up Ca’s with search model (core of protein)
Identify divergences (no nearby matches)
Fill in gaps with chains from new density
• Method
– Generate map around search model (MR solution)
– Run CAPRA to generate new backbone
– Assign Ca’s (closest match
between models, up to 3Å)
5.35Å
Deletion in model
– Assign new aa identities based on sequence alignment
supplied by user
ATAAEIAALPRQKVELVDPPFVHAHSQVAEGGPKVVEFTMVI----IVIDDAGTEVHAM...
-------ELPVIDAVTTHAPEVPPAI--DRDYPAKVRVKMETVEKTMKMDD-GVEYRYW...
• Format restricted (for now) to 2 long lines (or N pairs of lines for
N subunits in search model)
• Connect small gaps (len≤5)
– Common (including due to alignment errors)
– Method 1: Look for a bridge using existing Ca’s
– Method 2: Use a fragment library
• 4188 9-mers extracted from 238 non-homologous proteins
with min RMS of 1.25Å
• Superpose edges of each fragment on chain ends, with
expected number of missing Ca’s in middle
• Select top 25 fragments by RMS (typically in range of 1-2Å)
• Evaluate each fragment based on density measured every
0.5Å along fragment
• Score(frag) = S –exp(-(r-1))
–exp(-(r-1))
r
– Run patch to make any remaining
connections
• More indiscriminant; may skip residues or insert
extra atoms not consistent with alignment
• Can turn off via --connectivity=conservative
– Run ca_refine
• reduces variance in inter-Ca distances
– Run LOOKUP to build side-chains
– Run simulated annealing
Results
• 3 MR datasets from Phenix structure library:
native
reso
-----a2u-globulin
2.5 Å
human-otc
2.4 Å
nitrite-reductase 1.7 Å
search
model
----mup
a1s
kbv
perc
ident
----63%
48%
35%
sec
size
str
------158(x4) alpha
354
mixed
339
beta
MR map
Rtrue
corr
-------0.20/0.26 0.94
0.23/0.27 0.89
0.26/0.29 0.81
* Rtrue is R-factor after simulated annealing with refined structure
* MR map corr is density correl. between initial MR map and final 2Fo-Fc
• After building model with textal.build_mr and running simulated
annealing:
a2u-globulin
human-otc
nitrite-reductase
perc
num
built chains
---- ----93%
4/4
93%
2
84%
4
perc
ident
----98%
99%
93%
Rmod
---0.24/0.30
0.30/0.36
0.35/0.39
map
corr
----0.95
0.82
0.85
* Rmod is R-factor of model built by Textal, after simulated annealing
* Map corr is between model 2Fo-Fc and refined 2Fo-Fc density maps
* ideal sequence alignments were used based on structural alignments
generated using Shindyalov’s CE (Combinatorial Extension) algorithm
a2u-globulin (white)
Textal model (green)
11 res
N-term tail
not built
disordered
loop, res 60-64
human-otc (white)
Textal (red, green)
loop not built,
res 266-275
C-term not built,
res 345-352
human-otc (white)
Textal (red, green)
missing loop:
res 186-205
nitrite-reductase (white)
Textal model (colors)
missing term:
res 5-10
missing term:
res 334-342
missing loop:
res 159-170
missing loop:
res 29-36
nitrite-reductase (white)
kbv (MR solution, purple)
large
divergent
loop
small
differences
loop insertion
Initial Steps Toward Model Evaluation
Run SFCHECK on model built…
Identifying errors with SFCHECK
Thr-203
Gly-226
Glu-236
Thr-269
...
0.092
0.297
0.306
0.354
quality score (Sfcheck)
• Which combination of values correlates best
with errors in model?
• Use backbone_density_index from SFCHECK
as residue quality score
residues (sorted)
Residues in purple (50/284)
are those with low backbone
density index scores
(<0.92)
Re-running SA on editted models
Hypothesis: impact of completeness versus accuracy of
model on R-factor
random deletions
Num
residues
deleted
Rwork
Rfree
Rwork
Rfree
0
35.0
38.5
35.0
38.5
10
35.2
37.9
36.4
39.3
20
35.9
38.8
37.2
39.0
30
35.1
37.4
37.7
39.8
Issues:
40
35.2
37.5
38.3
42.6
50
35.9
38.6
39.0
41.6
• B-factors
• side-chains
• lack of HETATMs (2 Cu, 3Cd, 244 HOH in refined structure)
• avoid model bias (use omit maps?)
Availability
• Phenix command line:
textal.build_mr [-c] [--symmetry] [--amplitudes] [--phases]
<reflections> <search_model> <alignment_file>
textal.build_mr --symmetry=nitrite-redct.inp –amplitudes=FULL_MOD
nitrite-reduce.hkl kbv_mr_solution.pdb NR-KBV-align.txt
• Python API:
from textal.users.tom.textal_mr import MR_build
MR_build(reflections=rx,model=mod,alignment=algn,capra_only=True)
• Phenix GUI task: (textal/MR_Build):
Conclusion
• TEXTAL can build highly accurate models for
Molecular Replacement (completely automatically),
with almost perfect coordinates for backbone and
side-chains atoms (with the help of simulated
annealing), at least in the core (80-90%)
Future Work
• Handle missing domains in the search model
• Incorporate better model evaluation methods
• Automate the whole improvement cycle