Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
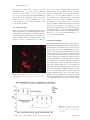
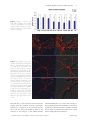
Addiction Biology PRECLINICAL STUDY: FULL ARTICLE doi:10.1111/j.1369-1600.2010.00218.x Altered architecture and functional consequences of the mesolimbic dopamine system in cannabis dependence Saturnino Spiga1, Alessandra Lintas2, Michele Migliore3 & Marco Diana2 Department of Animal Biology and Ecology, University of Cagliari, Italy1, “G.Minardi” Laboratory of Cognitive Neuroscience, Department of Drug Sciences, University of Sassari, Italy2 and Institute of Biophysics, National Research Council, Italy3 ABSTRACT Cannabinoid withdrawal produces a hypofunction of mesencephalic dopamine neurons that impinge upon medium spiny neurons (MSN) of the forebrain. After chronic treatment with two structurally different cannabinoid agonists, D9-tetrahydrocannabinol and CP55 940 (CP) rats were withdrawn spontaneously and pharmacologically with the CB1 antagonist SR141716A (SR). In these two conditions, evaluation of tyrosine hydroxylase (TH)-positive neurons revealed significant morphometrical reductions in the ventrotegmental area but not substantia nigra pars compacta of withdrawn rats. Similarly, confocal analysis of Golgi–Cox-stained sections of the nucleus accumbens revealed a decrease in the shell, but not the core, of the spines’ density of withdrawn rats. Administration of the CB1 antagonist SR to control rats, provoked structural abnormalities reminiscent of those observed in withdrawal conditions and support the regulatory role of cannabinoids in neurogenesis, axonal growth and synaptogenesis by acting as eu-proliferative signals through the CB1 receptors. Further, these measures were incorporated into a realistic computational model that predicts a strong reduction in the excitability of morphologically altered MSN, yielding a significant reduction in action potential output. These pieces of evidence support the tenet that withdrawal from addictive compounds alters functioning of the mesolimbic system and provide direct morphological evidence for functional abnormalities associated with cannabinoid dependence at the level of dopaminergic neurons and their postsynaptic counterpart and are coherent with recent hypothesis underscoring a hypodopaminergic state as a distinctive feature of the ‘addicted brain’. Keywords Cannabinoids, dopamine, nucleus accumbens, THC, VTA, withdrawal. Correspondence to: Marco Diana, Department of Drug Sciences, University of Sassari, Via Muroni 23/A, 07100 Sassari, Italy. E-mail: [email protected] INTRODUCTION Cannabis preparations have long been considered moderately harmful and mildly addictive possibly because until recently, no clear-cut description of withdrawal syndrome is provided. Clinical reports, however, recently described that chronic consumers of even low daily doses of cannabis derivatives experience, upon cessation of drug administration, overt abstinence signs (Haney et al. 1999, 2004; Budney & Hughes 2006), and these observations are paralleled by structural and functional neuroimaging studies of cannabis use (Iversen 2003; Quickfall & Crockford 2006). This abstinence syndrome is of clinical significance (Cooper & Haney 2008), similar to the onset and time course to that of other abused substances, and diagnostic criteria for cannabis withdrawal syndrome have been proposed (Haney et al. 1999, 2004). Likewise, overt somatic signs of cannabinoid withdrawal can be elicited in experimental models of cannabinoid dependence by administering the competitive antagonist of cannabinoid CB1 receptor SR141716A (SR) (Rinaldi-Carmona et al. 1994; Aceto et al. 1996; Diana et al. 1998; Gonzalez et al. 2004) to animals exposed to a chronic cannabinoid regimen, thereby offering the possibility to investigate neurobiological effects of addicting chemicals in a condition that mimics human addictive behaviour (Pulvirenti & Diana 2001; Melis, Spiga & Diana 2005). The morphological analysis of neurons and dendrites (Zito & Svoboda 2002; Zito et al. 2004) has recently seen © 2010 The Authors. Journal compilation © 2010 Society for the Study of Addiction Addiction Biology 2 Saturnino Spiga et al. an explosion in the studies of the consequences of longterm administration of drugs (Sklair-Tavron et al. 1996; Robinson & Kolb 1997, 2004; Spiga et al. 2003, 2005) since these measures are suggested to reflect plasticity of active synapses and therefore synaptic remodelling as a consequence of experience and drug exposure (Robinson & Kolb 2004). Addiction in this regard, as a condition characterized by long-term drug exposure, can be conceptualized as one example of experience-dependent plasticity, whereby experience (i.e. long-term exposure to addictive drugs) may long-lastingly affect behavioural, cognitive and psychological functions (Robinson & Kolb 2004; Melis et al. 2005). Cannabis withdrawal produces a marked reduction of electrophysiological activity in nucleus accumbens (NAcc)-projecting dopamine (DA) containing neurons of the rat midbrain (Diana et al. 1998) and a reduction of DA outflow in the NAcc shell (Tanda, Loddo & Di Chiara 1999). This functional evidence suggests that cannabinoid withdrawal may structurally alter cellular elements of the mesolimbic system as it was recently shown for opiate dependence (Spiga et al. 2003, 2005; Diana, Spiga & Acquas 2006). Accordingly, a chronic tetrahydrocannabinol (THC) treatment has been shown to increase the length of the dendrites as well as the number of dendritic branches in the shell of the NAcc and in the medial prefrontal cortex but not in other brain areas (Kolb et al. 2006). However, because morphological evaluations were made long after drug discontinuation (30 days), it is impossible to ascertain from these observations the relationship between structural changes and onset of withdrawal. Thus, in the present study, we investigated the morphological alterations affecting neurons of the rat ventrotegmental area (VTA), substantia nigra pars compacta (SNc) and their postsynaptic counterparts in the NAcc subregions. TH-positive neurons and Golgi–Cox-stained MSN were evaluated after chronic cannabinoid treatment and withdrawal in order to obtain further insights into the morphological features of the mesolimbic DA system and its involvement in cannabis dependence, whereas the role of endogenous cannabinoids was investigated through administration of the CB1 receptor antagonist SR. To further investigate on the functional relevance of the morphological changes after withdrawal, we used a biophysical model of MSNs to estimate the alteration in the spiking activity produced by cannabis dependence. MATERIALS AND METHODS Subjects, drugs and cannabinoid treatment Male Sprague-Dawley albino rats (n = 48; Charles River, Como, Italy), weighing 200–225 g at the beginning of treatment were used. The rats were kept on a 12-hour light/12-hour dark cycle with food and water available ad libitum. Experimental protocols were approved by the Ethical Committee (EC) of the University of Sassari and performed in strict accordance with the EC regulations for the use of experimental animals (CEE N°86/609), and recommended guidelines for the care and use of experimental animals approved by the Society for Neuroscience. The rats were administered twice daily (8 am and 8 pm). Time cycle changed at 8 am and 8 pm. Drug injections took place one in the light and one in the dark for 6.5 days with either D9-THC (Sigma, Milan, Italy) or CP 55 940 (Sigma-Aldrich, Milan, Italy) emulsified in 1% Tween 80 then diluted in a saline solution and administered i.p. in a volume of 3 ml/kg. The animals were assigned to the following groups: chronic saline (1% Tween) (CTRL) (n = 6); chronic CP (0.4 mg/kg) (CP-Chr) (n = 6); chronic D9-THC (15 mg/ kg) (THC-Chr) (n = 6); 24 hours spontaneous withdrawal from chronic CP (CP-Sw) (n = 6); 24 hours spontaneous withdrawal from chronic THC (THC-Sw) (n = 6); SR (5 mg/kg) precipitated withdrawal from chronic CP (0.4 mg/kg/administration) (CP-SR) (n = 6); SR (5 mg/ kg) precipitated withdrawal from chronic D9-THC (15 mg/kg/administration) (THC-SR) (n = 6); and SR (5 mg/kg) in CTRL (SR) (n = 6) (Diana et al. 1998). On the morning of day 7, the rats received the first daily administration (vehicle, CP or THC) and 1 hour before sacrifice were tested for signs of spontaneous and SR-precipitated withdrawal (not shown, see Diana et al. 1998). Histology The animals were anesthetized with urethane (1.3 g/kg i.p.) before transcardiac perfusion with 100 ml of ice-cold saline solution immediately followed by 400 ml of icecold 4% paraformaldehyde. The brains were divided into two parts at approximately -2 mm from bregma. VTA-SNc TH-immunolabelling The posterior (VTA- and SNc-containing) part of the brains (Fig. 1) was postfixed for 24 hours in the 4% paraformaldehyde solution and cryoprocteted in 30% sucrose in phosphate-buffered saline (PBS). Coronal sections between -5.80 mm and -6.30 mm for VTA-SN (25 mm thick) from bregma according to Paxinos & Watson (1998) (Fig. 1) were obtained with a cryostat (Micron Cryo-Star HM 560, Walldorf, Germany). The sections for TH-immunolabelling were washed for 3 ¥ 5 minutes in PBS, immersed for 30 minutes in 10% normal goat serum (NGS) in 0.1 M PBS added with 0.5% Triton X-100 (PBS-TX) and incubated for two hours with a mouse monoclonal anti-TH antibody (1:500; © 2010 The Authors. Journal compilation © 2010 Society for the Study of Addiction Addiction Biology Mesolimbic dopamine system in cannabis dependence 3 Chemicon, Temecula, CA, USA) in PBS-TX. The sections were then washed (3 ¥ 5 minutes) in PBS-TX and incubated with a biotinylated antimouse IgG (1:300, Vector Laboratories, Burlingame, CA, USA) in PBS-TX and 1% NGS for 30 minutes, rinsed (3 ¥ 5 minutes) in PBS-TX and incubated with avidin-TRITC (1:200, SigmaAldrich) in PBS-TX and 1% NGS overnight at 4°C. All of the sections were then washed (3 ¥ 20 minutes) in PBS-TX and cover-slipped with Glycergel mounting solution (Dako, Milan, Italy). Free-floating slices were rinsed in distilled water for one minute and placed in ammonium hydroxide (30%) in the dark. After rinsing (one minute), the slices were placed in Kodak Fix Film solution (Ilford, UK) for 40 minutes in the dark, then the, slices were rinsed again (two minutes) and fixed (for 10 minutes) with Kodak Paper fixative 1:7. After rinsing and dehydration by alcohol ascendant scale (from 50° to 100°), the slices were placed in xylene (three minutes). Finally, the slices were mounted and coverslipped by Canada balsam. NAcc Golgi-Cox staining Image processing: surface rendering techniques After perfusion, the anterior (NAcc-containing) part of the brains (Fig. 1) were immediately rinsed (15 minutes ¥ 3 times) in 0.1 M PBS and immersed in a Golgi– Cox solution (Glaser & Van der Loos 1981) composed by 5% potassium dycromate, 5% mercurium chloride and 5% potassium cromate (pH 6.5). The solution was changed once after two days, and the brains were then left in fresh Golgi–Cox solution for an additional 14 days. After this period, the brains were cryoprotected with a 30% sucrose solution for two to three days. Beginning at 1.70 mm and ending at 0.70 mm from bregma, 50 mm thick coronal slices, according to Paxinos & Watson (1998) (Fig. 1b), were obtained with a cryostat. Leica 4-D confocal laser scanning microscope (Leica Microsystems, Heidelberg, Germany) with an argon– krypton laser was used to analyze the TH-positive neurons and the Golgi–Cox-stained sections. Confocal images were generated using 40¥ oil (na = 1.00–0.5) and 100¥ oil (na = 1.3). Each frame was acquired eight times and then averaged to obtain noise-free images. Optical sections, usually at consecutive intervals of 0.5 mm in z-axis, were imaged through the depth of the labelled neurons and saved as image stacks as previously described (Tredici et al. 1993; Spiga et al. 2003, 2005). Maximum intensity algorithm (ImageJ) was used for three-dimensional (3-D) reconstructions of TH- Figure 1 Schematic illustration, modified from Paxinos & Watson (1998), of the location of sampled neurons. VTA-SNc (left panel) and NAcc’s shell/core (right panel). Distance from bregma is indicated © 2010 The Authors. Journal compilation © 2010 Society for the Study of Addiction Addiction Biology 4 Saturnino Spiga et al. immunolabelled cells, while extended focus algorithm was used for 3-D reconstructions of Golgi–Cox-stained neurons (Bitplane Imaris V.5.7.2). Morphometric determinations Cell body morphometry Morphometric analyses were performed by two independent observers blind to pharmacological treatments. TH-immunolabelled somata (n = 80/group) were collected from a square area (approximately 200 mm/side). When totally included in the sections, the TH-positive neurons were reconstructed in 3-D and used for measurements and statistical analysis using Bioscan Optimas software (v 6.5.1; Media Cybernetics Inc., MD, USA). The cell bodies were manually marked following their profile, excluding all dendritc trunks, to measure their area (mm2), perimeter, major length (MJ) and minor axis length (min). Spines’ Counts For each group (n = 80), dendritic segments (at least 20 mm long) of second-order dendrites were collected for analysis from 0.7–1.70 mm from bregma (58) and identified by confocal-rendered cells. The spines’ density was calculated by tracing a 10–15 mm long ‘sp.’ line along the dendritic trunk and counting the number of spines therein. The procedure was repeated along the entire dendritic length from the bifurcation from the first branch of primary dendrites to the next bifurcation. Statistical analysis was performed by means of oneway analysis of variance (ANOVA) followed by the Student t-test for post hoc comparisons. Computational modelling For computational modelling, MSNs were reconstructed using a modified version of Neuron Morpho plugin for ImageJ v1.1.6 and Neuromantic v1.6.3. All simulations were carried out with the NEURON program (v7, 61; Yale University, CT, USA). A custom 3-D reconstruction of a medium spiny neuron composed of 519 segments, explicitly including 216 spines (accounting for 31% of the dendritic membrane), was used for all simulations. Using the average experimental values for the membrane time constant [7 ms, obtained from nucleus accumbens shell neurons at physiological temperature (O’Donnell & Grace 1993) and a standard value for the membrane capacitance (1 mF/cm2], the model neuron resulted in an input resistance of 160 MW. To model the basic active properties, we used channel models downloaded from the public ModelDB repository (http://senselab.med.yale.edu/ModelDB/), implemented to reproduce the electrophysiological properties of NAcc medium spiny neurons (Wolf et al. 2005). Excitatory Figure 2 Histological sections depict sampled areas. TH-positive neurons in the rat midbrain coronal section (top) and MSN Golgistained in NAcc (bottom). Abbreviations indicate: cp, cerebral peduncle, basal part; VTA, vental tegmental area; SNc, substantia nigra pars compacta (amino - 3 - hydroxyl - 5 - methyl - 4 - isoxazole - propionate [AMPA]) synaptic input was implemented on all synapses using a double exponential conductance change mechanism with a rise and decay time constants of 0.5 and 3 ms, respectively. The same peak synaptic conductance (0.15 nS) was used for all synapses, and different values (0.2–0.6 nS) were tested obtaining the same qualitative results (not shown). The up and down states observed in these neurons in vivo (Gruber & O’Donnell 2009) were implemented, activating the synapses asynchronously and randomly (Poissonian) at an average frequency of 3 Hz for the down state (in the theta rhythm range) or at 40 Hz (in the gamma range). The up state was activated at 0.8 Hz with a duration of 400 ms (Gruber & O’Donnell 2009). Average spiking rates under different conditions were calculated from the last 20 seconds of simulations lasting 22 seconds. A movie of the first four seconds from simulations under control of after withdrawal is shown as supplemental material (see Supporting Information Movie S1). The complete model and simulation files are available for public download at the ModelDB database. RESULTS VTA and SNc cells size Confocal datasets of TH-immunolabelled neurons located in the VTA and in the dorsomedial portion of SNc (Figs 1 & 2) (Gonzalez-Hernandez & Rodriguez 2000) were mor- © 2010 The Authors. Journal compilation © 2010 Society for the Study of Addiction Addiction Biology Mesolimbic dopamine system in cannabis dependence 5 Table 1 Effect of treatments and one-way ANOVA results, on morphometrical changes in DA (a) neurons and (b) spine density of accumbal MSN. (a) Mesencephalic TH-positive neurons morphometry SNC CTRL CP-Chr THC-Chr CP-Sw THC-Sw CP-SR THC-SR SR VTA CTRL CP-Chr THC-Chr CP-Sw THC-Sw CP-SR THC-SR SR Area (mm2) (%) Perimeter (%) MJ length (%) Min length (%) 156.4 ! 3.4 (100.0) 158.2 ! 2.3 (101.2) 157.7 ! 4.5 (100.8) 156.6 ! 3.1 (100.1) 156.3 ! 3.9 (99.9) 157.1 ! 2.9 (100.4) 158.4 ! 2.8 (101.3) 150.6 ! 2.7 (96.3) 53.2 ! 0.7 (100.0) 52.8 ! 0.5 (99.2) 52.2 ! 0.8 (98.0) 53.3 ! 0.7 (100.1) 51.1 ! 0.7 (96.0) 52.9 ! 0.6 (99.4) 52.4 ! 0.6 (98.5) 52.0 ! 0.7 (97.7) 20.7 ! 0.3 (100.0) 21.2 ! 0.3 (102.8) 21.3 ! 0.4 (103.3) 20.9 ! 0.4 (101.1) 20.1 ! 0.3 (97.2) 21.3 ! 0.3 (102.9) 20.1 ! 0.3 (97.3) 20.7 ! 0.4 (100.2) 11.5 ! 0.2 (100.0) 11.1 ! 0.2 (97.0) 11.3 ! 0.2 (98.8) 11.1 ! 0.2 (96.7) 11.5 ! 0.2 (100.4) 10.9 ! 0.2 (95.2) 11.6 ! 0.2 (100.6) 11.0 ! 0.3 (96.2) 181.5 ! 3.6 (100) 177.4 ! 3.5 (97.7) 177.2 ! 4.2 (97.6) 116.3 ! 2.4 (64.1) 130.7 ! 3.0 (72.0) 124.1 ! 2.3 (68.4) 124.7 ! 2.9 (68.7) 128.4 ! 2.5 (70.7) 56.2 ! 0.7 (100) 56.5 ! 1.0 (100.7) 54.4 ! 0.7 (96.8) 44.5 ! 0.6 (79.2) 46.6 ! 0.8 (83.0) 45.4 ! 0.5 (80.9) 45.4 ! 0.6 (80.9) 46.1 ! 0.9 (82.1) 21.9 ! 0.4 (100) 21.2 ! 0.6 (96.8) 20.4 ! 0.3 (93.2) 17.2 ! 0.4 (78.2) 18.7 ! 0.3 (85.4) 17.5 ! 0.5 (79.7) 17.7 ! 0.2 (80.6) 17.7 ! 0.4 (80.7) 12.4 ! 0.3 (100) 12.5 ! 0.2 (101.0) 12.6 ! 0.3 (102.2) 10.0 ! 0.2 (80.6) 10.3 ! 0.2 (83.5) 10.2 ! 0.3 (82.4) 10.1 ! 0.3 (82.1) 10.4 ! 0.2 (84.2) (b) MSN Secondary dendrites spines density (spine/10 mm) CTRL CP-Chr THC-Chr CP-Sw THC-Sw CP-SR THC-SR SR Shell (%) Core (%) 8.065 ! 0.096 (100) 8.431 ! 0.164 (106.54) 7.885 ! 0.151 (97.76) 5.383 ! 0.132 (66.75) 5.121 ! 0.186 (63.49) 4.611 ! 0.144 (57.17) 6.113 ! 0.121 (75.79) 5.898 ! 0.149 (73.12) 10.741 ! 0.138 (100) 10.565 ! 0.143 (98.36) 10.590 ! 0.156 (98.59) 10.433 ! 0.162 (97.13) 10.443 ! 0.148 (97.22) 10.706 ! 0.216 (99.67) 10.617 ! 0.173 (98.85) 10.427 ! 0.131 (97.08) Data are expressed as mean ! standard error. phometrically analyzed in order to evaluate the effects of treatments. In accordance with previous reports, these neurons exhibited considerable variability in shape and size (ovoid, polygonal or fusiform) (Grace & Bunney 1983; Oades & Halliday 1987; Tepper, Sawyer & Groves 1987; Spiga et al. 2003) in both areas (Table 1, Fig. 3). ANOVA showed anatomical differences between the experimental groups in the VTA of the mean calculated area (F639 = 82.83; P < 0.0001), perimeter (F639 = 71.35; P < 0.0001), MJ (F639 = 37.71; P < 0.0001) and min (F639 = 32.76; P < 0.0001) length. Post hoc analysis revealed that cell bodies in the VTA exhibited smaller somata after both withdrawal conditions. In particular, a mean reduction as compared with CTRL was found for area [(t158 = 15.2: P < 0.0001); (t158 = 10.9; P < 0.0001)], perimeter [(t158 = 14.3 P < 0.0001); (t158 = 11.6; P < 0.0001)], MJ [(t158 = 11.6; P < 0.0001); (t158 = 7.6; P < 0.0001)] and min length [(t158 = 8.43 P < 0.0001); (t158 = 6.88; P < 0.0001)] of CP-Sw and THC-Sw, respectively. Further, we found similar results for precipitated withdrawal with the CB1 antagonist SR-treated rats after chronic CP [area (t158 = 12.3 P < 0.0001), perimeter (t158 = 12.8 P < 0.0001), MJ (CP-SR t158 = 9.45, P < 0.0001), min (CP-SR t158 = 7.06, P < 0.0001] and THC [area (t158 = 13.5, P < 0.0001), perimeter (t158 = 12.8, P < 0.0001), MJ (t158 = 11.1; P < 0.0001), min (t158 = 7.22; P < 0.0001)]. Specifically, CP-SR and THC-SR showed a statistically significant reduction of morphometric parameters compared with CTRL. Unexpectedly, we also found changes in the SR group qualitatively similar with the other withdrawal conditions [area (t158 = 12.1, P < 0.0001), perimeter (t158 = 13.2 P < 0.0001), MJ (t158 = 10.4, P < 0.0001), min (t158 = 6.9, P < 0.0001)]. On the contrary, no significant changes were observed in both the CP-Chr [area (t158 = 0.82, P = 0.41), perimeter (t158 = 0.39, P = 0.69), © 2010 The Authors. Journal compilation © 2010 Society for the Study of Addiction Addiction Biology 6 Saturnino Spiga et al. MJ (t158 = 0.29, P = 0.77), min (t158 = 0.39, P = 0.7)] and the THC-Chr [area (t158 = 0.70, P = 0.48), perimeter (t158 = 1.83, P = 0.068), MJ (t158 = 1.92, P = 0.055), min (t158 = 0.77, P = 0.44)] groups. No statistical differences were found for the TH-positive neurons from SNc [area (F639 = 0.58; P = 0.77), perimeter (F639 = 1.17; P = 0.31), MJ (F639 = 1.68; P = 0.11), Min (F639 = 1.41; P = 0.19)] (Fig. 4, Table 1). NAcc MSN spines density Figure 5 show the effects of all the treatments on dendritic spines density of accumbal MSN (shell and core) versus CTRL. The spine densities of the CTRL groups were significantly different between shell and core (Fig. 6). A one-way ANOVA revealed a significant effect on the shell (F639 = 107.2; P < 0.0001) but not on the core Figure 3 Representative confocal picture of TH-positive neurons in theVTA, obtained by a projection of 53 scans for a total of 26.5 mm in the z-axis (F639 = 0.58; P = 0.77) on spine density in the experimental groups. Post hoc analysis showed a selective reduction on spine density for spontaneous withdrawal [CP-Sw (t158 = 16.4 P < 0.0001), THC-Sw (t158 = 14.4 P < 0.0001)] and pharmacologically precipitated [CP-SR (t158 = 19.9 P < 0.0001), THC-SR (t158 = 12.6 P < 0.0001)] as well as SR(t158 = 12.2; P < 0.0001) groups in respect to CTRL. These reductions were found to be about 33% for CP-Sw, 36 % for THC-Sw, 42 % for CP-SR, 24% for THC-SR and 26% for SR. Further, the post hoc analysis failed to reveal any significant difference between spine density counts in the shell MSN for the CP-Chr (t158 = 1.2; P = 0.056) and the THC-Chr (t158 = 1; P = 0.32) groups versus CTRL. Computational modelling The possible functional consequences of the morphological changes were investigated using a realistic computational model of NAcc shell neurons to study how the average firing rate could be modified after drug withdrawal. To model the effects of drug withdrawal, we compared the simulation results under a control condition assuming that all synapses were active, with the results from two simulations in which 30% randomly chosen synaptic compartments were deleted from the morphology. Furthermore, to model the well-known changes in the dopaminergic D2 input, and especially its inhibitory effect on the AMPA currents (up to a 15% reduction of the peak current, Hernández-Echeagaray et al. 2004), we tested the possible effects of different amounts of reduction in the dopaminergic input by increasing the peak synaptic conductances up to 115% of the control value. Typical traces are shown in Fig. 7b, and the modelling results are summarized in Fig. 7c. A 30% reduction in the active synapses caused a ~45% decrease in the average Figure 4 Morphometric measures of VTA TH-positive neurons. Data are presented as % of control ! SEM. * indicates P < 0.05 versus CTRL © 2010 The Authors. Journal compilation © 2010 Society for the Study of Addiction Addiction Biology Mesolimbic dopamine system in cannabis dependence 7 Figure 5 Histograms represent the mean ! SEM of dendritic spines densities (number of spines/10 mm of second-order dendrites) of NAcc shell and core MSN. * indicates P < 0.05 versus CTRL (student t-test post hoc analysis) Figure 6 Representative confocal reconstructions synthesizing major findings in the NAcc. (a) Golgi–Cox-stained MSNs from core CTRL group, (b) MSN from shell CTRL group and (c) MSN from shell the THC-Sw group, using extended focus algorithm (left part) by Voxblast v3 (SGI Iris). Right panels show the same reconstructed neurons on the left, using ‘Surpass’ and ‘Filament Tracer’ by Bitplane Imaris v5.7.2 software. ‘Filament Tracer’ is used for the automatic detection of filaments in 3-D objects (in confocal microscopy), and it is based on interactive thresholding and anatomical and geometrical properties (length and radius) of the filaments as a dendritic branches. From the resulting database, it is possible to extract various information including the ‘terminal points’ number. This feature can offer a representation of dendritic spines (green dots) of entire scanned neurons firing rate (Fig. 7c, 100% DA input). However, the model suggests that this reduction could be progressively balanced by a reduction in the DA input because this would also reduce the inhibitory effect of the activation of D2 receptors on the AMPA channels (Hernández-Echeagaray et al. 2004). The reduction of the dopaminergic input after withdrawal could thus be considered as a kind of homeostatic signalling (reviewed in Davis 2006) to maintain the functional role of these neurons under pathological conditions. © 2010 The Authors. Journal compilation © 2010 Society for the Study of Addiction Addiction Biology 8 Saturnino Spiga et al. (a) (b) (c) Figure 7 Simulation results. (a) The 3-D reconstruction of a NAcc medium spiny shell neuron used in all simulations; (b) typical 5-second somatic traces during simulations under control conditions (top), and after 30% reduction of the active synapses (bottom); (c) average number of APs (with respect to control) as a function of dopaminergic input DISCUSSION The results of the present study show that withdrawal from a regimen of chronic cannabinoid administration profoundly affects the morphological characteristics of TH-positive neurons of the rat VTA and spines’ density of dendrites of MSN of NAcc shell. In contrast, SNc TH positive neurons and core spines density were unaffected. In particular, withdrawal from chronic administration of the synthetic CB1 agonist CP determined the shrinkage of the somatic region of TH-positive neurons of the VTA as assessed by the decrease of area, perimeter, MJ and min. These changes were paralleled by a reduction of the spines’ density in the NAcc shell. Thus, spontaneous withdrawal appears responsible for the morphological changes detected in the VTA and NAcc shell. The specificity of these effects can be reasonably attributed to abrupt removal of the chronic cannabinoid because when chronic administration was pharmacologically interrupted, by administration of the CB1 antagonist SR, such morphological measures were affected similarly. Further supporting this contention, THC’s spontaneous and precipitated withdrawal yielded similar results both in the VTA and NAcc shell. In contrast, chronic administration of both compounds fails to affect the morphology of VTA neurons and spines’ density in the NAcc, thus pointing to a critical role of cannabinoid withdrawal in the shrinkage of mesencephalic neurons and the spines’ loss in the NAcc shell. The spine loss described in the present study is in line and significantly extends previous findings (Kolb et al. 2006) that reported an elongation of the dendrites as well as the number of dendritic branches in the shell of the nucleus accumbens and in the medial prefrontal cortex but not in other brain areas. This further strengthens the view that withdrawal from chronic cannabinoids exerts powerful and long-lasting (Kolb et al. 2006) changes in key brain structures affected by addicting compounds. In particular, the spines’ loss observed here, besides contributing to reduce the already abated DA transmission, could help in explaining the down regulation of CB1 receptors after THC-withdrawal (Breivogel et al. 2003) as well as the CB1-mediated inhibition of excitatory synaptic transmission at the excitatory synapses between the prefrontal cortex and the NAcc (Mato et al. 2005). Administration of the CB1 antagonist SR in salinetreated rats produced effects qualitatively similar to those observed in the subjects treated with the exogenous cannabinoids CP and THC in both areas examined (i.e. VTA and NAcc shell). This unexpected finding would seem to suggest that endogenous cannabinoids are involved in the trophic control of key elements of the mesolimbic system such as VTA neurons and their physiological postsynaptic site. While further experiments are needed to corroborate this notion, the present finding supports the idea of an endocannabinoid trophic and protective role (Galve-Roperh et al. 2006, 2007) at the level of the DA system (Melis et al. 2006). Accordingly, endocannabinoids modulate synaptic plasticity in the VTA (Melis et al. 2004), SN pars reticulata (Szabo et al. 2000) and striatum (Gerdeman & Lovinger 2001). Alternatively, the possibility that SR might be acting as an inverse agonist (Rubino et al. 2000; Bass et al. 2002) should be considered and further experiments will be needed to clarify this issue. Irrespective of the mechanisms underlying SR-induced changes, the structural observations reported here are likely to have profound consequences on dopaminergic transmission in the shell of the NAcc, both pre- and postsynaptic levels. Indeed, the reduced DA © 2010 The Authors. Journal compilation © 2010 Society for the Study of Addiction Addiction Biology Mesolimbic dopamine system in cannabis dependence firing (Diana et al. 1998) is accompanied by a ‘shrinkage’ of the somatic region, thereby rendering the cell more excitable in line with the ‘size principle’ (Shepherd 1994; Spiga et al. 2003). At the postsynaptic side, the reduced number of spines would reduce total membrane surface (Gray 1959; Rall 1962), thereby decreasing membrane resistance (Wilson 1984) eventually leading to altered excitability. This possibility is in line with classical theoretical predictions (Chang 1952), and subsequent confirmatory experimental tests (Wilson 1984), which ascribed to the spine an attenuating effect on synaptic potentials. Importantly, recent studies (Hoffman et al. 2003) employing a chronic regimen of THC very similar to that employed here have reported that long-term exposure and subsequent withdrawal (recordings were performed 24 hours after last treatment) of the active ingredient of marijuana blocks synaptic plasticity in the NAcc and reduces the sensitivity of GABAergic and glutamatergic synapses. What could then be the functional consequence of losing about 30% of the synapses? There are two mechanisms directly related with the reduction in the number of synapses that can affect the overall firing rate of the neuron. On the one hand, the loss of neuronal membrane associated with the loss synapses (~9% of the total membrane in our model morphology) increases the input resistance of the neuron, and in principle, this results in a more excitable neuron; on the other hand, the neuron may decrease its firing rate because the overall excitatory input is reduced. The model’s result suggests that the reduced excitatory input upon withdrawal might be the dominant mechanism, at least under the conditions tested in this work. However, the model also suggests a possible compensatory mechanism: because of the reduced dopaminergic input on these neurons upon withdrawal, the inhibitory effect of D2 receptor activation on AMPA channels will also be reduced, amplifying the effect of the remaining synapses (Fig. 7). Overall, the present data suggest that the altered architecture of the DA system projecting to the shell of the NAcc documented here would profoundly alter the synaptic equilibrium affecting various neurotransmistters involved in the neurobiological mechanisms of cannabis dependence (Pulvirenti & Diana 2001; Melis et al. 2005). On the basis of the present and previous findings (Spiga et al. 2003, 2005; Diana 1996; Diana et al. 1995, 1999, 2006), we suggest that shrinkage of DA neurons and reduction of spines’ densities in their postsynaptic elements (i.e. MSN) upon withdrawal from chronic cannabinoids, might represent a morphological correlate of the functional deficits detected by electrophysiological (Diana et al. 1998) and neurochemical (Tanda et al. 1999) means that ultimately may contribute to negative 9 motivational properties of withdrawal from addictive drugs (Koob & Le Moal 2001; Di Chiara 2002; Melis et al. 2005). In general, the present data lends further support to the notion that drug addiction can be seen as a chronic drug-induced, aberrant form of neural plasticity (Nestler & Aghajanian 1997; Nestler 2001; Pulvirenti & Diana 2001; Robinson & Kolb 2004; Melis et al. 2005), whereby DA neurons originating in the VTA represent major cellular substrate involved at molecular (Nestler 2001), cellular (White 1996; Pulvirenti & Diana 2001; Diana & Tepper 2002) and behavioural levels (Koob & Le Moal 1997, 2001; Berridge & Robinson 1998; Di Chiara 1999; Robinson & Berridge 2001) and are coherent with recent hypothesis (Melis et al. 2005) that underscores a hypodopaminergic state as a distinctive feature of the ‘addicted brain’. Disclosure/Conflict of Interest The authors declare that over the past three years MD has received compensation from ASMED srl and ESSEX Italia for work unrelated to the material presented here. Acknowledgements This work was supported, in part, by a grant from MIUR (PRIN. N°2004052392) to M.D. The authors wish to thank G. Di Chiara for a gift of THC and William Dunn III for proofreading the article. Authors Contribution SS and AL performed research and analysed the date. MM contributed the computational evaluation and analysed the data. MD designed, researched and wrote the paper. All authors have critically reviewed content and approved final version submitted for publication. References Aceto MD, Scates S, Lowe JA, Martin BR (1996) Dependence on D9-Tetrahydrocannabinol: studies on precipitated and abrupt withdrawal. J Pharmacol Exp Ther 278:1290–1295. Bass CE, Griffin G, Grier M, Mahadevan A, Razdan RK, Martin BR (2002) SR-141716A-induced stimulation of locomotor activity. A structure–activity relationship study. Pharmacol Biochem Behav 74:31–40. Berridge KC, Robinson TE (1998) What is the role of dopamine in reward: hedonic impact, reward learning or incentive salience? Brain Res Rev 28:309–369. Breivogel CS, Scates SM, Beletskaya IO, Lowery OB, Aceto MD, Martin BR (2003) The effects of delta9-tetrahydrocannabinol physical dependence on brain cannabinoid receptors. Eur J Pharmacol 459:139–150. Budney AJ, Hughes JR (2006) The cannabis withdrawal syndrome. Curr Opin Psychiatry 19:233–238. Review. © 2010 The Authors. Journal compilation © 2010 Society for the Study of Addiction Addiction Biology 10 Saturnino Spiga et al. Chang HT (1952) Cortical neurons with particular reference to the apical dendrites. Cold Spring Harb Symp Quant Biol 17:189–202. Cooper ZD, Haney M (2008) Cannabis reinforcement and dependence: role of the cannabinoid CB1 receptor. Addict Biol 13:188–195. Davis GW (2006) Homeostatic control of neural activity: from phenomenology to molecular design. Annu Rev Neurosci 29:307–323. Di Chiara G (1999) Drug addiction as dopamine-dependent associative learning disorder. Eur J Pharmacol 375:13–30. Review. Di Chiara G (2002) Nucleus accumbens shell and core dopamine: differential role in behavior and addiction. Eur J Pharmacol 137:75–114. Diana M (1996) Dopaminergic neurotransmission and drug withdrawal: relevance to drug craving. In: Ohye VC, Kimura M, McKenzie J, eds. The Basal Ganglia Adv. Behav. Biol, pp. 123–130. New York: Plenum Press. Diana M, Melis M, Muntoni AL, Gessa GL (1998) Mesolimbic dopaminergic decline after cannabinoid withdrawal. Proc Natl Acad Sci U S A 95:10269–71023. Diana M, Muntoni AL, Pistis M, Melis M, Gessa GL (1999) Lasting reduction in mesolimbic dopamine neuronal activity after morphine withdrawal. Eur J Neurosci 11:1037–1041. Diana M, Pistis M, Muntoni A, Gessa G (1995) Profound decrease of mesolimbic dopaminergic neuronal activity in morphine withdrawn rats. J Pharmacol Exp Ther 72:781– 785. Diana M, Spiga S, Acquas E (2006) Persistent and reversible morphine withdrawal-induced morphological changes in the nucleus accumbens. Ann N Y Acad Sci 1074:446–457. Diana M, Tepper JM (2002) Electrophysiological pharmacology of mesencephalic dopaminergic neurons. In: Di Chiara G, ed. Handbook of Experimental Pharmacology 154/II, pp. 1–62. Berlin: Springer-Verlag. Galve-Roperh I, Aguado T, Palazuelos J, Guzmán M (2007) The endocannabinoid system and neurogenesis in health and disease. Neuroscientist 13:109–114. Galve-Roperh I, Aguado T, Rueda D, Velasco G, Guzmán M (2006) Endocannabinoids: a new family of lipid mediators involved in the regulation of neural cell development. Curr Pharm Des 12:2319–2325. Gerdeman G, Lovinger DM (2001) CB1 cannabinoid receptor inhibits synaptic release of glutamate in rat dorsolateral striatum. J Neurophysiol 85:468–471. Glaser EM, Van der Loos H (1981) Analysis of thick brain sections by obverse-reverse computer microscopy: application of a new, high clarity Golgi–Nissl stain. J Neurosci Methods 4:117–125. Gonzalez S, Fernandez-Ruiz J, Di Marzo V, Hernandez M, Arevalo C, Nicanor C, Cascio MG, Ambrosio E, Ramos JA (2004) Behavioral and molecular changes elicited by acute administration of SR141716 to Delta9-tetrahydrocannabinoltolerant rats: an experimental model of cannabinoid abstinence. Drug Alcohol Depend 74:159–170. Gonzalez-Hernandez T, Rodriguez M (2000) Compartmental organization and chemical profile of dopaminergic and GABAergic neurons in the substantia nigra of the rat. J Comp Neurol 421:107–135. Grace AA, Bunney BS (1983) Intracellular and extracellular electrophysiology of nigral dopaminergic neurons-2. Action potential generating mechanisms and morphological correlates. Neuroscience 10:317–331. Gray EG (1959) Axo-somatic and axo-dendritic synapses of the cerebral cortex: an electron microscope study. J Anat 93:420– 433. Gruber AJ, O’Donnell P (2009) Bursting activation of prefrontal cortex drives sustained up states in nucleus accumbens spiny neurons in vivo. Synapse 63:173–180. Haney M, Hart CL, Vosburg SK, Nasser J, Bennett A, Zubaran C, Foltin RW (2004) Marijuana withdrawal in humans: effects of oral THC or divalproex. Neuropsychopharmacology 29:158– 170. Haney M, Ward AS, Comer SD, Foltin RW, Fischman MW (1999) Abstinence symptoms following oral THC administration to humans. Psychopharmacology 141:395–404. Hernández-Echeagaray E, Starling AJ, Cepeda C, Levine MS (2004) Modulation of AMPA currents by D2 dopamine receptors in striatal medium-sized spiny neurons: are dendrites necessary? Eur J Neurosci 19:2455–2463. Hoffman AF, Oz M, Caulder T, Lupica CR (2003) Functional tolerance and blockade of long-term depression at synapses in the nucleus accumbens after chronic cannabinoid exposure. J Neurosci 23:4815–4820. Iversen L (2003) Cannabis and the brain. Brain 126:1252– 1270. Kolb B, Gorny G, Limebeer CL, Parker LA (2006) Chronic treatment with delta-9-tetrahydrocannabinol alters the structure of neurons in the nucleus accumbens shell and medial prefrontal cortex of rats. Synapse 60:429–436. Koob GF, Le Moal M (1997) Drug abuse: hedonic homeostatic dysregulation. Science 278:52–58. Koob GF, Le Moal M (2001) Drug addiction, dysregulation of reward, and allostasis. Neuropsychopharmacology 24:97– 129. Mato S, Robbe D, Puente N, Grandes P, Manzoni OJ (2005) Presynaptic homeostatic plasticity rescues long-term depression after chronic delta 9-tetrahydrocannabinol exposure. J Neurosci 25:11619–11627. Melis M, Pillolla G, Bisogno T, Minassi A, Petrosino S, Perra S, Muntoni AL, Lutz B, Gessa GL, Marsicano G, Di Marzo V, Pistis M (2006) Protective activation of the endocannabinoid system during ischemia in dopamine neurons. Neurobiol Dis 24:15–27. Melis M, Pistis M, Perra S, Muntoni AL, Pillolla G, Gessa GL (2004) Endocannabinoids mediate presynaptic inhibition of glutamatergic transmission in rat ventral tegmental area dopamine neurons through activation of CB1 receptors. J Neurosci 24:53–62. Melis M, Spiga S, Diana M (2005) The dopamine hypothesis of drug addiction: hypodopaminergic state. Int Rev Neurobiol 63:101–154. Nestler EJ (2001) Molecular basis of long-term plasticity underlying addiction. Nat Rev Neurosci 2:119–128. Nestler EJ, Aghajanian GK (1997) Molecular and cellular basis of addiction. Science 278:58–63. O’Donnell P, Grace AA (1993) Physiological and morphological properties of accumbens core and shell neurons recorded in vitro. Synapse 13:135–160. Oades RD, Halliday GM (1987) Ventral tegmental (A10) system: neurobiology. 1. Anatomy and connectivity. Brain Res 434:117–165. Paxinos G, Watson C (1998) The Rat Brain in Stereotaxic Coordinates, 3rd edn. Sydney: Academic Press. Pulvirenti L, Diana M (2001) Drug dependence as a disorder of neural plasticity: focus on dopamine and glutamate. Rev Neurosci 12:141–158. © 2010 The Authors. Journal compilation © 2010 Society for the Study of Addiction Addiction Biology Mesolimbic dopamine system in cannabis dependence Quickfall J, Crockford D (2006) Brain neuroimaging in cannabis use: a review. J Neuropsychiatry Clin Neurosci 18:318–332. Rall W (1962) Electrophysiology of a dendritic neuron model. Biophys J 2:145–167. Rinaldi-Carmona M, Barth F, Héaulme M, Shire D, Calandra B, Congy C, Martinez S, Maruani J, Néliat G, Caput D et al. (1994) SR141716A, a potent and selective antagonist of the brain cannabinoid receptor. FEBS Lett 350:240–244. Robinson TE, Berridge KC (2001) Incentive-sensitization and addiction. Addiction 96:103–114. Robinson TE, Kolb B (1997) Persistent structural modifications in nucleus accumbens and prefrontal cortex neurons produced by previous experience with amphetamine. J Neurosci 17:8491–8497. Robinson TE, Kolb B (2004) Structural plasticity associated with exposure to drugs of abuse. Neuropharmacology 47(Suppl. 1):33–46. Rubino T, Viganò D, Zagato E, Sala M, Parolaro D (2000) In vivo characterization of the specific cannabinoid receptor antagonist, SR141716A: behavioral and cellular responses after acute and chronic treatments. Synapse 35:8–14. Shepherd GM (1994) Neurobiology, 3rd edn. Oxford: University Press. Sklair-Tavron L, Shi WX, Lane SB, Harris HW, Bunney BS, Nestler EJ (1996) Chronic morphine induces visible changes in the morphology of mesolimbic dopamine neurons. Proc Natl Acad Sci U S A 93:11202–11207. Spiga S, Puddu MC, Pisano M, Diana M (2005) Morphine withdrawal-induced morphological changes in the nucleus accumbens. Eur J Neurosci 22:2332–2340. Spiga S, Serra GP, Puddu MC, Foddai M, Diana M (2003) Morphine withdrawal-induced abnormalities in the VTA: confocal laser scanning microscopy. Eur J Neurosci 17:605– 612. Szabo B, Wallmichrath I, Mathonia P, Pfreundtner C (2000) Cannabinoids inhibit excitatory neurotransmission in the substantia nigra pars reticulata. Neuroscience 97:89– 97. Tanda G, Loddo P, Di Chiara G (1999) Dependence of mesolimbic dopamine transmission on D9-tetrahydrocannabinol. Eur J Pharmacol 376:23–26. Tepper JM, Sawyer SF, Groves PM (1987) Electrophysiologically identified nigral dopaminergic neurons intracellularly labeled 11 with HRP: light-microscopic analysis. J Neurosci 7:2794– 2806. Tredici G, Di Francesco A, Miani A, Pizzini G (1993) Real complete three-dimensional reconstruction of Golgi-impregnated neurons by means of a confocal laser scanning microscope. Neuroimage 1:87–93. White FJ (1996) Synaptic regulation of mesocorticolimbic dopamine neurons. Annu Rev Neurosci 19:405–436. Wilson CJ (1984) Passive cable properties of dendritic spines and spiny neurons. J Neurosci 4:281–297. Wolf JA, Moyer JT, Lazarewicz MT, Contreras D, Benoit-Marand M, O’Donnell P, Finkel LH (2005) NMDA/AMPA ratio impacts state transitions and entrainment to oscillations in a computational model of the nucleus accumbens medium spiny projection neuron. J Neurosci 25:9080–9095. Zito K, Knott G, Shepherd GM, Shenolikar S, Svoboda K (2004) Induction of spine growth and synapse formation by regulation of the spine actin cytoskeleton. Neuron 44:321– 334. Zito K, Svoboda K (2002) Activity-dependent synaptogenesis in the adult Mammalian cortex. Neuron 35:1015–1017. Review. SUPPORTING INFORMATION Additional Supporting Information may be found in the online version of this article: Movie S1 Movie of the first four seconds from two simulations modelling control conditions (left plots) and after withdrawal (right plots). Top traces are somatic membrane potential, bottom plots illustrate the time course of the membrane potential over the entire neuron. Different colours correspond to different membrane potentials according to the colour scale Please note: Wiley-Blackwell is not responsible for the content or functionality of any supporting materials supplied by the authors. Any queries (other than missing material) should be directed to the corresponding author for the article. © 2010 The Authors. Journal compilation © 2010 Society for the Study of Addiction Addiction Biology