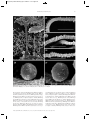
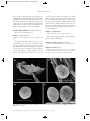
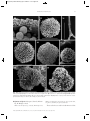
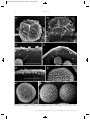
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
portada y contra 68(1) NOVEDADES2:portada y contra 66(1).qxd 16/06/2011 11:22 Página 1 ISSN: 0211-1322 Zamora, N.A. El caso de Lonchocarpus costaricensis (Leguminosae, Papilionoideae), una especie endémica de Costa Rica: un complejo taxonómico-nomenclatural, y una nueva especie / The case of Lonchocarpus costaricensis (Leguminosae, Papilionoideae), an endemic species of Costa Rica: a taxonomic-nomenclatural complex, and a new species ................................................................ Cano-Maqueda, J. & Talavera, S. A taxonomic revision of the Campanula lusitanica complex (Cam pa nulaceae) in the Western Mediterranean region / Una revisión taxonómica del com plejo Campanula lusitanica (Campanulaceae) en la región occidental mediterránea ........................................................................................................................................... Venhuis, C. & Oostermeijer, J.G.B. Distinguishing colour variants of Serapias perez-chiscanoi (Orchidaceae) from related taxa on the Iberian Peninsula / Distinción de variantes en color de Serapias perezchiscanoi (Orchidaceae) en relación con táxones de la Península Ibérica .......................................... Lado, C., Wrigley de Basanta, D. & Estrada-Torres, A. Biodiversity of Myxomycetes from the Monte Desert of Argentina / Biodiversidad de Myxomycetes en el Desierto de Monte (Argentina) ............... Peñas, J., Lorite, J., Alba-Sánchez, F. & Taisma, M.A. Self-incompatibility, floral parameters, and pollen characterization in the narrow endemic and threatened species Artemisia granatensis (Asteraceae) / Autoincompatibilidad, parámetros florales y caracterización de polen en la especie endémica y amenazada Artemisia granatensis (Asteraceae) ...................................................................................... Schneider, A.A. & Boldrini, I.I. Microsculpture of cypselae surface of Baccharis sect. Caulopterae (Asteraceae) from Brazil / Microescultura de la superficie de las cipselas de Baccharis sect. Caulopterae (Asteraceae) de Brasil ...................................................................................................................... Domínguez-Álvarez, S., Rico, J.M. & Gil-Rodríguez, M.C. Photosynthetic response and zonation of three species of Gelidiales from Tenerife, Canary Islands / Respuestas fotosintéticas y zonación de tres especies de Gelidiales de Tenerife, Islas Canarias ...................................................................... http://rjb.revistas.csic.es N.º 1 enero-junio 2011 Madrid (España) ISSN: 0211-1322 Madrid SUMARIO / CONTENTS Volumen 68 7/14 15/47 2011 Madrid (España) N.º 1 enero-junio 2011 49/59 61/95 97/105 107/116 117/124 Volumen 68 N.º 1 Anales del Jardín Botánico de Madrid Volumen 68 REAL JARDÍN BOTÁNICO www.publicaciones.csic.es CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS 01 primeras:01 primeras.qxd 13/06/2011 11:56 Página 1 Volumen 68 N.º 1 enero-junio 2011 Madrid (España) ISSN: 0211-1322 CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS 01 primeras:01 primeras.qxd 13/06/2011 11:56 Página 2 01 primeras:01 primeras.qxd 13/06/2011 11:56 Página 3 Volumen 68 N.º 1 enero-junio 2011 Madrid (España) ISSN: 0211-1322 ANALES DEL JARDÍN BOTÁNICO DE MADRID publica artículos originales e inéditos sobre taxonomía y sistemática vegetal y fúngica y campos relacionados, como biogeografía, bioinformática, conservación, ecofisiología, filogenia, filogeografía, florística, morfología funcional, nomenclatura o relaciones planta-animal, incluyendo trabajos de síntesis y revisión. ANALES DEL JARDÍN BOTÁNICO DE MADRID publishes original and unpublished articles in the taxonomy and systematics of all plant groups and fungi, including related fields like biogeography, bioinformatics, conservation, ecophysiology, phylogeny, phylogeography, functional morphology, nomenclature and plant-animal relations, as well as reviews and summary works. ANALES DEL JARDÍN BOTÁNICO DE MADRID cuenta con un equipo técnico de edición interno, lo que reduce al mínimo el tiempo necesario para procesar un original. En la mayoría de los casos un original puede estar publicado en un periodo de 7-8 meses. Los artículos serán publicados en formato PDF, el mismo en el que se entregarán las separatas a los autores. ANALES DEL JARDÍN BOTÁNICO DE MADRID has a technical editing team that allows reducing to a minimum the time needed to process manuscripts. Usually, articles are published within a period of 7-8 months. Articles are published in the Anales del Jardín Botánico de Madrid web page in PDF format. Se publica en Acceso Abierto en Internet. Su URL es: http://rjb.revistas.csic.es It publishes in Access Opened in Internet. Its URL is: http://rjb.revistas.csic.es Directora / Editor-in-Chief Inés Álvarez, Real Jardín Botánico, CSIC, España Secretaria / Secretary Blanca Landázuri, Real Jardín Botánico, CSIC, España Consejo de Redacción - Comité Editor / Editors Carlos Aedo, Real Jardín Botánico, CSIC, España Inés Álvarez, Real Jardín Botánico, CSIC, España Pilar Catalán, Universidad de Zaragoza, España Ana Crespo, Universidad Complutense, España José Luis Fernández, Real Jardín Botánico, CSIC, España María Candelaria Gil, Universidad de La Laguna, España Peter E. Gibbs, University of St. Andrews, Reino Unido Josep Guarro, Universitat Rovira i Virgili, España Juan Guerra, Universidad de Murcia, España Carlos Herrera, Estación Biológica de Doñana, CSIC, España Blanca Landázuri, Real Jardín Botánico, CSIC, España María Paz Martín, Real Jardín Botánico, CSIC, España Enrique Rico, Universidad de Salamanca, España Alfonso Susanna, Instituto Botánico de Barcelona, CSIC-ICUB, España Salvador Talavera, Universidad de Sevilla, España Consejo Asesor / Associate Editors Gerardo Aymard, Universidad Unellez-Guanare, Venezuela Stephan Beck, Universidad Mayor de San Andrés, Bolivia Ricardo Callejas, Universidad de Antioquia, Colombia Santos Cirujano, Real Jardín Botánico, CSIC, España Juan A. Devesa, Universidad de Córdoba, España Javier Diéguez, Real Jardín Botánico, CSIC, España Antonio Flores, Universidad de Málaga, España Javier Francisco-Ortega, Florida International University, EE.UU. Tomás Gallardo, Universidad Complutense, España Pedro D. Jordano, Estación Biológica de Doñana, CSIC, España Joachim K. Kadereit, Johannes Gutenberg-Universität, Alemania Carlos Lado, Real Jardín Botánico, CSIC, España Ginés López, Real Jardín Botánico, CSIC, España Clodomiro Marticorena, Universidad de Concepción, Chile Ramón Morales, Real Jardín Botánico, CSIC, España Jochen Müller, Friedrich Schiller-Universität, Alemania Daniel L. Nickrent, Southern Illinois University, EE.UU. Lourdes Rico, Royal Botanic Gardens, Kew, Reino Unido A. Jonathan Shaw, Duke University, EE.UU. Jaume Terradas, Universidad Autónoma de Barcelona, España Carmen Ulloa, Missouri Botanical Garden, EE.UU. Pablo Vargas, Real Jardín Botánico, CSIC, España Mauricio Velayos, Real Jardín Botánico, CSIC, España Robert Vogt, Botanisches Museum Berlin-Dahlem, Alemania 01 primeras:01 primeras.qxd 13/06/2011 11:56 Página 4 Coordinación y Gestión Editorial Técnica: Blanca Landázuri Maquetación y Preimpresión: Bernardo Fernández y Marta Díaz Corrección: Gabriel Páez de la Cadena Textos en inglés: Bill Murphy REDACCIÓN E INTERCAMBIO Real Jardín Botánico, CSIC Plaza de Murillo, 2. E-28014 Madrid Teléfono: +34 914 203 017 Correo electrónico: [email protected] DISTRIBUCIÓN, SUSCRIPCIÓN Y VENTA Departamento de Publicaciones del CSIC Vitruvio, 8. E-28006 Madrid Teléfonos: +34 915 612 833/ +34 915 681 619/620/640 Fax: +34 915 629 634 Correo electrónico: [email protected] Librería Científica Medinaceli Duque de Medinaceli, 6 E-28014 Madrid Teléfono: +34 914 295 684 Correo electrónico: [email protected] SERVICIOS DE INFORMACIÓN Bases de datos que indizan la revista: Agricola, Agris, Aquatic Science & Fisheries Abstracts, Biosis Previews, Biosis Toxicology, Cab Abstracts, Csa Life Sciences, Abstracts, Current Contents, Environmental Sciences, Global Health, Latindex, Pascal, Redalyc, Science Citation Index Expanded y Scopus. © CSIC, 2011 Las opiniones y hechos consignados en cada artículo son de la exclusiva responsabilidad de sus autores. El Consejo Superior de Investigaciones Científicas no se hace responsable en ningún caso de la credibilidad y autenticidad de los trabajos. Los originales de la revista Anales del Jardín Botánico de Madrid, publicados en papel y en versión electrónica, son propiedad del Consejo Superior de Investigaciones Científicas, siendo necesario citar la procedencia en cualquier reproducción parcial o total. ISSN: 0211-1322 eISSN: 1988-3196 NIPO (en papel): 472-11-010-8 NIPO (en línea): 472-11-009-5 Depósito legal: M. 697-1958 Impreso en España / Printed in Spain FARESO, S. A. (Madrid) El volumen 67(2) se publicó el 23 de diciembre de 2010 The opinions and facts expressed in each article are the sole responsibility of the authors. The Consejo Superior de Investigaciones Científicas accepts no responsibility whatsoever for the credibility or authenticity of said articles. The Consejo Superior de Investigaciones Científicas is the sole proprietor of the original copies of the magazine Anales del Jardín Botánico de Madrid, published both on hard copy and digitally, requiring citation thereof in any reproduction, be it partial or entire of the works therein. 01 primeras:01 primeras.qxd 13/06/2011 11:56 Página 5 Volumen 68 N.º 1 enero-junio 2011 Madrid (España) ISSN: 0211-1322 SUMARIO / CONTENTS Zamora, N.A. El caso de Lonchocarpus costaricensis (Leguminosae, Papilionoideae), una especie endémica de Costa Rica: un complejo taxonómico-nomenclatural, y una nueva especie / The case of Lonchocarpus costaricensis (Leguminosae, Papilionoideae), an endemic species of Costa Rica: a taxonomic-nomenclatural complex, and a new species ...... Cano-Maqueda, J. & Talavera, S. A taxonomic revision of the Campanula lusitanica complex (Campanulaceae) in the Western Mediterranean region / Una revisión taxonómica del complejo Campanula lusitanica (Campanulaceae) en la región occidental mediterránea ......................................................................................................................... Venhuis, C. & Oostermeijer, J.G.B. Distinguishing colour variants of Serapias perezchiscanoi (Orchidaceae) from related taxa on the Iberian Peninsula / Distinción de variantes en color de Serapias perez-chiscanoi (Orchidaceae) en relación con táxones de la Península Ibérica ........................................................................................................ Lado, C., Wrigley de Basanta, D. & Estrada-Torres, A. Biodiversity of Myxomycetes from the Monte Desert of Argentina / Biodiversidad de Myxomycetes en el Desierto de Monte (Argentina) ..................................................................................................................... Peñas, J., Lorite, J., Alba-Sánchez, F. & Taisma, M.A. Self-incompatibility, floral parameters, and pollen characterization in the narrow endemic and threatened species Artemisia granatensis (Asteraceae) / Autoincompatibilidad, parámetros florales y caracterización de polen en la especie endémica y amenazada Artemisia granatensis (Asteraceae) .................. Schneider, A.A. & Boldrini, I.I. Microsculpture of cypselae surface of Baccharis sect. Caulopterae (Asteraceae) from Brazil / Microescultura de la superficie de las cipselas de Baccharis sect. Caulopterae (Asteraceae) de Brasil ........................................................ Domínguez-Álvarez, S., Rico, J.M. & Gil-Rodríguez, M.C. Photosynthetic response and zonation of three species of Gelidiales from Tenerife, Canary Islands / Respuestas fotosintéticas y zonación de tres especies de Gelidiales de Tenerife, Islas Canarias .............. 7/14 15/47 49/59 61/95 97/105 107/116 117/124 La Serapias perez-chiscanoi (Orchidaceae) es una orquídea endémica que se puede observar al suroeste de la Península Ibérica. En general, tiene flores verde pálido, pero ocasionalmente se pueden encontrar ejemplares con flores rosadas o rojas. [Fotografía: Caspar Venhuis]. Serapias perez-chiscanoi (Orchidaceae) is a narrow endemic orchid of the southwestern part of the Iberian Peninsula. It generally has pale green flowers, but plants with pinkish or even red flowers are occasionally found. [Photograph: Caspar Venhuis]. 01 primeras:01 primeras.qxd 13/06/2011 11:56 Página 6 2255 lonchocarpus:Maquetación 1 13/06/2011 11:57 Página 7 Anales del Jardín Botánico de Madrid Vol. 68(1): 7-14 enero-junio 2011 ISSN: 0211-1322 doi: 10.3989/ajbm.2255 El caso de Lonchocarpus costaricensis (Leguminosae, Papilionoideae), una especie endémica de Costa Rica: un complejo taxonómico-nomenclatural, y una nueva especie por Nelson A. Zamora Unidad Estratégica de Botánica, Instituto Nacional de Biodiversidad (INBio), Costa Rica. Apdo. 22-3100, Santo Domingo [email protected] Resumen Abstract Zamora, N.A. 2011. El caso de Lonchocarpus costaricensis (Leguminosae, Papilionoideae), una especie endémica de Costa Rica: un complejo taxonómico-nomenclatural, y una nueva especie. Anales Jard. Bot. Madrid 68(1): 7-14. Zamora, N.A. 2011. The case of Lonchocarpus costaricensis (Leguminosae, Papilionoideae), an endemic species of Costa Rica: a taxonomic-nomenclatural complex, and a new species. Anales Jard. Bot. Madrid 68(1): 7-14 (in Spanish). La mezcla de colecciones, la ausencia de la designación de un espécimen tipo y la fuerte similitud morfológica entre colecciones citadas en el protólogo de la descripción de Derris costaricensis generaron una situación taxonómica y nomenclatural confusa alrededor de la entidad Lonchocarpus costaricensis (Donn. Sm.) Pittier; un estudio taxonómico detallado del caso reveló que una nueva especie (Lonchocarpus felipei N. Zamora), aquí descrita, está implicada. Asimismo se restablece aquí la lectotipificación original de Lonchocarpus macrocarpus Benth. Mixed collections, a lack of type specimen designation and strong morphological similarity between collections cited in the Derris costaricensis protologue has led to a very confused taxonomic and nomenclatural situation surrounding the species Lonchocarpus costaricensis (Donn. Sm.) Pittier; a detailed taxonomic study has revealed that a new species (Lonchocarpus felipei N. Zamora) here described, is involved. Also, the original lectotipification of Lonchocarpus macrocarpus Benth., is re-establish it here. Palabras clave: Taxonomía, nomenclatura, Leguminosae, Lonchocarpus, complejo de especies, lectotipificación, una nueva especie. Keywords: Taxonomy, nomenclature, Leguminosae, Lonchocarpus, species complex, lectotypification, a new species. Introducción citados por Pittier (1917) y aquellos citados en el protólogo de Derris costaricensis determinó que Pittier 3654 (fr) y Tonduz 2880 (fr) corresponden a un taxon distinto. Este taxon posee una gran semejanza en su follaje y frutos con Lonchocarpus costaricensis (Donn. Sm.) Pittier. Debido y a partir de esta situación de mezcla de colecciones, el concepto de L. costaricensis se adoptó (p.e., Standley, 1937: 543) y se ha venido aplicando en forma equivocada. Además, ha favorecido tal confusión la condición del verdadero L. costaricensis, al ser una especie endémica de Costa Rica relativamente escasa o rara, mientras la aquí descrita (L. felipei N. Zamora) es más común y conocida ampliamente de Nicaragua y Costa Rica. En la descripción original de Derris costaricensis Donn. Sm., Bot. Gaz. 44: 110-111. 1907, el autor citó dos colecciones (síntipos), Tonduz 2880 (CR!, US!) y Tonduz 13993 (BM!, NY!, US!), que fueron utilizadas para elaborar el protólogo, sin dar indicación de la colección tipo. Más tarde, Pittier (1917) hizo la transferencia al género Lonchocarpus y designó Tonduz 13993 (fl y fr) como el lectótipo; en esta misma publicación, Pittier citó otras colecciones: Pittier 3654 (fr) (NY!, US!), y Tonduz 13528 (fl y fr) (F!, US!) y 13532 (fl y fr) (BM!, NY!, P!, US!). Un estudio, aquí detallado, de todos los ejemplares 2255 lonchocarpus:Maquetación 1 13/06/2011 11:57 Página 8 8 N.A. Zamora Sousa (1990), consciente de esta situación (mezcla de colecciones) y aunque sin describirla y estudiarla en detalle, prefirió relectotipificar el nombre L. macrocarpus Benth., para acomodar la entidad aquí definida como distinta de L. costaricensis (Donn. Sm.) Pittier. Pero L. macrocarpus es un binomio heterogéneo compuesto de tres síntipos: Fendler 1861 (Venezuela), Orbigny 578 (Bolivia) y “New Spain, Herb Pavón” [lectótipo elegido por Sousa (1990)]. Sin embargo, ya previamente Pittier (1928), en ausencia de una designación de un tipo para L. macrocarpus, había seleccionado la colección Fendler 1861 (K!) como el lectótipo; designación con la que Sousa (1990) más tarde argumentó estar en desacuerdo con Pittier, señalando que fue una “escogencia mecánica”, y propuso relectotipificar L. macrocarpus con el ejemplar Herb. Pavón s.n. (G!). El síntipo “Pavón” lleva anotado por Bentham una etiqueta “Nueva España Herb. Pavón” y marcada por Pavón “Classis 17 N 204 dubia N E. Securidaca?”. La colección se compone de dos folíolos sueltos y frutos, cuya apariencia general (en especial por sus frutos) parece corresponder más bien a L. costaricensis. La validez de la lectotipificación de L. macrocarpus hecha por Pittier había sido respaldada y aceptada por Tozzi (1989); además, a su vez este autor analizó con mayor detalle la naturaleza del concepto de L. macrocarpus, restringiéndolo al síntipo Fendler 1861 (K!), apoyando con esto la decisión de Pittier (1928). Vale mencionar también que, siguiendo este concepto del tipo [Fendler 1861], la mayoría del material recolectado de L. macrocarpus proviene del área geográfica del tipo (Venezuela). Para el síntipo Orbigny 578 (G!, P!), Tozzi (1989) no resolvió sobre su identificación; pero dicha colección fue identificada como Lonchocarpus hedyosmus Miq., por H.H. Poppendieck (HBG) en julio de 1993. También, según Tozzi (1989) y confirmado por este estudio, la colección Spruce 4597 (K!, G!, NY!, P!, W!; tipo, K!) de L. macrocarpus var. sericophyllus Benth., citada por Bentham (1860) en el protólogo de L. macrocarpus, corresponde a L. hedyosmus Miq.; identificación a su vez respaldada por H.H. Poppendieck (HBG), en julio de 1993. Finalmente, Tozzi (1989) señaló que L. hedyosmus y L. macrocarpus están altamente emparentadas. Pero en esta última las inflorescencias tienden a ser más cortas; el cáliz, denticulado, y los folíolos, persistentemente denso y sedoso-pubescentes en el envés, versus inflorescencias más alargadas, el cáliz truncado y folíolos glabrescentes o no denso sedoso-pubescentes en el envés en L. hedyosmus; también existen algunas diferencias a nivel de frutos. Basándonos en la situación antes descrita, se concluye que la nueva lectotipificación establecida por Sousa (1990) para L. macrocarpus no es válida, y considero más apropiado proponer un nombre nuevo para la especie históricamente identificada en forma equivocada como Lonchocarpus costaricensis o L. macrocarpus sensu Sousa (1990, 2001). Además, el restablecer la lectotipificación de L. macrocarpus hecha por Pittier estaría en mayor concordancia con la descripción que se provee en el protólogo original de L. macrocarpus, tanto con el tamaño del fruto, al cual el epíteto hace referencia, como en el número de folíolos (917) y área geográfica (Sudamérica) de dos de los síntipos citados [Fendler 1861(Venezuela) y Orbigny 578 (Bolivia)], excepto la colección Herb. Pavón s.n., cuya procedencia específica sigue siendo aún dudosa o sin resolver (véase McVaugh, 2000: 322). Por lo que se conserva aquí el lectótipo seleccionado por Pittier (1928): Lonchocarpus macrocarpus Benth., J. Linn. Soc., Bot. 4 (Suppl.): 91. 1860. Tipo: “Fendler 1861 (Venezuela)” (lectótipo, K!, aquí redesignado). El restablecer la lectotipificación de Pittier (1928) significa a su vez oficializar aquí, por primera vez, que la especie Lonchocarpus margaritensis Pittier es sinónimo de Lonchocarpus macrocarpus Benth (véase Tozzi, 1989; Tozzi & Silva, 2007). La especie aquí descrita es ampliamente conocida (según abundante material de herbario citado y examinado) y estudiada por ecólogos y biólogos en la Provincia de Guanacaste (Costa Rica) desde mediados de los años 1960, mientras la verdadera (endémica, algo rara y más localizada) Lonchocarpus costaricensis permanece más o menos oculta o aislada en algunas colinas del Área de Conservación Guanacaste (especialmente Sector Murciélago) y algunos cerros de formaciones calcáreas en la Península de Nicoya. Lonchocarpus felipei N. Zamora, sp. nov. Tipo: Costa Rica. Puntarenas: Cantón de Puntarenas, ridges between Río Guacimal and Río Lagarto on road from Inter American Highway to Monteverde,10°16’N, 84°50’W, 800-1000 m, 20 April 1991 (fl), Haber & Zuchowski 10656 (holótipo, INB; isótipos, CR, K, MEXU, MO, PMA,). Figs. 1, 2. Lonchocarpus costaricensis similis, sed floribus minoribus [usque ad 9-10(12) mm, non 15-18(20) mm], vinaceis vel atropurpureis (non roseis) recedit; a Lonchocarpus retifer foliolis paucioribus, fructibus latioribus differt. Árbol de 7-14 m de alto, ramitas conspicuamente lenticeladas, denso a esparcido ferrugíneo pubescentes hacia el ápice o partes jóvenes; estípulas deciduas, no vistas. Hojas imparipinnado-compuestas, con 5-7(9) folíolos; pecíolo de 5,5-11 cm de largo; ra- Anales del Jardín Botánico de Madrid 68(1): 7-14, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2255 2255 lonchocarpus:Maquetación 1 13/06/2011 11:57 Página 9 Lonchocarpus costaricensis, especie endémica de Costa Rica 9 Fig. 1. Lonchocarpus felipei: A, rama con frutos (Zamora 2281, INB); B, rama con inflorescencias; C, flor; D, estambres, E, pétalos (Haber & Zuchowski 10656, INB). Ilustración de Claudia Aragón. Anales del Jardín Botánico de Madrid 68(1): 7-14, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2255 2255 lonchocarpus:Maquetación 1 13/06/2011 11:57 Página 10 10 N.A. Zamora quis de 4-10 cm de largo, levemente caniculado, glabrescente a ferrugíneo pubescente, peciolulos 5-9 mm de largo; folíolos distales (6,5)9-21 × 3,6-10,2 cm, oblongos a obovados, mediales (5)9-14,5 × (3)3,9-7,5 cm, oblongos a obovados, basales de (4)6-11 × (2,4) 3,6-7,7 cm, ovados a oblongos o suborbiculares, ápice redondeado, base aguda, obtusa a levemente asimétrica, glabros cuando adultos o esparcido pilosos cuando jóvenes en el haz, glabrescentes a ferrugíneo pilosos en el envés, nervios secundarios 9-17 por lado, venación terciaria conspicuamente reticulada. Inflorescencias racemosas, axilares, 2-16 cm de largo, eje principal glabrescente a diminuto estrigoso o esparcido seríceo, pedúnculos o ejes laterales secundarios de 1-5 mm de largo; brácteas deciduas, no vistas, bractéolas 0,5-0,6 mm, escuamiformes, decíduas. Flores de color marrón a púrpura oscuro, 9-10(12) mm de largo; pedicelo 3,5-4 mm de largo; cáliz (1,5)2-3 mm de largo, cupuliforme, diminuto seríceo, truncado o ligeramente ondulado-denticulado; pétalos 5, estandarte 8-11(12) × 9-10 mm, suborbicular y cóncavo, con los márgenes levemente involutos, ligeramente recurvado, nervado, seríceo por fuera y glabro por dentro, con una mácula verde basal y punteado cerca de la base en la cara interna, emarginado en ápice y auriculado en la base, la uña ca. 2 mm de largo; alas 5-7 × 2,5-3(3,5) mm, oblongo-oblicuas, redondeadas en el ápice y levemente auriculadas en la base, la uña ca. 3 mm; quilla 5-6 × 3-3,2 mm, oblongo-oblicua, unida distalmente, diminuto serícea por fuera, uña ca. 2,8-3 mm; tubo estaminal 6-6,5 mm, glabro; pistilo linear, 6-7,5 mm, denso seríceo, óvulos 1(2); estilo fuertemente recurvado; estigma inconspicuo; anteras basifijas. Frutos legumbres, 7-12(16) × 3-5 cm, elípticos a ovado-elípticos u obovado-elípticos a veces falcadoelípticos, redondeados en el ápice y obtusos a atenuados en la base, coriáceos, ambos márgenes afilados, pardo-amarillentos, glabros o glabrescentes cuando adultos y pardo-amarillento sedoso pubescentes cuando jóvenes, con verrugas evidentes a la altura de las semillas; semillas 1 ó 2, 1,5-1,6 × 0,8 cm, aplanadas, oblongo-reniformes, pardo-rojizas cuando secas. A pesar de la larga confusión en la que L. costaricensis estuvo inmersa, ésta difiere de L. felipei de manera notable, como se resume en el siguiente cuadro (Tabla 1). Ambas especies poseen hojas con 5-7 folíolos y éstos son muy semejantes en su apariencia general, por lo que es algo difícil diferenciarlas en forma vegetativa, aunque las ramitas y hojas (raquis y envés de los folíolos) de L. costaricensis poseen una pubescencia tomentosa pardo-rojiza o ferrugíneo-oscura; mientras, en L. felipei la pubescencia es más amarillenta a pardo-amarillenta. A nivel de flores (Fig. 2), tamaño y color, y en cierto grado los frutos, la mayor afinidad de L. felipei es más bien con la especie L. retifer Standl. & L.O. Williams, pero en ésta última sus flores son relativamente más pequeñas (5-6 mm), las hojas normalmente tienen más folíolos (comúnmente 9) y sus frutos son más angostos (2-2,6(2,9) cm). Además, L. retifer se da en climas más húmedos y su floración ocurre cuando el árbol tiene hojas. Fig. 2. A, estandarte; B, cáliz; C, frutos de: Lonchocarpus retifer (flores, Espinoza & al. 1472, INB; frutos, Morales 5559, INB), L. felipei (flores, Haber & Zuchowski 10656, INB; frutos, Zamora 2281, INB) y L. costaricensis (flores, Zamora & al. 2243, INB; frutos, González & Garita 3052, INB). Ilustración de Claudia Aragón. Hábitat, distribución, conservación y ecología: Lonchocarpus felipei es propio de bosques secos a húmedos, a lo largo de la costa pacífica de Nicaragua (Chontales) hasta Costa Rica (Valle Central); es de frecuente a común en vegetación caducifolia o semicaducifolia, de preferencia en terreno sedimentario, Anales del Jardín Botánico de Madrid 68(1): 7-14, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2255 2255 lonchocarpus:Maquetación 1 13/06/2011 11:57 Página 11 Lonchocarpus costaricensis, especie endémica de Costa Rica 11 Tabla 1. Principales caracteres diferenciadores entre Lonchocarpus felipei y L. costaricensis. Medidas de flores para L. felipei fueron tomadas de Haber & Zuchowski 10656 (fl) (INB!) y para L. costaricensis tomadas de Zamora & al. 2243 (fl) (INB!). Carácter L. felipei Flores, color marrón o púrpura oscuro L. costaricensis rosadas Flores, tamaño (mm) 9-10(12) 15-18(20) Estandarte, mácula presente, verde ausente o no diferenciada Cáliz, tamaño (mm) (1,5)2-3 4-5 Cáliz, forma cupuliforme campanulado Cáliz, margen truncado a ondulado-denticulado dentado, el vexilar 2-3 mm Bractéolas, forma escuamiformes filiformes Bractéolas, tamaño (mm) 0,5-0,6 mm, deciduas 5-7 mm, persistentes Inflorescencia, pubescencia diminuto estrigosa o serícea esparcida denso ferrugíneo tomentosa Frutos adultos verdoso-amarillentos o pardo-amarillentos, glabros, glabrescentes a tomentosos, con verrugas a la altura de las semillas pardo-rojizos o ferrugíneo-oscuros, denso tomentosos, lisos a la altura de las semillas Hábitat bosque seco a húmedo, prefiere terreno sedimentario bosque seco, prefiere terreno cálcareo Distribución Nicaragua-Costa Rica Endémica, Costa Rica desde el nivel del mar hasta los 800(1000) m. Al menos en Costa Rica la especie se encuentra protegida y poblaciones importantes se encuentran en el Área de Conservación Guanacaste (ACG: Sector Santa Rosa, Sector Santa Elena, Sector Murciélago, Sector Pocosol), Parque Nacional Palo Verde, Refugio de Vida Silvestre Macacona-Esparza y Zona Protectora El Rodeo-San José. Su floración ocurre de febrero a abril, siendo máxima cuando el árbol está completamente caducifolio. Es interesante añadir que he observado el fenómeno del albinismo en un individuo en el Sector Santa Rosa, ACG; fenómeno también visto en Lonchocarpus cultratus (Vell.) A.M.G. Azevedo & H.C. Lima (A.M.G. Azevedo Tozzi, com. pers., 2010). Sus frutos se han observado la mayor parte del año, aunque permanecen en el árbol durante toda la estación lluviosa y maduran y caen en el primer mes del verano siguiente; cada individuo reproductivo florece un año y tiene frutos hasta el siguiente, pero no sincronizadamente entre árboles (D.H. Janzen, com. pers., 2010). Ensayos de reproducción y propagación han dado buenos resultados, con importantes porcentajes de germinación y desarrollos en vivero y campo. Es importante mencionar que toda la literatura ecológica, química y molecular (p.ej., Chapman, 1989; Evans & al., 1985; Fellows & al., 1979; Janzen, 1980, 1982, 1983, 1986; Janzen & Liesner, 1980; Janzen & al., 1990; Navarro & al., 2005; Waterman & Mahmoud, 1985) generada para la especie (o donde se cita el nombre) Lonchocarpus costaricensis corresponde más bien a la especie aquí descrita. A menudo su follaje se encuentra altamente infestado de agallas, provocadas por insectos hemípteros del género Eu- phalerus, situación frecuente que padecen varias especies de Lonchocarpus (Hollis & Martin, 1997). Etimología: Dedico esta especie en honor a su Alteza Real el Príncipe Don Felipe de Borbón, por su apoyo al estudio de la biodiversidad de Costa Rica. Colecciones examinadas COSTA RICA. Guanacaste: Cantón de La Cruz, P.N. Guanacaste, Cordillera de Guanacaste, Camino a la Estación Maritza, a orillas del Río Espavelar, 10°58’10’’N, 85°33’35’’W, 300 m, 25-V1995 (fr), Zamora 2290 (MO); Bahía Salinas a Santa Cecilia, camino a la Estación Maritza, alrededores de la Quebrada Espavelar, 10°58’10”N 85°38’40’’W, 290 m, 5-VIII-1995 (fr), Ramírez & Soto 392 (INB); Cuajiniquil, camino entre Cuajiniquil y Junquillal, 10°57’20”N, 85°42’00”W, 0 m, 8-IX-1995 (fr), J. Sánchez 534 (CR). Cantón de Liberia, Parque Nacional Santa Rosa, Llano Jicaral, hacia la playa, 280 m, 25-I-1983 (fr), Sousa & al. 12688 (MO); Parque Nacional Santa Rosa, Llano Jicaral, hacia la playa, 280 m, 25-I-1983 (fr), Sousa 12688 (CR); Parque Nacional Santa Rosa, 10 m, 4-XII-1985 (fr), Zamora & al 1152 (CR, MO); Parque Nacional Santa Rosa, bosque seco, 317 m, 3-IV-1976, Tinney 236 (CR); Parque Nacional Santa Rosa, bosque secundario, 317 m, 3-I1976, Chazdon 210 (CR); Parque Nacional Santa Rosa, entrada al mirador Valle Naranjo, 10°48’00’N, 85°38’37’’W, 200 m, 11-IV2000 (str), Acosta & al. 847 (MO); Liberia, Parque Nacional Santa Rosa, alrededor de la entrada a Nancite y playa Naranjo, 10°48’30’’N, 85°40’55’’W, 10 m, 26-IV-2000 (fr), Acosta & al. 931 (INB, MO); faja costeña del golfo de Papagayo, Hacienda Horizontes, 10°42’25’’N, 83°34’30’’W, 130 m, 1-III-1995 (st), Zamora 2241 (MO). Cantón de Carrillo, Península de Nicoya, Sardinal Nuevo Colón, 2-3 km después del cruce a Zapotal, cerro Judas, camino a playa Guacamaya, 10°29’45”N, 85°44’05’’W, 29 m, 24-V1995 (fr), Zamora & al. 2285 (INB); faja costeña del golfo de Papagayo, Sardinal alrededores playa Monte del Barco, 10°36’30’’N, 85°38’20’’W, 20 m, 3-II-1996 (fr), Jiménez & al. 2065 (INB); bahía El Coco, bahía Playa Hermosa, and Sardinal, 10°32’00”N, 85°40’00”W, 0-150 m, 10-XI-1975 (fr), Burger & Baker 9933 (CR); Anales del Jardín Botánico de Madrid 68(1): 7-14, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2255 2255 lonchocarpus:Maquetación 1 13/06/2011 11:57 Página 12 12 N.A. Zamora a b c d e Fig. 3. Lonchocarpus felipei: a, b, flores maduras, mostrando el color vivo, el estandarte reflexo y mácula verde; c, rama con inflorescencias inmaduras, con hojas; d, rama en máxima floración, sin hojas; e, tallo, con la corteza lisa. Fotografías de D. Solano (Zamora & Solano 4913, INB). Anales del Jardín Botánico de Madrid 68(1): 7-14, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2255 2255 lonchocarpus:Maquetación 1 13/06/2011 11:57 Página 13 Lonchocarpus costaricensis, especie endémica de Costa Rica cerro el Hacha, 28-VII-1986 (fr), Chacón & Chacón 2038 (CR, MO); faja costeña del golfo de Papagayo, camino entre Nuevo Colón y Zapotal, cerca del cerro Matapalo. 10°30’30”N 85°46’20’’W, 300 m, 4-VIII-1995 (fr), Ramírez & Soto 384 (INB), 386 (INB). Cantón Santa Cruz, 4 km E. Santa Cruz, 130 m, XII1973, Solomon 667, (CR); Cañas, La Pacífica, 24-VIII-1990 (fr), Jiménez 54 (CR). Cantón de Bagaces, Parque Nacional Palo Verde, valle del Tempisque, cerro Jocote, sector Carreta, 10°22’45”N, 85°19’15’’W, 0-100 m, 11-VI-1993 (fr), Chavarría 822 (INB, CR); Parque Nacional Palo Verde, Estación Catalina, sendero Botija, saliendo al Cenicero, 10°21’N, 85°16’W, 10-20 m, 9-XII-1991 (fr), Chavarría 450 (INB, CR, MO); R.B. Lomas Barbudal, valle del Tempisque; Lomas Barbudal, Marañonal, Oja de Agua y Agua Fría, 10°26’25”N, 85°19’05’’W, 100-200 m, 2-V-1993 (fr), Chavarría 797 (INB); Cuenca del Temspisque, Hacienda Monteverde, 10°33’00’’N, 85°18’20’’W, 100-300 m, 9-VI-1996 (fl), Ronchi & Frankie 830 (INB); Cuenca del Tempisque, 3,5 km del cruce al Parque, sobre la carretera Interamericana, 10°29’50’’N, 85°15’40’’W, 70 m, 23-XI-2000 (fr), Acosta & al. 3009 (INB); Refugio de Vida Silvestre-Palo Verde, sin fecha (fl), Ramírez 220 (CR); finca La Pacífica, 2 miles N of Cañas along Pan-Am Hwy., gallery forest between río Corobicí and irrigation ditch, 5-VI-1971 (fr), Gentry 810 (CR, MO); Lomas Barbudal, Bagaces 100 m, 10V-1984 (fr), Gómez 23012 (CR, MO); road from Bagaces to Aguas Claras about 5 km North of Bagaces, 150-200 m, 7-VII-1976 (fr), Utley 5305 (MO, CR). Cantón de Cañas, Hacienda La Pacífica near Cañas, 14-VIII-1986 (fr), Seigler 12774 (MO), 12401A (MO); Comelco Ranch, Bagaces (MO), remnant forest along stream, 2 km southwest of La Cruz, 11°4’ N, 85°40’W, 10 m, 29-I-1978 (fr), Liesner 4635 (CR, MO); seasonal swamp, dry now, OTS Área A3 site a Comelco, 31 March 1972 (fl), Stone & Opler 3157 (MO); Comelco Ranch, 10°20-35’N, 85°18-25’W, V-1970 (fr), Hartshorn 910 (MO); Palo Verde, 4-VI-1969 (fr): valle del Tempisque, cerca del río Lajas, 174-175, 600 m antes de la Escuela de Buenos Aires, 10°19’40”N, 85°03’10’’W, 120 m, 3-VIII-1995 (fr), Ramírez & Soto 383 (INB). Cantón de Abangares, valle del Tempisque, Abangares, orillas de la carretera Interamericana ruta 1, 173-74, 1 km después del río Lajas, 10°19’10’’N, 85°02’50’’W, 100 m, 20-IV1991 (fr), Zamora 2281 (MO); valle del Tempisque, orillas de la carretera Interamericana, ruta 1, km 173-174, entre Lourdes y río Lajas, 10°19’10’’N, 85°02’50’’W, 100 m, 28-II-1995 (fl), Zamora & Mora 2239 (INB). Puntarenas: Cantón de Puntarenas, Hacienda Santa Marta, Cascajal, along ditch near Cascajal Station (25 km ESE of Puntarenas), 2-VI-1949 (fr), Holm & Iltis 221 (BM, MO, U); ridges between río Guacimal and río Lagarto on road from Inter American Highway to Monteverde, 10°16’N, 84°50’W, 8001000 m, 20-IV-1991 (fls), Haber & Zuchowski 10653 (INB, CR), 10656 (INB, CR); Monteverde, valle del río Guacimal, Lindora, vertiente pacífica, 10°18’N, 84°50’ W, 1000 m, 17-VI-1998 (fr), Bello 402 (CR, MO); Monteverde, road from Santa Elena to village of San Luis and Lagarto, Pacific slope, moist forest, 10°16’N, 84°50’W, 750-900 m, 10-VII-1990 (fr), Haber 9987 (CR, INB, MO); Santa Elena to Coyolar de Guacimal, Pacific Slope, roadside and remnant forest patches of dry forest, 10°15’N, 84°51’W, 250900 m, 16-VII-1988 (fr), Hammel 17111 (CR). R.N.A. Cabo Blanco, estación Cabo Blanco, bosque primario y secundario, 9°35’N, 85°06’W, 20-100 m, 4-XI-1991 (fr), Chavarría 295 (INB, CR, MO); R.N.A. Cabo Blanco, Camaronal, sendero el Barco, 9°34’42’’N, 85°08’10’’W, 0 m, 25-VI-2001 (fr), Chavarría & al. 2183 (INB); isla San Lucas, golfo de Nicoya, 9°57’N, 84°54’W, 0-400 m, 20-X-1984 (fr), Grayum & al. 4236 (MO); R.N.A. Cabo Blanco, península de Nicoya, sendero a El Atracadero de San Miguel, 9°35’00’’N,85°07’00’’W, 1-300 m, 17-XII-1993 (fr), Fernán- 13 dez & al. 1300 (INB); golfo de Nicoya, isla San Lucas, entre playa Cocos y playa Bellavista, 9°57’18’’N, 84°54’11’’W, 20 m, 17-III2005 (fl), Soto & González 539 (INB); Punta Morales, 1-3 m, 19VII-1984 (fr), Gómez-Laurito 10071 (CR). San José: Cantón de San José, Villa Colón, 800 m, 19-I-1972 (fr), Caffrey 86 (CR); vicinity of Villa Colón, 15-II-1965 (fl), Godfrey 66479 (MO). Cantón de Mora, Zona Protectora El Rodeo, camino que desciende de la Universidad a La Paz hacia el río Jaris, pequeños bosquetes a orilla del camino junto a potreros y cultivos, 9°54’00”N, 84°16’00”W, 500-1000 m, 8-VII-1996 (fr), Cascante 1040 (CR); Z.P. El Rodeo, valle del Tárcoles, bajo morales, 9°55’00’’N, 84°16’00’’W, 8001000 m, 08-VII-1995 (st), Jiménez & Ramírez 1886 (MO). Cantón de Acosta, valle del Candelaria, cuenca del río Candelaria, parches remanentes y potreros cerca del puente, 9°46’50’’N, 84°11’43’’W, 700 m, 19-XI-1994 (fr), Morales 3155 (INB, CR, MO); Cantón de Orotina, N outskirts of Orotina, 9°55’N, 84°32’W, 200 m, 6-IV1983 (fr), Judziewicz 4559 (CR). NICARAGUA. Chontales: 0,9 km NE of Hwy 7 on road to Comalapa, ca. 12°10’N, 85°33’W, 160 m, 12-VI-1982 (fr), Stevens & al. 21570 (MO). Granada: km 75, carretera Sur, 8 km de Nandaime, Llanos el Dorado, 11°41’N, 86°00’W, 70 m, 25-I-1984 (fr), Moreno & Stevens 22855 (MO); camino de Casa Tejas, 1,2 km antes de la finca San José del Mombacho, 11°46’N, 85°54’W, 40-60 m, 21-VI-1982 (fr), Moreno 16644 (MO); camino a Charco Muerto, 3 km al E de Casa de Tejas, sobre el camino, 100-200 m, 5-VI1980 (fr), Araquistain & Moreno 2855 (MO). Managua: carretera a Montelimar, comarca Aduana, al N del río Aduana, 80-100 m, 21VII-1980 (fr), Guzmán & al. 410 (MO). Rivas: along road SE from San Juan del Sur, 3-4 km NW of río La Flor, playa El Coco, quebrada El Coco, 11°09’N, 85°47’W, 0-95 m, 17-XII-1977 (fr), Stevens 5492 (MO). Agradecimientos Deseo expresar un profundo agradecimiento a mis colegas Claudia Aragón, por la preparación de las ilustraciones, y Henk van der Werff, por su asistencia en la elaboración de la diagnosis en latín. A Michael H. Grayum, A.M.G.A. Tozzi, Lourdes Rico, D.H. Janzen y los revisores de esta revista, por sus valiosos aportes, comentarios y sugerencias en una versión preliminar de este artículo. Además, a todos los herbarios citados, en especial aquellos que albergan colecciones históricas, por facilitarme acceso y ayuda para el estudio de las mismas. Esta investigación fue posible gracias al convenio de cooperación entre el Ministerio de Ambiente, Energía y Telecomunicaciones (MINAET) y el Instituto Nacional de Biodiversidad (INBio), al apoyo económico de particulares y a la red de especialistas taxónomos que contribuyen a completar el Inventario Nacional de Biodiversidad en Costa Rica. Referencias bibliográficas Bentham, G. 1860. Synopsis of Dalbergieae, a tribe of Leguminosae. Journal of Linnean Society Botany 4 (Suppl.): 1-28. Chapman, C.A. 1989. Primate Seed Dispersal: The Fate of Dispersed Seeds. Biotropica 21(2): 148-154. Evans, S. , Fellows, L.E., Janzen D.H., Chambers, J., & Hider, R.C. 1985. Erythro-gamma-hydroxyhomo-L-arginine: an amino acid from seed of Lonchocarpus costaricensis, and its preferential interaction with borate. Photochemistry 24: 12891292. Anales del Jardín Botánico de Madrid 68(1): 7-14, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2255 2255 lonchocarpus:Maquetación 1 13/06/2011 11:57 Página 14 14 N.A. Zamora Fellows, L.E., Bell, E.A., Lee, T.S. & Janzen, D.H. 1979. Tetrahydrolathyrine; a new amino acid from seeds of Lonchocarpus costaricensis. Phytochemistry 18: 1333-1335. Hollis, D. & J.H. Martin. 1997. Jumping plantlice (Insecta: Hemiptera) attacking Lonchocarpus species (Leguminosae), including “Black Cabbage Bark”, in Belize. Journal of Natural History 31: 237-267. Janzen, D.H. 1980. Specificity of seed-attacking beetles in a Costa Rican deciduous forest. Journal of Ecology 68: 929-952. Janzen, D.H. 1982. Weight of seeds in 1-3 seeded fruits of Lonchocarpus costaricensis (Leguminosae), a Costa Rican winddispersed tree. Brenesia 19/20: 363-368. Janzen, D.H. 1983. Costa Rican Natural History. University of Chicago Press, Chicago. 816 pp. Janzen, D.H. 1986. Mice, big mammals, and seeds: it matters who defecates what where. In: Estrada, A. & Fleming, T.H. (eds), Frugivores and seed dispersal. Pp. 251-271. Dr. W. Junk Publishers, Dordrecht. Janzen, D.H. & Liesner, R. 1980. Annotated check-list of plants of lowland Guanacaste Province, Costa Rica, exclusive of grasses and non-vascular cryptogams. Brenesia 18: 15-90. Janzen, D.H., Fellows, L.E. & Waterman, P.G. 1990. What protects Lonchocarpus (Leguminosae) seeds in a Costa Rican dry forest? Biotropica 22: 272-285. McVaugh, R. 2000. Botanical Results of the Sessé & Mociño Expedition (1787-1803). VII. A Guide to Relevant Scientific Names of Plants. Hunt Institute for Botanical Documentation. Carnegie Mellon University, Pittsburgh. Navarro, C., S. Cavers, N. Colpaert, G. Hernández, P. Breyne & A.J. Lowe. 2005. Chloroplast and Total Genomic Diversity in the Endemic Costa Rican Tree Lonchocarpus costaricensis (J. Donn. Smith) Pittier (Papilionaceae). Silvae Geniticae 54(6): 293-300. Pittier, H. 1917. The Middle American species of Lonchocarpus. Contributions from the United States National Herbarium 20: 37-93. Pittier, H. 1928. Contribuciones a la dendrología de Venezuela. Árboles y arbustos del orden de las Leguminosas. III-Papilionáceas. Trab. Mus. Com. Venezuela (Bol. Minist. R. R. E. E. n.º 4-7) 4: 179-259. Sousa, M. 1990. Adiciones a las Papilionadas de la flora de Nicaragua y una nueva combinación para Oaxaca, México. Annals of the Missouri Botanical Garden 77: 573-577. Sousa, M. 2001. Lonchocarpus. In: Stevens, W.D., Ulloa, C., Pool, A. & Montiel, O. (eds.), Flora de Nicaragua. Monographs in Systematic Botany from the Missouri Botanical Garden 85(2): 1-2666. Pp. 1017-1028. Standley. P. 1937-1938. Flora of Costa Rica. Field Museum of Natural History. Botanical Series 18: 1-1571. Tozzi, A.M.G.A. 1989. Estudos taxonômicos dos gêneros Lonchocarpus Kunth e Deguelia Aubl. no Brasil. Tesis doctoral. Universidade Estadual Campinas, Campinas. Tozzi, A.M.G.A. & Silva, M.J. 2007. Sinonimizações em Lonchocarpus Kunth (Leguminosae-Papilionoideae-Millettieae). Rodriguesia 58(2): 275-282. Waterman, P.G. & Mahmoud, E.N. 1985. Flavonoids from the seeds of Lonchocarpus costaricensis. Photochemistry 24(3): 571-574. Editor asociado: L. Rico Recibido: 17-III-2010 Aceptado: 1-XI-2010 Anales del Jardín Botánico de Madrid 68(1): 7-14, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2255 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 15 Anales del Jardín Botánico de Madrid Vol. 68(1): 15-47 enero-junio 2011 ISSN: 0211-1322 doi: 10.3989/ajbm.2274 A taxonomic revision of the Campanula lusitanica complex (Campanulaceae) in the Western Mediterranean region by Jara Cano-Maqueda & Salvador Talavera Departamento de Biología Vegetal y Ecología, Universidad de Sevilla, E-41080 Sevilla, Spain. Corresponding author: [email protected] Abstract Resumen Cano-Maqueda, J. & Talavera, S. 2011. A taxonomic revision of the Campanula lusitanica complex (Campanulaceae) in the Western Mediterranean region. Anales Jard. Bot. Madrid 68(1): 15-47. Cano-Maqueda, J. & Talavera, S. 2011. Una revisión taxonómica del complejo Campanula lusitanica (Campanulaceae) en la región occidental mediterránea. Anales Jard. Bot. Madrid 68(1): 15-47 (en inglés). The systematics of the annual species of the Campanula lusitanica complex in the Western Mediterranean is reviewed using ITS sequences, karyology and morphology of all species of the complex. The information provided by these three sources is consistent, and the species are reorganized into two groups with the rank of section: Campanula sect. Rapunculus Boiss. and Campanula sect. Decumbentes, the latter described as new in this work. These sections comprise two well-defined subclades in the phylogenetic analyses. Sect. Rapunculus is composed, in the W Mediterranean region, by C. lusitanica L. and C. matritensis A. DC., both with 2n = 18 chromosomes, and C. cabezudoi Cano-Maqueda & Talavera, C. specularioides Coss., C. transtagana R. Fern., and C. broussonetiana Schult., all with 2n = 20 chromosomes. In the Iberian Peninsula, Sect. Decumbentes comprises two endemic species, C. decumbens A. DC. with 2n = 32 chromosomes and C. dieckii Lange with 2n = 28 chromosomes. In C. decumbens a new subspecies is described: C. decumbens subsp. baetica Cano-Maqueda & Talavera, which occurs in the Guadalquivir valley. In the formal systematic part we provide a key to identify these annual species of the Western Mediterranean, with a description and typification, photographs of flowers and fruits, distribution maps, and comments on the habitat for each taxon. En este trabajo se revisa la sistemática de las especies anuales del complejo Campanula lusitanica en el occidente del Mediterráneo usando secuencias ITS, cariología y análisis de los caracteres morfológicos de todas las especies del complejo. La información proporcionada por estas tres fuentes de caracteres es congruente, y las distintas especies se reorganizan en dos grupos principales con categoría de sección, que se corresponden con los dos subclados bien definidos: Campanula sect. Rapunculus Boiss. y Campanula sect. Decumbentes, descrita como nueva en este trabajo. En el Mediterráneo occidental, la sect. Rapunculus está formada por las siguientes especies anuales: C. lusitanica L. y C. matritensis A. DC., con 2n = 18 cromosomas, y C. cabezudoi Cano-Maqueda & Talavera, C. specularioides Coss., C. transtagana R. Fern. y C. broussonetiana Schult., con 2n = 20 cromosomas. En la Península Ibérica la sect. Decumbentes está representada por C. decumbens A. DC., con 2n = 32 cromosomas, y C. dieckii Lange, con 2n = 28 cromosomas, ambas endémicas de la Península Ibérica; de C. decumbens se describe una subespecie nueva: C. decumbens subsp. baetica Cano-Maqueda & Talavera, taxon del valle del Guadalquivir muy bien diferenciado morfológicamente de la subsp. decumbens. En la parte sistemática se proporciona una clave para la identificación de estas especies anuales del oeste del Mediterráneo, así como una descripción y tipificación, fotografías de las flores y frutos, mapas de distribución, y aspectos de la ecología para cada uno de los táxones. Keywords: phylogeny, rn DNA ITS, karyology, systematic, typification. Palabras clave: filogenia, secuencias ITS, cariología, sistemática, tipificación. Introduction there are also some annual herbs, the latter mainly in the Mediterranean. This genus has a high morphological complexity that is reflected in the different classifications that have been proposed. Based mainly on the morphology of the calyx, A. de The genus Campanula comprises about 420 species distributed mainly in temperate regions of the Northern Hemisphere (Lammers, 2007a, b). They are usually perennial herbs, although some are shrubby, and 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 16 16 J. Cano-Maqueda & S. Talavera Candolle (1830, 1838) divided the genus Campanula into two sections: sect. Medium A. DC., with calycine appendages, and sect. Eucodon A. DC., without them. This classification was followed by most nineteenth century authors (e.g. Endlicher, 1838; Willkomm, 1868; Bentham & Hooker, 1876; Rouy, 1908). In contrast, Boissier (1875) divided the genus into two sections based on fruit dehiscence: sect. Medium A. DC., with capsules opening by basal pores or valves, and sect. Rapunculus Boiss., with apical or middle position pores or valves. This classification was followed with some modifications by Fedorov (1957, 1976). Perhaps the taxonomic treatment with most insight was that by Damboldt (1976) during the preparation of the Flora of Turkey. He divided the genus Campanula into six subgenera: Campanula, Megalocalyx Damboldt, Sicyocodon (Feer) Damboldt, Roucela (Dumort.) Damboldt, Brachycodonia (Fedorov) Damboldt and Rapunculus (Boiss.) Kharadze. The subgenus Campanula includes plants with 3 or 5 stigmas, with or without calycine appendages, and with the capsules opening by basal or middle position pores, or indehiscent. This is the most complex subgenus, with 12-15 recognized sections and it contains most of the species of the genus. According to Sáez & Aldasoro (2003), subgenera Megalocalyx and Sicyocodon are annual herbs with flowers with calycine appendages and with capsules opening by basal valves; the two subgenera differ in the size of style: very long (exceeding 35 mm) in the subgen. Sicyocodon, and short (less than 15 mm) in the subgen. Megalocalyx. The around 15 species of these two subgenera are distributed in the Mediterranean Basin, SW Asia and Macaronesia. The subgenus Roucela comprises annual plants, with flowers without calycine appendages and capsules opening by basal valves; it consists of 5 species distributed throughtout the E Mediterranean and SW Asia, with the exception of C. erinus L. which has a wider distribution. The subgenus Brachycodonia comprises only C. fastigiata Dufour, an annual plant with axillary inconspicuous flowers, without calycine appendages, and with capsules opening by three apical valves; C. fastigiata occurs on gypsum soils in Spain, N Africa, C Asia and Transcaucasia. The subgenus Rapunculus includes annual and perennial plants, without calycine appendages, with a large campanulate or infundibuliform corolla, and with capsules opening by apical or middle position pores. Damboldt (1976, 1978) divided this subgenus in three sections: Pterophyllum Damboldt, Alaria Damboldt and Rapunculus. Sect. Pterophyllum includes three species: C. primulifolia Brot. (Iberian Peninsula), C. alata Desf. (Algeria and Morocco), and C. peregrina L. (E Mediterranean, including Cyprus, Lebanon and SW Turkey and S Anatolia); they are hispid perennial plants with large infundibuliform flowers, arranged in spikes or panicles, with capsules opening by three pores of middle position, and with winged seeds. Sect. Alaria contains a single species, C. pterocaula Hausskn., endemic to N and C Anatolia (Turkey); it is a biennial glabrous plant, with winged stems, large flowers arranged in spiciform inflorescences, and capsules opening by apical pores. Sect. Rapunculus contains around 50 species distributed mainly in the Mediterranean region; it comprises perennial, biennial or annual plants, glabrous or tomentose, with wingless stems, the inflorescence a panicle, or sometimes spiciform, the capsules opening by apical or middle position pores, and wingless seeds. Recent molecular phylogenies (Eddie & al., 2003; Park & al., 2006; Roquet & al., 2008, 2009; Cellinese & al., 2009; Borsch & al., 2009; Haberle & al., 2009; Stefanović & Lakušić, 2009) have revealed that the genus Campanula is paraphyletic. Studies by Roquet & al. (2008) using ITS sequences, show that Campanula sect. Pterophyllum together with the genera Musschia Dumort. and Gadellia Schulkina form a clade (Musschia clade) that is sister to the Campanula core. The Campanula core consists of two subclades: one formed by the subgenera Campanula, Megalocalyx and Roucela, together with other genera of Campanulaceae; and a second subclade comprising subgenus Rapunculus (sections Rapunculus and Alaria) and subgenus Brachycodonia, again with some other genera of Campanulaceae. The annual species of sect. Rapunculus sensu Damboldt have two centers of diversity: the Iberian Peninsula and W Morocco, and the E Mediterranean (Greece and Turkey). The annual species of the Iberian Peninsula (which include many species names, viz. C. lusitanica L., C. broussonetiana Schult., C. transtagana R. Fern., C. matritensis A. DC., C. decumbens A. DC., C. specularioides Coss. C. cabezudoi CanoMaqueda & Talavera and C. diekii Lange) have received diverse treatments in recent taxonomic studies, i.e as varieties of Campanula patula L. (Pau, 1921; Cuatrecasas, 1929), as subspecies or varieties of C. lusitanica L. (Pau, 1924; Sáez & Aldasoro, 2001), or as varieties of C. decumbens A. DC. (López-González, 1979-1980). In all cases most taxa have been relegated to infraspecific status. However, our detailed morpho-geographical analysis of the exsiccata available for this group (which we call the Campanula lusitanica complex) lead us to con- Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 17 Taxonomic review of the Campanula lusitanica complex clude that not only are many of the species described by various authors valid, but that several new taxa can be recognised. Moreover, our analyses (Cano-Maqueda & al., 2008 and the present study) based on combined analysis of ITS sequences and trnT-L using samples from most of the annual species of the Western Mediterranean area not only support recognition of these species, but have also revealed that the annual species of sect. Rapunculus sensu Damboldt comprise a polyphyletic assemblage. In this study we present a formal taxonomic revision of the Campanula lusitanica complex from the Iberian Peninsula and Morocco, based on our studies of exsiccata, karylogical parameters, and molecular markers. And using information provided by the nrDNA ITS for a large number of Campanula species and allied taxa we have also attempted to place the species of Campanula sect. Rapunculus sensu Damboldt within the context of the ongoing molecular systematics of the genus Campanula. Materials and methods Molecular Phylogeny Fresh material and herbarium vouchers of seven samples of the Campanula lusitanica complex (four of C. lusitanica s.str., one of C. decumbens, one of C. sparsa Friv., and one sample of C. ramosissima Sibth. & Sm.), two samples of C. primulifolia and one of Walhenbergia hederacea L., were sequenced for this molecular survey. Additionally, 100 sequences were taken from GenBank (Eddie & al., 2003; Susanna & al., 2006; Roquet & al., 2008, 2009; Cano-Maqueda & al., 2008; Park & al., 2006). The list of taxa, with locality, herbarium vouchers or collector’s numbers, authorities and GenBank accession numbers is shown in the Appendix I. Genomic DNA was extracted from silica-gel-dried leaves collected in the field and from herbarium material using the DNeasy Plant Mini Kit (Qiagen) following the protocols provided by the manufacturer. Amplification of the ribosomal ITS region (ITS15.8S-ITS2) was performed in 25 µL reaction volume with 22.5 µL Thermo-Start ReddyMix Master Mix, 0.5 µL of each primer, 1 µL of DMSO (100%) and 0.5 µL of DNA. Forward ITS5 and reverse ITS4 primers (White & al., 1990) were used in the amplification and sequencing processes. The polymerase chain reaction (PCR) sequence profile was one cycle of 1 min at 96 ºC, followed by 35 amplification cycles of 10 s denaturing step at 96 ºC, 5 s annealing at 50 ºC, and 3 min elongation step at 60 ºC, plus an ending cycle of 8 min at 72 ºC. Amplified products were purified using the QIAquick PCR Purification Kit (Qiagen) according to 17 the manufacturers protocols. Purified products were sequenced in both directions. The sequence PCR profile was of a time of incubation of 15 min at 37 ºC and 15 min at 80 ºC. The direct and reverse sequences of each sample were compared and corrected using the program Geneious 4.8.3, obtaining the respective consensus sequence. Sequences were aligned using the algorithm of the program ClustalX, and then adjusted manually using the options of the program Se-Al v. 1.0 alpha 1 (Rambaud, 1996). Gap indels were coded as binary characters by their presence/absence (0/1 matrix). Only those gaps that were unambiguous and potentially informative (Torrecilla & Catalán, 2002) were added to their correspondent sequence matrix and used for parsimony-based analysis. Phylogenetic analyses The phylogenetic analyses were based on parsimony and Bayesian inference searches, which were respectively conducted with PAUP* v. 4.0 beta 10 (Swofford, 2002) and MRBAY ES v. 3.0 (Huelsenbeck & Ronquist, 2002) using Galactites tomentosa Moench. (Compositae) to root the trees. In the parsimony analysis, the data matrix was subjected to two heuristic searches (first search: closest, TBR, MULTIPARS ON; second search: randomorder-entry of 10,000 replicates, TBR, MULTIPARS OFF, saving no more than 10 trees of score > 10 per replicate) aimed at finding different putative islands of most-parsimonious trees. Bootstrap support for the best trees found under the parsimony criterion was estimated by heuristic search with 1,000 bootstrap replicates (Felsenstein, 1985) using the TBR and MULTIPARS OFF strategy proposed by DeBry and Olmstead (2000) to reduce the tree-search effort in bootstrap resampling analysis. Initial MaxTrees setting was 300,000 with an auto-increase of 100. Previous to the Bayesian inference search, 24 models of nucleotide substitution were tested for which the optimal model GTR + G + I. The Bayesian analysis was performed through 1,100,000 generations using the Markov chain Monte Carlo (MCMC), sampling trees every 100 generations and burn-in all sampled point from generations previous to convergence to a stable likelihood value (Huelsenbeck & Ronquist, 2002; Leaché & Reeder, 2002). From each search, a 50% majority-rule consensus tree that showed de posterior-probability values of branches was constructed. Karyological study Plants or seeds of each taxon of the Campanula lusitanica complex sensu Cano-Maqueda & al. (2008), Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 18 18 J. Cano-Maqueda & S. Talavera were collected from natural populations (Table 1) and cultivated in the greenhouses of the University of Seville. Chromosomes were observed from meristematic cells of root tips or meiosis in the anthers of flower buds. The root tips were treated with 8-hidroxiquinoleine 0,002M for three hours and a half at 4 ° C. Subsequently, roots and flower buds were fixed with Carnoy solution (3:1 ethanol 96%: glacial acetic acid) for a minimum of 24 hours. Staining of chromosomes was performed with-alcoholic-hydrochloric carmine (Snow, 1963). The images were taken with a Leica DC 300 inserted in a Zeiss Axiophot microscope with Plan-apochromatic objective 63/1.4 and an increase of 1.25. Levan & al. (1964) were followed for the morphological terminology and Stebbins (1938) for size terminology of the chromosomes. Systematic Treatment A morpho-geographical analysis of herbarium specimens of all relevant taxa associated with the C. lusitanica complex was performed, based on the following Herbaria: C, COI, FCO, G, HVR, LISE, LISU, MA, MGC, P, SALA, SALAF, SEV, W. All recognized species and most heterotypic synonyms have been typified. The initials of the provinces of Spain and Portugal which are cited in the description of each taxon follow those used in “Flora iberica”. Results and discussion Molecular Phylogeny The ITS region comprised 726 aligned nucleotide positions of which 469 were variable and 368 were parsimony informative. The first heuristic search found 101,914,528 equally parsimonious trees that were 2,480 steps long, with a consistency index of 0.353, excluding uniformative characters, and a retention index of 0.748. The second search did not find any other island of most-parsimonious trees, and trees from the first search were used to compute the strict consensus tree. The Bayesian analysis sampled 7,639 trees, which reached a stable likelihood value after burn-in 1,500 trees. The 50% majority-rule consensus tree of all sampled trees showed a topology totally concordant with that recovered from the parsimony analysis. The Bayesian tree was better resolved than the parsimony-based tree, so only the Bayesian tree with both bootstrap and posterior probability support values for branches is shown in Fig. 1. Topology within the ITS tree is consistent with results obtained in previous phylogenetic studies of the genus Campanula and allies (Eddie & al., 2003; Park & al., 2006; Roquet & al., 2008, 2009; Cellinese & al., 2009; Haberle & al., 2009; Stefanović & Lakušić, 2009). These show that the genus is paraphyletic and divided into two major clades, the ‘Campanula s.str.’ clade and the ‘Rapunculus’ clade, plus two small clades: the ‘Musschia’ clade (99% bootstrap support, BS; 100% posterior probability support, PPS), that includes Musschia Dumort., Gadellia Schulkina, Campanula peregrina and Campanula primulifolia; and the ‘Platycodon’ clade (79%BS; 100%PPS), that includes Codonopsis Wall., Cyananthus Wall., Canarina L. and Leptocodon Hook. The genus Jasione L. is sister to Campanula s. l. The ‘Campanula s. str.’ clade has good support (72%BS; 100%PPS) although it Table 1. Chromosome numbers for annual species of the Campanula luisitanica complex in the W Mediterranean area. Origin of material, gametic (n) or somatic (2n) chromosome number of and authors who have studied the different annual species in this complex. Taxa C. lusitanica Studied material Rivadavia (Orense, Spain) Serra de Lousã (Beira Litoral, Portugal) C. matritensis Hervás (Cáceres, Spain) Hinojos (Huelva, Spain) Coimbra (Beira Litoral) C. cabezudoi Venta de Zafarraya (Granada, Spain) C. specularioides C. transtagana C. broussonetiana Ubrique (Cádiz, Spain) Benaocaz (Cádiz, Spain) Montejaque (Málaga, Spain) Valverde (Huelva, Spain) Vila Velha de Rodão (Beira Baixa, Portugal) Don Benito (Badajoz, Spain) n 2n Authors 9 18 18 In this work Fernandes (1962) 18 18 Fernández et al. (2001) (as C. lusitanica) In this work Larsen (1954) (as C. loefingii) 9 10 10 Jbel Tazzeka (Taza, Morocco) C. decumbens Benaoján (Málaga, Spain) Villamartín (Cádiz, Spain) C. dieckii Alfarnate (Málaga, Spain) 20 In this work 20 García-Martín & Silvestre (1985) Gallego (1986) In this work 10 10 20 20 In this work Fernandes (1962) Gallego (1986) (as C. lusitanica subsp. transtagana) 20 In this work 16 16 In this work In this work 28 In this work Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 19 Taxonomic review of the Campanula lusitanica complex 19 Fig. 1. Bayesian 50% MR concensus tree topology of Campanulaceae ITS. Bootstrap and posterior probability values are indicated on corresponding branches. Symbols indicate the habit of the species: ●, annual; ⽧, biennial; 䊱, perennial. The two sections that are the focus of this article are marked in gray. The chromosome numbers shown for the species that are not are specified in the text, derive from indices of plant chromosome number (Castroviejo, S. & Valdés-Bermejo, E. (eds.). 1991. Archivos de Flora iberica I: números cromosomáticos de plantas vasculares ibéricas. CSIC. Madrid; Index to plant chromosome numbers. Monographs in Systematic Botany from the Missouri Botanical Garden [www.tropicos. org]; Moore, D.M. 1982. Flora Europaea. Checklist and chromosome index. Cambridge University press). Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 20 20 J. Cano-Maqueda & S. Talavera showed little internal resolution. This clade includes species of Campanula subgenera Campanula and Roucela and the genera Diosphaera Buser, Azorina Watson, Edraianthus A. DC., Michauxia L’Hér., Trachelium L., Symphyandra A. DC., and Feeria Buser. The two genera Heterocodon Nutt. and Githopsis Nutt. form a small clade, (the ‘Githopsis’ clade, 99%BS; 100%PPS) that appears between the ‘Campanula s. str.’ and ‘Rapunculus’ clades. The ‘Rapunculus’ clade forms a monophyletic and strongly supported group (100%,BS; 100%PPS), and as with the ‘Campanula s. str.’ clade, it includes taxa treated as separate genera [Hanabusaya Nakai, Adenophora Fisch., Legousia Durande, Triodanis Rafin., Campanulastrum Small, Asyneuma Griseb. & Schenk, Phyteuma L., Physoplexis (Endl.) Schur., Petromarula R. Hedw.] in addition to many species of Campanula. It comprises five principal well-resolved subclades: one subclade (100%BS; 100%PPS) corresponds to the ‘garganica’ subclade of Park & al. (2006); two subclades, ‘rotundifolia’ and ‘pulla’ subclade (81%BS; 100%PPS and 51%BS; 93%PPS) include the isophyllous and heterophyllous species of Park & al. (2006); And the other two subclades both include elements of the Campanula lusitanica complex of Cano-Maqueda & al. (2008) which we have called the ‘decumbens’ subclade and the ‘lusitanica’ subclade. The ‘decumbens’ subclade (67%PPS) is formed by C. ramosissima and C. hawkinsiana Hausskn. & Heldr., that are sister to C. decumbens and C. dieckii Lange. The ‘lusitanica’ subclade (100%BS; 100%PPS) corresponds to the ‘lusitanica’ lineage of Cano-Maqueda & al. (2008). In this subclade, C. patula, C. sparsa and C. rapunculus L. are sister to C. matritensis A. DC., C. lusitanica s. s., C. cabezudoi CanoMaqueda & Talavera, C. specularioides Coss., C. transtagana R. Fern. and C. broussonetiana Schult. The four samples of C. lusitanica s.str. (from three different populations, see Appendix I) collapse into a moderately supported subclade (66%BS; 77%PPS) and they appear as sister to C. matritensis in a well supported clade (100%BS; 100%PPS). Moreover, C. lusitanica s.str. presents morphological differences with C. matritensis, and it has a more restricted geographical distribution than the latter (see systematic treatment), so we have treated these two as separate species. Karyological study All species of ‘lusitanica’ subclade have 2n = 20 or n = 10 chromosomes, except C. matritensis and C. lusitanica with 2n = 18 or n = 9. The taxa of ‘decumbens’ subclade, C. decumbens and C. dieckii, have 2n = 32 or n = 16 and 2n = 28 chromosomes, respectively (Table 1). All species studied have small or moderately small chromosomes (1.2-4.1 µm), with the centromere located in the middle or submid region (Fig. 2). In C. specularioides, one pair of chromosomes with the centromere in the submiddle region has a satellite on the short arm. Chromosomes form bivalents in the meiotic metaphase. The basic number found in annual species of the ‘lusitanica’ subclade in the W Mediterranean, x = 10, coincides with that found in other species of this subclade: C. rapunculus, C. patula and C. sparsa (Larsen, 1956; Contandriopoulos, 1966). The basic number x = 9 found in C. lusitanica and C. matritensis can be considered as aneuploid of x = 10. The base number n = 10 is extremely rare in Campanulaceae, and has only been previously reported for C. ramosissima (Podlech & Damboldt, 1964). The chromosome number found in C. dieckii, 2n = 28, has been previously found in the subgenus Roucela sensu Damboldt, in C. erinus (Gallego, 1986) and C. drabifolia Sibth. & Sm. (Contandriopoulos, 1964a), and in C. edulis Forssk. (according to Goldblatt & Johnson, 1990), all species of ‘Campanula s. str.’ clade, and also in C. arvatica Lag. (Podlech & Damboldt, 1964), a species of ‘Rapunculus’ clade (Fig. 1). The chromosome number found in C. decumbens, 2n = 32, is also found in some species of the ‘Rapunculus’ clade, [C. raineri Perpenti (Podlech & Damboldt, 1964), C. fragilis Cyr. (Damboldt, 1965), C. isophylla Moertti (Merxmüller & Damboldt, 1962), C. herminii Hoffmans. & Link (Damboldt & Podlech, 1964), and C. stevenii Bieb. (according to Goldblatt, 1985), and Edraianthus graminifolius (L.) A. DC. (Contandriopoulos, 1964b), a taxon of the ‘Campanula s. str.’ clade, and Musschia aurea Dumort. (according to Goldblatt & Johnson, 1994), a taxon of ‘Musschia’ clade]. Since 2n = 34 is the most frequent number in Campanulaceae, it seems likely that 2n = 32 is an aneuploid of 2n = 34 (see Fig. 1). Systematic treatment After reviewing all specimens we found that the different species can be separated in two groups on basis on the morphology of the style and stigma: one formed by C. decumbens and C. dieckii, with the style glabrous and with three stigmas with arms straight and erect or erect-patent; and another formed by all other species, with the style hairy in the upper half, and a trifid stigma, with the stigmatic arms curved or circinate. These two groups are consistent with the molecular results and karyology, so that they are probably natural, and the most appropriate taxonomic treatment is to recognise them as sections. The second Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 21 Taxonomic review of the Campanula lusitanica complex 21 Fig. 2. Karyology of Campanula species. A-F and I, mitotic metaphase; G, meiotic anaphase I; H, meiotic metaphase I. A, C. lusitanica, 2n = 18 (Rivadavia, Orense, Spain, SEV 218947); B, C. matritensis, 2n = 18 (Hinojos, Huelva, Spain, SEV 216214); C, C. cabezudoi, 2n = 20 (Venta de Zafarraya, Granada, Spain, SEV 218873); D, C. specularioides, 2n = 20 (Montejaque, Málaga, Spain, SEV 216210); E, C. transtagana, 2n = 20 (Valverde del Camino, Huelva, Spain, SEV 216212); F, C. broussonetiana, 2n = 20 (Jbel Tazzeka, Taza, Morocco, SEV 216476); G, C. decumbens, n = 16 (Benaoján, Málaga, Spain, SEV 218875); H, C. decumbens, n = 16 (Villamartín, Cádiz, Spain, SEV 256653); I, C. dieckii, 2n = 28 (Alfarnate, Málaga, Spain, SEV 256652). The scale bar = 20 µm. Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 22 22 J. Cano-Maqueda & S. Talavera group, which includes Campanula rapunculus L., would constitute the sect. Rapunculus. The first group would establish a new section, sect. Decumbentes which is formally described below. Within these sections, the most significant characters are: the shape of the corolla (campanulate or infundibuliform), middle cauline leaves (petiolate or sessile), capsule morphology (obpyramidal, subglobose or subovoid), and ovary indumentum (setose, glabrous or papillose). We used this combination of characters to create a key for the identification of all annual species of the C. lusitanica complex in the Western Mediterranean. KEY TO THE SPECIES 1. Style 2.1-4(4.8) mm, glabrous, with three stigmas; stigmas straight, erect or erect-patent ........................................... 2 1. Style (4.5)5-14 mm, hairy in the upper half, with one trifid stigma; stigmatic branches curved or circinate, patent ....... 3 2. Stems glabrous or with few antrorse hairs, often scabrid near the flowers; upper cauline leaves cuneate or shortly petiolate; corolla 12-21 mm ................................... 7. C. decumbens 2. Stems densely pubescent with setose hairs, sometimes glabrescent towards the apex; upper cauline leaves sessile, subauriculate; corolla (6.6)8-13.7 mm ............. 8. C. dieckii 3. Corolla infundibuliform, with three purple nerves in each lobe; middle cauline leaves petiolate; plant decumbent ..... 4 3. Corolla campanulate, without purple nerves in the lobes; middle cauline leaves sessile or petiolate; plant erect or decumbent .......................................................................... 5 4. Capsule 2-3 × 3-4.5 mm, subspherical, generally glabrous, dehiscing by 3 pores of middle position, with 10 subwinged nerves; stems and leaves glabrous or glabrescent; calyx-teeth oblanceolate ...................................... 4. C. specularioides 4. Capsule 2.1-5.9 × 2.2-5 mm, ovoid or subspherical, usually densely hairy, dehiscing by three apical pores, with 10 ± acute but non-winged nerves; stems and leaves pubescent; calyx-teeth lanceolate or linear .................. 3. C. cabezudoi 5. Middle cauline leaves petiolate and elliptic or sessile and cuneate ............................................................................ 6 5. Middle cauline leaves sessile, ovate-lanceolate or lanceolate ................................................................................. 7 6. Upper half of the stem and leaves with short hairs 0.1-0.3 mm; middle cauline leaves with petiole 0.2-4.5 mm ............. ............................................................... 5. C. transtagana 6. Upper half of the stem and leaves with long hairs, the longest 0.4-2 mm, or rarely glabrous; middle cauline leaves sessile and cuneate or with petiole up to 2 mm and rounded at the base ................................................. 6. C. broussonetiana 7. Middle cauline leaves ovate-lanceolate, subauriculate, densely patent-hairy, with long hairs up to 0.8-1.1 mm; capsule subovoid, with 10 subwinged acute nerves ....... 1. C. lusitanica 7. Middle cauline leaves lanceolate, glabrous or with sparse hairs 0.1-0.5 mm; capsule obpyramidal, with 10 very wide nerves, like flat ribs .................................. 2. C. matritensis A. Sect. Rapunculus Boiss., Fl. Orient. 3: 895. 1875 Campanula subgen. Rapunculus (Boiss.) Kharadze in Zametki Sist. Geogr. Rast. Tiflis 28: 100. 1970 Type: C. rapunculus L. Annual or biennial plants. Lower leaves oblanceolate or spathulate, petiolate. Without calycine appendages. Corolla campanulate or infundibuliform. Stamens with white or blue anthers. Style with numerous pollen collecting hairs on the upper half, with a trifid stigma; stigmatic arms curved or circinate, patent, white or blue, with numerous pollen collecting hairs on the abaxial surface glabrous, and receptive on the adaxial surface. Capsule subspherical, ovoid or obpyramidal, dehiscing by three apical or middle position pores. x = 9, 10 (see karyology). Observations: This section includes: Campanula rapunculus and C. patula of the Mediterranean region; C. lusitanica, C. matritensis, C. cabezudoi, C. specularioides, C. transtagana and C. broussonetiana from the W Mediterranean; and C. sparsa and possibly C. spatulata Sibth. & Sm. of the E Mediterranean. The section is monophyletic (see lusitanica clade in Fig. 1). Experimental manual crosses showed that all annual species of the W Mediterranean had hybridization barriers at different levels: (1)- fruits were not produced, (2)- seeds do no germinated, (3)- germinated seeds formed chlorotic seedlings which did not survive and (4)- no chlorotic seedlings survived after the cotyledon state but they did not complete their development (Cano-Maqueda unpublished studies). 1. Campanula lusitanica L. in Loefl., Iter Hispan.: 111, 126. 1758 Campanula loeflingii Brot., Phytogr. Lusitan. Select. Fasc. I: 20. 1800, nom. superfl. C. patula var. lusitanica (L.) Pau in Bol. Soc. Iber. Ci. Nat. 20: 181. 1921. Ind. loc.: “Habitat in Lusitania ad Porto in collibus & muris”. Type: Portugal. Porto, June 1917, Mário de Castro s.n. [Sennen, Pl. Espagne n.º 3305] (neotype, here desinated, G 104248!, Fig. 3; see observations). C. duriaei Boiss., Voy. Bot. Espagne 2: 402. 1839. Ind. loc.: “Hab. In Hispaniâ circà Olyssiponem (L. Et Hoffm.), Hispaniâ septentr. In Asturiis (Durieu)”. Type: Spain. Asturias, Cangas de Tineo, 25 July 1835, Durieu s.n. [Plant. Select. Hispano-Lusit. Sect. 1ª n.º 280] (lectotype, here designated, G 104093!; see observations). C. loeflingii var. occidentalis Lange in Vidensk. Meddel. Dansk Naturhist. Foren. Kjøbenhavn 1861: 107, 108. 1862. Ind. loc.: “In Gallecia frequent!”. Type: Spain. Galicia [not found in the Lange herbarium (C)]. C. lusitanica var. puberula C. Vicioso in Anales Jard. Bot. Madrid 6: 78. 1946. Ind. loc.: “Hab. Ribadelago (Zamora)”. Type: Spain. Zamora, Ribaldelago, Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 23 Taxonomic review of the Campanula lusitanica complex 19 July 1945, C. Vicioso s.n. (lectotype, here designated, MA 121525!; see observations). Illustrations: Fig. 4 A, B. Herb 16-50 cm, annual, erect, branched from the base, rarely in the upper half, densely pubescent, not brittle. Stems angled, branched or simple, densely pubescent, rarely glabrescent in the uppermost portion, with short and long hairs, the longest 0.5-1.5 mm. Leaves somewhat coriaceous, entire, crenulate or toothed; middle cauline leaves 5-30 × 3.1-10.9 mm, sessile, ovate-lanceolate, subauriculate, entire or toothed, densely pubescent, ± long hairs, the longest 0.8-1.1 mm; upper cauline leaves 4-14.5 × 0.5-4.4 mm, sessile, lanceolate, truncate or somewhat auriculate, pubescent, with hairs up to 0.9 mm. Inflorescence paniculate, highly branched, lax or dense and spiciform. Flowers pedicellate or subsessile; pedicel (3)716 mm, glabrous or with hairs 0.1-0.2 mm. Calyxteeth 3.7-9(17.3) × 0.4-0.6(1.3) mm, linear, recurved in the female phase flower. Corolla (8.2)10-18.9 mm, campanulate, with tube longer or shorter than lobes; tube 3.5-9.7 mm, blue at the apex, white at the base; lobes 3.7-9.2 × 2.2-6.1 mm, elliptic-lanceolate, not re- Fig. 3. Neotype of Campanula lusitanica (G 104248). 23 flexed in the female phase flower, blue. Stamens with enlarged base of 0.6-1 × 0.4-0.5 mm; filaments 0.2-0.5 mm; anthers (1.5)3-5 mm, whitish. Ovary papillose or with setose hairs of 0.1-0.2 mm; style 7.5-11.8 mm, hairy in the upper half; stigma trifid, with stigmatic branches of 0.3-1.2(2.8) mm, patent, circinate, white. Capsule (2.4)4-6 × (2.5)3-4 mm, subovoid, longer than wide, papillose, rarely with some setose hairs 0.10.2 mm, with 10 subwinged and acute nerves, dehiscing by three apical pores. Seeds 0.3-0.5 × 0.2-0.3 mm, ellipsoid, shining, yellowish to brown. 2n = 18; n = 9. Habitat, phenology and distribution: Wet grasslands on generally acid substrates, sometimes draining and river banks; 15-1800 m. (V)VI-VIII. • Endemic to the NW of the Iberian Peninsula, common in Galicia and N Portugal, and also occurring in S Portugal, Monchique (Fig. 5). Portugal: Ag BA BL DL Mi TM. Spain: C Le Lu O Or Po Sa Za. Observations: López-González (in litere) has commented apropos the type of Campanula lusitanica: “Al parecer, Linneo no conoció nunca Campanula lusitanica, porque dicha planta no figura en la lista de materiales de Loefling enviados a Linneo en 1752 (Spanish list), pero en una carta de Loefling a Linneo, fechada el 7.VII.1751 y escrita en Oporto, le indicó que «Campanula caule angulato, ramoso, vago, calice corollae tubulosae aequali», la que Linneo denominaría C. lusitanica, habita aquí [Porto] en los caminos y en las tapias. Tampoco figura esta planta ni como Campanula lusitanica ni como «Campanula caule angulato…» en la lista del herbario de Loefling que redactó un capellán sueco, probablemente porque en esta lista solo figuraban las plantas identificadas, y «Campanula caule anguloso…» aún no tenía nombre. Es en el herbario de Loefling donde se podría encontrar la planta como Campanula sp., pero este herbario lo perdió Casimiro Gómez Ortega, que al parecer, según cuenta en una carta, lo prestó a un botánico francés y éste no lo devolvió”. Since it was impossible to find the type material of C. lusitanica in the different Linnean herbaria, the election of a neotype has been proposed. The plants in question have long and patent cauline hairs, and so do not correspond to the plant described by Loefling, who indicates “Caulis … leviter hispidus pilis pallidis, brevissimis”. The shape of the cauline leaves, ovateoblong or ovate-lanceolate, are however like those of the plants described by Loefling, who indicates “Folia caulina & subramorum ovato-oblonga, subglabra, sessilia, subserrata, alterna; ramorum superiora ovatolanceolata, vix serrata” and the whitish colour at the base of the corolla tube is also consistent with the description of Loefling: “Cor. caeruleis, tubulo infimo Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 24 24 J. Cano-Maqueda & S. Talavera brevísimo albo”. In accordance with these characteristics, we have chosen the plants from Porto as neotype of Campanula lusitanica. Brotero (1800: 20-21) gave a new name, C. loeflingii, to the plant collected in Porto by Loefling, but this name is illegitimate, since Linneus had named it as C. lusitanica in the work of Loefling. Brotero (1804: 287) subsequently not only described the Loefling plant, but also included characters from another species later described by Alphonse De Candolle Fig. 4. Flowers and fruits of Campanula species. A, B, C. lusitanica (Rivadavia, Orense, Spain, SEV 218947); C, D, C. matritensis (Hinojos, Huelva, Spain, SEV 261264); E, F, C. cabezudoi (Venta de Zafarraya, Granada, Spain, SEV 218873). The scale bar = 3 mm. Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 25 Taxonomic review of the Campanula lusitanica complex (1830) as C. matritensis. Indeed, the illustration that appears in his “Phytographia Lusitanieae Selectior” (Brotero, 1816, tab. 18) corresponds more closely to C. matritensis than to C. lusitanica. Hoffmannsegg & Link (1820-1824: 14) also provided a brief description of C. loeflingii, but like Brotero, they could be referring to both C. lusitanica and C. matritensis. Boissier (1839) described C. duriaei based on the characters of plants collected by Durieu in Cangas de Tineo (Asturias) distributed as the Exiccata No 280 in 1835. In the herbarium at Geneva there are three sheets of this collection (G 104093, G 104120 y G 104116). The first (G 104093) consists of 4 complete plants mounted on two sheets, and comes from the Boissier Herbarium. One of the sheets contains two plants of 30 cm and 22 cm respectively, and a Boissier handwritten label indicating “C. Duriaei/284”. We have chosen as lectotype the 30 cm plant because it is the closest to the description; the other plant, of 22 cm, is an isolectotype. The other sheet also contains two plants without any annotation by Boissier; they are also likely isolectotypes. The other two sheets (G 25 104120 and G 104116) are from the herbarium of Moïse-Etienne Moricand which were incorporated into the herbarium of the Conservatoire et Jardín Botanique de Genève in 1908; the plants contained in them, which were not seen by Boissier, are not part of the type material. Boissier was the first author to clearly describe the characteristic hairs of the Asturian plants: “Toute la plante est d’une consistance délicate et couverte de poils long et étalés, tandis que la C. loeflingii est u glabre ou hérissée de poils rudes et courts”. Boissier’s concept of C. loeflingii was very wide, since he included in this species most of the taxa of this group which had been collected by him in southern Spain or that he had seen in the Fouche or De Candolle herbaria. We identify Fouche herbarium plants as C. decumbens subsp. baetica Cano-Maqueda & Talavera and those in the De Candolle herbarium as C. broussonetiana, and the plants that he collected in Southern Spain as C. matritensis and C. dieckii (see these species below). The type material of C. lusitanica var. puberula consists of three plants mounted on the same sheet. We Fig. 5. Distribution map of Campanula lusitanica (䊱) and C. matritensis (䡬). Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 26 26 J. Cano-Maqueda & S. Talavera have chosen as lectotype the larger specimen mounted on the left, and the other two plants are isolectotypes Campanula lusitanica is very similar in the indumentum and the branching to C. broussonetiana, but C. lusitanica has 2n = 18 chromosomes, as does C. matritensis, while C. broussonetiana has 2n = 20 chromosomes, as do almost all species of sect. Rapunculus. The ITS phylogenetic tree shows that C. lusitanica and C. matritensis are sister species (see Fig. 1). Selected specimens PORTUGAL. Algarve: Serra de Monchique, entre Monchique e Alferce, Rouxinel, 27-VI-1978, M. Beliz (MA 270314); ibidem, de Pesos a Monchique, 24-VII-2009, S. & M. Talavera (SEV 248712). Beira Alta: Figueira de Castelo Rodrigo, Escalhão, 26-V-1996, M. Santos & M. Sequeira (HVR 8048). Serra da Estrela, without date, Romariz (LISU 2054); ibidem, Garganta de Loriga, VIII-1912, A.A. Silva Martins (LISU 36386). Vizeu, Santa Comba Dão, Pinheiro de Azer, arredores da ponte sobre a barragem, 20-VI-1982, A. Marques (MA 377387). Beira Litoral: Coimbra, Penacova, Oliveira do Mondego, 13-V-1982, A. Marques (MA 391392). Douro Litoral: entre Oliveira y San Joao de Madeira, dirección Oporto, 24-VI1986, J.A. Devesa & al. (SEV 161720). Porto, Castelo prox. Souto de Lafões, 27-V-1940, recolector ilegible (LISU 36362). Minho: Ancora, VI-1886, A.R. da Cunha (LISU 36367). Areosa, VI-1886, A.R. da Cunha (LISU 36404). Barcelos, VI-1886, A.R. da Cunha (LISU 36368). Caldas do Gerez, IX-1882, M.L. Enriques (COI). Caminha, Retorta, VI-1885, A.R. da Cunha (LISU 36379). Celorico de Basto, VI-1884, A.R. da Cunha (LISU 36380). Entre Ponte de Barca y Vila Verde, 27-VI-1982, M.J. Gallego & al. (SEV 161716). Fafe, Lameira, 7-VIII-1977, M. Beliz (MA 270317). Gondarem, VI-1885, A.R. da Cunha (LISU 36377). Melgaço, VI-1894, A. Moller (COI). Monção, Lavandeira, VI-1885, A.R. da Cunha (COI, LISU 36407). Ponte de Lima, VII-1894, M. Rodr. Maraes (COI). Povoa de Lanhoso, VI-1920, J. Sampaio (MA 121499). Senhora da Peneda, Serra do Soajo, VII-1890, A. Moller (COI). Serra Amarela, Mata do Cabril, Carvalhal do Sono, 8-VIII-1977, M. Beliz & J. Guerrero (MA 270316), Serra da Peneda, 880m, VI-1956, R. Bentos (LISE 48610). Serra do Gerez, 2-VII-1948, Romariz (LISU 922). Soajo, Serra do Soajo, VI-1890, A. Moller (COI). Vanlença, Olivar de Santa Barbara, VI-1885, A.R. da Cunha (LISU 36403). Veiga de Chaves, V-1910, F. Mendez & al. (LISU 36389). Veiga de Ganfei, VI-1885, A.R. da Cunha (LISU 36392). Vila Nova da Cerveira, VI1885, A.R. da Cunha (LISU 36402). Trás-os-Montes: arredores de Bragança, VI-1882, P.F.M. Var (COI). Bragança, 1877, Pereira Coutinho (LISU 36358). Entre Montalegre e Chaves, Sapiãos, VI1910, R. Palhinha & al. (LISU 36385). Estação do Pocinho, VI1915, Mendes & Palhinha (LISU 36387). Mogadouro, a montante da pte. de Ramondes, na margem dta. do rio Sabor., 18-V-1997, A. Castro & Tjarda De Koe (HVR 9475). Montalegre, 21-VII-1959, M. da Silva (G 104096). Montinho, Seixas, VI-1885, A.R. da Cunha (LISU 36401). Santa Marta de Penaguião, Veiga, Aldeia do Bertelo, 25-VI-1993, A. Coelho Costa & A.L. Crespí, A (HVR 10177). Vila Nova de Foz Côam, margem do Douro (esquerda) a jusante da Foz do Sabor, frente à Ilha, 30-IV-1995, M. Sequeira (HVR 5937). Vila Real Coêdo, 10-VII-1981, A. Além (HVR 3902, SANT 39046). Vimioso, VI-1888, G. Mariz (COI). SPAIN. Asturias: Cangas de Tineo, 2-VI-1864, E. Bourgeau, in E. Bourgeau, Pl. d’ Espagne 1864: nº 2657 (G 104119, G 104118, MA 121552, MA 152778). Corneliana, 16-VIII-1868, E. Boissier (G 104109). Pravia, without date, La Gasca (MA 121844). Taramundi, VII-1979, T.E. Diaz (MGC 14104). Asturias, VII-VIII- 1878, E. Boissier (G 104121, G 104117). La Coruña: Aranga, 30VI-1967, J. Dalda (SALA 35816). Cariño, Landoi, 4-VIII-1994, X. Soñora (SANT 31851). Carnota, 1-VI-1996, R.I. Louzán (FCO 24813, MA 581374, SANT 35758). Cuenca del río Deo, 1966-1968, J. Dalda (SANT 55699). Ferrol, 27-VI-1994, X. Soñora (SANT 29263). Mazaricos, 25-VI-1995, R.I. Louzán (SANT 35746). Santiago de Compostela, Pontepedriña, VI-1995, Rodríguez-Hergueta (SANT 42394). Sobrado, 24-VII-1882, J. Lange (G 104115). Somozas, 17-VII-1994, X. Soñora (SANT 31883). León: La Baña, Sierra de Cabrera, camino del Lago, 10-VII-1981, Lansac & Nieto (MA 317215, MA 317216, MA 317216). Palacios del Sil, Salentinos, 27-VIII-1996, Martín-Blanco (MA 597090). Ponferrada, Villanueva de Valdueza, VIII-1995, Rodríguez-Hergueta (SANT 42467). Riocastrillo de Ordás, 22-VI-1997, C. J. Martín-Blanco (MA 612392). Sobrado, Castropetre, 30-V-1990, J. Amigo & J. Giménez (SANT 26556). Valle del Bierzo, VI-1905, M. Gandoger (G 104142). Lugo: alrededores de la ciudad, 26-VI-1987, E. Carreira (MA 513456). Becerreá, El Cruzul, 18-VII-1989, S. Castroviejo & al. (MA 471686). Cabreira-Fonsagrada, VII-1957, E. Carreira (MA 204700). Cervantes, Correal, entre El Portelo y Piedrafita, 28-VI1994, M. A. Carrasco & al. (MA 543383). Chantada, 18-VI-1988, M. Buján (SANT 25112). Entre Esperante y Carbedo, 5-VII-1979, J. Amigo & al. (SANT 16230). Ferreiravella, 19-VI-1980, J. Amigo & al. (SANT 16229). Meiraos, 16-VII-1981, J. Amigo & al. (SANT 16226). Monforte, 18-V-1991, J. Amigo & M. Romero (SANT 25426). O Saviñao, Mourelos, 14-VII-1992, J. Amigo & M. Romero (SANT 22471, MA 517178). Palas de Rey, 16-VI-1951, Seijas (SANT 6124). Piornedo, 23-VII-1952, Bellot & Casaseca (SANT 6783). Quiroga, Hermida, San Victorio, 18-VI-1988, M. Buján & M. I. Romero (MA 546783, SANT 25107). Riberas de Lea, 25-VII1956, E. Carreira (MA 201352). Sierra del Caurel, Seoane, 28VII-1992, E. Blanco (MA 564613). Tardad. Villalba, 30-VII-1951, M. Orosa (G 104101). Vilamelle, 6-VI-1990, M.I. Romero (MA 546757. SANT 26231). Orense: Carballeda, 10-VII-1984, S. Ortiz (SANT 16523). Chandreja de Queija, pr. Paradaseca, 24-VII-1974, S.I. Laínz (MA 345998). Corrainzas, 12-VIII-2000, J. De Jesús & al. (SANT 46020). Entre Sobradelo y Casayo, 15-VI-1958, Bellot & Casaseca (SANT 9940). Leboreiro, 13-VII-1958, F. Bellot & B. Casaseca (MA 180705). Lobios, 9-VII-1993, I. Pulgar (SANT 56572). Río Lonia, 15-VII-2000, A.R. Romero & al. (SANT 45644). Río Sil, 15-VI-1958, Bellot & Casaseca (SANT 10066). Rivadavia, carretera a la Franqueirán, 24-VIII-2007, R. Pino (SEV 218947). Rubiá, Pardellán, Río Sil, 29-VI-1994, M.A. Carrasco & al. (MA 542966, SALA 115638). Serra do Invernadeiro, inter Rocín et Suacenza, 11-VII-1973, S. Castroviejo (MA 219730, MA 196712). Verín, 7-VI-1988, M. Buján & M. I. Romero (SANT 25110, SANT 25108). Pontevedra: Albeos, 14-VI-1988, M. Buján (SANT 25113). Cangas de Morrazo, 6-VIII-2007, S. Castroviejo (SEV 218948). Crecente, 24-V-1998, J. Amigo & al. (SANT 39787). Cuntis, 9-VI2007, J. García Devesa (SANT 57381). Ermelo, Bueu, 15-VII-1970, S. Castroviejo (MA 196710). Meaño, 16-III-1988, M. Buján & M.I. Romero (SANT 25109). Porteliña, 16-VIII-1983, S. Sivestre (SEV 161714). Prado, 15-VII-1947, Vieiter (SANT 213). Salvaterra de Miño, 24-V-1998, J. Amigo & al. (SANT 39806). Salamanca: Aldeadavila, 22-IV-1977, F. Amich (SALA 15518). La Fregeneda, 30-IV1977, F. Amich (SALA 15519). Zamora: Montelarreina, 31-V-1987, R. García Ríos (SALA 54045). Ribadelago, 14-VI-1987, A. Roa & P. García (MA 510330). 2. Campanula matritensis A. DC., Monogr. Campan.: 332. 1830 C. lusitanica subsp. matritensis (A. DC.) Franco, Nova Fl. Portugal: 2, 569. 1984. C. patula var. matritensis (A. DC.) Pau in Bol. Soc. Iber. Ci. Nat.: 20, 181. Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 27 Taxonomic review of the Campanula lusitanica complex 1921. C. loeflingii var. matritensis (A. DC.) Lange in Vidensk. Meddel Dansk Naturhist. Foren. Kjøbenhavn 1861: 108. 1862. Ind. loc.: “Habitat in Hispania circa Matritum (Lagasc.!)”. Type: Spain. Madrid, 1806, Lagasca s.n. (lectotype, here designated, G 138402! (G-DC.), Fig. 6; see observations). Illustrations. Gallego (1987: 564, as C. lusitanica); Brotero (1816, tab 18, as C. loeflingii); Hoffmannsegg & Link (1820, tab. 82, as C. loeflingii ); Sáez & Aldasoro (2001: 130, fig. 39b-h, as C. lusitanica subsp. lusitanica); Fig. 4 C, D. Herb (6)10-55 cm, annual, erect, branched at the base or in the upper half, often laxly pubescent, not brittle. Stems angled, little branched, laxly pubescent in the lower half with setose hairs 0.1-0.3(0.5) mm. Leaves not coriaceous, crenate, dentate or entire; middle cauline leaves 4.5-27 × 0.9-9.2 mm, sessile, lanceolate, entire or toothed, glabrous or laxly pubescent, with hairs 0.1-0.5 mm; upper cauline leaves 2-22 × 4.5-5.8 mm, sessile, lanceolate, glabrous or with some hairs of 0.1-0.4 mm. Inflorescence laxly paniculate. Flowers pedicellate; pedicel 3.5-123 mm, glabrous or with some hairs of 0.1-0.2 mm. Calyx-teeth (2.2)4-15 × 0.3-1(1.3) mm, linear, filiform. Corolla (8.8)11-26 mm, campanulate, with the tube much longer or much shorter than the lobes; tube (4.2)7-15.3 mm, bluish; lobes (5.5)6-16 × 2-7 mm, ovate, blue. Stamens with enlarged base of 0.6-1 × 0.3-0.7 mm; filaments 0.4-0.6 mm; anthers (2)3-5 mm, white, rarely bluish. Ovary glabrous, papillose or with scattered hairs of 0.1-0.2 mm; style 5-14 mm, hairy in the upper half; stigma trifid, with stigmatic branches of 1-2(3.1) mm, patent, circinate at the end of the female phase of the flower, white. Capsule (2.5)4-9.2 × 2-5 mm, obpyramidal, much longer than wide, glabrous, papillose or with some setose hairs less than 0.1 mm, with 10 very wide nerves like flat ribs, dehiscing by three apical pores. Seeds 0.4-0.6 × 0.2-0.3 mm, ovoid, shining, yellowish to brown. 2n = 18; n = 9. Habitat, phenology and distribution: Pine, cork oak and holm oak woodlands or their shrubby secondary communities, often on sandy substrates, but also on clay; 0-2200 m. V-VI(VII). • Endemic to the Iberian Peninsula. Distributed almost throughout the entire peninsula, but rare in limestone areas of C, E and N of Spain, and absent in NE of Spain (Fig. 5). Portugal: AAl Ag BA BAl BB BL E Mi R TM. Spain: Ab Al Av Ba Bu C Ca Cc Co CR Cu Gr Gu H J Le Lo Lu M Ma O S Sa Se Sg So Te To Va Vi Z Za. Observations: This species is very variable in size, flower size, indumentum and shape of the calyx, and 27 indumentum and size of the capsules. The plants that live in very shady banks are usually delicate, have small flowers and, in general, asymmetric capsules due to lack of fertilization in one or two of the three locules, probably caused by deficient pollination. In contrast, plants living in sunny and humid areas are vigorous, highly branched, with large flowers and perfectly symmetrical obpyramidal capsules, with all locules full of seeds, denoting a very efficient fertilization. On shallow soils, in montane areas, plants are very small, slightly branched, with small flowers and sometimes with ± ovoid and malformed capsules due to a deficient pollination, and these may be confused with C. transtagana or even with C. lusitanica. The three plants of type material of C. matritensis (Fig. 6), about 15 cm, attached to the label, have sessile cauline linear-lanceolate leaves, and obconical capsules. These agree with the characters described by Fig. 6. Lectotype of Campanula matritensis (plant located in the lower left corner) (G 138402, indicated by the arrow). The lectotype is the plant of the center, the other two plants are isolectotypes. The other two plants with their respective labels are not material type. Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 28 28 J. Cano-Maqueda & S. Talavera Alphonse De Candolle (1830). That these three plants include the type material is further indicated by an Alphonse De Candolle label at the bottom of the sheet with “Campanula matritensis Alph. DC.”. Of these three plants, the largest, located in the middle, is chosen as lectotype of Campanula matritensis A. DC. The other two plants are isolectotypes. In the same sheet there are other two plants which are not type materials (see Fig. 6). One [Herb. Prodr. (G-DC)G 138459] indicates “Campanula/ex hispania/m’ Guillemin 1827” and the plant, branched and young, is identified as C. matritensis; the other [Herb. Prodr. (G-DC)G 138462] indicates “C. pulla herb Lag./prope la Venta del Gorro in nov. Castell./1827 [1827? manuscript Lagasca?]/Londres 1828 [manuscript Lagasca]”. Coupled with this label there is another, probably handwritten by A. De Candolle, with the following inscription: “C. matritensis?/differt ab ómnibus affinibus lobis/calicinis marginem extus revolutis.”. Stitched to both labels there is a very small plant with a single flower, which we identify as C. dieckii Lange. Selected specimens PORTUGAL. Algarve: Broussailles à Monchique, 14-VI-1853, E. Bourgeau, in E. Bourgeau, Pl. d’ Espagne et de Portugal, 1853: nº 1943 (G 104070, G 104125, G 104140). Cabo de San Vicente, Chodat (G 104147). Castro Marim, Terras da Ordem, 24-IV-1975, M. Sequeira & al. (HVR 4734). Entre Corte-Figueira y Mù, VI/VII1885, J. Daveau (LISU 36408). Faro, Santo Antonio do Alto, V1889, J. Brandeiro (G 104123). Olhão, Quinta de Marim, 24-V-1986, A. Moura (MA 430492). Entre Santa Catharina e Sagres, V-1906, F. Mendes & Palhinha (LISU 36391). Serra da Picota, VII-1891, J. Brandeiro (COI). Serra de Monchique, Ribeira da Perna Negra, 30V-1979, M. Beliz & J. Guerrero (MA 270313). Alto Alentejo: Elvas, próximo da estrada Elvas-Badajoz, 1-I-1900, A. Forque (LISE 31816). Évora, V-1891, A. Moller (COI). Évora Monte, entre Estremoz y Évora, 28-V-1996, S. Castroviejo & al. (MA 588831). Gavião, 26-VI-1952, P. Silva, F. Fontes & B. Rainha (G 104102). Margen do rio Alirilungo, afluente de Xevora, V-1922, Fernandes (LISU 36417). Marvão, Serra de San Mamede, 16-V-1978, J.A. Devesa & J. Pastor (SEV 39778). Montargil, V-1883, G. Corteisão (COI). Mora, arredores nas arelas do leito da Ribeira da Raia, 13-V-1953, Bento Rainha (LISE 51183). Portalegre, Serra de Penha, VI-1882, A.R. da Cunha (LISU 36400). Portel, 15-V-1969, P. Silva & al. (LISE 93823). Povoa e Meadas, VI-1882, A.R. da Cunha (LISU 36370). Redondo, V-1892, D. Pita Simões (LISU 36361, LISU 36366). Reguengos, Berrocal, IV-1908, F. Mendes & Palhinha (LISU 36411). Serra d’Ossa, V-1891, A. Moller (COI). Baixo Alentejo: Cazevel, V-1888, A. Moller (COI). Cuba, entre Cuba e Vila de Frades, 4-V-1962, M. Silva (LISE 76879). Entre Garvão e Panóias, VI/VII-1885, J. Daveau (LISU 36393). Odemira, V-1915, F. Gomez & R. Machado (LISU 36415). Santiago de Cacém, 14-V-1958, B. Rainha, M. Silva & A.N. Teles (LISE 64924). Margen do rio Chança, VI-1913, F. Mendes, H. Navel & Palhinha (LISU 36414). Serra do Caldeirão, 12-V1905, M. Gandoger (G 104143). Sines, a 1 Km a norte de porto Covo, 21-V-1982, F. Catarino & al. (LISU 145357). Tarrão, 29-V1952, F. Fontes & al. (LISE 41814). Beira Alta: Águeda, perto de Escalhão, 26-V-1996, M. Santos & M. Sequeira (HVR 7845). Barca d’Alva, margen do Douro, VI-1915, F. Mendes & Palhinha (LISU 36416). Castelo Mendo, Moita do Carvalho, VII-1884, A.R. da Cunha (LISU 36399). Covilhã, VI/VII-1881, A.R. da Cunha (LISU 36395). Entre Guarda y Valhelhas, 16-VII-1983, S. Castroviejo & al. (MA 248455). Figueira de Castelo Rodrigo, Barca D’ Alva, 28-IV1943, J. Castro & A. Rozeira (MA 514273). Sabugal, 14-VI-1976, I. Melo & al. (LISU 69654). Serra da Estrela, VII-1887, A. Moller (MA 121501). Serra da Lapa e Mata da Vide, VI-1890, M. Ferreira (COI). Vale do Zêzere, 3-VIII-1949, C. Romariz (LISU 2053). Vouzela, 27V-1940, Palhinha (LISU 36363). Sabugal, à saída da vila, na estrada Castelo Branco (Miradouro), 14-VI-1976, I. Melo,& al. (SEV 121226). Beira Baixa: Alcaide, Barroca do Chorão, VI-1882, A.R. da Cunha (LISU 36369). Castelo Branco, VI-1881, A.R. da Cunha (LISU 36398). Covilhã, nos latudes da estrada prose da ribeira da Carapinheira, 23-VI-1946, B. Rainha (G 104146, LISE 21736, MA 121494). Monfortinho, prox. das termas, 14-VI-1948, B. Rainha (LISE 22693, MA 152780). Tavanca do Mondego. 31-V-1990. Z. Díaz & al. (SEV 161692). Beira Litoral: Arganil, Moita, V-1895, M. Ferreira (COI). Cantanhede, VI-1888, C.M. Ferreira (COI). Choupal prope Conimbricam, VI-1878, A. Moller (G 104124, G 104122). Coimbra,, 6-VIII-1883, A. Moller (G 104139). Guardinha, arred. do Louriçal, VI-1897, J.A. Vaz Serra (COI). Meco, 16-IV1995, A. Crespi & M. Sequeira (HVR 4604). Taboa, V-1883, A. Costa Carvalho (COI). Estremadura: Alfeite, V-1906, J. dos Santos (LISU 36384). Alrededores de Lisboa, Serra de Monsanto, VI-1884, A.R. da Cunha, in Flora Lusitanica (Soc. Brot. 7º. Anno) nº 910 (LISU 36360). Arredores de Setúbal, V-1906, F. Gomez (LISU 36409). Lagoa de Albufeira, V-1882, J. Daveau (LISU 36371). Moita, Arganil, V-1895, M. Ferreira (COI). Praia das Maçãs, V-1930, F. Matos (LISU 36364). S. Simão, Piedade, VIII/IX-1848, J. Daveau (LISU 36396). Sacavém, 1952, Duarte de Castro (LISE 40720). Seixal, V-1881, A.R. da Cunha (LISU 36397). Serra da Arrabida, Colina de Santa Margarida, 18-V-1942, C. Fontes & al. (LISE 15267, MA 121496). Serra de Montejunto, 31-I-1947, Romariz & Mendes (LISU 56937). Sesimbra, Alfarim, 2-VI-1971, M. Beliz & J. Guerreiro (MA 270318). Vila Nogueira, 25-V-1978, J.A. Devesa & al. (SEV 39777). Minho: Gerez, VII-1889, F. Loureiro (COI). Valença, Olivar de Sta. Barbara, VI-1885, A.R. da Cunha (COI). Ribatejo: Almeirim, 22-V1952, P. Silva & M. Silva (LISE 41804). Trás-os-Montes: arred. de Miranda do Douro, Iffanes, VI-1888, J. de Mariz (COI). Arredores de Bragança, 16-VI-1941, A. Carneiro (COI). Arredores de Moncorvo, Concelho de Mogadouro, Urrós, 20-V-1997, J. Hernándes & E. Rico (SALA 90849). Avelanoso, arred. de Vimioso, VI-1888, J. de Mariz (COI). Ligares, V-1884, J. de Mariz (COI). Valença, Olival de Santa Barbara, VI-1885, A.R. da Cunha (COI). Vila Real, Coêdo, 10VII-1981, A. Além (COI). SPAIN. Álava: Bernedo, Urturi, 1-VII-1987, J.A. Alejandre (MA 423674). Albacete: San Pedro, 11-VI-1984, J.M. Herranz (MA 318970). Almería: Abrucena, Las Rozas, 17-VI-1988, B. Valdés & al. (G 104090). Ávila: Arévalo, 20-VI-1971, J. Gómez (MA 432025). Castronuevo, 19-VI-1984, Barrera & al. (MA 314814, SALA 34644). Hoyocasero, Cueva del Moragato, 15-VI-1985, M. Luceño & P. Vargas (MA 407310). Hoyos del Espino, Las Chorreras, 9-VII-1988, Aizpuru & al. (MA 451362). Piedralaves, La Adrada, 22-VI-1982, F. de Diego Calonge (MA 538909, MA 538611). Poyales del Hoyo, 30VI-1917, J. Cuesta (MA 121528). Ramacastañas, Cerro de las Cuevas del Águila, 30-V-1987, Vargas (MA 655999). Sierra de Ojos Albos, Los Regajales, 3-VII-1984, A.R. Burgaz & al. (MA 389677). Sotillo de la Adrada, 15-VI-1973, G. López & E. Valdés Bermejo (MA 430972). Valle de Iruelas, 12-VI-1956, C. Vicioso (MA 170174). Venta del Obispo, 20-VI-1945, A. Caballero (MA 121586). Villanueva de Gómez, 19-VI-2003, M. Ladero (SALA 108263). Valle de Amblés, Villatoro, 12-VII-1974, Ladero & Fuertes (SALAF 23155). Badajoz: Campanario, V-1911, V. Lagares (MA 121534). Castuera, 14-IV-2001, P. Escobar García (MA 707044). El Berrocal, 25-V2001, P. Escobar García (MA 707042). El Toril, V-1951, Moreno Márquez (SEV 5005). Mérida, embalse de Proserpina, 16-IV-1994, F. Amich & al. (MA 717798). Oliva de la Frontera, 23-IV-1994, F. Amich & al. (MA 717195). Sierra del Palenque, 7-V-2000, P. Escobar Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 29 Taxonomic review of the Campanula lusitanica complex García (MA 766400). Talarrubias, 19-V-2001, P. Escobar García (MA 707043). Valdecaballeros, 17-III-1977, M. Figueroa & al. (MA 269851). Burgos: Aranda de Duero, V-1942, A. Caballero (MA 121572, MA 121526). Cardeñajimeno, 21-VI-1914, C. Pau (MA 121533). Ciruelos de Cervera, pie del Alto de la Cabeza, 11-VII1979, Pons Sorolla & Susanna (G 104082, MA 413025). De Quintanar de la Sierra a Neila, 1-VII-1925, M. Losa (MA 433608). La Revilla, camino de Ahedo, 17-VII-1979, Muñoz Garmendia & al. (G 104085, MA 414202). Miranda de Ebro, 15-VI-1912, H. Elías (MA 433606). Santa Gadea del Cid, 11-VII-1915, H. Elías, in F. Sennen, Plantes D’ Espagne, nº 2447 (MA 121573). Tejada, Pico Valdosa, 3VII-1979, J. Fernández Casas & al. (G 104084). Cáceres: Baños de Montemayor, 15-IV-1944, A. Caballero (MA 121543). Barrado, 19V-1988, A. Amor (SALAF 16729, SALAF 23633). Cañaveral, cerro de Cabezarrubias, 14-V-1988, Ladero & Santos (SALAF 16506). Casatejada, Las Cabezas, Sexta Suerte, Arroyo de Fresno, 7-V-1983, Ruiz Téllez (SALAF 6774). En las bajadas de Puerto Viejo, hacia Valverde del Fresno, 23-V-1982, A. Valdés Franzi (SALAF 12190). Guadalupe, 16-VI-1948, A. Caballero (MA 121539). Hervás, 12-X1987, R. González (SALA 101150). Jaraíz de la Vera, Las Costeras, 19-V-1988, A. Amor (SALAF 16703). La Bazagona, 1-V-1983, Ruiz Téllez (SALAF 6775). Losar de la Vera, Valle del Tiétar, 20-III1980, Meana & al. (MA 393441, SANT 18476). Mirabel, 4-V-1980, D. Belmonte (MA 344687). Montánchez, Sierra de Montánchez, carretera a Cerro Moro, 20-VI-1998, S. Castroviejo (MA 613395). Navalmoral de la Mata, 25-V-1984, Ruiz Téllez (SALAF 10761, MA 680829). Plasencia, 19-V-1988, Ladero & al. (SALAF 16507). Puerto de Hoyos, 17-VII-1978, A. Valdés Franzi (SALAF 12191). Puerto de Santa Clara, San Martín de Trevejo, 28-VI-1983, M. Ladero & A. Valdés (SALA 114203, SALAF 12192). Santiago de Alcántara, 26IV-1994, F. Amich & al. (MA 718573). Sierra de San Pedro, 17-V1909, M. Gandoger (G 104144). Torrecillas de la Tiesa, 22-V-1999, L. Medina (MA 624469). Valle del Jerte, 18-VI-1975, Carrasco & Castroviejo (SALA 25804). Cádiz: Algodonales, Sierra de Líjar, 11VII-1980, A. Aparicio & al (SEV 57983). Aprox. 5 km al E de Vejer de la Frontera, entre los pinos a lo largo de la carretera, 28-IV-1975, S. Holmdahl (MGC 50213). Bornos, 28-IV-1978, B. Molesworth Allen (SEV 53284). Chiclana de la Frontera, 22-V-1982, A. Charpin & C. Defferrand (G 104083, MA 243115). Grazalema, Puerto de las Palomas, 10-VI-1993, A. Aparicio & al. (MA 526987). Jeréz de la Frontera, 17-V-1985, A. Asensi & J. Cuenca (MA 570487). Los Barrios, Barranco del Arroyo de Valdeinfierno, 28-V-1978, J. Fernández Casas (G 104086, MA 226312, SALA 22564). Puerto de Santa María, Chiclana, et pr. Grazalema, V-1895, Porta & Rigo, Iter IV Hisp. 1895: 321 (G 104107). Sierra de Palma, 19-VII-1887, E. Reverchon, in E. Reverchon, Plantes de L’Andalousie, 1887: nº 17 (MA 121509). Tarifa, Sierra de Saladavieja, El Carrascal, 22-VII1980, J. Arroyo & J.M. Gil (SEV 64621). Ubrique, 30-V-1972, S. Holmdahl (MGC 50212). Cantabria: Valderredible, páramo de la Lora, 2-IX-1983, E. Loriente (MA 683841, MA 599290). Ciudad Real: Argamasilla de Calatrava, finca La Laguna de las Carboneras, 13-V-2001, M. Bellet & al. (MA 711976). Cabezarrubias del Puerto, 22-V-1998, R. García Ríos (MA 711940). Casas del Río, Navalagrulla, 1-VI-2001, S. Castroviejo & M. A. Carrasco (MA 692324). Sierra de Alhambra, 30-IV-1933, González Albo (MA 121583). Sierra Madrona, 29-V-1950, S. Rivas Goday & J. Borja (SALA 376). Sierra Morena, Venta de Cárdenas, 30-IV-1933, J. Cuatrecasas (MA 121520). Córdoba: Belalcázar, finca de Pedroche, 8-VII-1976, J.A. Devesa (SEV 33720). Cardeña, finca de Yegüerizo (UH-83), 30-V1976, J.A. Devesa (SEV 33723). Torrecampo, ribazos del río Guadamora, 16-V-1976, J.A. Devesa (SEV 33388). Trassierra, 14-V-1982, J. Arroyo (SEV 87091). Villaviciosa de Córdoba, Tres puentes, V1920, C. Pau (MA 121518). Cuenca: Ródenos de Cañete, 9-VI-1971, E. Valdés Bermejo & al. (MA 431991). Talayuelas, 18-VI-1979, G. Mateo (MA 256531). Granada: Cañar, Bco. río Chico, 20-VII1979, J. Molero Mesa (MA 432023). Capileira, 3-VII-1948, Vieiter 29 (SANT 211). In arenosis Regn. Granat. a littore, 1837, E. Boissier (G 104111). Jatar, Sierra Almijara, supra Jatar, 11-VI-1983, B. Cabezudo & J.M. Nieto (MGC 41527). Puerto de la Ragua, cruce con la carretera a la Alpujarra, 19-VI-1988, B. Valdés & al. (G 104089). Sierra Nevada, 21-VII-1879, Huter & al. Extinere hispanico 1879: 234 (G 104137); ibidem, Cáñar, 28-VII-1930, C. Vicioso (MA 121558). Guadalajara: Campillejo, VII-1973, Demetrio (FCO 4103). Cantalojas, Valle del Lillas, 20-VI-1985, Burgos & Cardiel (MA 487000). Checa, 25-VI-1997, L.M. Ferrero & L. Medina (MA 595634). El Pedregal, VII-1894, J. Benedicto (MA 121566). La Fuensaviñán, 26-V1994, J. Castillo & al. (MA 543882). Prado Redondo, Monte del Condado (MA 153134). Valverde de los Arroyos, 13-VII-1998, F. Lamata (MA 615546). Guadalajara, VI-1994 (MA 546347). Huelva: Almonte, 28-IV-1943, C. Vicioso (MA 121510); ibídem, 12-V-2006, A. Quintanar (MA 772195). Ayamonte, 5-V-1902 (MA 121515). Calañas, 28-IV-1921, Gros (MA 121513). Chucena, Cerro de las Palomas, 19-IV-2001, B. Cabezudo & al. (MGC 48566). Higuera de la Sierra, 24-V-1988, E. Bayón & E. Villanueva (MA 438689). Hinojos, 26-V-2004, J. Cano-Maqueda & al. (SEV 216214). La Barra de Huelva, 22-IV-1943, C. Vicioso (MA 121512). Sierra de Aracena, Aracena, 15-V-1979, J. Rivera (SEV 48340). Jaén: Andújar, El Abogado, 11-V-1985, E. Cano (MA 716641). Baños de la Encina, mina Matacabras, 23-IV-1988, C. Fernández & al. (FCO 21720, G 104155, MA 553899, MGC 38654, SALA 59048, SANT 29973). Guarromán, camino hacia la mina de los Dolores, 5-V-1966, S. Silvestre (SEV 19712). Sierra Morena, Arroyo de Oruga, 7-VI-1923, Fernández & al. (MA 121519). La Rioja: Ezcaray, barranco Reoyo, pr. Urdanta, 16-VII-1998, Gracía-Baquero & al. (SALA 100212). Ocón, 27-VII-1930 (MA 121531). San Millán de la Cogolla, 16-VII1981, S. Castroviejo & Fernández Quirós (MA 433427). Sierra de la Demanda, de Anguiano a Tabladas, 26-VII-1995, L. Loidi & A. Berastegui, in Lambinon, Pl. Europe Occid.-Bas. Méd., 1997: nº 17459 (G 104092, MA 589694). León: Castroquilame, 12-V-1973, Andrés & Carbó (SEV 16643). La Gotera, 8-VII-1944, Rojas (MA 121550). Ponferrada, 10-VII-1933, W. Rothmaler, in W. Rothmaler, Plantae Hispaniae Boreali-Occidentalis, nº 128 (MA 121551); ibidem, carretera de San Esteban de Valdueza a San Pedro de Montes, 21-VI1981, Alamillo & Nieto (MA 317259). Trabadelo, San Fiz do Seo, valle del río Barjas, 18-VII-1998, Martín-Blanco (MA 641637). Lugo: Tardad, Villalba, 30-VII-1951, M. Orosa (G 104101). VillardíazFonsagrada, 22-VII-1953, E. Carreira (G 104197). Madrid: Madrid, VI-1841, Reuter (G 104130, G 104131). Buitrago, 1-VI-1918, C. Vicioso (MA 121559). Collines à la base de la Sierra de Gredos, 8-VII1863, E. Bourgeau, in E. Bourgeau, Pl. d’Espagne et de Portugal, 1863: nº 2442 (G 104127, G 104132, MA 721360, MA 121530, MA 121529, MA 152777). Casa de campo pres Madrid, 7-VI-1854, E. Bourgeau (G 104128, G 104145, MA 720519). Chamartín, 17-V, Isern (MA 153136). Chinchón, cerros de Butarrón, VI-1919, C. Vicioso (MA 121576). El Escorial, VI-1914, C. Vicioso (MA 121577). El Pardo, 4-VI-1936, M. Martínez & A. Rodríguez (MA 432685). Guadalix de la Sierra, 29-VI-1983, F. Gómez Manzaneque (MA 450150). Hoyo de Manzanares, VI-1998, A. Izuzquiza & al. (MA 615516). La Pedriza, V-1932 (MA 432723). Miraflores, 25-VI-1945, L.C. & A.R. (MA 201400). Monte del Pardo, 20-V-1917, C. Vicioso (MA 121578). Montejo de la Sierra, 2-VII-1954, A. Rodríguez (G 104103). Navacerrada, VI-1915, C. Vicioso & Beltrán (MA 121580). Puerto de la Cruz Verde, 22-VI-1973, Rivas-Martínez & Costa (MA 432036). Robledo de Chavela, 19-VI-1988, M. Costa Tenorio & H. Sainz Ollero, in J. Lambinon, Pl. Europe Occ.-Bas. Méd. 1993: nº 15575 (G 104153, MA 532060, SALA 88527). San Martín de Valdeiglesias, Rozas de Puerto Real, 6-VI-1992, P. Vargas (MA 515427). Sierra de Guadarrama, 4-VII-1968, O. Polunin (MGC 5003). Sierra de la Cabrera, 4-VII-1993, A. Izuzquiza & al. (MA 529567). Torrelaguna, V-1912, C. Vicioso (MA 121582). Málaga: Benaoján, Sierra del Palo, 27-V-2007, J. Cano-Maqueda (SEV 218872). Benarrabá, carril de los Pepes, 29-V-2004, O. Gavira Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 30 30 J. Cano-Maqueda & S. Talavera (MGC 60964). Casares, Monte del Duque, 20-V-1988, B. Cabezudo & A.V. Pérez-Latorre (MGC 35466). Estepona, 16-V-1919, E. Gros (MA 121506). Gaucín, VI-1916, E. Gros (MA 121508). Gobantes, garganta de El Gaitán, 11-VI-1930, C. Vicioso (MA 121486). Los Villares, 12-V-1988, B. Cabezudo & al. (MGC 24576). Ronda, 07VI-1889, E. Reverchon, in E. Reverchon, Plantes de L’Andalousie 1889: nº 17 (G 104260). Sedella, Los Picaricos, 30-V-2003, B. Cabezudo & al. (MGC 60070). Sierra Bermeja, 18-V-1919, E. Gros (MA 121507). Sierra Tejeda, 26-VI-1982, J.M. Nieto (MGC 18110). Tolox, Castañar de los Hornillos, 13-VI-1932, L. Ceballos (MA 121505). Torrox, VI-1909, Domingo (G 104138). Orense: Lovios, 6-VI-1993, I. Pulgar (FCO 21431, MA 551003). Serra do Invernadeiro, entre Rocín y Suacenza, 11-VII-1973, S. Castroviejo (G 104080). Salamanca: Cantalapiedra, 25-V-1987, X. Giráldez & Aragón (SALA 46173). Ciudad Rodrigo, 4/6-VII-1932, C. Pau (MA 121549). Embalse del río Águeda, 6-VI-1976, E. Rico (SALA 9431). Entre Fuentes de Béjar y Navas de Béjar, 21-VI-1978, J.A. Devesa & J. Pastor (SEV 39776). Entre La Alberca y Sotoserrano, 20-VII1972, Fernández Díez (SALA 5451). Guijuelo, 7-VI-1987, E. Rico & J. Serradilla (MA 476539, SALA 46836). Hurdes, 5-VII-1946, A. Caballero (MA 121547). Ledesma, 19-V-1976, J. Sánchez (SALA 17419). Linares de Riofrío, VII-1980, J. L. Fernández Alonso (MA 519040). Masueco, laderas del río Uces, 16-VI-1976, F. Amich (SALA 15517). Monleras, 22-X-1976, J. Sánchez (SALA 17428). Montemayor del Río, 3-VII-1983, A. Guillén (SALA 36167). Navacarros, 15-VII-1983, F. Amich & F. Herrero (SALA 34909, SALA 34910). Pelabravo, 2-VI-1990, A. Pastor (SALA 57034). Peñamecer, 23-V-1976, J. Sánchez (MA 219749, SALA 17427). Puerto Seguro, 9-V-1976, E. Rico (SALA 9432). San Esteban de la Sierra, 20-VI-1971, Fernández Díez (SALA 5595). Topas, 9-VI1967, B. Casaseca (SALA 375). Valdelageve, 28-IV-1996, J. Barrios Pérez (SALA 121888, SALA 121889). Segovia: Aguilafuente, Cotarra de Juriñas, 6-VI-2002, M. Pérez Valero (MA 756780). Entre Riaza y Ayllón, cruce de la carretera hacia Pajares del Fresno, 28-V1992, A. Izuzquiza (MA 508024). Fresno de la Fuente, 20-VI-1985, A. Izuzquiza (MA 348898). Fuentidueña, 15-VI-1983, T. Romero (SALA 40297). Lastras de Cuéllar, Molino Ladrón, 24-V-1998, P. Bariego Hernández & A. Gastón González (MA 754632). Ochando, 19-VI-2002, M. Sanz Elorza (MA 687112). Puerto de los Cotos, 5VII-1989, A. Charpin & P.A. Loizeau (G 104151). Riofrío de Riaza, 7-IX-1975, F.J. Fernández Casas (MA 347715). Siguero, 1-VII-1983, T. Romero (MA 567697). Torrecilla del Pinar, 12-VII-1984, T. Romero (SALA 40298). Sevilla: al S de Villamanrique de la Condensa, 7-V-1988, E. Bayón & al. (MA 438338). Aznalcázar, 20-IV-2003, Z. Díaz (MA 707168). Castilblanco de los Arroyos, Los Melonares, 1-VI-2006, J. Cano-Maqueda & al. (SEV 218870). Constantina, Hermita Virgen de Robledo, 1-VI-2006, J. Cano-Maqueda & al. (SEV 218869). Dehesa de Covarrubias, 6-IV-2001, B. Cabezudo & al. (MGC 48568). Gerena, 3-V-2001, B. Cabezudo & al. (MGC 48568). Paradas, 5-V-1933, C. Vicioso (MA 121517). Soria: Berlanga de Duero, 18-VI-1977, A. Segura Zubizarreta (G 104087, MA 226315, SALAF 437, SEV 74840). Canredondo, 22-VI-1990, A. Segura Zubizarreta (MA 581101). Cañón del río Lobos, 30-VI-1983, A. Buades (MA 504440). Carbonera de Frentes, 2-VII-1989, A. Charpin (G 104152). Herrera de Soria, 30-VI-1983, A. Buades (MA 504408). Molinos de Duero, 21-VIII-1987, G. Mateo (MA 383713). Quintana Redonda, 30-VI-1974, A. Segura Zubizarreta (SEV 41098). Tozalmoro, 6-VII-1935, C. Vicioso (MA 121527). Teruel: Orihuela del Tremedal, 18-VI-1907, C. Pau (MA 433607). Segura de los Baños, VII-1894, J. Benedicto (MA 121566b). Toledo: Almorox, El Pinar, 3-VII-1982, M. Luceño (MA 430416). Nacimiento del río Estena, 19-VI-1986, J. Assens & al. (MA 430409). Valladolid: Castronuño, Sendero de los Ladrones, 22-V-1988, G. Balbás (SALAF 23342). Pedrajas de San Esteban, 7-VII-1975, F.J. Fernández Díez (FCO 4601, SALA 7575). Zamora: Aspariegos, La Salgada, 10-V-1990, R. García Ríos (SALA 54046). Corrales, 15-VI-1951, Casaseca (SANT 5565). Cubo del Vino, 19-VI-1981, X. Giráldez (SALA 31612). Escober, Los Linares, 15-VII-1996, P. Bariego Hernández (MA 651736). Lago de Sanabria, 25-VIII-1953, A. Rodríguez (MA 201347). San Cebrián de Castro, El Barrucal, 28-V-1990, R. García Ríos (SALA 54048). Villaseco del Pan, La Era del Campo, 30-IV1988, R. García Ríos (SALA 54047). Zaragoza: Atea, 29-V-1909, C. Vicioso (MA 121565). Calatayud, VI-1987, C. Vicioso (MA 121560). Moncayo, 19-VII-1893, C. Vicioso (MA 121560). Belmonte, 4-V1931, Gros, in F. Sennen, Plantes d’ Espagne 1932: nº 8206 (G 104149, MA 121514). 3. Campanula cabezudoi Cano-Maqueda & Talavera in Acta Bot. Malacitana 32: 254. 2007 C. decumbens var. pseudospecularioides G. López in Bol. Soc. Brot. ser. 2, 53: 301 (1979-1980) [syn. subst.]. Ind. loc.: “Habitat locis umbrosis, inter rupes calcareas, loco dicto Boquete de Zafarraya, Sierra Tejeda (Málaga-Granada), ubi cum P. Cubas et J.M. Moreno, die 28-VI-1976, legi. Holotype: MA 210975, Fig. 7; Isotype: MA 210994”. Illustrations: Gallego (1987: 564, as C. decumbens); Fig. 7. Holotype of Campanula cabezudoi (MA 210975). Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 31 Taxonomic review of the Campanula lusitanica complex Sáez & Aldasoro (2001: 130, fig. 39a, as C. lusitanica subsp. lusitanica); Fig. 4 E, F. Herb 8-30(45) cm, annual, decumbent, branched from the base, pubescent, not brittle. Stems slightly angled, branched, pubescent, sometimes glabrescent at the apex, with setose hairs of 0.1-1.4 mm. Leaves subcoriaceous, entire or toothed, petiolate; middle cauline leaves 6.2-18.2 × (2.9)5-9 mm, petiolate, elliptical, ± pubescent, with hairs to 1.1 mm, with petioles up to 2 mm; upper cauline leaves 4.5-14.5 × 1.3-3.9 mm, oblanceolate, ± hairy, with hairs up to 1.2 mm, with petioles of c. 0.5 mm. Inflorescence laxly paniculate. Flowers pedicellate; pedicel (9.8)20-68 mm, glabrous or setose, with hairs of 0.1-0.7(1.5) mm. Calyx-teeth (4)5-11 × 0.4-1.2 mm, lanceolate or linear. Corolla (7.3)8-16 mm, infundibuliform, with the tube shorter than the lobes; tube 3.1-6.9 mm, white; lobes (3.6)5-10.3 × 3.1-6.2 mm, elliptic, blue, with three purple nerves. Stamens with enlarged base of 0.6-1 × 0.5-0.7 mm, deltoid; filaments 0.4-0.5 mm; anthers (2.5)3-6.4 mm, whitish. Ovary densely hispid, with setose hairs 0.2-1.8 mm; style (5)7-12.3 mm, hairy in the 31 upper half; stigma trifid, with stigmatic branches 11.7 mm, patent, arched, white. Capsule 2.1-5.9 × 2.25 mm, subspherical or ovoid, generally wider than long, densely hairy, with papillose hairs up to 1.5 mm, with 10 nerves ± acute but not winged, dehiscing by three apical pores. Seeds 0.5-0.6 × 0.2-0.3 mm, ovoid, shining, yellowish to brown. 2n = 20. Habitat, phenology and distribution: Fissures of calcareous rocks; 400-1600 m. VI-VII. • Endemic to the Sierra Subbética from Seville to Jaén, and the Penibética, where it is very frequent, especially in the Sierra de Loja and Tejeda (Fig. 8). Spain: Ca Co Gr J Ma Se. Observations: Sáez & Aldasoro (2001) comment with reference to C. decumbens var. pseudospecularioides G. López: “existen formas intermedias entre esta subespecie [which they refer to C. specularioides] y C. decumbens [which they refer to C. dieckii], que han recibido reconocimiento taxonómico”. It is very likely that they are referring to C. cabezudoi. In fact, the vegetative characters of decumbent habit and the presence of petiolate middle leaves of the stem, Fig. 8. Distribution map of Campanula cabezudoi (䊱) and C. specularioides (䡬). Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:09 Página 32 32 J. Cano-Maqueda & S. Talavera are similar in C. cabezudoi and C. decumbens, but C. cabezudoi has hairy style in the upper half and a single trifid stigma with curved and patent stigmatic lobes, while C. decumbens has a glabrous style and three straight and erect-patent stigmas. Moreover, in the topology of the ITS analysis, C. cabezudoi and C. decumbens are in two different clades (Fig. 1) and they have different chromosomes number (Table 1, Fig. 2). Selected specimens SPAIN. Cádiz: Grazalema, Puerto de las Palomas, El Pinar, 5VII-1984, A. Aparicio (SEV 161693). Córdoba: Almedinilla, Sierra de Albayate, 7-VI-1980, J. Muñoz (MA 749306). Priego de Córdoba, Sierra Horconera, 4-VII-1980, J. Muñoz & R. Tormo (MA 749309). Rute, Sierra de Rute, Pico Las Cruces, 26-VI-1978, J. Muñoz (MA 749308, SEV 161894). Granada: entre Venta de Zafarraya y Zafarraya, Sierra Gorda, VI-2007, Cano-Maqueda & al. (SEV 218873). Sierra Harana, cercanías de la Cueva del Agua, 16VI-1982, Casares & al. (MA 432054). Sierra Tejeda, 7-VII-1935, M. Laza (MA 121470). Jaén: Mágina, 3-VII-1925, J. Cuatrecasas (MA 121522). Sierra de Cazorla, El Tranco, 30-VI-1988, S. Talavera & al. (G 104061, SEV 161662). Málaga: Alfarnate, Sierra de la Torca, 7-VI-2006, B. Cabezudo & al. (MGC 63804). Álora, Sierra de Huma, 26-VI-1986, B. Cabezudo & R. Suan (MGC 36711). Antequera, 14-VI-1930, C. Vicioso (MA 121488). De Ronda a El Burgo, 18-VI-1972, L. Bernardi (G 104157). Los Alazores, Puerto de los Alazores, Sierra de Alhama, Tres Mogotes, 5-VII-1973, B. Cabezudo & B. Valdés (SEV 161891). Puerto del Viento, 29-VI1849, E. Bourgeau in Bourgeau, Pl. Espagne 1849: nº 320 (G 104256). Sierra Prieta, 07-VI-1879, Hunter, Porta & Rigo in Iter. Hisp. 1879: nº 232 (G 104184). Ronda-Sierra de las Nieves, entrando al Sabinar, Monte de la Peineta, 19-VI-1974, S. Talavera & B. Valdés (SEV 161889). Sierra de Peñarrubia, 12-VI-1930, C. Vicioso (MA 121487). Valle de Abdajalís, cortado del Cuervo, 15-VI1973, S. Talavera & B. Valdés (SEV 161888). Villanueva del Rosario, Sierra Camorolos, 5-VII-1973, B. Cabezudo & B. Valdés (SEV 161887). Yunquera, P. N. Sierra de las Nieves, entre el Puerto Bellina y el cruce del camino con la Cañada de los Hornillos, 18VI-1998, B. Cabezudo & al. (MGC 46750). Sevilla: Algámitas, Sierra del Tablón, 5-VII-1978, B. Cabezudo & E. Ruíz de Clavijo (SALA 13425, SEV 31778). angled, branched, glabrous, rarely with setiform hairs to 1.2 mm. Leaves fleshy, entire or toothed, petiolate; middle cauline leaves 7.5-13.4 × 4.5-20 mm, broadly elliptical, glabrous or glabrescent, with hairs 0.1-0.2 mm, and petiole 1-6.4(14) mm; upper cauline leaves 3.5-14.5 × (1)1.9-6.4 mm, elliptic, glabrous, with petiole 0.4-1.7 mm. Inflorescence laxly paniculate. Flowers pedicellate; pedicel (5.7)7-35 mm, glabrous or setose hairs 0.1-0.8 mm. Calyx-teeth (1.7)3-7.6 × 0.7-1.5 mm, oblanceolate. Corolla (6.6)8-10(14) mm, broadly infundibuliform, with the tube shorter than lobes; tube 2.6-4.9 mm, whitish; lobes 4.1-7.5(10.3) × 2.25.4 mm, elliptical, pinkish or bluish, with three purple nerves. Stamens with enlarged base of 0.5-1 × 0.2-0.5 mm, deltoid; filaments 0.6-0.8 mm; anthers 2.1-3.2 mm, blue or whitish. Ovary glabrous, rarely densely hairy; style (4.7)6-7.3(8.5) mm, hairy in the upper half; trifid stigma, with stigmatic branches 0.6-1(1.4) mm, patent, curved, white or blue. Capsule 2-3 × 3-4.5 mm, subspherical, wider than long, with 10 subwinged, glabrous nerves, rarely with setose hairs up to 1.5 mm, dehiscing by three pores of middle position. 4. Campanula specularioides Coss., Notes Pl. Crit.: 41. 1849 C. lusitanica subsp. specularioides (Coss.) Aldasoro & L. Sáez in Anales. Jard. Bot. Madrid 59: 173. 2001. Ind. loc.: “In fissuris rupium regionis montanae regni Granatensis, loco dicto Cortijo blanco prope Ronda, a Bourgeau inventa”. Type: Spain. Málaga, Ronda, Cortijo Blanco, 20 June 1849, Bourgeau s.n. [Pl. D’Espagne, 1849] (lectotype, here designated, P 185468!, Fig. 9; see observations). Illustrations: Gallego (1987: 565); Sáez & Aldasoro (2001: 130, fig. 39i-m, as C. lusitanica subsp. specularioides); Fig. 10 A, B. Herb 6-26 cm, annual, decumbent, branched from the base, glabrous or glabrescent, very brittle. Stems Fig. 9. Lectotype of Campanula specularioides (P 185468, herb. Cosson). Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 33 Taxonomic review of the Campanula lusitanica complex Seeds 0.5-0.7 × 0.2-0.3 mm, ovoid, shining, yellowish to brown. 2n = 20; n = 10. Habitat, phenology and distribution: In crevices in walls or limestone rocks; 500-1650 m. V-VI(VII). •En- 33 demic to Sierra de Grazalema and Serranía de Ronda (Fig. 8). Spain: Ca Ma. Observations: Despite its narrow distribution, this species is markedly variable in the colour of anthers Fig. 10. Flowers and fruits of Campanula species. A, B, C. specularioides (Montejaque, Málaga, Spain, SEV 218871); C, D, C. transtagana (Valverde del Camino, Huelva, Spain, SEV 216212); E, F, C. broussonetiana (Jbel Tazzeka, Taza, Morocco, SEV 216476). The scale bar = 3 mm. Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 34 34 J. Cano-Maqueda & S. Talavera and stigmas, and in the indumentum of calyx and capsule. since both can be very hairy or completely glabrous, even in the same population, although more commonly all plants of a population have glabrous calyx, and likewise the capsule. Within populations there are plants with white anthers and stigmas, and others with blue anthers and stigmas, or even plants with white anthers and blue stigma or vice versa. All these colour morphs, which certainly have a genetic basis, seem to be interfertile so this trait has little taxonomic value. Based on the middle position of the pores in the capsule, this species was included by Fedorov (1976) in the sect. Campanula. The phylogenetic tree of nr DNA ITS shows that C. transtagana and C. broussonetiana are sister species of C. specularioides (see Fig. 1). The type material of C. specularioides consists of three complete plants and several fragments. The largest of the three plants, placed at the top of the sheet was chosen as lectotype, since it is consistent with the description of E. Cosson (see Fig. 9). In the general herbarium of the “Conservatoire et Jardín Botanique de Genève” there is a sheet (G 104263) with several plants of the same collection of E. Bourgeau, that are also possible type materials. C. lusitanica subsp. transtagana (R. Fern.) Fedorov in Bot. J. Linn. Soc. 67: 281. 1973. Ind. loc.: “Habitat in Lusitania, regione Transtagana, ad marginem sinistram fluminis Tagis pr. pagum dictum Vila Velha de Ródâo, ubi super declives solo argilloso et sicco inter sepes copiosa, 21-VI-1959, A. Fernandes, J. Matos & A. Sarmiento 2923 (COI, holotype)”, (Fig. 11; see observations). C. loeflingii var. filiformis Lange in Vidensk. Meddel. Dansk. Naturhist. Foren. Kjøbenhavn 1861: 108. 1862. Ind. loc.: “La Carolina (Sierra Morena) 10 Mai.”. Type : not found in the Lange herbarium (C). Illustrations: Fig. 10 C, D. Herb 6-45 cm, annual, decumbent or erect, branched from the base, rarely in the upper half, very laxly pubescent, not brittle. Stems angled, branched, glabrescent, with hairs 0.1-0.2, commonly very laxly located in the stem angles. Leaves not coriaceous, crenate, entire or toothed, petiolate; middle cauline leaves (4.6)9-20(30) × 2.2-8(13.2) mm, elliptic, laxly pubescent, with hairs 0.1-0.3 mm scattered over the limb and margin of the leaf, and with petiole 0.2-4.5 Selected spcimens SPAIN. Cádiz: Benaocaz, Km 16-17 a Ubrique, 22-VI-1984, A. Aparicio & J. G. Rowe (MA 490917, SEV 161738). Entre Benaocaz y Ubrique, 13-VII-1978, J. Devesa & al. (MA 465708, MA 111772, SEV 103546). Entre Ubrique y Grazalema, 13-VI-1970, E.F. Galiano & B. Valdés (SEV 108955). Entre Villaluenga del Rosario y Benahocaz, 26-VI-1988, Férnandez Díez & Mochales (SALA 47474, MA 476540). Grazalema, 21-VI-1890, E. Reverchon, in E. Reverchon, Plantes de L’ Andalousie, 1890: nº 331 (G 104191). Idem, 9-VII-2003, J. Cano-Maqueda (SEV 216211). Cerro de San Cristóbal, V-1961, J. Borja (MA 177095, SEV 5016). Manga de Villaluenga, 22-VI-1983, A. Aparicio & J.G. Rowe (SEV 161737). Sierra de la Silla, 21-VI-1983, A. Aparicio & S. Silvestre (G 104188, MA 490918, SEV 161731). Sierra de Zafalgar, Puerto de la Miera, 28-VI-1984, A. Aparicio & al. (SEV 161726). Sierra del Caillo, Navazo Alto, 30-VI-1983, A. Aparicio (SEV 161730). Sierra del Endrinal, Pozo de las Presillas, 12-VII-1984, A. Aparicio & S. Silvestre (SEV 161736). Ubrique, 26-VI-1925, P. Fonti Quer & E. Gros (MA 702195, MGC 53268, SALA 114797). Villaluenga del Rosario, 24-VI-1973, A. Asensi & B. Diez (MGC 81). Málaga: Benaoján, 17-VI-2007, J. Cano-Maqueda (SEV 218871). Cartajima, Cancha Almola, 12-VI-2004, O. Gavira (MGC 60974, MGC 60975). De Ronda a Montejaque et circa Cueva de la Pileta, 17/18VI-1972, L. Bernardi (G 104194, MA 269887). Jimera de Líbar, Alto del Conio, 4-VII-2004, O. Gavira (MGC 60868). Montejaque, 8-VI-2005, J. Cano-Maqueda (SEV 216210). Serranía de Ronda, 23-VII-1888, E. Reverchon, in E. Reverchon, Plantes de L’Andalousie, 1889: nº 331 (G 104264, G 104185, G 104186, G 104187, G 104192, G 104193, LISU 50130, MA 121476, MA 121477). 5. Campanula transtagana R. Fern. in Bot. Soc. Brot. ser. 2, 36: 121-126. 1962 Fig. 11. Holotype of Campanula transtagana (COI). Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 35 Taxonomic review of the Campanula lusitanica complex mm; upper cauline leaves (3)5-11(15) × 0.6-3.4 mm, elliptic, laxly pubescent, with hairs 0.1-0.3 mm scattered over the limb and leaf margin, and with petiole 0.2-0.3 mm. Inflorescence laxly paniculate. Flowers pedicellate; pedicel 9.6-64 mm, glabrous or with some hairs of c. 0.5 mm. Calyx-teeth (2.9)3.5-16.6 × 0.3-0.7 mm, linear. Corolla (6.5)8-12(14) mm, campanulate, with the tube greater or equal, rarely shorter, than the lobes; tube (2.9)3.7-7 mm, light blue with a white base; lobes 3.6-7 × 2.2-3.5 mm, elliptic, bluish. Stamens with enlarged base of 0.5-1 × 0.3-0.6 mm; filaments 0.2-0.6 mm; anthers 1.6-2.7 mm, whitish. Ovary glabrous, rarely with setose hairs 0.1-0.2 mm; style 4.5-6.4 mm, hairy in the upper half; stigma trifid, with estigmatic branches 0.6-1.2 mm, patent, curved, white. Capsule 2-4.4 × 1.5-3 mm, subovoid or subspherical, longer than wide, glabra, rarely with some setose hairs of 0.1-0.2 mm, with 10 angled nerves, but not winged, dehiscing by three pores of middle or subapical position. Seeds 0.4-0.6 × 0.1-0.2 mm, ovoid, shining, yellowish to brown. 2n = 20; n = 10. Habitat, phenology and distribution: Wet grasslands on sandy or slate-rich substrates; 50-800 m. V-VI (VII). • Endemic to SW Iberian Peninsula, from the River Tajo to the Guadiana, Sierra de Guadalupe and almost all Sierra Morena (Fig. 12). Portugal: AAl Ag BAl BB. Spain: Ba Cc Co H J Se. Observations: C. transtagana is rare in Portugal and in Sierra de Guadalupe but very common throughout the Sierra Morena, especially in shady slopes and valley bottoms, where it normally co-habits with C. matritensis. The profuse branching, small corollas with white background tube and petiolate leaves of the stem, clearly differentiate it from C. matritensis. The holotype of C. transtagana consists of five cultivated plants that are very branched from the base, between 20 and 35 cm, and placed on three sheets in Coimbra herbarium. 35 bar García (MA 707042). Oliva de la Frontera, riberas del río Ardilla, 23-IV-1994 VI itinera Mediterranea nº 557 (MA 717195). Cáceres: Guadalupe, 31-V-1958, E. Guinea (MA 432639). Córdoba: Alcolea, margen derecha del Guadalquivir, 21-V-1987, Z. Díaz & C. López (SEV 130952). Cardeña, 24-VI-2005, J. Cano-Maqueda & al. (SEV 216480). Entre Azuel y el río de las Yeguas, 28-V-1982, J. Devesa & García (SEV 161896). Entre Montoro y Adamuz, proximidades a Montoro, 4-V-1982, M.J. Díez & I. Fernández (SEV 161898). Río Zújar, cruce con la carretera Los BlázquezPeraleda de Zaucejo, 6-VI-1979, J.M. Muñoz & E. Ruiz de Clavijo (SEV 161897). Trassierra, derecha del Guadiato, Cerro del Trigo, pantano de la Breña, 16-VI-1978, J.A. Varela (SEV 161900). Villafranca, central eléctrica, río Guadalquivir, 21-V-1987, Z. Díaz & C. López (SEV 130876). Huelva: Alosno, 26-V-1942, C. Vicioso (MA 121571). Aracena, entre la Cefiña y la N433, 23-V-2005, J. Cano-Maqueda & al. (SEV 216213). Corterrangel, 18-VI-1978, J. Rivera (SEV 48336). Entre Aroche y Las Cantiendas, 6-VI-1979, J. Rivera & B. Cabezudo (MGC 8832, SEV 48900). Escacena del Campo, Reserva de la Pata del Caballo, arroyo del Chacho, 31-V-2001, B. Cabezudo & al. (MGC 48565). Higuera de la Sierra, 400 m, 24-V-1988, E. Bayón & E. Villanueva (MA 438691). Linares de la Sierra, 22-VI-1942, C. Vicioso (MA 121570). Río Múrtiga, 8-VI-1984, F.J. García & al. (G 104154). Valverde del Camino, El Manzanito, 23-V-2005, J. Cano-Maqueda (SEV 216212). Jaén: Andújar, Arroyo de los Santos, 22-VII-1992, J.M. Mancebo (MA 651609). Baños de la Encina, Charca de la Enea, 29-VII-1992, J.M. Mancebo & J.R. Molina (MA 651610). Despeñaperros, 1-VII-1975, J. Fernández Casas & al. (MA 412612). Sevilla: Castilblanco de los Arroyos, 1-VI-2006, J. Cano-Maqueda & al. (SEV 232786). El Castillo de las Guardas, V-1914, C. Vicioso (MA 121569). El Garrobo, junto a la carretera, 15-V-1982, J.M. Rodríguez & al. (SEV 161950). Entre el Ronquillo y Almadén de la Plata, 22-VI-1976, E.F. Galiano (SEV 161899). Guillena, Arroyo Herrero, without date, C. Fernández & J.A. Fariña (SEV 109874). Selected specimens PORTUGAL. Algarve: Serra da Picota, VII-1891, J. Brandeiro (COI). Alto Alentejo: Redondo, V-1892, A. Fernandes & al. (COI). Idem, VI-1893, A. Fernandes & al. (COI). Riveira da Sapatoa, a 5 km de Montoito e a 16 de Reguengos de Monsaraz, 10-VI1962, A. Fernandes & al. (COI). Baixo Alentejo: Concelho de Moura, margens do rio Guadiana, 2-IV-1999, P. Bringe & T. Rego, in Exsiccata Flora Iberomacaronesica Selecta, centuria IV: nº 340: (FCO 25015, MA 632684, SALA 99329, SANT 42025). Entre Eiras Altas e Claúdia, 11-VI-1962, M. da Silva (LISE 76991, MA 199926). Margem da ribeira de Chança, prox. Vila Verde de Ficalho, 11-VI-1962, A. Fernandes & al. (COI). Beira Baixa: Encosta da margem esquerda do Tejo, entre a estrada e o rio Tejo, 8-VI-1962, A. Fernandes & al. (COI). Salvaterra do Extremo, 14-VI-2005, A. Charpin (G 104159). SPAIN. Badajoz: Don Benito, La Zafrilla, 28-VI-1985, M.J. Gallego (SEV161715). Magacela, El Berrocal, 25-V-2001, P. Esco- Fig. 12. Distribution map of Campanula transtagana (䡬) and C. broussonetiana (䊱). Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 36 36 J. Cano-Maqueda & S. Talavera 6. Campanula broussonetiana Schult. in Roem. & Schult., Syst. Veg. 5: 104. 1819-1820 C. lusitanica var. broussonetiana (Schult.) Pau in Mem. Real Soc. Esp. Hist. Nat. 12: 356. 1924. Ind. loc.: “In Mogador. Broussonet”. Type: Morocco. Safi, Mogador, Broussonet s.n. (lectotype: here designated, W 03798!; see observations). C. loeflingii var. maura Murb., Contr. Fl. Maroc. 2: 50. 1923. Ind. loc.: “Pentes herbeuses entre Amismiz et Oucheffine; Ighen Draa près Demnat. 900-1000 m.- J’ai vu la même plante des environs de Casablanca (leg. Mellerio)”. Type: not studied. C. lusitanica f. pallidiflora Maire in Jahand. et Maire, Cat. Pl. Maroc: 735. 1934. Ind. loc.: “Mont Tazzeka (Humbert et Maire)”. Type: not studied. C. lusitanica f. tenuis Caball. in Trab. Mus. Ci. Nat. Madrid, Ser. Bot. 30: 5. 1935. Ind. loc.: “Legi basi montis Tamarrut dicto, 6-VII-1934”. Type: Morocco. Agadir, Tamarrut, 6 July 1934, Caballero s.n. (lectotype, here designated, MA 121492!; see observations). C. vincaeflora Pau in Bol. Soc. Esp. Hist. Nat. 21: 278. 1921, nom. illeg., non Vent. 1804. Ind. loc.: “Tiguisar, 26-IV[-1921]”. Type: Morocco. Tittáguen, Tiguisar (Gomara), 26 April 1921, Vidal López s.n. (lectotype, here designated, MA 121493!; see observations). Illustrations: Fig. 10 E, F. Herb 6-45 cm, annual, erect or decumbent, branched from the base, pubescent, not brittle. Stems angled, branched, pubescent, rarely glabrous at the apex, with long hairs up to 0.4-2 mm. Leaves scarcely coriaceous, crenulated, entire or toothed; middle cauline leaves 8-37 × (4)6-15.5 mm, broadly elliptical or obovate, sessile and cuneate or with petiole up to 2 mm and rounded at the base, entire or toothed, ± pubescent, with hairs up to 1.2 mm; upper cauline leaves (4.2)6-13.5 × 1-7.6 mm, elliptical or lanceolate, generally entire, sessile, cuneate, pubescent, with hairs up to 0.4 mm or rarely glabrous, without a differentiated petiole. Inflorescence laxly paniculate. Flowers pedicellate; pedicel 7-67 mm, glabrous or with hairs 0.21.2 mm. Calyx-teeth (3.6)5.2-13.5 × (0.4)0.7-2(2.7) mm, oblanceolate, rarely linear. Corolla (7.2)8.8-17 mm, campanulate, with the tube usually shorter or equal in length to the lobes, rarely longer than lobes; tube 4,1-8,3 mm, with the base whitish and the apex light blue; lobes (2.4)3.5-8 × 2.1-5,2 mm, ellipticlanceolate, bluish. Stamens with enlarged base of 0.60.7 × 0.4-0.6 mm; filaments 0.6-0.7 mm; anthers 2.1-4 mm, bluish or white. Ovary glabrous, rarely with hairs 0.1-0.8 mm; style 5-8.2 mm, hairy; stigma trifid, with stigmatic branches of 1-2 mm, patent, curved, white or blue. Capsule 3-6.6 × 2-4.1 mm, subovoid, longer than wide, glabrous, rarely with setose hairs, with 10 nerves ± angled but not winged, dehiscing by three subapical pores. Seeds 0.4-0.6 × 0.2-0.3 mm, ellipsoid, shining, yellowish to brown. 2n = 20. Habitat, phenology and distribution: Shady hollows and the understory of cork woodlands, on limestone, or slate-rich or sandy substrates, from sea level to high mountains; 0-2000 m. IV-VII(VIII). • Endemic to W and N Morocco, along most of the Atlantic coastlands from Sidi Ifni to the Mamora forests; also the Rif, Middle Atlas and Great Atlas (Fig. 12). Observations: This species has the same color morphs for anthers and stigmas as those found in C. specularioides, and similary, the morphs seem to be interfertile. Alphonse De Candolle (1830) syno nymized this species with C. loeflingii Brot., possibly influenced by the illustration 18 of the “Phytographia Lusitaniae Selector” (Brotero, 1816) under this name. Certainly, C. lusitanica and C. broussonetiana are very similar morphologically. Both have long hairy indumentum that is straight and patent on the stem, and wide leaves, but in C. broussonetiana the middle cauline leaves are attenuate at the base or petiolate and in C. lusitanica are sessile and subauriculate. The molecular phylogeny has shown that C. broussonetiana is more closely related to C. transtagana than any other species of sect. Rapunculus (see Fig. 1). The sheet of type material of C. broussonetiana (W03798) contains a complete highly branched plant with many flowers. Schultes (1819-1820: 104) already indicated that the type material was in the Willdenow herbarium under the name Campanula ramosissima. In the herbarium of the Prodromus of De Candolle there are two sheets (G 138472 and G 138271G 138478), one (G 138472) with a complete plant of c. 30 cm and the other (G 138271, G 138478) with three smaller plants (15-22 cm), all belonging to the expedition by Broussonet in 1804, collected at Mogador (Essaouira, Morocco). This last sheet contains two labels. In one (G 138478), handwritten by A. De Candolle, is indicated “Voyage de Broussonet 1804” at the base of two of the plants, and in the other (G 138271), with orthography of Broussonet, is written: “Campanula/Mogador” and also, with orthography of Alphonse De Candolle, “Campanula loeflingii Brot./ _ Broussonetiana Roem. Et Sch./ A. DC.”. These materials were probably not studied by Schultes, and therefore they can not be considered type material (Fig. 13). The sheet of type material of C. lusitanica f. tenuis contains two complete plants with flowers in postan- Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 16/06/2011 11:25 Página 37 Taxonomic review of the Campanula lusitanica complex thesis and two single flowered small stems. The larger sized plant (c. 20 cm) is chosen as lectotype. The sheet also contains three determinavit labels, one handwritten by C. Pau, which reproduces all the information of the name, the diagnosis and the protological description from Caballero; the second label was identified as C. lusitanica by Yvonne Nyman in 1987 and the third label by Juan José Aldasoro and Llorenç Sáez who identified the specimen as C. broussonetiana The type material of C. vincaeflora consists of a single whole plant in flower that we have chosen as lectotype. The sheet contains some other stems and a label printed with the original description. Selected specimens MOROCCO. Agadir: Col du Kerdous (Anti-Atlas), 26-V1980, A. Charpin & al. (G 104179, MA 227542). Ida Ouehembal, Sud-ouest du Maroc, 1875, Mardochée (G 104254, G 104168, G 104255). O. Querat, Sous, 29-IV-1923, E. Jahandiez, in E. Jahandiez. Plantes Marocaines, 1923: nº 211 (G 104173). 3 Km Sidi Ifni, 16-IV-1989, F. Jacquemoud (G 104160). Beni Mellal: entre le Fig. 13. Materials of Campanula broussonetiana. Collected by Broussonet in Mogador (Essaouira, Morocco) during his campaign in Morocco in 1805. The type material of this collection is located in Willdenow herbarium (B). 37 Col du Tizi Mlil et Beni Mellal, 5-VI-1980, A. Charpin & al. (G 104180, MA 243114). Entre Oulad M’ Barek y Ouaouizarhte, cerca de Beni Mellal, 12-VI-1982, J. Fernández Casas & al. (MA 633188, MA 430482). Gran Atlas, entre Afourer y Bin-el-Ouidane, 9-VII-1996, S. Cirujano & al. (MA 625034). Subida al Jbel Tassemit, 23-IV-2003, S. Talavera & al. (SEV 216216). Dar el Beïda: Sidi Abd-Er-Rahman, 1886, Ibrahim (G 104250). El Jadida: 20 Km NE Benahmed an der Strase nach El-Khatouat (1419), 6-V1989, D. Podlech (G 104182). Er Ribat: Arbaa-Sehoul, en la carretera hacia Romani, 2-V-2007, S. Talavera & al. (SEV 224279). Fes: subida a Jbel. Zalagh, al NE de Fes, 16-V-2006, S. Talavera & al. (SEV 217539, SEV 217537, SEV 217536, SEV 217535, SEV 217534). Bab Zitouna, collado del Jbel Zalagh, en la ruta de Fes a Ouezzane, 10-V-1994, A. Achhal & al. (SEV 161661). Jbel Zalagh, al NE de Fes, 17-V-2006, S. Talavera & al. (SEV 216499). Kenitra: entre Sidi-Slimane y Khemisset, en saladares, 9-IV-1983, J.A. Devesa & al. (SEV 161658). Forèt de la Mamora, entre Khemisset y Tiflet, a la izquierda en dirección a Máaziz, 25-V-2006, S. Talavera & al. (SEV 234710). Mamora, Dar Salem, sables, 29-IV-1924, E. Jahandiez, in E. Jahandiez, Plantes Marocaines, 1924: nº 211 (G 104174, MA 121503). Montes de Zaïan, entre Tiddas y Jbel, Bouchchene, 24-V-2006, S. Talavera & al. (SEV 234709). Salé, 25IV-1888, Grant (G 104251). Marrakech: cerca de Toufliht, entre Marrakech y Taddert, 14-VI-1982, J. Fernández Casas & al. (MA 430483). De Marrakech a Oukaimeden, Ogaionar, a 53 km de Marrakech, 1-VI-2007, S. Talavera & al. (SEV 223071). Gran Atlas, V-1871, Hooker (LISUG 50012). Krifla, 16/18-IV-1887, Grant (G 104167). Medna Ben Abou, 11-IV-1921, G. Wibagek (G 104175). Tabgourt, 20-VII-1884, Ibrahim (G 104259). Sidi-Ouasmin, 12-VI-1889, Ibrahim (G 104252). Meknés: Atlas Medio, entre Oulmés y Aguelmous, 24-V-2006, S. Talavera & al. (SEV 234711). Entre Meknés et Fes, 2 Km avant le croisement à Ain Taoujdate, 15-V-1989, B. Valdés & al (SEV 161657). Zerhoun, c. 25 Km due NE of Meknès road from Moulay Idriss to Nzaia-desBeni-Ammar, 5-VI-1994, S.L. Jury, M. Ait Lafkih & B. Tahiri (SEV 161660). Oudjda: Berkane, Refuge Zegzel, 23-V-1928, A. Faure (G 104165). Env. de Martimprey-du-Kiss, Pelouses et broussailes, 14V-1930, A. Faure (G 104177, G 104166, MA 121481). Monts des Beni-Snassèn, Djbel Fourhal, près de Taforalt, 23-V-1994, J. Lambinon & G. Van Den Sande (MA 562384). Safi: environs de Mogador, 1867, B. Balansa (G 104253). Falaises da Cap Safi, 17-IV1924, in E. Jahandiez, Pl. Maroc. 1924: nº 82 (MA 121482). Mogador, 1804, Broussonet (G 138271, G 138472, G 138478). Tandja: Chaouia, 25-IV-1935, Gattefossé (G 104163). Taza: c. 32 Km from Taza on minor road near Bab-Bou-Idir, 6-VII-1993, S.L. Jury & al. (SEV 161656). 11 Km from Taza on minor road below RasEl-Ma, 4-VII-1993, M. Ait Lafkih & al. (MA 577072, SEV 161659). Entre Bab-Azhar y Bab-Bou-Idir, 17-VI-2003, S. Talavera & al. (SEV 216476, SEV 216478). Entre Sidi-Abdallah-desRhiata y Bab-Azhar, 17-VI-2003, S. Talavera & al. (SEV 216475). Tittáguen: Armautah, lower part of Jbel Bouhalla, 3-VII-1993, J.A. Mejías & S. Silvestre (SEV 139107). Bou-Ahmed, pista entre Souk-el-Had e Imazerdane, 29-IV-1995, M.A. Mateos & al. (SEV 139102). C. Iaarguit ( Beni Hosmar), 26-V-1930, Font Quer, in Font Quer, iter maroc. 1930: nº 637 (G 104162, MA 121489). Entre Bou-Ahmed y Targha, 3-V-1996, M.A. Mateos & al. (SEV 155222). Kaa Asras, Imarsboutene, 5-V-1996, M.A. Mateos & al. (SEV 155672). Oued-el-Kannar, 7-VI-1930, Font Quer (SEV 139142). Oued Laou, 9-IV-1995, A.J. Caruz & al. (SEV 139100). SE of Chefchaouen, E of Bab Taza, on road from Cherafat to Bab Berred, Matrasse Lakhmasse, 28-V-2002, M. Ait Lafkih & al. (MA 698258). Sok-el-Jemis (Beni Selman), pr. Tiguisas, 7-VI-1930, Font Quer, in Font Quer, Iter Maroc., 1930: nº 638 (G 104164, MA 121484). Talembote, 1 Km vor dem Ort in Flussbett des Oued Talembote, zw. Tamarix u. Oleander, 7-VII-1971, M. Dittrich (G 104178, G 104172). Targha, 7-IV-1995, A.J. Caruz & al. (SEV Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 16/06/2011 11:25 Página 38 38 J. Cano-Maqueda & S. Talavera 139141); ibidem, valle en la ladera W del Djebel Azenti, 8-IV1995, A.J. Caruz & al. (SEV 154716). Tarsif , próximo a OuedLaou, 30-IV-1995, M.A. Mateos & al. (SEV 139101). Tétouan, entre Chechaouen y Talembote, 31-V-1981, (MA 236533). Xauen, 14-V-1921, Font Quer, in Font Quer, iter maroc., 1928: nº 391 (G 104161, MA 121483, MA 121502, MA 121491). B. Sect. Decumbentes Cano-Maqueda & Talavera, Sect. nov. Type (here designated): Campanula decumbens A. DC. Plantae annuae vel perennes. Sinus calycis exappendiculati. Corolla infundibuliformis. Antherae albae. Stylus glaber. Stigma tripartitum erectum vel erectopatens rectum album, in pagina abaxiali multos pilos pollen collectores exhibet, pagina adaxiali glabra in qua pollen germinat. Capsula obpyramidali vel ovata 5angulari, poris tribus versus apicem vel subapicem sitis dehiscente. Annual or perennial plants. Without calycine appendages. Corolla infundibuliform. Anthers white. Style glabrous with a tripartite stigma. Stigma erect or erect-patent, straight, white, with numerous pollen collecting hairs on the abaxial surface, glabrous and receptive in the adaxial side. Capsule obpyramidal or ovoid, dehiscing by three apical or middle position pores. “Habitat in Hispaniâ propè Aranjuez … [description]. Specimina numerosa apud dominum Delessert, in herbario Ventenatii vidi. Circâ Aranjuez in Hispania lecta fuerunt”. Type: Spain. Madrid. Aranjuez, 1827, Delessert s.n. [lectotype, here designated, G 138256! (G-DC), Fig. 14; see observations]. Herb 8(14)-38 cm, annual, decumbent or erect, branched in the upper half or from the base, glabrescent. Stems angled, branched only in the inflorescence, glabrous or with a few antrorse hairs 0.10.3(1.4) mm long, often scabrid near the flowers. Leaves not coriaceous, entire, toothed or crenulate; middle cauline leaves 9(13)-28 × 4.7-10 mm, elliptic or lanceolate, petiolate or sessile, toothed or entire, glabrous or with some hairs 0.1-0.2 mm scattered on the underside nerves, and petioles up to 7-12 mm; upper cauline leaves 3(5)-15 × (0.5)2.5-8 mm, elliptic or lanceolate, cuneate or shortly petiolate, glabrous or glabrescent, with some hairs 0.1-0.3 long on the Observations: This section consists of two species from the Iberian Peninsula (C. decumbens and C. dieckii), and two species from the Eastern Mediterranean (C. ramosissima and C. hawkinsiana). In the topology of the ITS tree (Fig. 1), this section forms a moderately supported clade (67% PPS), where C. decumbens appears as sister of C. dieckii (63% PPS), and C. ramosissima and C. hawkinsiana join into a well-supported subclade (99% BS; 100% PPS). This section thus includes both annual (C. decumbens, C. dieckii, C. ramosissima) and perennial (C. hawkinsiana) taxa, and also shows great variability in chromosome number: with 2n = 32 in C. decumbens (in this work), 2n = 28 in C. dieckii (in this work), 2n = 20 in C. ramosissima (Damboldt & Podlech, 1964) and 2n = 22 in C. hawkinsiana (Contandriopoulos, 1964a). The morphological characters that define this section (glabrous style with three straight, erect or erectpatent stigmas) are very rare in the Campanulaceae, and only co-occur in Campanula sect. Pterophyllum Damboldt. 7. Campanula decumbens A. DC., Monogr. Campan.: 334. 1830 C. patula var. decumbens (A. DC.) Cuatrec. in Trab. Mus. Ci. Nat. Barcelona 12: 441. 1929. Ind. loc.: Fig. 14. Lectotype and isolectotype of Campanula decumbens (G 138256). The lectotype is the plant located in the center, on the right, the only one with a flower in anthesis. The other plants are isolectotypes. Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 39 Taxonomic review of the Campanula lusitanica complex nerves of the underside, and petioles up to 2 mm. Inflorescence laxely paniculate. Flowers pedicellate; pedicel (10.5)40-60(206) mm, glabrous or with some setose hairs near the ovary. Calyx-teeth (6)8-15 × 0.81.6 mm, lanceolate or closely oblanceolate, with the apex obtuse or acute, entire. Corolla 12-21 mm, infundibuliform, with the tube shorter than the lobes; tube 3-8 mm, light blue, with white base; lobes (7)910(14) × 5-8.1 mm, broadly triangular or elliptical, blue, with three darker nerves. Stamens with enlarged base of 0.7-1.5 × 0.5-1.5 mm; filaments c. 1 mm; anthers 1.6-3(3.8) mm, whitish. Ovary glabrous or papillose, rarely hairy; style (3)3.5-4(4.8) mm, glabrous, with 3 stigmas; stigmas (2)2.3-5(5.5) × 0.35-0.4 mm, erect-patent at anthesis, ± straight, flat in the adaxial side, convex and with numerous pollen collecting hairs on the abaxial side. Capsule (2.2)3-8 × 2.4-4(5.7) mm, ovoid or obpyramidal, longer than wide, glabrous or papillose, with 10 thick nerves, dehiscing by three apical or middle position pores. Seeds 0.40.7 × 0.2-0.3 mm, ovoid or ellipsoid, shining, brownyellowish. 2n = 32. Habitat, phenology and distribution: Grassland and wet meadows, on basic substrates (limestone and dolomite); 10-1250 m. VI-VII(VIII). •Endemic to the S of Spain, in the Guadalquivir valley, Sierra de Grazalema and Serranía de Ronda, cited only once from Aranjuez. Spain: Ca M? Ma, Se. Observations: The identity of C. decumbens has been discussed by various authors (Pau, 1896; Cuatrecasas, 1929; Caballero, 1942; Fedorov, 1976; LópezGonzález, 1979-1980); but in fact most authors do not treat with the decumbent plant from Aranjuez described by Alphonse De Candolle. These authors looked for this species near Aranjuez, but they only found an upright and very hairy plant with crenate leaves. This upright plant is the species described by Lange (1893) as C. dieckii and later by Pau (1896) as C. semisphaerica. Only Fedorov (1976) indicated that C. decumbens A. DC. is indeed decumbent. From the type he thought it endemic to Aranjuez, and that it might be better treated as subspecies or variety of C. patula L., as proposed by Cuatrecasas (1929). The type material of Campanula decumbens (G 138256) contains two complete plants of 20 and 25 cm and two stems, all with flower buds and only one plant with an open flower. This flowering plant is chosen as lectotype. The other materials are isolectotypes (see Fig. 14). In the general herbarium at Geneva there is another sheet (G 104141), also from the Ventenat herbarium, containing an incomplete plant of 20 cm with two flowers. The label indicates “Campanula/ affinis vincaeflorae/ Aranjuez près Madrid”. The 39 plant contained in this sheet may also form part of type material. The type material has petiolate middle leaves of the stem, and a small and infundibuliform corolla, in which it resembles C. cabezudoi [described by LópezGonzález (1979-1980) as C. decumbens var. pseudospecularioides]. But the style with three ± straight stigmas is similar to that of C. dieckii and so, it is clear that C. decumbens A. DC. belongs to sect. Decumbentes. Cano-Maqueda & al. (2008) indicated that this species, apparently only known only from the Delessert type materials, could be extinct. However, several populations have been located in Sierra de Grazalema and Serrania de Ronda, with the same characters as the type material of C. decumbens A. DC. Plants from two of these populations were cultivated in the greenhouses of the University of Seville. They retained the decumbent habit, are self-incompatible (Cano-Maqueda & al., unpublished data) and the meiotic number of chromosomes is n = 16. Due to this discovery that C. decumbens occurs on diverse localities on alkaline substrates in the Serranía de Ronda and surrounding areas, it is likely that the material studied by Alphonse de Candolle came from the Southern Betic Cordillera and not from Aranjuez as stated in the sheet of the type. Subsequently, a number of exsiccata by previous collectors in this area and also from the Guadalquivir valley and Cádiz coastland were also identified as C. decumbens. However, the Guadalquivir valley plants are morphologically rather different from those of the Sierras Béticas, although the molecular phylogeny has shown that both groups of plants do not present any changes in the nucleotide sequence (Fig. 1). They also have the same chromosome number (n = 16; Table 1 and Fig. 2H). However, since in addition to the morphological differences, the populations present a different ecology and distribution, we have considered it most appropriate to treat these two groups of populations as two subspecies. KEY TO THE SUBSPECIES 1. Middle cauline leaves petiolate; capsule (2.2)3-4(4.4) × 2,4-4 mm, ovoid ............... a. C. decumbens subsp. decumbens 1. Middle cauline leaves sessile; capsule 4-8 × 3-5.7 mm, obpyramidal ................... b. C. decumbens subsp. baetica a. Campanula decumbens A. DC. subsp. decumbens Illustrations: De Candolle (1830, tab. 12 fig. A); Fig. 15 A, B. Herb 20-36 cm, decumbent. Middle cauline leaves petiolate. Calyx-teeth (6)8-10 × 0.8-1.1 mm, lanceolate, with the apex obtuse. Corolla 12-14 mm; tube 3- Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 40 40 J. Cano-Maqueda & S. Talavera Fig. 15. Flowers and fruits of Campanula species. A, B, C. decumbens subsp. decumbens (Benaoján, Málaga, Spain, SEV 218875); C, D, C. decumbens subsp. baetica (Villamartín, Cádiz, Spain, SEV 256653); E, F, C. dieckii (Alfarnate, Málaga, Spain, SEV 256652). The scale bar = 3 mm. Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 41 Taxonomic review of the Campanula lusitanica complex 4 mm; lobes 9-10 × 5-6 mm, broadly triangular. Stigmas 2-3.2 mm. Capsule (2.2)3-4(4.4) × 2.4-4 mm, ovoid. 2n =32. Habitat, phenology and distibution: Wet meadows, on basic substrates (limestone and dolomite); (480) 1000-1250 m. VI-VII(VIII). • Endemic to the Sierra de Grazalema and Serranía de Ronda (Fig. 16). Spain: Ca M? Ma. Selected specimens SPAIN. Cádiz: Benaocaz, Manga de Villaluenga, 22-VI-1984, A. Aparicio & S. Silvestre (SEV 161834). Grazalema, 11-VI-1890, E. Reverchon, in E. Reverchon, Plantes de l ‘Andalousie, 1889: nº 17 (G 104066). Sierra del Endrinal, 10-VII-1925, Font i Quer & E. Gros (G 104158, MA 702573, MGC 53045); ibidem, 29-VI-1849, P. Fonti i Quer & E. Gros (SALA 114817). Málaga: Benaoján, Carretera hacia Ronda, 15-V-1988, D. Montilla (MGC 40574); ibidem, Sierra del Palo, 17-VI-2007, J. Cano-Maqueda (SEV 218875); ibidem, 6-VI-2001, M. Becerra & al. (MGC 52844, MGC 51636); ibidem, Puerto España, 25-V-2002, M. Becerra & F. Sánchez (MGC 51972). Ronda, 18-VI-1889, E. Reverchon, in E. Reverchon, Plantes de l’Andalousie, 1889 nº 17 (G 104072). b. Campanula decumbens subsp. baetica CanoMaqueda & Talavera, subsp. nov. 41 A Campanula decumbens subsp. decumbens foliis caulinis sessilibus non petiolatibus, caule erecto non decumbente, capsula obpyramidali non ovata differt. A Campanula dieckii foliis glabrescentibus integris vel serratis, calyce, corolla stigmatibusque majoribus differt. This subspecies differs from Campanula decumbens subsp. decumbens in its non-petiolate, sessile cauline leaves, erect rather than decumbent stems, and obpyramidal capsule, not ovoid. It differs of Campanula dieckii by its entire or toothed and glabrescent leaves, and by the larger size of the calyx, corolla and stigmas. Type: Spain. Sevilla. El Coronil, bujeos, June 1990, Aparicio & Silvestre s.n. (holotype: SEV 217838!, Fig. 17; isotype: SEV 238167!). Illustrations: Boissier (1839, tab. 120a, as C. erinoides); Fig. 15 C and D. Herb (8)14-38 cm, erect. Middle cauline leaves sessile. Calyx-teeth (8)8.5-15 × 1-1.6 mm, closely oblanceolate, with the apex acute. Corolla (12)14.5- Fig. 16. Distribution map of Campanula decumbens subsp. decumbens (䡬), C. decumbens subsp. baetica (夡) and C. dieckii (䊱). Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 42 42 J. Cano-Maqueda & S. Talavera decabras i Provinsen Cuenca, 1.juni 1892 blomstrencle (Dieck)!”.Type: Spain. Cuenca, Uña, 1 June 1892, Dieck s.n. (lectotype, here designated, C, herb. J. Lange, Fig. 18; see observations). C. matritensis var. nevadensis A. DC. in DC., Prodr. 7(1): 481. 1838. Ind. loc.: “in editioribus SierraeNevadae. (Boiss.!)”. Type: Spain. Granada, Sierra Nevada, 1838, Boissier s.n. (lectotype, here designated, G138396!; see observations). C. semisphaerica Pau, Not. Bot. Fl. Españ. fasc. 6: 76. 1896. Ind. loc.: “Sacañet, á 1100 m de alt. en compañía de la C. dichotoma; 8 julio 1895”. Type: Spain. Castellón, Sacañet, 8 July 1895, Pau s.n. (lectotype, here designated, MA121465!; see observations). C. argutidens Porta et Rigo, Iter. Hisp. III: n.º 309 (1891), in sched., nom. nud. C. specularioides var. argutidens Porta et Rigo, Iter. Hisp. III: nº 309 (1891), in sched., nom. nud. Illustrations: Sáez & Aldasoro (2001: 132, fig. 40, as C. decumbens); Fig. 15 E, F. Fig. 17. Holotype of Campanula decumbens subsp. baetica Cano-Maqueda & Talavera (SEV 217838). 21 mm; tube (3.4)4-8 mm; lobes 7-14 × 5-8 mm, elliptical. Stigmas (4)4.3-5.5 mm. Capsule 4-8 × 3-5.7 mm, obpiramidal. n = 16. Habitat, phenology and distribution: Grasslands on basic substrates, usually very clay soils; 10-200 m; (V)VI-VIII. • Endemic to the S of Spain, in the Guadalquivir valley (Fig. 16). Spain: Ca Se. Selected specimens SPAIN. Cádiz: Cádiz, without date, Chaubert (G 104126). Cerca de Villamartín, 31-V-1969, E.F.Galiano & al. (SEV 161833); ibidem, en la carretera a El Bosque, 17-VI-2009, S. Talavera & al. (SEV 248711). Vejer de la Frontera, Caños de Meca, 15-V-1959, D.M.C. Brinton-Lee (SEV 81177). Villamartín, 8-V-2010, J. CanoMaqueda (SEV 256653). Sevilla: Carmona, J.M. Triguero (SEVhistórico 1077). 8. Campanula dieckii Lange in Overs. Kongel. Danske Vidensk. Selsk. Forh. Medlemmers Arbeider 1893: 195. 1893 Ind. loc.: “Ciudad Encantada mellem Uña og Val- Herb (3)8-20(30) cm, annual, erect, usually branched in upper half, often densely pubescent, at least in the lower half. Stem angled, little branched, densely pubescent, with ± antrorse setose hairs, 0.10.7 mm, sometimes glabrescent at the apex. Leaves somewhat fleshy, crenate, lobed or pedate, sometimes the uppermost subentire; middle cauline leaves (4)718 × 3-9(14) mm, ± elliptical, sessile, subauriculate, densely pubescent, with hairs 0.1-0.7 mm; upper cauline leaves 3-13.5 × 0.5-2.5(7.7) mm, ± lanceolate, sessile, subauriculate, glabrous or more often with scattered hairs 0.1-0.6 mm. Inflorescence paniculate, lax. Flowers pedicellate; pedicel (11.9)19-90(155) mm, glabrous or papillose on top. Calyx-teeth (2.3) 3-10.5 × 0.7-1.5 mm, lanceolate, obtuse, with thickened margins. Corolla (6.6)8-13.7 mm, infundibuliform, with the tube much shorter than the lobes; tube (1.5)3-4.5 mm, light blue with white base; lobes 712.3 × (3)4-5.5(7.6) mm, ovate-lanceolate, blue, with three purple nerves. Stamens with enlarged base of 1.1-1.9 × 1.5-1.9 mm; filaments 0.6-0.7 mm; anthers 2.1-3(3.8) mm, whitish. Ovary glabrous, papillose or densely hairy, with setose hairs of (0.2)0.7-2.2 mm; style 2.1-4(4.7) mm, glabrous, with three stigmas; stigmas (2.1)2.5-4 mm, erect-patent at anthesis, ± straight. Capsule (3.5)4.9-9(11.5) × 3.5-6.5 mm, obpyramidal, longer than wide, from papillose to densely hairy, with setose hairs of 0.3-1 mm, with 10 very wide nerves like flat ribs, dehiscing by three middle position pores. Seeds 0.5-0.8 × 0.2-0.3 mm, ovoid, shining, yellowish to brown. 2n = 28. Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 43 Taxonomic review of the Campanula lusitanica complex Habitat, phenology and distribution: Wet meadows and kermes oak woodlands on basic substrates (gypsum, limestone and dolomite); 600-2300 m. VIVII(VIII). • Endemic to the Iberian Peninsula, C, E and SE Spain, where it is frequent, and rare in CW Portugal, Serra do Sintra (Fig. 16). Portugal: E. Spain: Ab Al Bu Cs CR Cu Gr Gu J Lo M Ma Mu Sa Sg So To Va Z Za. Observations: This species has been confused with C. decumbens by several authors. Cuatrecasas (1929) commented: “Las diferencias que separan a la planta de De Candolle [C. decumbens] de la de Loefling [C. lusitanica] son mínimas. La primera son simplemente formas de tallos menos ramificados y lacinias calicinales más anchas que en la última, la cual, en general, también difiere por su mayor estrechez de la corola. Estos caracteres están, sin embargo, sujetos a variaciones, lo mismo que la vestidura del tubo de los cálices entre unos muy híspidos y otros muy lampiños”. However, in this comment Cuatrecasas is referring to three different species from the Sierra de Má- 43 gina (Jaen): C. matritensis with a glabrous ovary, and C. cabezudoi and C. dieckii, both with very hairy, almost hispid ovary. We have seen material of this species from two collections from Sintra (Estremadura, Portugal). One collected by F. Fernandes in V-1914 (G 104249, 104150; LISU 36374; MA 121500), distributed in exiccata by F. Sennen (in F. Sennen Pl. Esp. n.º 6006), and another by W. Rothmaler on 13-V-1938 (G 104148; LISE 4368). Since this plant has not been collected again at this locality, the presence of this species as a native in Portugal may be considered doubtful. However, this kind of disjunction is not uncommon in other species of the Iberian Peninsula, i.e., Silene distichia Willd., a common species in the Eastern half of Spain and rare in the W Portugal (Talavera, 1990). The type material of Campanula dieckii is composed by 7 whole plants at anthesis of 10-20 cm with sessile, elliptic, crenate cauline leaves, infundibuliform corolla, glabrous style, straight stigma and densely setose ovary. The sheet also contains two flowers in an envelope. The first plant, in the upper left corner of the sheet is chosen as lectotype, because it is the one that best fits the description of the author. The other six plants are isolectotypes (see Fig. 18). In the Geneva herbarium there is a sheet (G104244) with three plants that are also isolectotypes. The sheet of type material of C. matritensis var. nevadensis contains 6 plants in flower. We have chosen as lectotype the plant placed at the top right, about 10 cm, very hairy, with crenate leaves and two flowers in anthesis. The remaining plants are isolectotypes. The type material of C. semisphaerica consists of two very small plants, each with one flower open. The plant on the right is the lectotype. On the sheet there is a typed label by Ginés López-González dated 1311-1979 indicating the material contained on the sheet as holotype. Pau’s indication “genuine” may be understood that this material was chosen as type by the author of the binomial. Selected specimens Fig. 18. Lectotype (in the upper left corner) and isolectotypes (the rest of plants) of Campanula dieckii (C, herb. Lange). PORTUGAL. Estremadura: Sintra, V-1914, F. Fernandes, in F. Sennen, Plantes D’Espagne, 1926, nº 6006 (G 104150, G 104249, LISU 36374, MA 121500, MA 474598); ibidem, 13-V-1938, W. Rothmaler, in W. Rothmaler, Flora Lusitanica nº 13133 (G 104148, LISE 4368). SPAIN. Albacete: Alcaraz, La Molata, 24-V-1993, B. Casaseca & M.A. Carrasco (MA 531238). Alrededores de Santa Elena de Ruidera, 24-V-1933, González Albo (MA 121567). El Ballestero, 28-VI-1935, González Albo (MA 432730). El Cascajal, 24-V-1933, González Albo (MA 121480, MA 121479). Pontegruelos, 28-VI1935, González Albo (MA 432731). Tús, vertiente SE del Calar del Mundo, 29-V-1987, E. Villanueva & al. (MA 393151). Villaverde Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 44 44 J. Cano-Maqueda & S. Talavera de Guadalimar, 18-VI-1969, P.E. Gibbs (SEV 161752). Almería: Bacares, 4-VI-1929, E. Gros (MA 433604). De Bacares a la Venta de Lleiva, 5-VI-1929, E. Gros (MA 433605). Burgos: Ciruelos de Cervera. Pie del Alto de la Cabeza, 11-VII-1979, Pons Sorolla & Susanna (MA 413025). Hortezuelos, 17-VI-1982, M.A. Carrasco & M. Velayos (MA 312710, SALA 32983). Huidobro, 5-VII-1987, Galán Cela & A. Martín (MA 639955). Tejada. Pico Valdosa, 3VII-1979, J. Fernández Casas & al. (MA 413017). Ciudad Real: La Molata, 2-VI-1934, González Albo (MA 201345). Cuenca: Barajas de Melo, valle del río Calvache, pr. Urbanización Valderíos, 580 m, 30-V-1998, V.J. Arán & M.J. Tohá (MA 614852. MA 620787). Pinar de Beteta, 8-VII-1932, A. Caballero (MA 121464). Talayuelas, 1100m, 18-VI-1979, G. Mateo (MA 256531). Granada: Alhama de Granada, Barranco del Malinfierno, 17-VI-2004, B. Cabezudo & al. (MGC 59291). Arenas del Rey, El Cenacho, 3-VI2004, B. Cabezudo & al. (MGC 59289). Ascenso a Sierra Nevada, falla del Purche, 8-V-1966, S. Silvestre (SEV 19702). Guadix, 11VI-1921, E. Gros (MA 31778). Sierra de Albuñuelas, 15-VI-1976, A. Asensi & P. Díez (MGC 3617). Sierra de Baza, ascenso a Santa Bárbara, 21-VI-1988, S. Talavera & al. (G 104091). Sierra de Castril, VI-1903, E. Reverchon., in E. Reverchon, Plantes d’Espagne 1903: nº 1209 (G 104074, MA 121523). Sierra de Cázulas, 23-VI1976, Ladero & al. (MA 204705, SALA 8787). Sierra de Guillimona, Cuerda de los Mirabetes, 23-VI-1988, S. Talavera & al. (G 104059). Huéscar, Sierra de la Sagra, cara sur, 14-VI-1995, B. Cabezudo & al. (MGC 39893). Padul, 4-VI-1980, M. Ladero & al. (SALA 92232). Sierra del Pinar, without date, Reverchon (G 104076). Sierra Nevada, Trevenque, VI-1973, J. Fernández-Casas (G 104078, SEV 19866). Guadalajara: Codes, 21-VI-1988, M.A. Carrasco & M. Velayos (MA 711892). Sacecorbo, 9-VI-1973, A. Segura Zubizarreta (G 104064, MA 269833). Tamajón, alrededores de la ermita de los Enebrales, 4-VII-1978, M.A. Rivas & C. Soriano (MA 385718). Torremocha del Pinar, 19-VI-1995, M.A. Martín Ballesteros (SALA 59846). Jaén: between Tobos and Vites, bed of river Zumeta, 25-VI-1988, S. Talavera & al. (G 104058). Cambil, Sierra de Mágina, Matabegí, 15-VI-1995, B. Cabezudo & al. (MGC 39707). Cazorla, aledaños de la C. F. Fuente del Oso, 31-V-1976, F. Muñoz Garmendia & C. Soriano (MA 454856). La Iruela, barranco de Guadahornillos, 16-VI-1976, F. Muñoz Garmendia & C. Soriano (MA 454825). Las Altarillas, 16-VI-1941, E. Guinea (MA 432654). Orcera, 6-VI-1980, C. Soriano (MA 592112). Pozo Alcón, pico de Cabañas, 20-VI-1975, F. Muñoz Garmendia & C. Soriano (MA 454823). Santiago de la Espada, cabecera del arroyo del Membrillo, 5-VI-1975, F. Muñoz Garmendia & C. Soriano (MA 454816). Segura de la Sierra, 17-VI-1850, E. Bourgeau, in E. Bourgeau, Pl. d’ Espagne, 1850: nº 992 (G 104100). Sierra del Pozo, VI-1905, E. Reverchon, in E. Reverchon, Plantes d’ Espagne 1905: nº 1209 (G 104134). Sierra Mágina, entre el Cortijo de los Prados y Cerro Carceles, 11-VI-1987, E. Villanueva & al. (MA 401189). Siles, VI-1850, M. Blanco (G 104113). Madrid: Algodor, 31-V-1925, A. Caballero & González Guerrero (MA 432742, MA 432744). Aranjuez, V-1897, C. Pau (MA 121457). Idem, 25-V1919, C. Vicioso (MA 121459). Arganda, IV-1932, C. Pau (MA 432687). Piul de Rivas, V-1915, C. Vicioso (MA 121456). Venta del Gorro, 1802, Lagasca (G 138462). Málaga: Alcaucín. Sierra Tejeda, loma de las Víboras, 7-VI-2002, D. Navas & al. (MGC 52239). Alfarnate, 30-III-2007, (SEV256652). Canillas de Albaida, Los Horcajos, puerto de la Orza, 5-VI-1919, E. Gros (MA 121468). Castillo de Frigiliana, VI-1919, E. Gros (MA 121467). Cómpeta, Navachica, 22-VI-2004, B. Cabezudo & al. (MGC 59293). Sedella, 14-VI-1994, A. Aparicio & al. (MA 543877). Sierra Tejeda, V1914, E. Gros (MA 121471). Murcia: Sierra de Moratalla, Revolcadores, 15-VII-1974, A. Charpin & J. Fernández. Casas (G 104079). Salamanca: Ledesma, 19-V-1976, J. Sánchez (MA 219750). Molinillo, 1859, Sainz (MA 153143). Segovia: Cedillo de la Torre, 5-VI-1985, A.R. Burgaz & A. Izuzquiza (MA 306519). Fuentidueña, 24-VI-1983, T. Romero (SALA 41198). Lastras de Cuéllar, Molino Ladrón, 840 m, 24-V-1998, P. Bariego Hernández & A. Gastón González (MA 754470). Villaseca, 12-VI-1983, T. Romero (SALA 41951). Soria: Andaluz, 26-VI-1975, A. Segura Zubizarreta (FCO 7873, SEV 69199). Cañón del río Lobos, 17-VI1982, A. Buades (MA 502178). Herrera de Soria, 30-VI-1983, A. Buades (MA 504408). Ucero, cuesta de la Galiana, 28-V-1983, A. Buades (MA 504043). Toledo: Toledo, 22-V-1897, C. Pau (MA 121458). Valladolid: Castronuño, 20-VI-1984, F.J. González & C. J. Valle (SALAF 7361). Encinas de Esgueva, 7-VII-1983, J.L. Fernández Alonso (MA 517691). Rábano, 15-VI-1983, T. Romero (SALA 41197). Urueña, 20-VI-1980, Fernández Díez (SALA 22002). Valladolid, 17-VI-1906, Sennen (MA 121463). Zamora: Castrillo de la Guareña, 30-V-1983, X. Giráldez (SALA 31601). Zaragoza: Calatayud, 15-VI-1910, C. Vicioso (G 104095, MA 121462). Cerros de Andrés, VI-1898, C. Vicioso (MA 121460). Acknowledgements We thank Drs. B. Cabezudo (Málaga, Spain), the late S. Castroviejo (Madrid, Spain) and E. Rico (Salamanca, Spain) for their help in field collections. We are grateful to curators of the cited herbaria for the loan of specimens. We are also grateful to Dr. Peter Gibbs (St. Andrews, UK) for reviewing the manuscript. This work has been financed by grants from the Ministerio de Educación y Ciencia to M. Arista (REM2002-04354-C02-02 and CGL2005-01951), to S. Talavera (CGL2006-00817 and CGL2009-08178) and by a predoctoral grant to J. Cano-Maqueda (BES-2003-0332). References Bentham, G. & Hooker, J.D. 1876. Genera Plantarum, vol. 2. London. Boissier, E. 1839. Voyage botanique dans le midi de L’Espagne pendant l’année 1837, vol. 2: 1-480. Paris. Boissier, E. 1875. Flora Orientalis, vol. 3: 893-945. Genève, Basel. Borsch, T., Korotkova, N., Raus, T., Lobin, W. & Löhne, C. 2009. The petD group II intron as a species level marker: utility for tree inference and species identification in the diverse genus Campanula (Campanulaceae). Willdenowia 39: 7-33. Brotero, F.A. 1800. Phytographia Lusitaniae Selectior, fasc. I. Olissipone. Brotero, F.A. 1804. Flora Lusitanica, vol. 1. Olissipone. Brotero, F.A. 1816. Phytographia Lusitaniae Selectior, vol. 1. Olissipone. Caballero, A. 1942. Apuntes para una flórula de la Serranía de Cuenca. Anales del Jardín Botánico de Madrid 2: 236-265. Cano-Maqueda, J., Talavera, S., Arista, M. & Catalán, P. 2008. Speciation and biogeographical history of the Campanula lusitanica complex (Campanulaceae) in the Western Mediterranean region. Taxon 57: 1252-1266. Cellinese, N., Smith, S.A., Edwards, E.J., Kim, S.T., Haberle, R.C., Avramakis, M. & Donoghue, M.J. 2009. Historical biogeography of the endemic Campanulaceae of Crete. Journal of Biogeography 36: 1253-1269. Contandriopoulos, J. 1964a. Recherches sur la flore endémique de la Corse et sur ses origines II. Revue Générale Botanique (Paris) 71: 361-384. Contandriopoulos, J. 1964b. Contribution a l’etude caryologique des Campanulaceas de Grece. Bulletin de la Société Botanique de France 111: 222-235. Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 45 Taxonomic review of the Campanula lusitanica complex Contandriopoulos, J. 1966. Contribution a l’etude caryologique des Campanulaceas de Grece II. Bulletin de la Société Botanique de France 113: 453-474. Cuatrecasas, J. 1929. Estudios sobre la flora y la vegetación del Macizo de Mágina. Trabajos del Museo de Ciencias Naturales de Barcelona, Ser. Bot. 12: 7-510. Damboldt, J. 1965. Campanula tommasiana Koch und C. waldsteiniana R. et S.: Zur Zytotaxonomie zweier mediterraner Reliktsippen. Österreichische Botanische Zeitschrift 112: 392-406. Damboldt, J. 1976. Materials for a Flora of Turkey XXXII: Campanulaceae. Notes from the Royal Botanical Garden, Edinburgh, 35: 39-52. Damboldt, J. 1978. Campanula. In: Davis, P.H. (ed.), Flora of Turkey and East Aegean Islands, vol. 6: 2-64.University Press, Edinburgh. DeBry, R.W. & R.G. Olmstead. 2000. A simulation study of reduced tree-search effort in bootstrap resampling analysis. Systematic Biology 49: 171-179. De Candolle, A. 1830. Monographie des Campanulées. Paris. De Candolle, A. 1838. Campanulaceae. In: De Candolle, A.P. (ed.), Prodromus Systematis Naturalis Regni Vegetabilis, vol. 7: 414-496. Paris. Eddie, W.M.M., Shulkina, T., Gaskin, J., Haberle, R.C. & Cansen, R.K. 2003. Phylogeny of Campanulaceae s. str. inferred from its sequences of nuclear ribosomal DNA. Annals of the Missouri Botanic Garden 90: 554-575. Endlicher, S. 1838. Genera Plantarum. Vindobonae. Fedorov, A.A. 1957. Campanulaceae. In: Shiskin, B.K. (ed.). Flora USSR, vol. 24: 126-450. Moskva-Leningrad. Fedorov, A.A. 1976. Campanula. In: Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M. & Webb, D.A. (eds.). Flora Europaea, vol. 4: 74-93. Cambridge University Press, Cambridge. Felsenstein, J. 1985. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39: 783-791. Fernandes, A. 1962. Sobre a cariologia de Campanula lusitanica L. ex Loefl. e C. transtagana R. Fernandes. Boletim da Sociedade Broterian, sér. 2, 34: 129-137. Fernández, M.P., Narbona, F.E., Ortiz, M.A., Talavera, S. & Salgueiro, F.J. 2001. Números cromosomáticos de plantas occidentales. Anales Jardín Botánico de Madrid 58: 341-345. Gallego, M.J. 1986. Números cromosómicos para la flora española, nº 464-470. Lagascalia 14: 286-288. Gallego, M.J. 1987. Campanula. In: Valdés, B., Talavera, S. & Galiano, E.F. (eds.). Flora de Andalucía Occidental, vol. 2: 564265. Ketrès, Barcelona. García Martín, F. & Silvestre, S. 1985. Números cromosómicos para la flora española nº 409-421. Lagascalia 13: 313-318. Goldblatt, P. 1985. Index to plant chromosome numbers 19821983. Monographs in Systematic Botany from the Missouri Botanical Garden 13: 1-224. Goldblatt, P. & Johnson, D.E. 1990. Index to plant chromosome numbers 1986-1987. Monographs in Systematic Botany from the Missouri Botanical Garden 30: i-ix, 1-124. Goldblatt, P. & Johnson, D.E.. 1994. Index to plant chromosome numbers 1990-1991. Monographs in Systematic Botany from the Missouri Botanical Garden 51: i-ix, 1-267. Haberle, R.C., Dang, A., Lee, T., Penaflor, C., Cortes-Burn, H., Oestreich, A., Raubenson, L., Cellinese, N., Edwards, E.J. & Kim, S.T. 2009. Taxonomic and biogeographic implications of a phylogenetic analysis of the Campanulaceae based on three chloroplast genes. Taxon 58: 715-734. 45 Hoffmannsegg, J.C.C. & Link, H.F. 1820-1824. Flore Portugaise, vol. 2 (17-18): 14 Berlin. Hoffmannsegg, J.C.C. & Link, H.F. 1820. Flore Portugaise, vol. 2: pl. 82. Berlin. Huelsenbeck, J.P. & Ronquist, F. 2002. MRBAY ES. Bayesian inference of phylogeny. V. 3.0. Bioinformatics 17: 754-755. Lammers, T.G. 2007a. Campanulaceae. In: Kadereit, J.W. & Jeffrey, C. (eds.), The families and genera of vascular plants. Flowering Plants. Monocots. Asteridae, vol. 8: 26-56. Springer-Verlag, Berlin. Lammers, T.G. 2007b. World Checklist and bibliography of Campanulaceae. Royal Botanic Gardens, Kew. Lange, J. 1893. Nye Bidrag til Spaniens Flora Overs-Kongel. Danske Videnskabernes Selskabs Forhandlinger og dets Medlemmers Arbeider 1893: 191-207. Larsen, K. 1954. Chromosome numbers of some European flowering plants. Botanisk Tidsskrift 50: 163-174. Larsen, K. 1956. Chromosome studies in some mediterranean and south European flowering plant. Botaniska Notiser (Lund) 109: 293-307. Leaché, A.D. & Reeder, T.N. 2002. Molecular systematics of the Eastern fence lizard (Sceloporus undulatus): a comparision of parsimony, likelihood, and bayesian approaches. Systematic Biology 51: 44-68. Levan, A., Fredga, K. & Sandberg, A. 1964. Nomenclature for centromeric position on chromosomes. Hereditas 52: 201-220. López-González, G. 1979-1980. Dos campánulas españolas: C. decumbens DC. y «C. mollis L.». Boletim da Sociedade Broteriana, sér. 2, 53: 299-308. Merxmüller, H. & Damboldt, J. 1962. Die chromosomenzahlen einiger rupicoler mediterraner Campanulae. Berichte der Deutschen Botanischen Gesellscaft 75: 233-236. Park J.M., Kovačič, S., Liber, Z., Eddie, W.M.M. & Schneeweiss, G.M. 2006. Phylogeny and Biogeography of Iophyllous Species of Campanula (Campanulaceae) in the Mediterranean area. Systematic Botany 31: 862-880. Pau, C. 1896. Notas botánicas a la flora española, fasc. 6. Segorbe. Pau, C. 1921. Notas sueltas sobre la flora matritense. Boletín de la Sociedad Ibérica de Ciencias Naturales 20: 176-191. Pau, C. 1924. Plantas del norte de Yebala (Marruecos). Memorias de la Real Sociedad Española de Historia Natural 12: 263-401. Podlech, D. & Damboldt, J. 1964. Zytotaxonomische Beiträge zur Kenntnis der Campanulaceen in Europa. Berichte der Deutschen Botanischen Gesellschaft 7(9): 360-369. Rambaud, A. 1996. Sequence Alignment Editor (Se-Al). Ver. 1.0 alpha 1. Oxford University, Oxford. Roquet, C., Sáez, L., Aldasoro, J.J., Susanna, A., Alarcón, M.L. & García-Jacas, N. 2008. Natural delineation, molecular phylogeny and floral evolution in Campanula. Systematic Botanic 33: 203-217. Roquet, C., Sanmartín, I., García-Jacas, N., Sáez, L., Susanna, A., Wikström, N. & Aldasoro, J.J. 2009. Reconstructing the history of Campanulaceae with a Bayesian approach to molecular dating and dispersal-vicariance analyses. Molecular phylogenetics and Evolution 52: 575-587. Rouy, G. 1908. Flore de France 10: 59-82. Paris. Sáez, L. & Aldasoro, J.J. 2001. Campanula. In: Paiva, J., Sales, F., Hedge, I., Aedo, C., Aldasoro, J.J., Castroviejo, S., Herrero, A. & Velayos, M. (eds.), Flora iberica, vol. 14: 105-136. CSIC, Madrid. Sáez, L. & Aldasoro, J.J. 2003. A taxonomic revision of Campanula L. subgenus Sicyocodon (Feer) Damboldt and subgenus Megalocalyx Damboldt (Campanulaceae). Botanical Journal of the Linnean Society 141: 215-241. Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 46 46 J. Cano-Maqueda & S. Talavera Schultes, J.A. & Roemer, J.J. 1819-1820. In: Roemer, J.J. & Schultes, J.A. (eds.), Systema Vegetabilum, vol. 5: 104. Stuttgart. Snow, R. 1963. Alcoholic hydrochloric acid-carmine as a stain for chromosomes in squach preparations. Biotechnic & Histochemistry 38: 9-13. Stebbins, G.L. 1938. Cytological characteristics associated with the different growth habits in the dicotyledons. American Journal of Botany 25: 189-198. Stefanović, S. & Lakušić, D. 2009. Molecular reappraisal confirms that the Campanula trichocalycina-pichleri complex belongs to Asyneuma (Campanulaceae). Botanica Serbica 33: 21-31. Susanna, A., García-Jacas, N., Hidalgo, O., Vilatersana, R. & Garnatje, T. 2006. The Cardueae (Compositae) revisited: insights from ITS, trnL-trnF, and matK nuclear and chloroplast DNA analysis. Annals of the Missouri Botanical Garden 93: 150-171. Swofford, D. L. 2002. PAUP*: Phylogenetic Analysis Using Parsimony and other methods. Ver. 4.0 beta 10. Sinauer, Sunderland. Talavera, S. 1990. Silene. In: Castroviejo, S. & al. (eds.), Flora iberica 2: 313-406, C.S.I.C., Madrid. Torrecilla, P. & Catalán, P. 2002. Phylogeny of broad-leaved and fine-leaved Festuca linajes (Poaceae) based on nuclear ITS sequences. Systematic Biology 27: 241-251. Valdés, B., Talavera, S. & Galiano, E.F. (eds.). 1987. Flora Vascular de Andalucía Occidental, vol. 2: 564. Editorial Ketrès, Barcelona. White, T.J., Bruns, T., Lee, S. & Taylor, J.W. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis, M.A., Gelfand, D.H., Sninsky, J.J. & White, T.J. (eds.), PCR protocols; a guide to methods and applications, Academic Press, San Diego, California. pp.: 315-322. Willkomm, H.M. 1868. Campanulaceae. In: Willkomm, H.M. & Lange, J. (eds.), Prodromus Flora Hispanicea, vol. 2: 279-298. Stuttgartiae. Appendix 1 Species, origin of plant material, voucher, collector or reference and ITS GenBank accession no. Adenophora divaricata Franch. & Sav.; cultivated in Royal Botanic Gardens Edinburgh; Eddie & al. (2003); AY 322005 & AY 331418. Asyneuma japonicum (Miq.) Briq.; Korea; Kim & al. (1999); AF 183437 & AF 183443. Azorina vidalii (Wats.) Feer.; cultivated in University of Edinburgh; Eddie & al. (2003); AY 322007 & AY 331420. Campanula alliarifolia Willd.; cultivated in Royal Botanic, Garden, Kew; Eddie & al. (2003); AY 322008 & AY 331421. C. alpina Jacq.; Austria, Niedere Tauern; Park & al. (2006), DQ 304573. C. armazica Kharadze; Caucasus; Eddie & al. (2003); AY 322009 & AY 331422. C. arvatica Lag.; cultivated in University of Edinburgh; Eddie & al. (2003); AY 322010 & AY 331423. C. barbata L.; Italy; Eddie & al. (2003); AY 32211 & AY 331424. C. beckiana Hayek; Austria, Northeastern Alps; Park & al. (2006); DQ 304619. C. bellidifolia Adams; Caucasus; Eddie & al. (2003); AY 322012 & AY 331425. C. bononiensis L.; Austria, Leithagebirge; Park & al. (2006); DQ 304571. C. broussonetiana Schult.; Morocco, Tazzeka (Middle Atlas); Cano-Maqueda & al. (2008); FM 212711. C. cabezudoi Cano-Maqueda & Talavera; Spain, Málaga, Junquera; Cano-Maqueda & al. (2008); FM 212727. C. carpatica Jacq.; USA, Illinois; Eddie & al. (2003); AY 322013 & AY 331426. C. cenisia L.; Austria, Lechtaler Alps; Park & al. (2006); DQ 304622. C. cespitosa Scop.; Austria, Northeastern Alps; Park & al. (2006); DQ 304621. C. decumbens A. DC. subsp. decumbens; (1): Spain, Málaga: Benaoján; Cano-Maqueda & al. (2008); FM 212735. C. decumbens subsp. baetica Cano-Maqueda & Talavera; (2): Spain, Cádiz, Villamartín; Cano-Maqueda; HQ 407547. C. dichotoma L.; Italy, Calabria, NW of Nicótera; Park & al. (2006); DQ 304579. C. dieckii Lange; (1) Spain, Granada, Alhama de Granada; Cano-Maqueda & al. (2008); FM 212733. C. dieckii Lange; (2) Spain, Cuenca, Barajas de Melo; Roquet & al. (2008) as C. decumbens; EF 090526 & EF 090567.C. divaricata Michx.; USA, Virginia; Eddie & al. (2003); AY 322014 & AY 331427. C. drabifolia Sibth. & Sm.; Greece, Ionian islands, Atokos; Park & al. (2006); DQ 304578. C. edulis Forssk.; Saudi Arabia; Eddie & al. (2003); AY 233015 & AY 331428. C. elatines L.; Italy, Alpi Cozie; Park & al. (2006); DQ 304624. C. elatinoides Moretti; Italy, Southern Alps; Park & al. (2006); DQ 304625. C. erinus L.; Spain, Sevilla, Sevilla City; Cano-Maqueda & al., (2008); FM 212737. C. fenestrellata Feer subsp. fenestrellata; Croatia, Velebit, Velika Paklenica; Park & al. (2006); DQ 304592. C. fragilis subsp. cavolinni (Ten.) Damb.; Italy, Abruzzo; Park & al. (2006); DQ 304629. C. garganica Ten. subsp. garganica; cultivated in Botanical Garden Zagreb (material from Italy); Park & al. (2006); DQ 304596. C. glomerata L.; cultivated in University of Edinburgh; Eddie & al. (2003); AY 322017 & AY 331430. C. grossheimii Kharadze; Caucasus; Eddie & al. (2003); AY 322018 & AY 331431. C. hawkinsiana Hausskn. & Heldreich; Eddie & al. (2003); AY 322019 & AY 331432. C. hercegovina Degen & Fiala; Bosnia & Herzergovina, Blidinje; Park & al. (2006); DQ 304616. C. herminii Hoffmans. & Link.; Portugal; Eddie & al. (2003); AY 322020 & AY 331433. C. isophylla Moertti; cultivated in Botanical Garden Zagreb (material from Italy); Park & al. (2006); DQ 304630. C. justiniana Witasek; Croatia, Čabranka river; Park & al. (2006); DQ 304613. C. kolenatiana C.A. Mey.; Caucasus; Eddie & al. (2003); AY 322022 & AY 331435. C. lanata Friv.; cultivated in University of Edinburgh; Eddie & al. (2003); AY 322023 & AY 331436. C. latifolia L.; Eddie & al. (2003); AY 322024 & AY 331437. C. lusitanica L.; (1): Spain, Orense, Rivadavia, R. Pino; HQ 407553. C. lusitanica; (2): Portugal, Sierra de Monchique, Talavera & al.; HQ 407550. C. lusitanica; (3): Portugal, Sierra de Monchique; Talavera & al.; HQ 407551. C. lusitanica L. (4): Spain, Pontevedra, Cangas de Morrazo, S. Castroviejo; HQ 407552. C. marchesettii Witasek; Croatia, Učka; Park & al. (2006); DQ 304612. C. matritensis A. DC.; Spain, Huelva, Hinojos; Cano-Maqueda & al. (2008) as C. lusitanica; FM 212703. C. mirabilis Albov; Royal Botanic Gardens Edinburgh; Eddie & al. (2003); AY 322026 & AY 331439. C. mollis L.; Spain; Eddie & al. (2003); AY 322027 & AY 331440. C. morettiana Rchb.; Italy, Dolomites; Park & al. (2006); DQ 304602. C. ossetica Bieb.; Caucasus; Eddie & al. (2003); AY 322028 & AY 331441. C. patula L.; Spain, Huesca, Hecho (Pyrenees); Cano-Maqueda & al. (2008); FM 212739. C. peregrina L.; Turkey; Eddie & al. (2003); AY 322029 & AY 331442. C. persicifolia L.; cultivated in University of Edinburgh; Eddie & al. (2003); AY 322030 & AY 331443. C. petraea L.; France; Eddie & al. (2003); AY 322031 & AY 331444. C. portenschlagiana Schultes; Croatia, Biokovo; Park & al. (2006); DQ 304600. C. poscharskyana Degen; Croatia, Dubrovnik region; Park & al. (2006); DQ 304601. C. primulifolia L.; (1): Portugal, Sirerra de Monchique, between Monchique and Odemira; Talavera & al.; HQ 407555. C. primulifolia Brot.; (2): Spain, Huelva, Sierra de Aracena; Aldea de las Veredas, Arroyo del Acebuche; Talavera & al.; HQ 407554. C. pulla L.; Austria; northeastern Alps; Park & al. (2006); DQ 304605. C. punctata Lam.; cultivated in University of Edinburgh; Eddie & al. (2003); AY 322033 & AY 331446. C. pyramidalis L.; cultivated in University of Edinburgh; Eddie & al. (2003); AY 322034 & AY 331447.C. raddeana Trautv.; Caucasus; Eddie & al. (2003); AY Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 47 Taxonomic review of the Campanula lusitanica complex 322035 & AY 331448. C. raineri Perpenti; Italy, Alpi Bergamaschi; Park & al. (2006); DQ 304604. C. ramosissima Sibth. & Sm.; Greece, Lakonia; SALA 135597; Aedo & al.; HQ 407548. C. rapunculus L.; Spain, Huelva, Hinojos; Cano-Maqueda & al. (2008); FM 212738. C. reatina Lucchese; Italy, Turano Valley; Park & al. (2006); DQ 304599. C. reverchonii A. Gray; USA, Texas; Eddie & al. (2003); AY 322036 & AY 331449. C. rotundifolia L.; Spain, Cádiz, Grazalema; Cano-Maqueda & al. (2008); FM 212736. C. sarmatica Ker-Gawl.; Caucasus; Eddie & al. (2003); AY 322038 & AY 331451. C. siegizmundii Fed.; Caucasus; Eddie & al. (2003); AY 322039 & AY 331452. C. sosnowskyi Charadze; Caucasus; Eddie & al. (2003); AY 322040 & AY 331453. C. sparsa Friv.; Greece, Grevená, Palaiokastro; SALA 135596; Aedo & al.; HQ 407549. C. specularioides Coss.; Spain, Cádiz, Grazalema; Cano-Maqueda & al. (2008); FM 212705. C. spicata L.; Italy, Southern Alps; Park & al. (2006); DQ 304574.C. stenocodon Boiss. & Reuter; Italy, Alpi Cozie; Park & al. (2006); DQ 304620. C. steveni Bieb.; Caucasus; Eddie & al. (2003); AY 322041 & AY 331454. C. thyrsoides L.; cultivated in University of Edinburgh; Eddie & al. (2003); AY 322042 & AY 331455. C. tommasiana Koch; Croatia, Učka; Park & al. (2006); DQ 304611. C. transtagana R. Fern.; Spain, Córdoba, Azuel (Sierra Morena); Cano-Maqueda & al. (2008); FM 212721. C. tridentata Schreb.; Caucasus; Eddie & al. (2003); AY 322043 & AY 331456. C. uniflora L.; Noeway, Sor-Trondelag; Park & al. (2006); DQ 304588. C. versicolor Andrews; Greece, Ionian Islands, Kefallinía; Park & al. (2006); DQ 304607; C. waldsteiniana Schultes; Croatia, Velebit Mtns.; Park & al. (2006); DQ 304610. C. zoysii Wulfen; Slovenia, Kamniške Alps; Park & al. (2006); DQ 304603. Campanulastrum americanum (L.) Small.; Eddie & al. (2003); AY 322044 & AY 331457. Canarina canariensis (L.) Vatke; Spain, Canary Islands; Eddie & al. (2003); AY 322045 & AY 331458. Codonopsis dicentrifolia W. W. Sm.; Nepal; Eddie & al. (2003); AY 322046 & AY 331459. Craterocapsa congesta Hilliard & B.L. Burtt; Lesotho; Eddie & al. (2003); AY 322049 & AY 331462. Cynanthus lobatus Wall. ex Benth; Eddie & al. (2003); AY 322050 & AY 331463. Diosphaera rumeliana (Hampe) Bornm.; cultivated in University of Edinburgh; Eddie & al. (2003); AY 47 322051 & AY 331464. Edraianthus graminifolius (L.) A. DC.; cultivated in Royal Botanic Gardens Edinburgh; Eddie & al. (2003); AY 322052 & AY 331465. Feeria angustifolia (Schousb.) Buser; Morocco; Eddie & al. (2003); AY 322054 & AY 331467. Gadellia lactiflora (M. Bieb.) Schulkina; cultivated in Royal Botanic Gardens Edinburgh; Eddie & al. (2003); AY 322055 & AY 331468. Galactites tomentosa Moench.; Sussana & al. (2006); AY 826285. Githopsis diffusa A. Gray; Eddie & al. (2003); AY 322056 & AY 331469. Hanabusaya asiatica Nakai; South Korea; Eddie & al. (2003); AY 322057 & AY 331470. Heterocodon rariflorum Nutt.; USA, California; Eddie & al. (2003); AY 322058 & AY 331471. Jasione montana L.; Spain; Eddie & al. (2003); AY 322062 & AY 331475. Legousia falcata (Ten.) Fritsch; cultivated in Royal Botanic Gardens Edinburgh and University of Texas; Eddie & al. (2003); AY 322064 & AY 331477. Leptocodon gracilis Lem.; Nepal; Eddie & al. (2003); AY 322066 & AY 331479. Michauxia tchihatcheffii Fisch. & C.A. Mey.; cultivated in Royal Botanic Gardens Edinburgh; Eddie & al. (2003); AY 322068 & AY 331480. Musschia aurea Dumort.; cultivated in Royal Botanic Gardens Edinburgh and University of Texas; Eddie & al. (2003); AY 322067 & AY 331481. Petromarula pinnata (L.) A. DC.; Greece; Eddie & al. (2003); AY 322069 & AY 331482. Physoplexis comosa (L.) Schur; cultivated in Royal Botanic Gardens Edinburgh; Eddie & al. (2003); AY 322070 & AY 331483. Phyteuma orbiculare L.; Eddie & al. (2003); AY 322071 & AY 331484. Roella ciliata L.; Eddie & al. (2003); AY 322074 & AY 331487. Symphyandra armena (Stev.) A. DC.; cultivated in Royal Botanic Gardens Edinburgh; Eddie & al. (2003); AY 322075 & AY 331488. Trachelium caeruleum L.; cultivated in University of Edinburgh; Eddie & al. (2003); AY 322078 & AY 331491. Triodanis leptocarpa (Nutt.) Nieuwl.; USA, Texas; Eddie & al. (2003); AY 322079 & AY 331492. Wahlenbergia hederacea L.; Portugal, Sierra de Monchique, Talavera & al.; HQ 407556. Anales del Jardín Botánico de Madrid 68(1): 15-47, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2274 Associate Editor: C. Aedo Received: 27-X-2010 Accepted: 25-I-2011 2274 campanula:Maquetación 1 13/06/2011 12:10 Página 48 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 49 Anales del Jardín Botánico de Madrid Vol. 68(1): 49-59 enero-junio 2011 ISSN: 0211-1322 doi: 10.3989/ajbm.2269 Distinguishing colour variants of Serapias perez-chiscanoi (Orchidaceae) from related taxa on the Iberian Peninsula by Caspar Venhuis 1 & J. Gerard B. Oostermeijer 2 Derde Goudsbloemdwarsstraat 21, 1015 KA Amsterdam, The Netherlands. [email protected] Institute for Biodiversity and Ecosystem Dynamics (IBED), Universiteit van Amsterdam, Science Park 904, 1098 XH Amsterdam, The Netherlands. [email protected] 1 2 Abstract Resumen Venhuis, C. & Oostermeijer, J.G.B. 2011. Distinguishing colour variants of Serapias perez-chiscanoi (Orchidaceae) from related taxa on the Iberian Peninsula. Anales Jard. Bot. Madrid 68(1): 49-59. Venhuis, C. & Oostermeijer, J.G.B. 2011. Distinción de variantes en color de Serapias perez-chiscanoi (Orchidaceae) en relación con táxones de la Península Ibérica. Anales Jard. Bot. Madrid 68(1): 49-59 (en inglés). Serapias perez-chiscanoi has a stable and uniform appearance with green flowers. Throughout its distribution area, however, plants have been found with deviant pink to red flowers that show similarities with other taxa that are occasionally pale flowered. S. perez-chiscanoi is easy to differentiate from S. cordigera subsp. cordigera by the colour of the flowers (S. cordigera subsp. cordigera has red to purple flowers) and the fact that the hypochile dimensions of S. perez-chiscanoi are significantly smaller. It is, however, more difficult to distinguish it from individuals of S. cordigera subsp. gentilii with pale flowers, which occur frequently. The two taxa differ in colour pattern and floral dimensions, especially the hypochile length, which is shorter in S. perez-chiscanoi. Pale-flowered individuals of another species, S. parviflora, are easily distinguished by their significantly smaller flowers. S. perez-chiscanoi occurs in Spain in the autonomous regions of Extremadura and Castilla-La Mancha and in Portugal, S. cordigera subsp. gentilii seems to occur along the coastal regions of SW Portugal, while S. cordigera subsp. cordigera and S. parviflora are distributed throughout the Iberian Peninsula. La Serapias perez-chiscanoi tiene una apariencia estable y uniforme con flores verdes. Sin embargo, a lo largo de su área de distribución, se han encontrado ejemplares de flores con coloraciones desviantes de color rosa hasta rojo que muestran similitudes con otros táxones que presentan ocasionalmente flores pálidas. Serapias perez-chiscanoi es fácil de diferenciar con respecto a S. cordigera subsp. cordigera por el color de las flores (S. cordigera subsp. cordigera tiene flores de rojizas hasta púrpura) y por las dimensiones significativamente más pequeñas del hipoquilo de S. perez-chiscanoi. Sin embargo es más difícil hacer una distinción con respecto a S. cordigera subsp. gentilii, ya que los individuos de este taxon presentan flores pálidas, lo que ocurre con frecuencia. Estos dos táxones se diferencian por el patrón de colores y por las pequeñas dimensiones de las piezas florales, especialmente el tamaño del hipoquilo más corto en S. perez-chiscanoi. Los individuos de flores pálidas de S. parviflora se pueden distinguir fácilmente gracias a sus flores de pequeño tamaño. Serapias perez-chiscanoi se localiza en España en las comunidades autónomas de Extremadura y Castilla-La Mancha y en Portugal, S. cordigera subsp. gentilii se puede encontrar en las zonas costeras del suroeste de Portugal, mientras que S. cordigera subsp. cordigera y S. parviflora se encuentran en la mayor parte de la Península Ibérica. Keywords: Orchidaceae, Serapias perez-chiscanoi, Serapias cordigera subsp. gentilii, flower colour, floral dimensions, Spain, Portugal. Palabras clave: Orchidaceae, Serapias perez-chiscanoi, Serapias cordigera subsp. gentilii, color de las flores, dimensiones de las flores, España, Portugal. 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 50 50 C. Venhuis & G. Oostermeijer Introduction In 1976 Jose Luis Pérez Chiscano discovered deviant Serapias plants along the Guadiana river basin in Extremadura (Spain). After a twelve-year study, Pérez Chiscano (1988) described these plants as a new species, Serapias viridis Pérez Chiscano. Acedo (1990), however, found that the same name had been used for a Brazilian species by Vellozo (1825). To avoid confusion, the Spanish species was renamed S. perez-chiscanoi C. Acedo. Pérez Chiscano & al. (1991) reported that only some eight populations of this species were known, all located in the Guadiana river basin in Extremadura. However, due to an increased interest in this species, many new populations were found in Extremadura during the past ten years (Venhuis & al., 2006). Furthermore, the species was also found in Castilla-La Mancha (Venhuis & al., 2006), and also in Portugal (Jansen, 1993). Observations on these recently found populations have increased our knowledge of the species. One new aspect is that plants with deviant reddish flower colours were found among the “normal” S. perez-chiscanoi individuals that have pale green flowers, or pale green flowers with a red venation. In this article, we describe the variation in flower colour in S. perez-chiscanoi and the differences and similarities in morphology and geographic distribution with other Serapias taxa. ed as this species is morphologically readily distinctive from the other taxa. According to the analyses by Venhuis & al (2007), the dimensions of the epichile and hypochile are the most distinctive characters, and so in each population we measured the width and length of both the epichile and hypochile (Fig. 1). Results and discussion Variation in flower colour Serapias perez-chiscanoi in Extremadura has a fairly uniform morphology and flower colour. The plants can be divided into two extremes, which present “green” or “red veined” variants. In the green variation (Fig. 2a, b), the leaves, stem, bracts, ovary, gynostegium, lamellae, flowers and veins are all pale green, with whitish hairs on the labellum. The lateral lobes of the flowers are yellowish and greenish. The “red veined” variation (Fig. 2c, d) is also greenish but with a red venation on the leaves, stem, bracts and ovary. It also has red veins and reddish hairs on the labellum and pinkish to reddish lateral lobes and lamellae. Intermediate colour variations occur very frequently (Venhuis & al., 2004). In Portugal, most of the known populations mainly comprise the “red veined” variation. A population of about 80 flowering plants in C Portugal contained in- Material and methods a In 2004 and 2010, we obtained morphological data for populations of Serapias cordigera subsp. cordigera L., S. cordigera subsp. gentilii C. Venhuis, P. Venhuis & Kreutz, S. perez-chiscanoi and S. parviflora Parl. in Spain and Portugal. For both subspecies of S. cordigera, we measured 25 plants, from one population of subsp. cordigera in Extremadura, and likewise for subsp. gentilii in the Algarve. With S. perez-chiscanoi we measured 75 plants from three populations in Extremadura (Spain), and 5 plants from a population in mid-western Portugal. For S. parviflora, 50 plants were measured from two populations (Algarve and Extremadura) (Table 1). The latter data is not includ- d b Table 1. Sampled populations of the studied Serapias taxa. Species Location S. cordigera subsp. cordigera S. cordigera subsp. gentilii S. perez-chiscanoi S. perez-chiscanoi S. perez-chiscanoi S. perez-chiscanoi Badajoz Cotifo Badajoz Aljucén Trujillanos Ereiras Region Country Extremadura Spain Algarve Portugal Extremadura Spain Extremadura Spain Extremadura Spain Beira Litoral Portugal c Fig. 1. Measured floral dimensions of the labellum: a, hypochile width; b, epichile lenght; c, epichile width; d, hypochile lenght. Anales del Jardín Botánico de Madrid 68(1): 49-59, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2269 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 51 Colour variation in Serapias perez-chiscanoi a b c d 51 Fig. 2. a, Serapias perez-chiscanoi, Obando, Extremadura, Spain, 27-IV-2007; b, S. perez-chiscanoi, Trujillanos, Extremadura, Spain, 29-IV-2010; c, S. perez-chiscanoi, Vila Nova da Baronia, Baixo Alentejo, Portugal, 23-IV-2007; d, S. perez-chiscanoi, Alange, Extremadura, Spain, 30-IV-2010. All photographs: C. Venhuis. Anales del Jardín Botánico de Madrid 68(1): 49-59, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2269 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 52 52 C. Venhuis & G. Oostermeijer a b c d Fig. 3. a, Serapias perez-chiscanoi, Ereiras, Beira Litoral, Portugal, 1-V-2010; b, S. perez-chiscanoi, Ereiras, Beira Litoral, Portugal, 1-V-2010; c, S. perez-chiscanoi, Ereiras, Beira Litoral, Portugal, 1-V-2010; d, S. perez-chiscanoi, Ereiras, Beira Litoral, Portugal, 25-IV-2011. All photographs: C. Venhuis. Anales del Jardín Botánico de Madrid 68(1): 49-59, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2269 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 53 Colour variation in Serapias perez-chiscanoi dividuals with three flower colour variations. About ten plants were of the “green” variation, whilst some 50 plants were quite similar to the “red veined” variation, but differed from it by a bright red hypochile and lateral lobes, an epichile that was both greenish and pinkish, and the sepals, petals and bracts were also often slightly pinkish (Fig. 3a, b). The most deviant plants, however, about 20 individuals, had a bright red labellum (hypochile, lateral lobes and epichile), red petals and red veins on all plant parts. In addition, the bracts and sepals were pinkish (Fig. 3c, d). Hybridization Although hybridization cannot be excluded, the occurrence of hybrids is unlikely. The pollinia of S. perez-chiscanoi flowers disintegrate very rapidly onto their own stigmatic surface, often before the flowers open (Perez Chiscano & al., 1991), and so the possibility that intact pollinia are transferred to another flower is limited. Moreover, S. perez-chiscanoi, is a diploid (Bernardos & al., 2004), while S. lingua L., a species with which it is frequently sympatric, is tetraploid (D’Emerico & al., 2000), so that the difference in ploidy level makes cross-fertility unlikely. In contrast, S. perez-chiscanoi very rarely co-occurs with the diploid S. cordigera, which makes hybridisation between these two species also highly unlikely. Nevertheless, hybrids between S. perez-chiscanoi and both S. lingua and S. cordigera have been reported. A hybrid between S. perez-chiscanoi and S. lingua was mentioned by Wallenwein & Breier (1992) and was described subsequently as S. × venhuisia by Vázquez (2009). However, the photo in Wallenwein & Breier (1992), is clearly of S. lingua; Vázquez did not provide any photographs. Furthermore, Venhuis & al. (2004) and Vázquez (2009) suggested hybridization between S. perez-chiscanoi and S. cordigera. This putative hybrid was based on a plant near Aljucén (Venhuis & al., 2004), from the same population as the photographs presented in this paper (Fig. 4a, b). The inflorescences of these very rare individuals contained salmon-pink and pink flowers, and it is noteworthy that flower colour varied within single plants. However, morphological measurements on the flowers of these ‘hybrid’ individuals revealed floral dimensions identical to S. perez-chiscanoi, and since the other putative parent, S. cordigera, did not occur in the vicinity, and no morphological character of any other Serapias species was present, we now conclude that these plants probably represent a pink-flowered form of S. perez-chiscanoi rather than a hybrid. 53 Related species In the south-western part of the Iberian Peninsula about seven taxa of Serapias occur, which can be divided into two main groups based on their flower size: the S. vomeracea group and the S. parviflora group (Venhuis & al., 2007). In this region, the S. vomeracea group includes S. cordigera subsp. cordigera, S. cordigera subsp. gentilii, S. perez-chiscanoi and S. occidentalis C. Venhuis & P. Venhuis. The occurrence of S. vomeracea subsp. vomeracea in SW Spain and Portugal is uncertain (Venhuis & al., 2007), and the taxonomic status of the new species, S. maria F.M. Váquez (Váquez, 2008), needs further study since the dimensions of morphological characters of this species overlap to a large extent with those of S. occidentalis. Further research on these taxa is necessary to determine their occurrence and taxonomic status respectively. Taxa belonging to the S. parviflora group are S. parviflora, S. strictiflora Welwitsch ex Vega and S. lingua. Furthermore, two varieties of S. strictiflora are found in our region: var. elsae (P. Delforge) C. Venhuis & P. Venhuis, and var. distenta Presser. In the field, S. perez-chiscanoi is easy to distinguish from most other co-occurring Serapias taxa on the basis of flower colour or floral dimensions. However, some taxa that occasionally have pale flowers, such as S. cordigera subsp. gentilii and S. parviflora, resemble S. perez-chiscanoi, and the differences between such individuals and S. perez-chiscanoi are clarified below. Differences with S. cordigera subsp. cordigera Serapias cordigera subsp. cordigera is presumed to be the parental species of S. perez-chiscanoi, and both taxa are similar morphologically (Venhuis & al., 2007) and closely related according to molecular studies (Bellusci & al., 2008). Nevertheless, S. cordigera subsp. cordigera, with its dark red to purple flowers with a large, heart-shaped epichile (Fig. 4c, d) is easily distinguished in the field from S. perez-chiscanoi. It can also be distinguished on the basis of three other features: 1) epichile position, 2) emergence of the lateral lobes and 3) inflorescence architecture. The epichile of S. cordigera subsp. cordigera is normally positioned parallel to the stem (i.e., pointing downwards), whereas the epichile of S. perez-chiscanoi is generally positioned at an angle of about 45-90 degrees to the stem (pointing more or less outwards). In S. cordigera subsp. cordigera, the lateral lobes protrude from the casco, whereas in S. perez-chiscanoi the lateral lobes remain hidden inside the hood. The flowers in S. cordigera subsp. cordigera are placed more or less opposite each other, whilst those of S. perezchiscanoi are positioned close together and in a spi- Anales del Jardín Botánico de Madrid 68(1): 49-59, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2269 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 54 54 C. Venhuis & G. Oostermeijer a b c d Fig. 4. a, Serapias perez-chiscanoi, Aljucén, Extremadura, Spain, 28-IV-2010; b, S. perez-chiscanoi, Aljucén, Extremadura, Spain, 28-IV-2010; c, S. cordigera subsp. cordigera, Badajoz, Extremadura, Spain, 30-IV-2010; d, S. cordigera subsp. cordigera, Badajoz, Extremadura, Spain, 30-IV-2010. All photographs: C. Venhuis. Anales del Jardín Botánico de Madrid 68(1): 49-59, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2269 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 55 Colour variation in Serapias perez-chiscanoi a b c d 55 Fig. 5. a, Serapias cordigera subsp. gentilii, Barranco do Velho, Algarve, Portugal, 13-IV-1995; b, S. cordigera subsp. gentilii, Galaxos, Algarve, Portugal, 27-IV-2010; c, S. cordigera subsp. gentilii, Barranco do Velho, Algarve, Portugal, 26-IV-2010; d, S. cordigera subsp. gentilii, Barranco do Velho, Algarve, Portugal, 17-IV-1995. Photographs: a, d, D. Tyteca; b, c, C. Venhuis. Anales del Jardín Botánico de Madrid 68(1): 49-59, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2269 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 56 56 C. Venhuis & G. Oostermeijer a b Fig. 6. a, Serapias parviflora, Aldeia dos Palheiros, Baixo Alentejo, Portugal, 23-IV-2007; b, S. parviflora, Aldeia dos Palheiros, Baixo Alentejo, Portugal, 23-IV-2007. Photographs: C. Venhuis. ral, which gives the inflorescence a “twisted” appearance. Although there is a small overlap in the epichile dimensions between S. cordigera subsp. cordigera and S. perez-chiscanoi: length (18)23-30(36) mm and (14)16-18(21) mm respectively, and width (13)1724(29) mm and (10)12-13(15) mm respectively, S. cordigera subsp. cordigera can be distinguished from S. perez-chiscanoi on the basis of the non-overlapping hypochile dimensions: length (10)11-14(17) mm and (6)7-8(9) mm respectively, and width (18)21-27(31) mm and (14)16-18(19) mm respectively (Fig. 7). Differences with S. cordigera subsp. gentili Serapias cordigera subsp. gentilii (Fig. 5a-d) is sometimes difficult to differentiate from S. perez-chiscanoi because it occasionally has pale flowers that resemble the latter species. Most flowers of S. cordigera subsp. gentilii are red, but in many populations some plants with pale flowers occur, which vary from red with white edges (resembling S. nurrica Corrias), or completely pink, pink with yellow and pink with reddish veins, to yellowish, greenish or whitish (resembling S. perez-chiscanoi). In addition, several features that are present in S. cordigera subsp. cordigera are absent in S. cordigera subsp. gentilii and in S. perez-chiscanoi: the position of the epichile in the latter taxa generally points more or less outwards (although frequently downwards), and the inflorescence is, when there are many flowers, quite dense and spiralled and the edges of the epichile are often curled upwards. Furthermore, S. cordigera subsp. gentilii, like S. perez-chiscanoi, seems to be autonomously self-pollinating (Venhuis & al., 2007). It differs from S. perez-chiscanoi, however, by lateral lobes that generally emerge from the hood in contrast to S. perez-chiscanoi in which the lateral lobes are always completely hidden inside it. Although the flowers of S. cordigera subsp. gentilii generally have a colour pattern quite different from S. perez-chiscanoi, some individuals have flowers with a greenish epichile that may superficially look similar. But in comparison with the “green” variation of S. perez-chiscanoi, these flowers have a red hypochile, while in comparison with the “red” variation of that taxon, they lack the reddish hairs on the labellum Anales del Jardín Botánico de Madrid 68(1): 49-59, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2269 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 57 Colour variation in Serapias perez-chiscanoi 57 Fig. 7. Boxplots of Serapias cordigera subsp. gentilii, S. cordigera subsp. cordigera, S. perez-chiscanoi from Extremadura and S.perezchiscanoi from Portugal (Ereiras). Outliers and extremes were not removed. a, epichile width; b, epichile length; c, hypochile width; d, hypochile length. Anales del Jardín Botánico de Madrid 68(1): 49-59, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2269 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 58 58 C. Venhuis & G. Oostermeijer and the red venation on all plant parts. The red-flowered S. perez-chiscanoi, which until now was known from only one locality, is also very similar to the flowers of some plants of S. cordigera subsp. gentilii but differs from the latter subspecies by a striking red venation on all plant parts. Most floral features are unhelpful to distinguish between these taxa, and the only character that separates S. cordigera subsp. gentilii from S. perez-chiscanoi is the length of the hypochile, which in S. cordigera subsp. gentilii is (9)11-12(13) mm, and in S. perezchiscanoi is (6)7-8(9) mm (Fig. 7). All floral dimensions as well as the flower colour of S. cordigera subsp. gentilii are more or less intermediate between S. cordigera subsp. cordigera and S. perez-chiscanoi. Differences to S. parviflora Serapias parviflora is generally easily distinguished from S. perez-chiscanoi by its very small flowers. In the studied area, specimens of S. parviflora with pale pink and yellowish/greenish flowers (Figs. 6a, b) frequently occur, but these resemble S. perez-chiscanoi in colour only. Both taxa can be readily distinguished on the basis of three of the four labellum dimensions: Epichile length in S. parviflora ranges from (7)9- 11(12) mm and in S. perez-chiscanoi from (14)1618(21) mm, the epichile width in S. parviflora falls between (2)4-6(6) mm and in S. perez-chiscanoi between (10)12-13(15) mm. Furthermore, S. parviflora differs from S. perez-chiscanoi in hypochile width (8)1011(13) mm and (14)16-18(19) mm respectively. Hypochile length does not differ: (5)7-8(9) mm and (6)78(9) mm, respectively. Distribution Serapias perez-chiscanoi is a rare tongue-orchid, which was previously only known from the Guadiana river basin in Extremadura (Spain) (Pérez Chiscano, 1988; Pérez Chiscano & al., 1991; Delforge, 2002). After a field study, Venhuis & al. (2004) reported six new populations, and after intensive searches during the last five years by, amongst others, employees of the regional government of Extremadura, several new populations were found along the river basin of the Tajo and also south of the Guadiana river basin in Extremadura (Venhuis & al., 2006), which increases the total number of populations known in Extremadura to around 30 (Fig. 8). Yet another population was found in Castilla-La Mancha (Venhuis & al., 2006). In the Algarve (Portugal), Jansen (1993) found a population that disappeared soon after its discovery. And we have seen a population in the Baixo Alentejo province that was discovered by M. Pereira, and also four other populations that were discovered by either J. Moura, J. Pessoa and J. Monteiro, in the provinces of Beira Litoral and Ribatejo in the central part of Portugal and in the province of Trás-os-Montes e Alto Douro in northern Portugal (Fig. 8). Serapias cordigera subsp. cordigera is found throughout the Iberian Peninsula, sometimes only locally but often abundantly. Serapias parviflora also occurs in the entire Iberian Peninsula, but is much more widespread and often abundant. Serapias cordigera subsp. gentilii is found predominantly along the coastal regions of the Algarve but also extends further north. The distribution map (Fig. 8) is based on populations seen by us, and photos, and also on literature, in which it was cited as a variety of S. cordigera. Identification key Fig. 8. Distribution map of Serapias perez-chiscanoi and S. cordigera subsp. gentilii. All known populations are presented, including populations that have already disappeared ( S. perezchiscanoi; S. cordigera subsp. gentilii). Here, we present a concise key for the identification of species from the large flowered S. vomeracea group in the SW Iberian Peninsula. It will be clear from this article that the identification of the different species is not too difficult, despite the considerable variation in flower morphology, flower colours and venation patterns. More research on the relationships Anales del Jardín Botánico de Madrid 68(1): 49-59, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2269 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 59 Colour variation in Serapias perez-chiscanoi between the morphological variation and the pollination ecology in S. perez-chiscanoi is underway. IDENTIFICATION KEY FOR THE SERAPIAS VOMERACEA GROUP IN THE SOUTHWESTERN PART OF THE IBERIAN PENINSULA 1. Ratio epichile width/hypochile length = 0.6-1.2 ................... ................................................................... S. occidentalis 1. Ratio epichile width/hypochile length = 1.3-2.1 ................ 2 2. Hypochile length (6)7-8(9) mm ............. S. perez-chiscanoi 2. Hypochile length (9)10-14(17) mm ................................... 3 3. Epichile broad and heart-shaped, purple, with no divergent edges; pollinia coherent ...... S. cordigera subsp. cordigera 3. Epichile slender, usually pale, often with divergent edges; pollinia friable ......................... S. cordigera subsp. gentilii Acknowledgements We thank Marizia Pereira, Daniel Tyteca, Joaquim Pessoa, Jorge Moura and Karel Kreutz for their kind help with providing localities and/or photos of the studied species. Marisela Cornado Garcia is kindly acknowledged for translating the summary into Spanish. References Acedo, C. 1990. Serapias perez-chiscanoi, nom. nov. Anales del Jardin Botánico de Madrid 47(2): 510. Bellusci, F., Pellegrino, G., Palermo, A.M. & Musacchio, A. 2008. Phylogenetic relationships in the orchid genus Serapias L. based on noncoding regions of the chloroplast genome. Molecular Phylogenetics and Evolution 47: 986-991. Bernardos, S., Tyteca, D. & Amich, F. 2004. Cytotaxonomic study of some taxa of the subtribe Orchidinae (Orchidoideae, Orchidaceae) from the Iberian Peninsula. Israel Journal of Plant Sciences 52: 161-170. Delforge P. 2002. Guía de las Orquídeas de España y Europa. Lynx Ediciones, Barcelona, Spain. 59 D’Emerico, S., Pignone, D. & Scrugli, A. 2000. Giemsa C-banded karyotypes in Serapias L. (Orchidaceae). Botanical Journal of the Linnean Society 133: 485-492. Jansen H. 1993. Serapias viridis Pérez Chiscano in Portugal! Mitteilungsblatts des Arbeitskreises Heimische Orchideen BadenWürttemberg 10: 50-53. Pérez Chiscano, J.L. 1988. Nueva especie de Serapias L. en Extremadura (España). Monografias del Instituto Pirenaico de Ecologia, Homenaje a Pedro Montserrat, Jaca y Huesca: 305-309. Pérez Chiscano, J.L., J.R. Gil Llano & F. Duran Oliva. 1991. Orquídeas de Extremadura. Fonda Natural, Madrid. Tyteca, D. 1997. The orchid flora of Portugal. Journal Europäischer Orchideen 29(2/3): 185-581. Vázquez Pardo, F.M. 2008. Annotations to the Orchidaceae of Extremadura (SW Spain). Journal Europäischer Orchideen 40(4): 699-725. Vázquez Pardo, F.M. 2009. Revisión de la familia Orchidaceae en Extremadura (España). Folia Botanica Extremadurensis 3: 5-368 Vellozo, J.M. da C. 1825. Florae fluminensis. Senefelder, Rio de Janeiro, Brasil. Venhuis, C., Oostermeijer, J.G.B. & Tonk, J.Th.P. 2004. Conservation biology of Serapias perez-chiscanoi Acedo in the Guadiana river basin in Extremadura (Spain). Eurorchis 16: 49-63. Venhuis, C., Oostermeijer, J.G.B. & Cornado Garcia, M. 2006. Serapias perez-chiscanoi: legal protection and distribution. Eurorchis 18: 88-91. Venhuis, C., Venhuis, P., Oostermeijer, J.G.B. & van Tienderen, P.H. 2007. Morphological systematics of Serapias L. (Orchidaceae) in Southwest Europe. Plant Systematics and Evolution 265: 165-177. Wallenwein, F. & Breier, W. 1992. Bemerkungen zu einigen Arten der Gattung Serapias L. aus Spanien. Mitteilungsblatts des Arbeitskreises Heimische Orchideen Baden-Württemberg 24(1): 115-121. Anales del Jardín Botánico de Madrid 68(1): 49-59, enero-junio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm: 2269 Associate Editor: L. Sáez Received: 27-IX-2010 Accepted: 7-II-2011 2269 serapias:Maquetación 1 13/06/2011 12:11 Página 60 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 61 Anales del Jardín Botánico de Madrid Vol. 68(1): 61-95 enero-julio 2011 ISSN: 0211-1322 doi: 10.3989/ajbm.2266 Biodiversity of Myxomycetes from the Monte Desert of Argentina by C. Lado1, D. Wrigley de Basanta1 & A. Estrada-Torres2 Real Jardín Botánico, CSIC, Plaza de Murillo 2, E-28014 Madrid, Spain. [email protected] Centro de Investigación en Ciencias Biológicas, Universidad Autónoma de Tlaxcala, km 10.5 carretera Texmelucan-Tlaxcala, Ixtacuixtla, 90122, Tlaxcala, México. [email protected] 1 2 Abstract Abstract Lado, C., Wrigley de Basanta, D. & Estrada-Torres, A. 2011. Biodiversity of Myxomycetes from the Monte Desert of Argentina. Anales Jard. Bot. Madrid 68(1): 61-95 A biodiversity survey for myxomycetes was carried out in the Monte Desert (Argentina) and surrounding areas in November 2006 and late February and March 2007. Specimens were collected in seven different provinces (Catamarca, Jujuy, La Rioja, Salta, San Juan, San Luis and Tucumán), between 23º and 33º S latitude, and a total of 105 localities were sampled. Cacti and succulent plants were the most common type of substrate investigated, but shrubs and herbs characteristic of this biome were also included in the survey. Almost six hundred specimens of myxomycetes from 72 different species in 22 genera were collected either in the field, or from moist chamber cultures prepared with samples of plant material obtained from the same collecting sites. The results include 1 species new to science, Macbrideola andina three more species recently described based on material from this survey, 5 species cited for the first time for the Neotropics, 11 new records for South America and 38 new records for Argentina. Taxonomic comments on rare or unusual species are included and illustrated with photographs by LM and SEM. Data are presented on the development of some species and microenvironmental factors are discussed. An analysis of the biodiversity of myxomycetes in this area, and a comparison with other desert areas, are included. Lado, C., Wrigley de Basanta, D. & Estrada-Torres, A. 2011. Biodiversidad de Myxomycetes en el Desierto de Monte (Argentina). Anales Jard. Bot. Madrid 68(1): 61-95 (en inglés). Con el objetivo de estudiar la biodiversidad de Myxomycetes en el Desierto de Monte (Argentina) y áreas circundantes, se realizó un muestreo en los meses de noviembre de 2006 y febrero y marzo de 2007. Se recolectaron especímenes en un total de 105 localidades pertenecientes a siete provincias (Catamarca, Jujuy, La Rioja, Salta, San Juan, San Luis y Tucumán), situadas entre los paralelos 23º y 33º de latitud sur. Los cactus y plantas suculentas fueron los tipos de sustratos más estudiados, pero también se analizaron arbustos y plantas herbáceas características de este bioma. Casi 600 especímenes de mixomicetes pertenecientes a 72 especies y 22 géneros fueron recolectados en el campo o se obtuvieron en el laboratorio, por cultivo en cámara húmeda, a partir de plantas procedentes de las mismas localidades. Los resultados incluyen una nueva especie, Macbrideola andina, otras tres recientemente descritas y basadas en material de este estudio, 5 especies que se citan por primera vez para el Neotrópico, 11 nuevos registros para América del Sur y 38 nuevos registros para Argentina. Se añaden comentarios taxonómicos e ilustraciones fotográficas, tanto con microscopía óptica como electrónica, de aquellas especies raras o poco comunes. Se discuten nuevos datos sobre el desarrollo de algunas especies y cómo influyen determinados factores microambientales. También se incluye un análisis de la biodiversidad de mixomicetes en esta zona árida y se compara con la obtenida en otros desiertos de América. Keywords: Amoebozoa, arid environments, Neotropics, Protista, SEM, slime mould, taxonomy. Palabras clave: Amoebozoa, hongos mucilaginosos plasmodiales, MEB, Neotrópico, Protista, taxonomía, zonas áridas. Introduction free-living amoebae, and the social amoebae (dictyostelids), in a protists-like group called the Amoebozoa (Adl & al., 2005; Baldauf, 2008). They live in almost all terrestrial ecosystems and are particularly abundant in temperate and tropical forests (Ing, 1994; Rojas & Stephenson, 2008; Kosheleva & al., 2008), but The myxomycetes are a group of holotrophic eukaryotic organisms of worldwide distribution that for many years have been regarded as related to fungi, but are now included, together with several groups of 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 62 62 C. Lado & al. many species are also known to be present in warm dryland ecosystems (Evenson, 1961; Blackwell & Gilbertson, 1980; Novozhilov & al., 2003; Lado & al., 2007a; Estrada-Torres & al., 2009; Ndiritu & al., 2009). Most of the literature describes studies of deserts from North America, and has indicated that a special myxobiota, adapted to arid conditions, and much more numerous and varied than previously imagined, may develop in these environments (Lado & al., 1999; Estrada-Torres & al., 2009). To see if the patterns of distribution continue into South America, and to further investigate the relationships that exist between myxomycetes and the plants that live in these drylands, an intensive study was undertaken of a selected arid area of the Monte Desert. From a biogeographical point of view, the Monte Desert, with an area of about 467,000 km2, forms part of the ecoregion of Neotropic deserts (Roig & al., 2009). The Monte Desert, a subtropical to warm temperate desert and semidesert, is located entirely in Argentina. It extends approximately from 23° to 42° South latitude along the eastern border of the Andes, from close to Bolivia curving down to the Atlantic coast at Peninsula Valdés. There are very few published studies of myxomycetes from this region of Argentina. Spegazzini (1899) and Fries (1903) reported on the first data from this area, and Digilio (1946, 1950), and Deschamps (1972, 1976) compiled catalogues including some data from Catamarca, Jujuy, Salta and Tucumán, but few of these were from the Monte Desert. This information forms part of the catalogue of the country published by Crespo & Lugo (2003). In addition, a biodiversity inventory of myxomycetes, from the Chilean Atacama desert, that runs parallel to the northern part of the Monte Desert, on the other side (West) of the Andes, was completed by Lado & al. (2007a). Study area The area studied included the northern and central parts of the Monte Desert in Northwest Argentina, and the bordering transition zones of prepuna and puna in the provinces of Jujuy, Salta, Catamarca, Tucumán, La Rioja, San Juan and San Luis (Fig. 1). The elevation gradient was from 500 m to 4500 m, from the valleys and endorheic basins to the pre-Andean and Andean mountains. The soils of the Monte Desert are very poor, sandy or rocky, with very little humus. The climate of the area is semiarid to arid, with a mean annual rainfall between 50 and 450 mm, being among the most arid areas of Argentina, and the mean annual temperature is between 10 and 18 °C. The area is in the rain shadow of the Andes, and rapid evaporation, increased even more by windy condi- tions in the South, also contributes to the aridity of the region (Abraham & al., 2009). The vegetation of the Monte (Figs. 2-9) is composed of shrub steppes dominated by Larrea spp., spiny shrubs such as Chuquiraga spp., and including several tree species of Prosopis (Roig & al., 2009). In addition, several kinds of cacti of the genera Denmoza, Maihueniopsis, Cereus, Echinopsis, Opuntia, Tephrocactus and Trichocereus are present in the area or among the bordering vegetation types at higher altitudes, where the grasslands of the puna (species of Festuca, Poa and Stipa) also formed part of the present study. In the basins extensive salt flats form, due to the constant evaporation caused by intense solar radiation and the lack of precipitation, and in those areas typical halophytic vegetation is found. Material and methods Sampling was carried out in 105 localities (Table 1) in two expeditions during November to December 2006 and February to March 2007 to coincide with the austral Spring and the end of the summer of the same phenological year. At each locality, the microhabitats in which myxomycetes are known or suspected to occur were examined carefully. All localities were geo-referenced with a portable GPS unit (Magellan eXplorist 600 5.1, Datum WGS84). Samples were collected in the field and substrate samples were also removed for moist chamber culture. Methods used for collecting myxomycetes in the field and substrates for laboratory culture can be found in Stephenson (1989), Rossman & al. (1998). The field work was done by the three authors and approximately one hour was spent in each collecting locality. Moist chamber cultures were prepared using sterile disposable plastic Petri dishes (9 cm diameter), in the manner described by Wrigley de Basanta & al. (2009). The pH of each culture was determined with a portable pH meter after 24 hours, and then excess water in each dish was poured off. Cultures were maintained at room temperature (21-25 ºC) in diffuse daylight and examined at regular intervals with a dissecting microscope for a period of three months. As the myxomycetes matured, the portion of the substrate upon which they occurred was removed from the moist chamber culture, allowed to dry slowly in a closed empty petri dish and then glued in a small cardboard box. All sporophores of a given species that developed in the same culture, were considered to represent a single record. All specimens are deposited in the MA-Fungi herbarium (sub Lado) with duplicates in TLXM herbarium (sub aet), or in the private collection of Wrigley Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 63 Biodiversity of Myxomycetes 63 Fig. 1. Map of Argentina showing the collecting localities (numbers). de Basanta (dwb). Differential interference contrast (DIC) microscopy was used to obtain descriptive data. The light photomicrographs were made using a Nikon AZ100 microscope and combining sequential images. Specimens were examined and photographed at 10-15 kV, with a Hitachi S-3000N scanning electron microscope (SEM), in the Real Jardín Botánico, CSIC. For all SEM-photographs the critical point dried material technique was employed. Colour notations in parenthesis are from the ISCC-NBS Color Name Charts Illustrated with Centroid Colors (Anonymous, 1976). Taxonomic diversity was examined using the mean number of species per genus (S/G),which has been Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 64 64 C. Lado & al. Figs. 2-9. Some characteristic plants of the Monte Desert and sourrounding areas: 2, desert scrub with Larrea spp. (jarillas). 3, Trichocereus sp. and the rosette-leaved plant Puya sp. 4, cardonal of Trichocereus pasacana and Opuntia sp. 5, Prosopis sp. 6, Xerophyllous scrubland with “retamo” (Bulnesia retama) at El Leoncito National Park. 7, Mahiueniopsis sp. 8, Grassland near to the saltflats in the Andes. 9, Grassland of the puna (species of Festuca, Poa and/or Stipa). Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 Jujuy, Tumbaya, on route RN-52, Salinas Grandes Salta, La Poma, Salinas Grandes, Cerro Negro, on route RN-52, km 43 Jujuy, Susques, Angosto del Taire, on route RN-52 Jujuy, Susques, Paso de Jama, customs office, on route RN-52 ARG-06-12 ARG-06-13 ARG-06-14 ARG-06-15 Andean puna with Stipa sp. and salt flats Andean puna with Stipa atacamensis Salt flat with leguminous tree Andean puna with Gramineae Andean puna with Stipa sp. and cacti Andean puna with Stipa sp. Xerophyllous scrubland with Puya sp. Desert scrub with Echinopsis atacamensis, and Opuntia sp. Desert scrub with Tephrocactus and Puya sp. 21 Nov. 2006 21 Nov. 2006 20 Nov. 2006 20 Nov. 2006 20 Nov. 2006 20 Nov. 2006 20 Nov. 2006 20 Nov. 2006 20 Nov. 2006 19 Nov. 2006 19 Nov. 2006 19 Nov. 2006 19 Nov. 2006 18 Nov. 2006 18 Nov. 2006 18 Nov. 2006 Date Biodiversity of Myxomycetes 23°14’14”S 67°01’45”W, 4102 m ± 4 m 23°25’05”S 66°29’54”W, 4001 m ± 8 m 23°25’28”S 66°10’38”W, 3513 m ± 6 m 23°35’10”S 65°53’58”W, 3425 m ± 8 m 23°42’22”S 65°40’20”W, 4147 m ± 7 m 23°41’39 , 65°38’58”W, 4149 m ± 9 m Jujuy, Tumbaya, Abra de Potrerillos pass, on route RN-52, East of Saladillas 23°52’12”S 65°27’50”W, 2112 m ± 8 m ARG-06-11 Jujuy, Tumbaya, Volcán, Huajra ARG-06-08 Pre-puna with Echinopsis atacamensis, Opuntia sulphurea and leguminous trees Pre-puna with leguminous plants, Compositae and Oreocereus trollii Desert scrub with Echinopsis atacamensis and Opuntia sulphurea Spiny scrubland with Echinopsis atacamensis, Cereus uruguayensis and Trichocereus thelegonus Spiny scrubland with Echinopsis atacamensis and Cereus uruguayensis Spiny scrubland with Acacia sp. and Opuntia quimilo Vegetation 23°21’33”S 65°20’42”W, 2780 m ± 11 m Desert scrub with Echinopsis atacamensis, Opuntia sulphurea and leguminous trees Jujuy, Tumbaya, Abra de Potrerillos pass, on route RN-52, El Quemado Jujuy, Humahuaca, 16 km South of Humahuaca, on route RN-9, Chucalesna ARG-06-07 23°06’28”S 65°22’26”W, 3218 m ± 6 m ARG-06-10 Jujuy, Humahuaca, 12 km North of Humahuaca, on route RN-9 ARG-06-06 23°02’03”S 65°22’50” W, 3419 m ± 7 m 23°40’08”S 65°34’33”W, 2972 m ± 6 m Jujuy: Humahuaca, 20 km North of Humahuaca, on route RN-9, 6 km from Hornaditas, by Sapaua stream ARG-06-05 23°40’47”S 65°26’50” W, 2346 m ± 8 m Jujuy, Tumbaya, Purmamarca, on route RN-52, 12 Km West of Purmamarca, Paso de Lipán Jujuy: Tumbaya, North of Volcán, on route RN-9, Cieneguillas ARG-06-04 25°04’42”S 65°00’12”W, 818 m ± 8 m ARG-06-09 Salta: General Güemes, Virgilio Tedín, on route RN-9, Km 1515 ARG-06-03 26°01’24”S 65°07’53”W, 965 m ± 9 m 23°41’58”S 65°32’53”W, 2678 m ± 8 m Salta: Candelaria, Rosario de la Frontera, on route RN-9, km 1393 ARG-06-02 26°31’23”S 65°18’18”W, 751 m ± 7 m Coordinates, elevation ARG-06-08bis Jujuy, Tumbaya, Purmamarca, La Ciénaga, on route RN-52 Tucumán: Trancas, Vipos, on route RN-9, km 1331 ARG-06-01 Locality Table 1. List of collecting localities. 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 65 65 Jujuy, Susques, Paso de Jama, Salar de Jama, on route RN-52 Jujuy, Susques, southern end of the Salar de Jama, on route RN-52 Jujuy, Susques, Archibarca, 68 Km Southwest of Susques, on route RN-52 Jujuy, Susques, Salar de Olaroz, 43.6 km Southwest of Susques, on route RN-52 Jujuy, Susques, 9 km East of Susques, on route RN-52 Jujuy, Susques, 20 km East of Susques, on route RN-52, Quebrada Malpaso Salta, La Poma, Río Negro, on route RP-75 to Cobres, km 24 Salta, Los Andes, San Antonio de los Cobres, La Polvorilla viaduct, mine Concordia Salta, La Poma, San Antonio de los Cobres, Abra Blanca, 129 km Northwest of Salta, on route RN-52 Salta, Rosario de Lerma, Tastil, 119 km Northwest of Salta, on route RN-51, La Encrucijada Salta, San Carlos, San Fernando de Escoipe, cemetery, on route RP-33, km 33 Salta, Cachi, San Martín, Cuesta del Obispo, on route RP-33, km 50 Salta, Cachi, Los Cardones National Park, Tin Tin straight Salta, Cachi, on route RN-40 from Payogasta to La Poma, km 4513 Salta, Cachi, on route RN-40 from Payogasta to La Poma, km 4539, Pueblo Viejo, 2 km North of Rodeo ARG-06-16 ARG-06-17 ARG-06-18 ARG-06-19 ARG-06-20 ARG-06-21 ARG-06-22 ARG-06-23 ARG-06-24 ARG-06-25 ARG-06-26 ARG-06-27 ARG-06-28 ARG-06-29 ARG-06-30 Locality Table 1. List of collecting localities. (Continuation). 24°50’58”S 66°09’12”W, 2703 m ± 6 m 25°02’32”S 66°05’28”W, 2490 m ± 6 m 25°12’08”S 66°58’27”W, 2911 m ± 5 m 25°10’27”S 65°49’31”W, 2744 m ± 13 m 25°09’54”S 65°44’07”W, 1900 m ± 8 m 24°21’06”S 66°04’18”W, 3559 m ± 5 m 24°19’30”S 66°07’02”W, 4001 m ± 7 m 24°12’21”S 66°24’05”W, 4160 m ± 6 m 24 Nov. 2006 23 Nov. 2006 23 Nov. 2006 23 Nov. 2006 22 Nov. 2006 22 Nov. 2006 22 Nov. 2006 22 Nov. 2006 22 Nov. 2006 21 Nov. 2006 21 Nov. 2006 21 Nov. 2006 21 Nov. 2006 21 Nov. 2006 Date Spiny scrubland with Echinopsis atacamensis and cacti 24 Nov. 2006 Cultivated area with Prosopis sp. Desert scrub with Echinopsis atacamensis and bushes Dry gully with bushes (Compositae, Solanaceae and Sambucus sp.) and grasses Spiny scrubland with Puya sp., leguminous plants and Cereus uruguayensis Desert scrub with Echinopsis atacamensis Andean puna with Cumulopuntia sp. and grasses Spiny scrubland with Cumulopuntia sp. and Compositae Andean puna with Prosopis sp., bushes and grasses Scrubland with Cumulopuntia sp. Borders of a stream with Cortaderia sp. and bushes High elevation wetland with Stipa sp. High elevation wetland with Stipa sp. Andean puna with Stipa sp. and salt flats Andean puna with Stipa sp. and salt flats Vegetation 66 23°36’43”S 66°13’55”W, 3442 m ± 5 m 23°25’51”S 66°16’01”W, 3723 ± 7 m 23°26’39”S 66°19’00”W, 3760 ± 9 m 23°33’29”S 66°39’36”W, 3931 m ± 5 m 23°37’19”S 66°51’18”W, 4043 m ± 8 m 23°24’38”S 66°57’08”W, 4117 m ± 8 m 23°15’15”S 67°00’27”W, 4100 m ± 7 m Coordinates, elevation 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 66 C. Lado & al. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 Salta, Cachi, Buena Vista, Potrero river, on route RN-40, km 4519 Salta, Cachi, Los Cardones National Park, Tin Tin straight, 2 km up an unmarked path Salta, Cachi, Los Cardones National Park, Tin Tin straight, up a path to a copse Salta, Cachi, Los Cardones National Park, on route RP-42, km 17 Salta, Cachi, Los Cardones National Park, on route RP-42, km 5 Salta, Cachi, El Algarrobal Salta, Molinos, on route RN-40, km 4472 Salta, Molinos, Seclantás, on route RN-40, km 4467 Salta, Molinos, Molinos river, on route RN-40, km 4458 Salta, Molinos, Angostura, on route RN-40, km 4445 Salta, San Carlos, Corte la Flecha, on route RN-40, km 4406 Salta, San Carlos, Los Sauces, on route RN-40, km 4380 Salta, Cafayate, on route RN-40, km 4332 Tucumán, Tafí del Valle, Quilmes, Quilmes archeaological ruins Catamarca, Santa María, Punta de Balasto, on route RN-40, km 4219 Catamarca, Santa María, Guanaco Yacu, on oute RN-40, km 4206 Catamarca, Santa María, Guanaco Yacu, on route RN-40, km 4203 ARG-06-31 ARG-06-32 ARG-06-33 ARG-06-34 ARG-06-35 ARG-06-36 ARG-06-37 ARG-06-38 ARG-06-39 ARG-06-40 ARG-06-41 ARG-06-42 ARG-06-43 ARG-06-44 ARG-06-45 ARG-06-46 ARG-06-47 Locality Table 1. List of collecting localities. (Continuation). Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 26°59’56”S 66°15’13”W, 219 1m ± 7 m 27°00’09”S 66°14’00”W, 2188 m ± 8 m 26°56’46”S 66°07’51”W, 2132 m ± 6 m 26°28’06”S 66°01’56” W, 1849 m ± 6 m 26°08’40”S 65°57’30”W, 1640 m ± 9 m 25°47’06”S 65°58’04”W, 1861 m ± 13 m 25°42’29”S 66°07’17”W, 1951 m ± 8 m 25°29’29”S 66°14’10”W, 1986 m ± 7 m 25°25’56”S 66°17’11”W, 2059 m ± 10 m 25°21’47”S 66°16’52”W, 2238 m ± 6 m 25°18’49”S 66°14’50”W, 2135 m ± 6 m 25°05’22”S 66°04’57”W, 2684 m ± 9 m 25°15’06”S 66°06’30”W, 2660 m ± 8 m 25°12’47”S 66°01’06”W, 2805 m ± 5 m 25°11’22”S 65°59’29”W, 2880 m ± 7 m 25°10’13”S 66°00’11”W, 2907 m ± 6 m 25°00’13”S 66°06’03”W, 2504 m ± 6 m Coordinates, elevation Spiny scrubland on sand dunes with grasses and succulent-leaved bushes Spiny scrubland on sand dunes with grasses and succulent-leaved bushes Spiny scrubland on sand with Prosopis sp. Desert scrub with Trichocereus sp., Echinopsis sp. and leguminous plants Desert scrub with Puya sp. in a rocky area Cultivated land with Prosopis sp. Spiny scrubland Spiny scrubland with Gymnocalycium sp., Austrocylindropuntia sp. and Denmoza sp. Prosopis sp. woodland Spiny scrubland with Puya sp. Spiny scrubland with Opuntia sulphurea Desert scrub with Echinopsis atacamensis and a copse of Prosopis alba and Prosopis nigra Desert scrub with Echinopsis atacamensis Desert scrub with Echinopsis atacamensis, Cortaderia sp. and Asteraceae Desert scrub with Echinopsis atacamensis and leguminous plants Desert scrub with Echinopsis atacamensis and bushes Scrubland along a dry stream Vegetation 26 Nov. 2006 26 Nov. 2006 26 Nov. 2006 26 Nov. 2006 25 Nov. 2006 25 Nov. 2006 25 Nov. 2006 25 Nov. 2006 25 Nov. 2006 25 Nov. 2006 25 Nov. 2006 24 Nov. 2006 24 Nov. 2006 24 Nov. 2006 24 Nov. 2006 24 Nov. 2006 24 Nov. 2006 Date 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 67 Biodiversity of Myxomycetes 67 Catamarca, Santa María, Sierra del Hombre Muerto, on route RN-40, km 4167 Catamarca, Belén, Hualfin, Los Nacimientos, on route RN-40, km 4151 Catamarca, Belén, on route RN-40 from Belén to Hualfin, 4 km South of Hualfin Catamarca, Belén, 7 km North of Belén, on route RN-40, Reserva Natural Morro de los Cóndores Catamarca, Balén, Londres, up a track 5 km West from route RP-3 to Tinogasta, La Aguada Catamarca, Tinogasta, Andaluca, on route RN-60, km 1277 Catamarca, Belén, Hualfin, Los Nacimientos, on route RN-40, km 4143 Catamarca, Tinogasta, Fiambalá, on route RN-60, 59 km Southeast of San Francisco pass, Las Losas Catamarca, Tinogasta, Fiambalá, on route RN-60, 34 km Easth of San Francisco pass, Las Peladas Catamarca, Tinogasta, Fiambalá, on route RN-60, 35 km Easth of San Francisco pass, Las Peladas Catamarca, Tinogasta, Fiambalá, Valle de Chaschuil, on route RN-60 to San Francisco pass, 50 km West of Fiambalá Catamarca, Tinogasta, Fiambalá, Valle de Chaschuil, on route RN- 60 to San Francisco pass, Cañón de Angosturas Catamarca, Tinogasta, on route RN-60, km 1317, 10 km South of La Puntilla Catamarca, Tinogasta, Costa de Reyes, on route RP-3 ARG-06-48 ARG-06-49 ARG-06-50 ARG-06-51 ARG-06-52 ARG-06-53 ARG-06-54 ARG-06-55 ARG-06-56 ARG-06-57 ARG-06-58 ARG-06-59 ARG-06-60 ARG-06-61 Locality Table 1. List of collecting localities. (Continuation). 28°16’18”S 67°38’51”W, 1437 m ± 7 m 28°06’13”S 67°30’52”W, 1184 m ± 8 m 27°42’16”S 67°56’47”W, 2644 m ± 11 m 27°47’09”S 68°04’55”W, 3085 m ± 10 m 27°01’45”S 68°04’00”W, 3928 m ± 8 m 26°55’33”S 68°04’49”W, 4141 m ± 5 m 27°12’28”S 68°06’25”W, 3764 m ± 7 m 27°12’36”S 66°47’20”W, 1938 m ± 7 m Desert scrub with rosette-leaved succulent plants . (Puya sp.), Opuntia sp. and Prosopis sp. Desert scrub with Puya sp. Rocky outcrop with Opuntia sp., Parodia sp. . and Cumulopuntia sp Scrub with halophytes, bushes, Parodia sp. and Cumulopuntia sp. Andean puna with Stipa sp. Andean puna Pre-puna with Cumulopuntia sp. Spiny scrubland with Gymnocalycium sp., Prosopis sp. and Cercidium sp. Spiny scrubland on sand dunes with Leguminosae and opuntioid cacti Thorn forest with Leguminosae (Brea sp.) Desert scrub with Puya sp., and grasses Desert scrub with Puya sp., and cacti (Trichocereus sp.) Source of a spring with herbaceous plants and grasses (Cortaderia sp.) Scrubland with Asteraceae Vegetation 29 Nov. 2006 29 Nov. 2006 28 Nov. 2006 28 Nov. 2006 28 Nov. 2006 28 Nov. 2006 28 Nov. 2006 26 Nov. 2006 27 Nov. 2006 27 Nov. 2006 27 Nov. 2006 27 Nov. 2006 26 Nov. 2006 26 Nov. 2006 Date 68 28°18’37”S 67°17’36”W, 1035 m ± 9 m 27°45’09”S 67°12’26”W, 1328 m ± 6 m 27°34’13”S 67°00’10”W, 1308 m ± 16 m 27°36’55”S 67°01’06”W, 1305 m ± 6 m 27°10’02”S 66°44’14”W, 2049 m ± 7 m 27°05’14”S 66°37’01”W, 2305 m ± 5 m Coordinates, elevation 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 68 C. Lado & al. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 La Rioja, Arauco, Aimogasta, Villa Mazán, on route RN-60 Catamarca, Capayán, 7 km West of Chumbicha, on route RN-60 ARG-06-75 ARG-06-76 La Rioja, Capital, La Rioja, Los Padercitos, on route RN-75, km 6 5ARG-06-70 La Rioja, Arauco, Aimogasta, 2 km South of Aimogasta, on route RN-75 San Juan, Valle Fértil, El Agua de Arriba, on route RP-510, km 21 ARG-06-69 ARG-06-74 San Juan, Valle Fértil, Ischigualasto Provincial Park, on route RP-510 to the Park, km 104 ARG-06-68 La Rioja, Castro Barros, Pinchas, on route RN-75 La Rioja, Independencia, Talampaya National Park, on route RP-26, km 99 ARG-06-67 ARG-06-73 La Rioja, General F. Varela, Villa Unión, Talampaya National Park, on route RP-26, km 143 ARG-06-66 La Rioja, Sanagasta, 3 km West of Sanagasta, on route RN-75 La Rioja, General F. Varela, Villa Unión, Pagancillo, on route RP-26, km 174 ARG-06-65 ARG-06-72 La Rioja, Vinchina, Famatina, El Potrerillo, on route RN-78, km 3 ARG-06-64 La Rioja, Sanagasta, Dique Los Sauces, on route RN-75, km 15 La Rioja, Vinchina, Campanas, on route RP-11, km 51 ARG-06-63 ARG-06-71 Catamarca, Tinogasta, Costa de Reyes, 34 km South of Tinogasta, on route RP-3 ARG-06-62 Locality Table 1. List of collecting localities. (Continuation). 28°51’42”S 66°23’58”W, 582 m ± 8 m 28°39’02”S 66°33’05”W, 950 m ± 10 m 28°34’37”S 66°49’10”W, 906 m ± 5 m 28°55’46”S 66°57’32”W, 1370 m ± 9 m 29°20’27”S 67°00’25”W, 914 m ± 10 m 29°22’41”S 66°58’58”W, 858 m ± 8 m 29°24’20”S 66°56’35”W, 681 m ± 9 m 30°07’54”S 67°03’27”W, 599 m ± 7 m 30°10’44”S 67°48’56”W, 1374 m ± 12 m 30°07’42”S 67°44’19”W, 1378 m ± 8 m 29°49’03”S 67°59’06”W, 1229 m ± 9 m 29°34’00”S 68°05’17”W, 1162 m ± 8 m 28°59’00”S 67°30’51”W, 1390 m ± 7 m 28°36’32”S 67°38’25”W, 1777 m ± 10 m 28°23’12”S 67°39’44”W, 1647 m ± 5 m Coordinates, elevation Dry forest with Opuntia quimilo Desert scrubland with Opuntia sp. and Trichocereus sp. Disturbed zone with palm trees Disturbed zone with lianas Desert scrub with Prosopis sp. and Puya sp. Desert scrub with rosette-leaved succulent plants of Puya sp. Spiny scrubland with Stetsonia coryne Desert scrubland Desert scrub with Puya sp., Opuntia sp. and Trichocereus sp. Desert scrub with Puya sp., Tunilla sp. and Trichocereus sp. Sand dunes Cultivated area with Prosopis alba and Brea sp. Desert scrub with Puya sp., Gymnocalycium sp. and cultivated area with leguminous plants and lianas Spiny scrubland Spiny scrubland with Trichocereus sp. and other cacti Vegetation 1 Dec. 2006 1 Dec. 2006 1 Dec. 2006 1 Dec. 2006 1 Dec. 2006 1 Dec. 2006 1 Dec. 2006 30 Nov. 2006 30 Nov. 2006 30 Nov. 2006 30 Nov. 2006 30 Nov. 2006 29 Nov. 2006 29 Nov. 2006 29 Nov. 2006 Date 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 69 Biodiversity of Myxomycetes 69 San Juan, Calingasta, Barreal, El Leoncito National Park, 50 km South of Barreal, on route RP-412 San Juan, Calingasta, Barreal, El Leoncito National Park, 25 km South of Barreal, on route RP-412 San Juan, Calingasta, Villa Nueva, on route RP-412 towards Tocota and Iglesia San Juan, Iglesia, Rodeo, on route RN-150, km 380, Paso del Agua Negra, Quebrada Sarmiento San Juan, Iglesia, Rodeo, on route RN-150, km 370, Los Corrales San Juan, Iglesia, Rodeo, on route RN-150, km 366 San Juan, Iglesia, Rodeo, on route RN-150, km 350.5, Ojo de Agua,17 km West of Arrequintín San Juan, Iglesia, Rodeo, on route RN-150, km 326 San Juan, Iglesia, Rodeo, on route RN-150, km 313 San Luis, La Capital, on route RN-7, km 858, Alto Pencoso San Luis, La Capital, San Luis, on route RN-147, km 818, San Jerónimo San Luis, Belgrano, San Antonio, on route RN-147, km 868 San Luis, Belgrano, Hualtarán, Sierra de las Quijadas National Park, viewpoint San Luis, Belgrano, Hualtarán, Sierra de las Quijadas National Park, 2 km East of the viewpoint San Luis, Belgrano, Hualtarán, Sierra de las Quijadas National Park, 3 km East of the viewpoint ARG-07-06 ARG-07-07 ARG-07-08 ARG-07-09 ARG-07-10 ARG-07-11 ARG-07-12 ARG-07-13 ARG-07-14 ARG-07-45 ARG-07-46 ARG-07-47 ARG-07-48 ARG-07-49 ARG-07-50 Locality Table 1. List of collecting localities. (Continuation). 32º29’35”S 66º59’23”W, 777 ± 7 m 32º29’49”S 66º59’44”W, 780 ± 10 m 32º29’47”S 67º00’22”W, 805 ± 9 m 33º14’14”S 66º23’45”W, 663 ± 6 m 33º14’14”S 66º23’45”W, 663 ± 6 m 33º25’07”S 67º04’38”W, 512 ± 6 m 30º22’05”S 69º23’38”W, 2387 ± 5 m 30º23’12”S 69º31’11”W, 2730 ± 8 m 30º21’24”S 69º42’04”W, 3710 ± 10 m Spiny scrubland with Echinopsis sp. Spiny scrubland with Echinopsis sp. Spiny scrubland with Echinopsis sp. Spiny scrubland with Larrea spp. and Prosopis flexuosa Spiny scrubland Spiny scrubland with Prosopis sp. Xerophyllous scrubland Xerophyllous scrubland Xerophyllous scrubland Andean puna Andean puna Andean puna Xerophyllous scrubland Xerophyllous scrubland Xerophyllous scrubland Vegetation 7 Mar. 2007 7 Mar. 2007 7 Mar. 2007 7 Mar. 2007 7 Mar. 2007 6 Mar. 2007 26 Feb. 2007 26 Feb. 2007 26 Feb. 2007 26 Feb. 2007 26 Feb. 2007 26 Feb. 2007 24 Feb. 2007 24 Feb. 2007 24 Feb. 2007 Date 70 30º16’04”S 69º47’43”W, 4100 ± 10 m 30º13’30”S 69º47’33”W, 4305 ± 10 m 30º13’36”S 69º48’26”W, 4615 ± 9 m 31º02’49”S 69º27’50”W, 1668 ± 5 m 31º51’23”S 69º26’03”W, 1867 ± 9 m 31º53’20”S 69º25’08”W, 1892 ± 7 m Coordinates, elevation 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 70 C. Lado & al. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 San Juan, Jáchal, Niquivil, on route RN-40, km 3525, 20 km North of Talacasto San Juan, Jáchal, Niquivil, on route RN-40, km 3572, Tucunuco 30º37’23”S 68º38’21”W, 909 ± 9 m San Juan, Jáchal, San Roque, on route RN-40, km 3619 San Juan, Jáchal, San José de Jáchal, Dique Pachimoco, on route RN-150, km 243 San Juan, Ullum, on route RP-436, 79 km Northwest of the ARG-07-54 ARG-07-55 ARG-07-56 ARG-07-57 ARG-07-58 30º41’41”S 69º01’19”W, 2105 ± 10 m 30º42’41”S 68º58’22”W, 1900 ± 6 m 30º50’58”S 68º57’01”W, 1660 ± 10 m 31º00’46”S 68º45’58”W, 1370 ± 10 m San Juan, Ullum, on route RP-436, 72 km Northwest of the crossroads with route RN-40 and 1 km from the turn to La Invernada San Juan, Ullum, on route RP-436, 66 km Northwest of the crossroads with route RN-40, Minas de Gualilán San Juan, Ullum, La Ciénaga, on route RP-436, 50 km Northwest of the crossroads with route RN-40 San Juan, Ullum, Quebrada de Las Burras, on route RP-436, 18 km Northwest of the crossroads with route RN-40 31º01’41”S 68º45’44”W, 1333 ± 6 m San Juan, Ullum, Termas de Talacasto, on route RP-436, 16.2 km Northwest of from the crossroads with route RN-40 ARG-07-60 ARG-07-61 ARG-07-62 ARG-07-63 30º38’47”S 69º03’56”W, 2445 ± 8 m 30º11’52”S 68º49’23”W, 1240 ± 7 m 30º21’03”S 68º38’07”W, 1054 ± 5 m 31º02’36”S 68º38’09”W, 990 ± 8 m 31º19’14”S 68º36’23”W, 1040 ± 10 m ARG-07-59 crossroads with route RN-40 and 3 km from the turn to Tocota San Juan, Ullum, Matagusanos, on route RN-40, km 3491 ARG-07-53 31º23’07”S 68º35’41”W, 830 ± 10 m San Juan, Albardón, on route RN-40, km 3483 ARG-07-52 31º45’02”S 68º01’56”W, 748 ± 5 m San Juan, Caucete, Vallecito, Difunta Correa, Valle Fértil Natural Park, Sierra Pie de Palo, on route RN-141, km 185 Coordinates, elevation ARG-07-51 Locality Table 1. List of collecting localities. (Continuation). Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 Desert scrub with rosette-leaved succulent plants of Puya sp. Desert scrub with rosette-leaved succulent plants of Puya sp. Spiny scrubland with Eriosyce sp. Spiny scrubland with Tunilla corrugata Spiny scrubland with Tephrocactus sp. Spiny scrubland Spiny scrubland Desert scrub with rosette-leaved succulent plants of Puya sp. Spiny scrubland Spiny scrubland Spiny scrubland with Echinopsis sp., Opuntia sp. and Tephrocactus sp. Spiny scrubland with Tephrocactus sp., Opuntia sp. and Puya sp. Spiny scrubland with Tephrocactus sp. Vegetation 9 Mar. 2007 9 Mar. 2007 9 Mar. 2007 9 Mar. 2007 9 Mar. 2007 9 Mar. 2007 8 Mar. 2007 8 Mar. 2007 8 Mar. 2007 8 Mar. 2007 8 Mar. 2007 8 Mar. 2007 7 Mar. 2007 Date 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 71 Biodiversity of Myxomycetes 71 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 72 C. Lado & al. 72 used in other studies of myxomycetes (Stephenson & al., 1993). To examine community similarity, the Sørensen coefficient of community (CC) index was used, which considers the presence or absence of species in the study areas compared using the formula CC = 2z / (x + y), where z = the number of species in common to both communities, and where x and y equal the number of species in communities A and B, respectively. The completeness of the sampling effort was evaluated using the ACE and CHAO1 abundance indices (Colwell & Coddington, 1994; Colwell & al., 2004). Each collecting site was used as the unit of collecting effort, using the total number of species found with the programme EstimateS v 7.5.2 (http.://viceroy.eeb.uconn.edu/estimates). Results This survey produced a total of 594 collections of myxomycetes, including 372 that had developed under natural conditions in the field, as well as 222 collections obtained from 127 moist chamber cultures. The collections represent 72 different species from 22 genera of myxomycetes. The following list of species includes one species new to science, three species recently described based on material from this survey, 5 more are new records for the Neotropics and a further 11 species previously unknown from South America. The survey has added a total of 38 species to the catalogue of Argentina. Annotated list of species In the list that follows, all the myxomycetes observed are arranged alphabetically by genus and species. Information is provided on the source of each record, first the locality from which the specimen itself or the sample of dead plant material used to prepare the moist chamber culture was collected (Table 1), followed by the substrate upon which it was collected or cultured, and the collection number. A collection obtained from a moist chamber culture is indicated by [mc] followed by the pH of the culture in which the specimen appeared. Additional comments are included for records of particular interest. Nomenclature follows Lado (2005-2010). The abbreviation ‘cf.’ in the name of a taxon indicates that the specimen representing the source of the record could not be identified with certainty. Unless otherwise indicated, the data on Neotropical distribution of myxomycete species is from Lado & Wrigley de Basanta (2008). Arcyria afroalpina Rammeloo. (Figs. 10-21). ARG-06-38: On Puya sp. leaves, MA-Fungi 80230, 80231; on dead leaf base of Puya sp. (mc, pH 7), dwb 2831; (mc pH 7.1), dwb 2854. ARG-06-50: On Puya sp. leaves, MA-Fungi 80232; on dead leaf bases of Puya sp (mc, pH 6.8), dwb 2979. ARG-06-60: On dead leaf bases of Puya sp. (mc, pH 6.9), dwb 2853. ARG-06-61: On dead leaf bases of Puya sp. (mc, pH 7.4), dwb 2973. ARG-0667: On dead leaf bases of Puya sp. (mc, pH 7.2), dwb 2844; (mc, pH 6.8), dwb 2866; (mc, pH 6.9), dwb 3152. ARG-06-68: On dead leaf bases of Puya sp. (mc, pH 7.1), dwb 2849. ARG-07-62: On dead leaf bases of Puya sp. (mc, pH 6.7), dwb 2924. These collections have globose to subglobose sporocarps, 225-500 µm in diameter and 875-1050 µm in total height (Figs. 10-11). The stipe is long and thin (45-67-70 µm wide) and filled with spore-like cysts 8(12.1)-15.5 µm diameter, the peridium is partially evanescent, and the remaining flattened calyculus is slightly narrower than the diameter of the sporotheca and finely warted on the inner surface by LM and SEM (Figs. 12-13). The capillitium is tubular, slightly elastic, firmly attached to the calyculus (Fig. 11), the tubules are 2.5-(3.5)-4.5 µm in diameter, lightly ornamented with warts by LM, the warts fused to form small crests by SEM (Figs. 14, 15, 17, 18). The spores are yellowish by LM, 7.3-(8.3)-9.2 µm diameter, finely warted with groups of more prominent warts readily visible by SEM (Figs. 16, 19-21). In general the specimens agree with the original description of A. afroalpina by Rammeloo (1981a, 1981b) except for the size of the spores, which are 9-11 µm diameter for that species. Rammeloo however comments on the presence of collections (Rammeloo 4061 and Z109) with smaller spores, which he called A. aff. afroalpina but he describes the colour as grey, not yellowish like A. afroalpina and our specimens. In South America only previously reported from Ecuador. New for Argentina. Arcyria cinerea (Bull.) Pers. ARG-06-22: On twigs, MA-Fungi 80406. ARG-06-38: On dead leaf bases of Puya sp. (mc, pH 7), dwb 2859. ARG-06-43: On dead leaf bases of Puya sp. (mc), dwb 2806. ARG-06-51: On dead leaf bases of Puya sp. (mc, pH 6.9), dwb 2868. ARG-06-61: On Puya sp. leaves, MA-Fungi 80233, 80234, 80235. ARG-06-64: On bark of dead liana (mc, pH 7), dwb 2990; (mc, pH 7.1), dwb 3003. ARG-06-73: On dead liana, (mc, pH 7), dwb 2977. ARG-06-76: On Opuntia quimilo bark, (mc, pH 7.4), dwb 2995. ARG-07-48: On dead leaf bases of Puya sp. (mc, pH 6.6), dwb 2907. Arcyria denudata (L.) Wettst. (Figs. 22-27). ARG-06-46: On dead leaves of Cortaderia sp., MA-Fungi 80236, 80237, 80238, 80239, 80240, 80241. ARG-06-47: On dead leaves of Cortaderia sp., MA-Fungi 80404, 80405. The material has small sub-cylindrical sporocarps with the capillitium firmly attached to the calyculus (Fig. 22). The calyculus is plicate with a warted-reticulate ornamentation; the capillitium ornamented with rings, half rings and a reticulum (Figs. 23-25); spores Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 73 Biodiversity of Myxomycetes 73 Figs. 10-11. Sporocarps of Arcyria afroalpina [dwb 2853]: 10, before spore dispersal. 11, after spore dispersal. The capillitial threads firmly attached to the calyculus and barely expanding. globose, 7-8 µm diameter, warted with groups of larger warts (Figs. 26, 27). Arcyria insignis Kalchbr. & Cooke ARG-06-22: On twigs, MA-Fungi 80242. The collection confirms the presence of this species in Argentina (Wrigley de Basanta & al., 2010b). Arcyria minuta Buchet ARG-06-52: On wood, MA-Fungi 80243. Badhamia affinis Rostaf. ARG-06-42: On bark of living Prosopis sp., MA-Fungi 80149, 80244. ARG-07-45: On Prosopis flexuosa bark (mc, pH 5.8), dwb 2901. The field collections were plasmodiocarpic mixed with a few short-stalked sporocarps. The collections all had a delicate capillitium in the form of columns arising from the base of the sporotheca, and spores with a pale band as mentioned by Martin & Alexopoulos (1969). Badhamia foliicola Lister ARG-06-40: On decayed Denmoza rhodocantha, MA-Fungi 80245. Badhamia macrocarpa (Ces.) Rostaf. ARG-06-04: On Opuntia sulphurea cladodes, MA-Fungi 80246. ARG-06-08bis: On Opuntia sp. cladodes, MA-Fungi 80247. ARG-06-27: On twigs, MA-Fungi 80248. ARG-06-52: On wood, MA-Fungi 80249. ARG-06-62: On Opuntia sulphurea, MAFungi 80250. ARG-06-63: On wood, MA-Fungi 80251. ARG-0664: On dried legume fruit, MA-Fungi 80252. Two collections (MA-Fungi 80247, 80248) had physaroid capillitium, but they keyed out to B. macrocarpa. A physaroid capillitium is also mentioned by Martin & Alexopoulos (1969). Badhamia melanospora Speg. ARG-06-01: On Opuntia quimilo cladodes, MA-Fungi 80071, 80072, 80073, 80074, 80075, 80076, 80077, 80078, 80079, 80080. ARG-06-03: On Opuntia sp. cladodes, MA-Fungi 80081, 80082. ARG-06-04: On decayed Echinopsis atacamensis, MA-Fungi 80083; on decayed Gymnocalycium sp., MA-Fungi 80084, 80085; on decayed Puya sp. leaves, MA-Fungi 80089; on decayed Pyrrhocactus sp., MA-Fungi 80086, 80087; on Opuntia sulphurea cladodes, MA-Fungi 80088; on cactus litter on agar, dwb 2815. ARG-0605: On decayed Orocereus trolli, MA-Fungi 80090; (mc, pH 7.4), dwb 2813. ARG-06-06: On Opuntia sp. cladodes, MA-Fungi 80091, 80092, 80093, 80094, 80095, 80096, 80097, 80099, 80100. ARG-06-07: On decayed Echinopsis atacamensis, MA-Fungi 80101, 80102, 80103, 80104, 80105, 80106, 80107, 80108, 80109, 80110; on decayed legume tree wood, MA-Fungi 80113; on deca- Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 74 74 C. Lado & al. Figs. 12-21. Arcyria afroalpina by SEM: 12, 13, detail of the lightly ornamented inner surface of the calyculus. 14, 15, 17, 18, Capillitial threads. 16, 19-21, Spores. [12, 13, 15, 17, 18, 20, 21: dwb 2866. 14, 16, 19: MA-Fungi 80232]. Bar: 12, 13, 17 = 20 µm; 14-16, 18-21 = 10 µm. yed Pyrrhocactus sp., MA-Fungi 80114, 80115, 80116, 80117; on decayed Thillandsia sp. leaves, MA-Fungi 80111, 80112. ARG-0608: On decayed Tephrocactus sp., MA-Fungi 80118; on dead leaf bases of Puya sp. (mc, pH 7.3), dwb 2949. ARG-06-08bis: On decayed Echinopsis atacamensis, MA-Fungi 80119, 80121; on Opuntia sp. cladodes, MA-Fungi 80120. ARG-06-21: On Cumulopuntia boliviana bark (mc, pH 7.5), dwb 2810; (mc, pH 8.1), dwb 2827; (mc, pH 8.1), dwb 2812. ARG-06-24: On decayed Acanthocalycium sp., MA-Fungi 80122. ARG-06-25: On decayed Echinopsis atacamen- sis, MA-Fungi 80123, 80124, 80125. ARG-06-26: On decayed Cereus uruguayensis, MA-Fungi 80129; on decayed Austrocylindropuntia sp., MA-Fungi 80130; on Opuntia sulphurea cladodes, MAFungi 80126, 80127, 80128. ARG-06-28: On decayed Echinopsis atacamensis, MA-Fungi 80131, 80132. ARG-06-30: On decayed Echinopsis atacamensis, MA-Fungi 80133; on Opuntia sulphurea cladodes, MA-Fungi 80134; on Denmoza rhodocantha epidermis (mc, pH 7.6), dwb 3052; (mc, pH 7.8), dwb 3081, dwb 3082. ARG06-32: On decayed Echinopsis atacamensis, MA-Fungi 80136; on Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 75 Biodiversity of Myxomycetes 75 Figs. 22-27. Arcyria denudata by SEM [MA-Fungi 80405]: 22, stalk and remains of the calyculus and capillitial threads. 23-25, capillitial threads. 26, 27, spores. Bar: 22 = 300 µm; 23-25 = 20 µm; 26, 27 = 10 µm. Opuntia sulphurea cladodes, MA-Fungi 80135. ARG-06-34: On decayed Echinopsis atacamensis, MA-Fungi 80137, 80138. ARG-0635: On decayed Echinopsis atacamensis, MA-Fungi 80139; on twigs, MA-Fungi 80140. ARG-06-37: On decayed Echinopsis atacamensis, MA-Fungi 80141; on Opuntia sulphurea cladodes, MA-Fungi 80142. ARG-06-38: On Puya sp. leaves, MA-Fungi 80143, 80144. ARG-06-40: On decayed Austrocylindropuntia sp., MA-Fungi 80145, 80148; on decayed Denmoza rhodocantha, MA-Fungi 80146, 80147. ARG-06-42: On Prosopis sp. bark, (mc, pH 5.1), dwb 3074, dwb 3073. ARG-06-43: On Opuntia sulphurea cladodes, MA-Fungi 80150. ARG-06-44: On decayed Trichocereus thelego- nus, MA-Fungi 80151; on Opuntia sulphurea cladodes, MA-Fungi 80152. ARG-06-45: On decayed Trichocereus sp., MA-Fungi 80153; on Opuntia sulphurea cladodes, MA-Fungi 80154. ARG-0646: On Opuntia sp. cladodes, MA-Fungi 80155. ARG-06-50: On decayed Trichocereus sp., MA-Fungi 80157, 80158, MA-Fungi 80159. ARG-06-52: On Opuntia sp. cladodes, MA-Fungi 80160. ARG-06-54: On decayed Gymnocalycium sp., MA-Fungi 80156. ARG-06-58: On decayed Parodia sp., MA-Fungi 80161. ARG-0659: On decayed Parodia sp., MA-Fungi 80162. ARG-06-62: On decayed Trichocereus sp., MA-Fungi 80163, 80166, 80167; on Opuntia sulphurea cladodes, MA-Fungi 80164, 80165; on Trichocereus Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 76 C. Lado & al. 76 sp. cortex, (mc, pH 7.4), dwb 2984. ARG-06-63: On decayed Trichocereus sp., MA-Fungi 80169, 80170; on Opuntia sulphurea cladodes, MA-Fungi 80168. ARG-06-64: On decayed Gymnocalycium sp., MA-Fungi 80171. ARG-06-68: On Opuntia sulphurea cladodes, MA-Fungi 80172. ARG-06-75: On Tephrocactus sp. remains, MA-Fungi 80173. ARG-06-76: On Opuntia quimilo cladodes, MAFungi 80174, 80175; on Opuntia quimilo bark, (mc, pH 7.8), dwb 2998. ARG-07-08: On decayed Tephrocactus aoracanthus, MAFungi 80176, 80177; on decayed Tephrocactus aoracanthus, (mc, pH 7.7), aet 11925; (mc, pH 8.3), aet 11923; (mc, pH 8.5), aet 11921, aet 11924; (mc, pH 7.4), dwb 2899; (mc, pH 7.3), dwb 2926; (mc, pH 7.2), dwb 2934. ARG-07-11: On twigs, (mc, pH 7.8), aet 11937; on succulent stem of Compositae, (mc, pH 8.0), aet 11942; (mc, pH 8.2), aet 11947. ARG-07-12: On decayed cladodes of Cumulopuntia boliviana, (mc, pH 8.1), aet 11931; on twigs (mc, pH 5.7), aet 11950. ARG-07-13: On decayed Denmoza rhodacantha, MA-Fungi 80178; on Opuntia sp. internal tissue (mc, pH 9.2), aet 11949, aet 11920, aet 11930; (mc, pH 9.0), aet 11919, aet 11926. ARG-07-47: On decayed Echinopsis candicans, MA-Fungi 80181; on Opuntia sulphurea cladodes, MA-Fungi 80179, 80180, 80182; (mc, pH, 8.4), aet 12012, aet 12013. ARG-07-48: On decayed Echinopsis candicans, MA-Fungi 80183. ARG-07-50: On decayed Echinopsis candicans, MA-Fungi 80184; on decayed Tephrocactus articulatus, (mc, pH 8.2), aet 12025. ARG-07-51: On decayed Tephrocactus articulatus, (mc, pH 8.2), aet 12024, aet 12033. ARG-07-52: On decayed Echinopsis strigosa (mc, pH 9.0), aet 12029; on decayed Tephrocactus sp., MA-Fungi 80185, 80186, 80187. ARG-07-53: On decayed Echinopsis sp., MA-Fungi 80188. ARG-07-56: On Puya sp. leaves, MA-Fungi 80189. ARG-07-59: On decayed Eriosyce sp., MA-Fungi 80192, 80193; on decayed Tephrocactus articulatus, MAFungi 80190; on Opuntia sulphurea cladodes, MA-Fungi 80191. ARG-07-60: On decayed Tunilla corrugata, MA-Fungi 80194. ARG-07-61: On decayed Eriosyce sp., MA-Fungi 80195. Calomyxa metallica (Berk.) Nieuwl. ARG-06-71: On Puya sp. leaves, MA-Fungi 80253. Comatricha laxa Rostaf. ARG-06-39: On Prosopis sp. bark (mc, pH 5.5), dwb 2819. Comatricha pulchelloides Nann.-Bremek. ARG-06-38: On dead leaf base of Puya sp. (mc, pH 7), dwb 2867. The sporocarps in this large (>40 sporocarps) moist chamber collection are typical of this species, except for the shorter stalks measuring 1/5 to 1/3 of the total height rather than 1/3 to 1⁄2 as in the description by Nannenga-Bremekamp (1985), and the spores are 9-11 µm diameter, rather than 8-9 µm. A similar species, C. longipila Nann.-Bremek. has shorter stalks, but smaller spores (6-7 µm diameter) and swollen tips of some free ends of the capillitium, a character not present in our specimens. This is the first time this species has been recorded in the Neotropics. Craterium leucocephalum (Pers. ex J.F. Gmel.) Ditmar ARG-06-20: On Cortaderia sp., MA-Fungi 80254, 80255, 80256. ARG-06-24: On grasses, MA-Fungi 80257. ARG-06-50: On Puya sp. leaves, MA-Fungi 80258, 80259, 80260, 80261. ARG06-51: On Puya sp. leaves, MA-Fungi 80262, 80263, 80264. ARG06-52: On wood, MA-Fungi 80265. ARG-06-61: On Puya sp. leaves, MA-Fungi 80266, 80267, 80268, 80269. ARG-06-67: On Puya sp. leaves, MA-Fungi 80270. ARG-06-68: On dead leaf base of Puya sp. (mc, pH 6.9), dwb 2841. ARG-06-71: On dead leaf base of Puya sp. (mc, pH 6.8), dwb 2975. ARG-07-63: On dead leaf base of Puya sp. (mc, pH 6.4), dwb 2895. Some of the specimens (MA-Fungi 80254, 80255, 80256, 80257, dwb 2841, dwb 2975, dwb 2895) were the variety scyphoides (Cooke & Balf. f.) G. Lister, differentiated by the more globose sporotheca and slightly larger spores. Cribaria lepida Meyl. (Figs. 28-31). ARG-07-56: On Puya sp. leaves, MA-Fungi 80271; on dead leaf base of Puya sp. (mc, pH 6.9), dwb 2886; (fc) dwb 2896. ARG-0661: On dead leaf base of Puya sp. (mc, pH 7.4), dwb 3015. ARG07-63: On Puya sp. leaves, MA-Fungi 80272. This purple Cribraria, is similar in colour to C. violacea but with longer stalks, up to 8 times the diameter of the sporotheca. The specimens have a shallow calyculus occupying approximately one third of the diameter of the sporotheca (Fig. 28), finely dotted with calcic granules (Fig. 28), the upper margin scalloped giving rise at the points to a fine net with few thickened pulvinate nodes. The spores are pale violet, 6-7.5 µm diameter and have a smooth but pitted appearance in transmitted light with Nomarski optics, but are in fact densely and minutely warted by SEM (Figs. 29-31). The plasmodium was described as probably white by Meylan (1927) but he may have seen later stages of the morphogenesis. Pale purple protoplasmodia were observed during the development of these specimens in moist chamber culture, giving rise to a pale stalk with the white ball of the forming sporotheca on top, dotted with purple as the nodes form, also mentioned by Meylan, then all turning completely purple and exuding water droplets with the maturation of the spores. These suppose new records of the species for South America. It has been cited from dry areas of Mexico (Estrada-Torres & al., 2009). Cribraria violacea Rex ARG-06-27: On twigs, MA-Fungi 80273. Dianema corticatum Lister Comatricha tenerrima Nann.-Bremek. ARG-06-27: On twigs, MA-Fungi 80274. ARG-06-64: On bark of dead liana (mc, pH 7.1), dwb 2986; (mc, pH 7), dwb 2988. In this collection the specimens were predominate- Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 77 Biodiversity of Myxomycetes ly sporocarpic to short plasmodiocarps, and the sparse capillitium has branched slender threads, often twisted spirally at the ends and with a roughened surface, and typical clustered spores but with 5-15 spores per cluster, not 2-6 as stated in Martin and Alexopolous (1969). This is the first record for South America. In the Neotropics it has been reported from Mexico. 77 ceum in the spores, which are subglobose, 13-16 µm diameter, densely and minutely warted, with a pattern of tiny white lines where there are fewer warts, the spores appearing almost angular with Nomarski optics because of the lines, whereas in D. crustaceum the spores are dark and spiny, mostly 12-14 µm diameter. In the Neotropics, this species has been reported from Mexico. Dictydiaethalium plumbeum (Schumach.) Rostaf. ARG-06-27: On twigs, MA-Fungi 80275. Diderma cf. deplanatum Fr. ARG-06-51: On leaves of Puya sp., MA-Fungi 80276. Diderma cf. crustaceum Peck ARG-06-50: On dead leaf bases of Puya sp. (mc, pH 6.8), dwb 3023. The specimens in this collection are macroscopically similar to D. crustaceum, with white sessile sporocarps, which are closely gregarious to heaped, and distorted in shape by mutual pressure. They differ from D. crustaceum in having a brown and membranous hypothallus, not white and limy as in that species. Our specimen also differs from D. crusta- The weathered material in this collection did not permit a definite identification. Diderma hemisphaericum (Bull.) Hornem. ARG-06-27: On twigs, MA-Fungi 80277, 80278, 80279, 80280. Didymium anellus Morgan ARG-06-03: On decayed Echinopsis atacamensis cf., MA-Fungi 80281. ARG-06-38: On dead leaf base of Puya sp. (mc, pH 7.1), dwb 2855; (mc, pH 7.1), dwb 2870; (mc, pH 7), dwb 2856. Figs. 28-31. Cribraria lepida by SEM [dwb 2886]: 28, remains of the peridium forming a shallow calyculus. 29-31, spores. Bar: 28 = 100 µm; 29-31 = 10 µm. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 78 78 C. Lado & al. The field collection differs from the typical D. anellus in having spores without groups of warts. Didymium clavus (Alb. & Schwein.) Rabenh. Didymium obducens P. Karst. ARG-06-08: On dead leaf bases of Puya sp. (mc, pH 7.3), dwb 2940. ARG-06-76: On litter, MA-Fungi 80098; on wood, MA-Fungi 80282, MA-Fungi 80283. Our specimen fits the description of this species by Härkönen (1979: 3-5) exactly. It is the first record for the Neotropics. Didymium dubium Rostaf. Didymium quitense (Pat.) Torrend. (Figs. 32-36). ARG-06-03: On decayed Echinopsis sp., MA-Fungi 80284. ARG-06-64: On the wood of a leguminous tree, MA-Fungi 80285. Didymium infundibuliforme D.Wrigley, Lado & Estrada ARG-06-08: On dead leaf bases of Puya sp., dwb 2942. ARG06-50: On Puya sp. leaves, MA-Fungi 78321, 80286; on dead leaf bases of Puya sp. (mc, pH 6.8), dwb 3019. ARG-06-51: On dead leaf base of Puya sp. (mc, pH 6.8), dwb 2829, dwb 2834. ARG-0660: On dead leaf base of Puya sp. (mc, pH 7.1), dwb 2851. ARG06-61: On Puya sp. leaves, MA-Fungi 78322, 78323, 78320. ARG06-67: On dead leaf base of Puya sp. (mc, pH 6.9), dwb 2825; (mc, pH 7.2), dwb 2843; (mc, pH 6.8), dwb 2845; (mc, pH 6.9), dwb 3154. ARG-07-50: On dead leaf base of Puya sp. (mc, pH 6.9), dwb 2918. ARG-07-52: On dead leaf base of Puya sp. (mc, pH 7.1), dwb 2927. ARG-07-56: On Puya sp. leaves, MA-Fungi 78324, 78325. ARG-07- 63: On dead leaf base of Puya sp. (mc, pH 6.4), dwb 2894. These collections with the exception of MA-Fungi 80286, dwb 2918 and dwb 3154, were included in the original description of the species by Wrigley de Basanta & al. (2009). Didymium cf. listeri Massee ARG-06-27: On twigs, MA-Fungi 80288. This collection has white sporocarps to short plasmodiocarps, with a double peridium, the outer layer like an egg-shell, formed of tiny closely packed crystals (Fig. 32), the inner layer membranous and iridescent. The dehiscence is irregular, leaving a flat base attached to the substrate. The capillitium is short and rigid, brown with pale ends, of uniform diameter and with cross connections forming a loose net, with a granular surface by SEM (Fig. 34). The spores are dark purplish brown and polyhedral with the angles slightly lighter in colour, 13-15 µm diameter, covered with warts united to form a sub-reticulum by LM, much more clearly seen by SEM. In the SEM micrographs (Figs. 33, 35, 36) the ornamentation can be seen to be a dense network of irregular muri. This rare species was described by Patouillard & Lagerheim (1895) as Chondrioderma quitense Pat. from Quito (Ecuador), on leaves. This is the first time the species has been recorded from Argentina. Didymium squamulosum (Alb. & Schwein.) Fr. ARG-07-11: On decayed wood, (mc, pH 7.0), aet 11956. This collection has many sessile, pulvinate sporocarps to short effuse plasmodiocarps, with irregular dehiscence. The peridium is double, the outer layer like an egg-shell, formed of closely packed crystals. The inner layer is membranous, iridescent and hyaline by LM. The capillitium is rigid, formed by hyaline tubules arising from the rudimentary columella. The spores are globose, 10-11.5 µm diameter, faintly warted and with a paler area. This species is macroscopically similar to D. quitense (see below) but has smaller, paler globose spores, and a hyaline capillitium. It differs from the original description in the pale area of the spores and the paler capillitium. If confirmed this would be the first record of the species for Argentina. In the Neotropics it has been found in Mexico and Ecuador. ARG-06-27: On twigs, MA-Fungi 80289, 80290. ARG-06-64: On legume dried fruits, MA-Fungi 80291, 80292; on legume tree wood, MA-Fungi 80293, 80294, 80295; on Puya sp. leaves, MAFungi 80296. ARG-06-70: On decayed Stetsonia coryne, MA-Fungi 80297, 80298; on legume tree wood, MA-Fungi 80299. ARG06-76: On Prosopis sp. litter, MA-Fungi 80300; on Opuntia quimilo bark (mc, pH 6.9), dwb 2993. Some of the collections had sub-sessile sporocarps, and the stipes of some stipitate sporocarps were less robust. Some also did not have an obvious white hypothallus. Didymium vaccinum (Durieu & Mont.) Buchet ARG-06-07: On decayed Echinopsis atacamensis, MA-Fungi 80287. ARG-06-24: On Cumulopuntia boliviana epidermis (mc, pH 8), dwb 3076; (mc, pH 7.5), dwb 3077. ARG-06-38: On Puya sp. leaves, MA-Fungi 80301, 80302, 80303, 80304; on dead leaf base of Puya sp. (mc, pH 7), dwb 2832. ARG-06- 50: On Puya sp. leaves, MA-Fungi 80305, 80306, 80307, 80308; on dead leaf bases of Puya sp. (mc, pH 7.1), dwb 2978. ARG-06-57: On Stipa atacamensis, MA-Fungi 80309. ARG-06-60: On Puya sp. leaves, MA-Fungi 80310. ARG-07-56: On Puya sp. leaves, MA-Fungi 80311, 80312, 80313. ARG-06-64: On bark of dead liana (mc, pH 7), dwb 3020. ARG-06- 67: On dead leaf base of Puya sp. (mc, pH 6.9), dwb 3153. This is the first record of this species for South America. In the Neotropics it has been found in Mexico. These represent the first records of this species for Argentina. Didymium mexicanum G. Moreno, Lizárraga & Illana Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 79 Biodiversity of Myxomycetes 79 Figs. 32-36. Didymium quitense by SEM [MA-Fungi 80288]: 32, detail of the peridium with the outer layer formed of tiny closely packed crystals. 33, 35, 36, spores with a dense network of irregular muri. 34, fragment of a capillitial thread. Figs. 37-39. Physarum synsporum by SEM [MA-Fungi 80377]: 37, spore with irregular ornamentation. 38, 39, cluster of spores, arrows show the contact points between the spores. Bar: 32, 34 = 20 µm; 33, 35-39 = 10 µm. Didymium wildpretii Mosquera, Estrada, BeltránTej., D. Wrigley & Lado ARG-06-76: On Opuntia quimilo bark (mc, pH 7.4), dwb 2994; (mc, pH 7.8), dwb 2996; (mc, pH 6.9), dwb 3005. ARG-06-03: On Opuntia sp. cladodes, MA-Fungi 80208. These are the first records for South America of this Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 80 80 C. Lado & al. species. It was recently described from material from arid areas of Mexico and from the Canary Islands, Spain (Lado & al., 2007b). Echinostelium arboreum H.W. Keller & T.E. Brooks. (Fig. 40). ARG-06-08: On dead leaf bases of Puya sp. (mc, pH 7.3), dwb 2937; (mc, pH 6.9), dwb 2939. ARG-06-30: On Denmoza rhodocantha epidermis (mc, pH 7.5), dwb 3078. ARG-07-08: On Tephrocactus aoracanthus remains (mc, pH 8.3), aet 11918. Described from Mexico this species is characterized by pale yellow sporocarps with a persistent shiny peridium (Fig. 40), which dehisces at the base of the sporotheca leaving a distinct wide collar. The columella is stout and it has dichotomously branched capillitial threads. This is the first record of the species from Argentina. Echinostelium colliculosum K.D. Whitney & H.W. Keller ARG-06-08: On dead leaf bases of Puya sp. (mc, pH 6.9), dwb 2938. ARG-06-20: On bark and twigs of unidentified shrub (mc, pH 4.4), dwb 2786; (mc, pH 4.5), dwb 2801 ARG-06-30: On Denmoza rhodocantha epidermis, (mc, pH 7.5), dwb 3042; (mc, pH 7.8), dwb 3043. ARG-06-36: On Prosopis sp. bark (mc, pH 6.1), dwb 2787 ARG-06-39: On Prosopis sp. bark (mc, pH 6.1), dwb 3039; (mc, pH 6), dwb 3040 ARG-06-51: On dead leaf bases of Puya sp. (mc, pH 6.9), dwb 2823; (mc, pH 6.8), dwb 2824 ARG06-52: On Brea sp. bark (mc, pH 6.4), dwb 2963. ARG-07-08: On Tephrocactus aoracanthus remains (mc, pH 7.3), dwb 2887; (mc, pH 8.5), aet 11913; (mc, pH 8.3), aet 11915. ARG-07-50: On Opuntia sulphurea remains (mc, pH 8.8), aet 12010; on Tephrocactus articulatus remains (mc, pH 8.4), aet 12009. This species was reported from arid areas of Mexico (Estrada-Torres & al., 2009) from cacti and tree bark. The authors commented on the small size and the slightly different spore-like body and spore shape. These collections appear to be the same ecotype, and also measure from 60-130 µm in total height with a sporotheca of 20-29 µm diameter. Whitney (1980), in the original description, gives the measurements as 70-150 µm, and 30-50 µm respectively. It was reported for the first time from Argentina from Santa Cruz (Wrigley de Basanta & al., 2010b). Echinostelium minutum de Bary ARG-06-38: On dead leaf bases of Puya sp. (mc, pH 7), dwb 2863. ARG-06-42: On Prosopis sp. bark (mc, pH 5.6), dwb 3045. Hemitrichia minor G. Lister ARG-06-01: On Opuntia quimilo cladodes, MA-Fungi 80196, 80197. Fig. 40. Echinostelium arboreum [dwb 2937]: Two sporocarps showing the persistent shiny peridium. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 81 Biodiversity of Myxomycetes Licea belmontiana Nann.-Bremek. ARG-07-63: On Puya sp. leaves, MA-Fungi 80314. ARG-0748: Dead leaf base of Puya sp. (mc, pH 6.6), dwb 2916. 81 The species is cited here for the first time for the Neotropics. These are the first records of this species in South America. In the Neotropics it has been cited from Mexico. Licea scyphoides T.E. Brooks & H.W. Keller. (Figs. 49-53). Licea denudescens H.W. Keller & T.E. Brooks The circumscissile equatorial dehiscence leaving a basal calyculus (Figs. 49, 50), the clearly ornamented inner peridial surface (Fig. 51) and the very closely punctate spores (Figs. 52, 53) are characteristics of this species. Reported previously from Ecuador and Peru in South America, and on the bark of another living cactus, Opuntia sp. from Mexico (Wrigley de Basanta & Lado, 2005) but these are the first records from Argentina. ARG-07-45: On Prosopis flexuosa bark (mc, pH 6), dwb 2902, dwb 2905. In South America the species has been recorded from Brazil, but these represent the first records for Argentina. Licea eremophila D. Wrigley, Lado & Estrada ARG-06-60: On Puya sp. leaves, MA-Fungi 79159; on dead leaf bases of Puya sp. (mc, pH 7.1), dwb 2837; (mc, pH 6.9), dwb 2888. ARG-06-61: On dead leaf bases of Puya sp. (mc, pH 7.4), dwb 2982. ARG-06-62: On Trichocereus sp. cortex (mc, pH 7.4), dwb 3002. ARG-06-67: On Puya sp. leaves, MA-Fungi 79160, 79161; on dead leaf bases of Puya sp. (mc, pH 6.9), dwb 2826, dwb 3151; (mc, pH 7.2), dwb 2885. ARG-07-62: On dead leaf bases of Puya sp. (mc, pH 6.7), dwb 2920. These collections were included in the paper describing this as a new species (Wrigley de Basanta & al., 2010a). Licea pygmaea (Meyl.) Ing. (Figs. 41-48). ARG-06-06: On bark of living Prosopis sp. (mc, pH 6.2), dwb 3035. ARG-06-10: On Stipa atacamensis (mc, pH 7.1), dwb 2874; (mc, pH 7), dwb 2875. ARG-06-14: On Stipa atacamensis (mc, pH 6.5), dwb 3006. ARG-06-22: On Stipa sp. (mc, pH 5.5), dwb 3026; (mc, pH 4.8), dwb 3037. ARG-06-24: On Stipa atacamensis (mc, pH 5.5), dwb 2999. These specimens belong to the subgenus Licea, according to the revision of the genus by Nannenga-Bremekamp (1965), dehiscing by platelets (Figs. 41, 42). They have baculate spore ornamentation by SEM (Figs. 46-48) and teeth-like protuberances on the edge of the platelets (Figs. 43-45). The double peridium is slightly warted on edge of the inner surface at high magnification by SEM (Figs. 43, 44), otherwise smooth. These are the first records of this species in South America. In the Neotropics it has been cited from Mexico. ARG-06-02: On bark of living Echinopsis atacamensis (mc, pH 6.5), dwb 2778. Licea succulenticola Mosquera, Lado, Estrada & Beltrán-Tej. ARG-06-50: On dead leaf bases of Puya sp. (mc, pH 6.8), dwb 3008. ARG-06-60: On dead leaf bases of Puya sp. (mc, pH 7.1), dwb 2871. ARG-07-50: On internal tissue of Opuntia sulphurea (mc, pH 8.8), aet 12021. ARG-07-56: On Puya sp. leaves, MAFungi 80343; on dead leaf bases of Puya sp. (mc, pH 6.9), dwb 2921. These represent the first records of the species for Argentina. It was recently described from material from arid areas of Mexico, from the Canary Islands, Spain and from USA (Mosquera & al., 2003). In South America it is also known from Chile and Ecuador. Lycogala epidendrum (L.) Fr. ARG-06-27: On wood, MA-Fungi 80315. Macbrideola andina D. Wrigley, Lado & Estrada, sp. nov. (Figs. 54-58). Holotype: Argentina. Salta, Molinos, Molinos river, route RN-40, km 4458, 25°25’56”S 66°17’11”W, 2059 m ± 10m, on bark of living Prosopis sp. collected 25-XI-2006, A. Estrada-Torres, C. Lado & D. Wrigley de Basanta, dwb 3048 (MA-Fungi 79883). ARG-06-06: On bark of living Prosopis sp. (mc, pH 6.2), dwb 3036, 3026a. Species Macbrideola oblonga Pando & Lado proxima, sed ab ea densis capillitium atque uniforme diametros, cum acuminis spinosis, sporothecae sine torquis basalis, sporis densis spinulis ornatis primo icto discernibilis. The characteristic hyaline tubercles on the platelet edges of the transparent peridium permit the identification of this tiny species, described by Mitchell & McHugh (2000), from the British Isles and USA. Sporocarps scattered or in small groups, stipitate, 0.4-0.9 mm in total height. Sporotheca 0.25-0.6 mm in height by 0.2-0.4 mm wide, ellipsoidal (Figs. 54, 57), rarely subglobose, greyish brown (61. gy. Br-62. d. gy. Licea sambucina D.W. Mitch. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 82 82 C. Lado & al. Figs. 41-48. Licea pygmaea by SEM: 41, 42, sporocarp showing the dehiscence by plates. 43-45, edge of the peridial platelets showing teeth-like protuberances. 46-48, spores. [41, 47: dwb 3037. 42-46, 48: dwb 2999]. Bar: 41, 42 = 100 µm; 43, 44 = 20 µm; 45-48 = 10 µm. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 83 Biodiversity of Myxomycetes 83 Figs. 49-53. Licea scyphoides by SEM [dwb 2778]: 49, 50, dehisced sporocarps. 51, detail of the ornamentation of the inner peridial surface. 52, 53, spores with a dense ornamentation of minute warts. Bar: 49, 50 = 100 µm; 51-53 = 10 µm. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 84 84 C. Lado & al. Br). Hypothallus membranous, translucent, concolourous with the base of the stalk. Stalk 0.1-0.3 mm in length, hollow, tubular, widening into the hypothallus, brownish black (65. br Black), with a paler area, orange-yellow to yellowish brown (72. d. OY-75. deep y Br) at the base. Peridium membranous, completely evanescent. Columella brownish black (65. br Black), almost reaching the apex of the sporotheca. Capillitium greyish brown (61. gy. Br-62. d. gy. Br), arising from all parts of the columella, dichotomously branched, forming a dense internal net, the threads of uniform diameter, not tapering towards the periphery, making frequent anastomoses with numerous free ends appearing as spines at the periphery, at the base of the sporotheca these anastomoses making an irregular surface net (Figs. 55, 58). Spores greyish brown (61. gy. Br-62. d. gy. Br) in mass, lighter by transmitted light (63. l. br Gy), globose, 9-11 µm diam., warted by LM, with closely, evenly distributed spinules by SEM (Fig. 56). Plasmodium not observed. Etymology: The epithet andina refers to the geographical area where the species was found. Habitat: Bark of living Prosopis sp. Known distribution: Salta and Jujuy, Argentina. Other specimens examined ARG-06-06: On bark of living Prosopis sp. (mc, pH 6.0), dwb 3055. ARG-06-36: On bark of living Prosopis sp. (mc, pH 6.1), dwb 2795. ARG-06-39: On bark of living Prosopis sp. (mc, pH 6), dwb 3041; (mc, pH 6.1), dwb 3065. Macbrideola oblonga Pando & Lado. Spain: Soria, Calatañazor, Dehesa de Carrillo, 1050 m, 30TWM1417, on bark of Juniperus thurifera (mc), MA-Fungi 16008 (Holotype). The distinctive characters of this new species are its ellipsoid sporotheca (Fig. 54), a short tubular stalk, a columella gradually tapering upwards to the apex, a dense, robust, capillitium with threads of uniform diameter, and no collar (Figs. 54, 55, 57, 58). This species also has a distinctive ornamentation of the spores by SEM (Fig. 56). The overall habit is most similar to Macbrideola oblonga from which the new species differs in its denser capillitium, with spiny free ends (Figs. 55, 58), and its cylindrical stalk (Figs. 54, 57). In M. oblonga the capillitium was described as “… hardly or not anastomosing inside ... free ends blunt slightly swollen or club-shaped” (Pando & Lado, 1988) and in M. oblonga has 4-6 meshes per radius of the sporotheca, whereas in M. andina there are 10-12 meshes in a much denser net. The stalk in M. oblonga is conical (Pando & Lado, 1988: fig. 2). The new species also differs in its total lack of a collar (transparent red-brown in M. oblonga) and the ornamentation of the spores by SEM. This ornamentation, in M. oblonga, is of small rounded warts, not spinules, and they are less uniformly distributed than in M. andina. Another species that lacks a collar and has anastomoses on the periphery of the capillitium is Macbrideola dubia Nann.-Bremek. & Y.Yamam. (NannengaBremekamp & Yamamoto, 1990), but in this species the columella only reaches around the middle of the sporotheca, not almost to the apex as in our species, the lax capillitium attenuates towards the periphery, and it has a netted fibrous stalk base, absent in the new species. Macbrideola dubia also has an extensive hypothallus common to groups of sporocarps, and darker spores. The shape of M. andina is somewhat similar to M. ovoidea Nann.-Bremek. & Y. Yamam. (Nannenga-Bremekamp & Yamamoto, 1983) but in this species the capillitium does not anastomose, which easily distinguishes it from the new species. Macbrideola ovoidea also has smaller spores, 7-8.5 µm diameter vs. 9-11 µm diameter in M. andina, and the spiny ornamentation by SEM has stellate apices (Moreno & al., 2006: figs. 29, 30), absent in M. andina. Another species with a dense capillitium is Macbrideola lamprodermoides G. Moreno, Lizárraga, & Illana (Moreno & al., 2006) but it can be easily distinguished from M. andina by its persistent silvery peridium, evanescent in the latter, and the presence of a collar, absent in M. andina. Macbrideola herrerae Lizárraga, G. Moreno & Illana (Lizárraga & al., 2006) has a well-developed capillitium but it does not anastomose at the periphery and is made of rigid and parallel threads with free dichotomously branched ends, totally different from the dense and robust net of the capillitium of M. andina. Macbrideola reticulospora Hooff & Nann.-Bremek. is distinguished by its reticulate spores. The specimens developed on the bark of living Prosopis sp. at over 2000 m elevation at a slightly acidic pH, in two different provinces (Jujuy and Salta) of the North of Argentina. The development time in moist chamber was 4-71 days, probably dependent on whether the species was in the form of sclerotia, microcysts or spores, when the substrate was wetted. Macbrideola scintillans H.C. Gilbert ARG-06-73: On dead liana (mc, pH 7), dwb 2966. This represents the first record of the species for South America. Perichaena calongei Lado, D. Wrigley & Estrada ARG-06-08: On dead leaf base of Puya sp. (mc, pH 6.9), dwb 2957. ARG-06-38: On Puya sp. leaves, MA-Fungi 78678, 78679, 78680, 78681; on dead leaf base of Puya sp. (mc, pH 7), dwb 2833; (mc, pH 7.1), dwb 2865; (mc, pH 7), dwb 2857. ARG-06-50: On Puya sp. leaves, MA-Fungi 78682, 78683, 78684. ARG-06-51: On Puya sp. leaves, MA-Fungi 78685, 78686, 78687, 78688. ARG-0660: On dead leaf base of Puya sp. (mc, pH 6.9), dwb 2838; (mc, pH Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 85 Biodiversity of Myxomycetes 85 Figs. 54-58. Macbrideola andina by SEM [dwb 3048]: 54, 57, sporocarps. 55, detail of the base of the sporotheca with the capillitium attached along the length of the columella. 56, spores. 58, detail of the base of another sporotheca showing anastomoses and spiny free ends. Bar: 54 = 300 µm; 55, 57, 58 = 100 µm; 56 = 10 µm. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 86 86 C. Lado & al. 7.1), dwb 2852. ARG-06-61: On Puya sp. leaves, MA-Fungi 78689; on dead leaf base of Puya sp. (mc, pH 7.4), dwb 3009. ARG-06-67: On Puya sp. leaves, MA-Fungi 78690. ARG-06-68: On dead leaf base of Puya sp. (mc, pH 7.1), dwb 2850; (mc pH 6.9), dwb 2873. ARG-07-56: On Puya sp. leaves, MA-Fungi 78691, 78692, 78693. ARG-07-63: On Puya sp. leaves, MA-Fungi 78694. These collections were included in the paper describing this as a new species (Lado & al., 2009). Perichaena depressa Lib. ARG-06-38: On Puya sp. leaves, MA-Fungi 80316, 80317, 80318, 80319, 80320. ARG-06-50: On Puya sp. leaves, MA-Fungi 80321. ARG-06-51: On Puya sp. leaves, MA-Fungi 80322. ARG06-60: On Puya sp. leaves, MA-Fungi 80323. ARG-06-70: On decayed Stetsonia coryne, MA-Fungi 80324. ARG-07-56: On Puya sp. leaves, MA-Fungi 80325, 80326. Perichaena quadrata T. Macbr. ARG-06-06: On bark of living Prosopis sp., (mc, pH 6), dwb 3058. ARG-06-08: On dead leaf base of Puya sp. (mc, pH 7.2), dwb 2955; (mc, pH 6.9), dwb 2956. ARG-06-27: On twigs, MAFungi 80327. ARG-06-38: On Puya sp. leaves, MA-Fungi 80328; on dead leaf base of Puya sp. (mc, pH 7.1), dwb 2864; (mc, pH 7), dwb 2858, dwb 2860. ARG-06-39: On Prosopis sp. bark (mc, pH 5.5), dwb 3094. ARG-06-42: On dead leaf base of Puya sp. (mc, pH 7.2), dwb 3091, ARG-06-61: On dead leaf base of Puya sp. (mc, pH 7.4), dwb 2981. ARG-06-64: On bark of dead liana (mc, pH 7), dwb 3021, (mc, pH 7.1), dwb 3010. These represent the first records of the species for South America. In the Neotropics, it has been reported from Mexico. for the Neotropics, if confirmed. The species was described by Martin (1948), from the mountains of California. Physarum bitectum G. Lister ARG-06-06: On bark of Prosopis sp. (mc, pH 5.9), dwb 3038. ARG-06-27: On twigs, MA-Fungi 80344, 80345, 80346, 80347, 80348. ARG-06-38: On Puya sp. leaves, MA-Fungi 80349, 80350 80351, MA-Fungi 80352. ARG-06-39: On bark of Prosopis sp. (mc, pH 5.5), dwb 3071. ARG-06-42: On bark of Prosopis sp. (mc, pH 5.8), dwb 3068. These collections are new records of the species for Argentina. Physarum compressum Alb. & Schwein. ARG-06-01: On Opuntia quimilo cladodes, MA-Fungi 80198, 80199, 80200. ARG-06-03: On decayed Echinopsis sp., MA-Fungi 80203, 80204, 80205; on Opuntia sp. cladodes, MA-Fungi 80201, 80202, 80206. ARG-06-08: Isolated on agar from remains of a bromeliad, dwb 2814. ARG-06-38: On dead leaf base of Puya sp. (mc, pH 7), dwb 2861. ARG-06-50: On Puya sp. leaves, MA-Fungi 80207. ARG-06-76: On Opuntia quimilo bark (mc, pH 6.9), dwb 2992; (mc, pH 7.8), dwb 2997. Physarum decipiens M.A. Curtis ARG-06-39: On bark of living Prosopis sp. (mc, pH 6), dwb 3070; (mc, pH 5.5), dwb 3072. These collections are new records of the species for Argentina. Physarum didermoides (Pers.) Rostaf. Perichaena vermicularis (Schwein.) Rostaf. ARG-06-05: On Oreocereus trollii remains (mc, pH 6.8), dwb 2811. ARG-06-07: On decayed Echinopsis atacamensis, MA-Fungi 80329, 80330, 80331, 80332; on decayed legume tree wood, MAFungi 80333. ARG-06-24: On decayed Acanthocalycium sp., MAFungi 80334. ARG-06-38: On Puya sp. leaves, MA-Fungi 80335. ARG-06-50: On Puya sp. leaves, MA-Fungi 80336, 80337, 80338. ARG-06-51: On Puya sp. leaves, MA-Fungi 80339, 80340. ARG-0652: On Brea sp. bark (mc, pH 6.5), dwb 3013. ARG-06-62: On decayed Trichocereus sp., MA-Fungi 80341; on Trichocereus sp. cortex (mc, pH 7.4), dwb 2985; (mc, pH 7.2), dwb 3017. ARG-06-63: On decayed Trichocereus sp., MA-Fungi 80342. ARG-06-71: On dead leaf bases of Puya sp. (mc, pH 6.8), dwb 2983. ARG-07-08: On Tephrocactus aoracanthus remains (mc, pH 8.3), aet 11960; (mc, pH 8.5), aet 11924b. ARG-07-47: On Opuntia sulphurea remains (mc, pH 8.2), aet 12030; (mc, pH 8.4), aet 12031. ARG-07-50: On Tephrocactus articulatus (mc, pH 8.2), aet 12026; (mc, pH 8.4), aet 12027. ARG-07-51: On Tephrocactus articulatus (mc, pH 8.2), aet 12035. Physarum cf. auripigmentum G.W. Martin ARG-07-56: On Puya sp. leaves, MA-Fungi 80407. The specimen is close to P. auripigmentum but the spores are slightly larger [10-12 µm diameter vs. (8)910(11) µm diameter] and have a paler area at one pole. This would represent the first record of the species ARG-06-03: On decayed Echinopsis sp., MA-Fungi 80353; on Opuntia sp. cladodes, MA-Fungi 80401. Physarum hongkongense Chao H. Chung ARG-06-38: On Puya sp. leaves, MA-Fungi 80354. The laterally compressed, pale yellow, sessile plasmodiocarps with a double peridum, which separates at dehiscence, are typical of Physarum hongkongense. However the spores in our collection are larger (10.512 µm diameter vs. 7.5-9 µm diameter). This species can be distinguished from P. bogoriense primarily on the basis of the yellow colour of the plasmodiocarps (Chung & Tzean, 1998), but the authors have found the two species occurring together on a single leaf. We have also seen the same in material from Brazil (unpublished data). Physarum hongkongense, apart from its colour, differs in the apical fissure of dehiscence, which is by irregular fracture into fragments in P. bogoriense. As has been stated before (Wrigley de Basanta & al., 2008), these species may be conspecific. This collection represents the first record of the species for South America. In the Neotropics, it has been reported from Mexico. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 87 Biodiversity of Myxomycetes Physarum leucophaeum Fr. ARG-06-27: On twigs, MA-Fungi 80355. Physarum licheniforme (Schwein.) Lado ARG-06-16: On Stipa atacamensis, MA-Fungi 80356, 80402, 80357; (mc, pH 7), dwb 2794; (mc, pH 7.1), dwb 2800. ARG-0620: On Cortaderia sp., MA-Fungi 80358. ARG-06-24: On grasses, MA-Fungi 80359. ARG-06-50: On dead leaf bases of Puya sp. (mc, pH 6.8), dwb 2980. ARG-07-12: On twigs, MA-Fungi 80360. These collections represent the first records of the species for South America. In the Neotropics, it has been reported from Mexico and Cuba. 87 2796. ARG-07-45: On bark of living Prosopis flexuosa (mc, pH 6), dwb 2930, dwb 2910. Physarum spectabile Nann.-Bremek., Lado & G. Moreno ARG-06-03: On decayed Echinopsis sp., MA-Fungi 80373. ARG-06-07: On Echinopsis atacamensis internal tissue (mc, pH 8.9), dwb 2798; (mc, pH 8.6), dwb 2799; (mc, pH 9.1), dwb 2809. ARG-06-30: On decayed Echinopsis atacamensis, MA-Fungi 80374. ARG-06-35: On twigs, MA-Fungi 80375. ARG-06-62: On decayed Trichocereus sp., MA-Fungi 80376. ARG-06-70: On Stetsonia coryne, (mc, pH 7.4), dwb 3007. ARG-07-11: On succulent stem of Compositae, (mc, pH 8.0), aet 11946; on twigs of an unidentified shrub (mc, pH 7.8), aet 11953. ARG-07-12: On decayed Cumulopuntia boliviana, (mc, pH 8.1), aet 11959. Physarum megalosporum T. Macbr. ARG-06-63: On wood, MA-Fungi 80361. ARG-06-70: On wood of a leguminous plant, MA-Fungi 80362. ARG-06-76: On wood, MA-Fungi 80363. These collections are the first records of this species for Argentina. Physarum notabile T. Macbr. ARG-06-02: On Echinopsis atacamensis remains (mc, pH 6.5), dwb 2802. ARG-06-61: On leaves of Puya sp., MA-Fungi 80364, 80365, 80366. ARG-06-62: On Trichocereus sp., MA-Fungi 80367 ARG-06-64: On a legume tree wood, MA-Fungi 80368, 80369, 80370. ARG-06-67: On dead leaf base of Puya sp. (mc, pH 6.5), dwb 2842; on leaves of Puya sp., MA-Fungi 80371. ARG-06-68: On dead leaf base of Puya sp. (mc, pH 6.8), dwb 2872. ARG-0676: On wood, MA-Fungi 80372. ARG-07-12: On twigs of a Compositae, (mc, pH 5.8), aet 11944. Physarum pusillum (Berk. & M.A. Curtis) G. Lister ARG-06-03: On decayed Trichocereus thelegonus, MA-Fungi 80209. ARG-06-38: On Puya sp. leaves, MA-Fungi 80210; on dead leaf base of Puya sp. (mc, pH 7), dwb 2830. ARG-06-42: On Prosopis sp. wood, MA-Fungi 80212, 80213; on twigs, MA-Fungi 80211; on Prosopis sp. bark (mc, pH 6.5), dwb 3075. ARG-06-43: On dead leaf base of Puya sp. (mc), dwb 2777. ARG-06-50: On Puya sp. leaves, MA-Fungi 80214, 80215, 80216; on dead leaf base of Puya sp. (mc, pH 6.8), dwb 2972. ARG-06-51: On Puya sp. leaves, MA-Fungi 80217, 80218, 80219; on dead leaf base of Puya sp. (mc, pH 6.8), dwb 2869. ARG-06-52: On Brea sp. bark (mc, pH 6.4), dwb 3011; (mc, pH 6.1), dwb 3012. ARG-06-60: On dead leaf base of Puya sp. (mc, pH 6.9), dwb 2835. ARG-06-61: On Puya sp. leaves, MA-Fungi 80220, 80221. ARG-06-64: On legume tree wood, MA-Fungi 80222; on Puya sp. leaves, MA-Fungi 80223; on bark of dead liana (mc, pH 7.1), dwb 2987; (mc, pH 7), dwb 2989. ARG-06-67: On Puya sp. leaves, MA-Fungi 80224. ARG-06-68: On dead leaf base of Puya sp. (mc, pH 6.8), dwb 2846; (mc, pH 6.9), dwb 2847; (mc, pH 7.1), dwb 2848. ARG-0672: On Puya sp. leaves, MA-Fungi 80225. ARG-06-73: On dead liana (mc, pH 7), dwb 2991. ARG-07-48: On dead leaf base of Puya sp. (mc, pH 6.6), dwb 2906, dwb 2897. ARG-07-52: On Puya sp. leaves, MA-Fungi 80226. ARG-07-56: On Puya sp. leaves, MAFungi 80227, 80228, 80229. Physarum serpula Morgan ARG-06-36: On bark of living Prosopis sp. (mc, pH 6.1), dwb These collections represent the first records of the species for Argentina. In South America, it has been reported from Chile. Physarum synsporum S.L. Stephenson & Nann.-Bremek. (Figs. 37-39). ARG-06-27: On twigs, MA-Fungi 80377. This collection has sessile, elongated plasmodiocarps, with translucent, smooth, membranous peridium, with scanty lime granules on the surface. The capillitium is typically physaroid, with large, white, amorphous lime nodes connected by hyaline threads. The spores are in tight clusters of 2-4 slightly ovoid spores (Figs. 38, 39), 11-12.5 × 8-9 µm, spinulose on the outer surface, almost smooth on the inner surface (Fig. 37). By SEM, some clusters of spores show a double line of warts along the points of spore to spore contact (Figs. 38, 39). This is one of the few species of Physarum with clustered spores. These collections represent the first record of the species for the Neotropics. Described from West Virginia, USA by Stephenson & Nannega-Bremekamp (1990). Stemonaria irregularis (Rex) Nann.-Bremek., R. Sharma & Y. Yamam. ARG-07-45: On Prosopis flexuosa bark (mc, pH 6), dwb 2909, dwb 2903. These collections are very similar to the description of Stemonaria irregularis (Nannenga- Bremekamp & al., 1984), except for the size of the spores, which are larger (10-12.5 µm diameter) in our specimens than the size range given for that species (7.5-9.5 µm diameter). Stemonitis mussooriensis G.W. Martin, K.S. Thind & Sohi ARG-06-62: On decayed Trichocereus sp., MA-Fungi 80378. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 88 88 C. Lado & al. This collection represents the first record of the species for Argentina. Stemonitopsis gracilis (G. Lister) Nann.-Bremek. ARG-06-02: On Echinopsis atacamensis remains (mc, pH 6.5), dwb 2790. This collection represents the first record of the species for Argentina. Trichia affinis de Bary ARG-06-10: On grasses, MA-Fungi 80379, 80380. ARG-06-46: On grasses, MA-Fungi 80381. ARG-06-47: On grasses, MA-Fungi 80382, 80383. ARG-06-49: On grasses, MA-Fungi 80384, 80403, 80385, 80386; on Cortaderia sp. (mc, pH 4.6), dwb 3000. Trichia agaves (G. Moreno, Lizarraga & Illana) Mosquera, Lado, Estrada & Beltran-Tej. ARG-06-50: On Trichocereus sp., MA-Fungi 80387. This collection represents the first record of the species for South America. In the Neotropics, it has been reported from Mexico Trichia contorta (Ditmar) Rostaf. ARG-06-20: On Cortaderia sp., MA-Fungi 80388. Trichia scabra Rostaf. ARG-06-27: On twigs, MA-Fungi 80389, 80390, 80391, 80392, 80393, 80394, 80395. Willkommlangea reticulata (Alb. & Schwein.) Kuntze ARG-06-14: On Stipa atacamensis, MA-Fungi 80396. ARG-0622: On grasses, MA-Fungi 80397; on twigs, MA-Fungi 80398. ARG-06-24: On grasses, MA-Fungi 80399. ARG-06-27: On twigs, MA-Fungi 80400. Discussion This biodiversity survey in the Monte Desert and surroundings has produced almost six hundred myxomycete collections from 105 localities. They represent 72 species from 22 genera, of which the genera Dianema and Macbrideola and 38 species, are new to Argentina, an increase of almost 22% to the country catalogue (Lado & Wrigley de Basanta, 2008; Wrigley de Basanta & al., 2010b). Among these 38 species, 11 are new for South America, 5 for the whole Neotropical region and 4 species are new to science, one described herein, Macbrideola andina, and three recently described based on material from this survey, Didymium infundibuliforme (Wrigley de Basanta & al., 2009), Perichaena calongei (Lado & al., 2009), and Licea eremophila (Wrigley de Basanta & al., 2010a). This brings the total number of myxomycete species known from Argentina to 211, and from this desert area the results represent 8% of the number of species known worldwide (Lado, 2005-2010), a notable number for such a dry environment. Among the interesting species from the survey, apart from those newly described, are Arcyria afroalpina, Comatricha pulchelloides, Didymium mexicanum, D. obducens, D. wildpretii, Licea sambucina, Physarum auripigmentum and Ph. synsporum, either because they are exclusive to arid ecosystems or because they are rare species. The results from the Monte Desert in these seven provinces show a high biodiversity of myxomycetes. Although for the whole survey, the species to genus ratio, a measure used to compare taxonomic diversity, with the lower numbers indicating greater diversity (Stephenson & al., 1993), is quite high (3.27), it is lower than the results (3.9) obtained in the dryland ecosystem in Mexico (Estrada-Torres & al., 2009) or (3.6) in Colorado, USA (Novozhilov & al., 2003), and comparable with other results (2.2-4.6) for temperate and tropical forests (Stephenson & al., 1993). When the results are separated by province it can be seen that some provinces have much higher taxonomic diversity than this (Table 2). The total number of species, now known from each province, indicates that Salta (57 species), Jujuy (47) and Tucumán (43) are currently the most species-rich. The table also shows the first data on myxomycetes from La Rioja, San Juan and San Luis, provinces previously unexplored for myxomycetes. In order to directly compare the results from each province, the total number of collections was adjusted to take into account the number of collecting sites that had identifiable collections of myxomycetes included in the annotated species list (positive site). Catamarca in this case had the greatest number of collections per positive collecting site, followed by Salta and Tucumán. The mean result was almost 8 species recovered from each locality, irrespective of the province. The most abundant species (Table 3), by number of collections, were Badhamia melanospora, Physarum pusillum, Perichaena calongei, P. vermicularis, Craterium leucocephalum, Didymium infundibuliforme, D. vaccinum and Echinostelium colliculosum. Many of these are recognised as characteristic species of arid environments, and here more abundant than other species such as Didymium squamulosum, Physarum bitectum and Arcyria cinerea of broad distribution, usually among the common species in studies of other environments. The assemblage of abundant species coincides with that obtained in other studies of warm arid areas such as the Tehuacán-Cuicatlán valley in Mexico (Estrada-Torres & al., 2009), although there Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 89 Biodiversity of Myxomycetes 89 Table 2. Summary data on the Myxomycetes in different Provinces of the survey area. Species a) from the literature-all vegetation types, b) this survey-arid vegetation. Province Localities Total (positive) Collections Catamarca 19 (15) 151 Jujuy 18 (12) La Rioja Collections per positive site b Different species total Genera a S/G ratio 10.07 6 33 37 13 2.85 87 7.25 27 21 47 24 1.96 11 (8) 62 7.75 0 20 20 10 2.00 Salta 25 (19) 166 8.74 10 48 57 23 2.48 San Juan 24 (18) 84 4.67 0 21 21 9 2.33 San Luis 6 (4) 28 7.00 0 12 12 8 1.50 Tucumán 2 (2) 17 8.50 41 3 43 18 2.34 are some differences probably due to different substrate species (Wrigley de Basanta & al., 2010a). Surprisingly, in spite of the intense work done over recent decades in the Neotropics (Lado & Wrigley de Basanta, 2008), almost 30% (27.8) of the species found in the Monte Desert are new records for South America and 12.5% are new records for the Neotropics, confirming the exclusive myxobiota of this dryland ecosystem. A relatively high number of species (Didymium infundibuliforme, D. mexicanum, D. wildpretii, Licea eremophila, L. succulenticola, Macbrideola andina, Perichaena calongei, Physarum spectabile, Trichia agaves) also belong to the succulenticolous species group. The most abundant species were also the most widespread, as can be seen (Table 3) from the number of localities from which they were recovered. Of the total 72 species listed above, 25 were only found once which may indicate that these are the rarer species such as Didymium obducens, or that at the time of collecting there were not the ideal phenological conditions for some species or adequate substrate for others, such as Lycogala epidendrum, a typical lignicolous species. The most common genera belonged to the order Physarales which made up almost 50% of the collections. This has been noted in other arid areas, for instance the Tehuacán valley in Mexico (Estrada-Torres & al., 2009), and Atacama desert in Chile (Lado & al., 2007a). In the Monte Desert however, the percentage of species of the genus Physarum was greater than that of Didymium species, unlike the results from both Mexico and Chile where the genus Didymium was predominant. In all three places, the species within the genera differed, except for the most common species, indicating precise microhabitat preferences, since the substrate species were also different. This was Species particularly the case with the genus Didymium which had only 5 of the 30 species of the genus from Tehuacán in common. The species in the order Physarales made up only 32% of the total found in the Colorado plateau, USA (Novozhilov & al., 2003), but different woody vegetation was mainly sampled there. The sequence of orders was also basically the same in Mexico, and in a review of all Neotropical myxomycetes (Lado & Wrigley de Basanta, 2008), where results were compared to the percentage of known species from each order. The order Trichiales were slightly better represented in these results from Argentina, and in spite of the known number of species in the order being below that of the Liceales and Stemonitales, more species of Trichiales were found than of the other two orders. The most productive substrates of this study were the more than twenty species of cacti, which produced 37% of all the specimens. The leafy substrates were the next most productive with 35% of the results. This substrate group includes mainly the leaf bases of the rosette-leaved succulent plant Puya sp. (Fig. 3), a particularly productive substrate both in the field and in moist chamber culture. The number of species however was greater in the latter group with 32 different species compared to 25 different species from cacti. Similar rosette-leaved succulent plants, but of other genera such as Agave, Beaucarnea, and Hechtia, were the most productive group of substrata in the Tehuacán valley in Mexico (Estrada-Torres & al., 2009). As has been noted previously (Wrigley de Basanta & al., 2010a) the rosettes form a water trap where any available moisture, even from condensation, in these extreme desert environments is channeled towards the base of the plant. The overlapping leaves prevent evaporation and, in the field, dead leaves Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 90 90 C. Lado & al. Table 3. Summary data on the most common Myxomycete species. Species Field collections Moist chamber Total collections collections Percentage Nº localities of Records found Arcyria afroalpina 3 10 13 2.20 7 Arcyria cinerea 4 8 12 2.02 9 123 36 159 26.72 50 17 3 20 3.36 10 7 12 19 3.19 10 Didymium squamulosum 12 1 13 2.20 4 Didymium vaccinum 13 6 19 3.19 8 Echinostelium colliculosum 0 16 16 2.69 9 Licea eremophila 3 8 11 1.85 5 Perichaena calongei 17 9 26 4.37 10 Perichaena depressa 11 0 11 1.85 6 Perichaena quadrata 2 11 13 2.20 8 14 12 26 4.37 14 9 3 12 2.02 5 10 4 14 2.35 6 Physarum notabile 9 4 13 2.20 8 Physarum pusillum 17 12 29 4.87 17 Physarum spectabile 4 7 11 1.85 8 Trichia affinis 9 1 10 1.68 4 284 162 446 75.18 – Badhamia melanospora Craterium leucocephalum Didymium infundibuliforme Perichaena vermicularis Physarum bitectum Physarum compressum TOTAL (19 species) around the base were often moist even in the middle of the day in mid summer. On bark, sixteen species were recovered, in spite of the paucity of woody substrates in these arid areas. The majority of bark was from species of the leguminous trees of the genus Prosopis, which was also a productive substrate in Mexican drylands (Estrada-Torres & al., 2009). In the grasslands of the puna and surrounding areas, in spite of the high elevation (above 3000 m), the high levels of solar radiation and extreme temperature differences day and night, the different species of grasses also produced a reasonable number of collections (almost 8%), and 12.5% of the species found in this survey. In order to compare the results according to elevation and latitude of the sampling area, the results were expressed as collections and species of myxomycetes per locality, to correct for differences in sampling ef- fort (Fig. 59). In the case of elevation there was a trend towards decreasing number of specimens with increasing elevation in the sampling range of 500 m to over 4000 m, as was found in other areas of Argentina (Wrigley de Basanta & al., 2010b). This was not the case for the number of species, as this remained more or less constant over the range. These values could only reflect the moment of sampling, which may represent a better time for myxomycetes at lower elevations and not at higher elevations, since sampling was done over the same period. The climatic conditions over this large gradient can vary considerably and affect the substrates for myxomycete development. The range of latitudes in this survey was from South latitude 23º to 33º. However the results according to latitude, corrected for differences in collecting effort, were fairly uniform, without a significant trend. The Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 91 Biodiversity of Myxomycetes 91 Fig. 59. Number of Myxomycetes from different elevations (positive sites only). largest number of collections (16%), were made at latitude 28ºS and the combined number of collections at the lower latitudes (23º-27ºS), was much greater (60%) than the 24% at the higher latitudes (29º33ºS). This is consistent with data from other studies in Argentina (Wrigley de Basanta & al., 2010b). The results include 28 species only found in the field, 19 species only produced in moist chamber culture and 25 species occurring in both field collections and from moist chamber culture. The 127 moist chamber cultures were 82% positive for myxomycete plasmodia or fruiting bodies. As in other studies (Estrada-Torres & al., 2009; Wrigley de Basanta & al., 2010b) some plasmodia only produced poor or malformed specimens, not included in the results, and in some cases only formed sclerotia that never fruited in the time span of the cultures. The productivity is high for moist chambers made with plants from such a dryland area, and higher than that of 250 moist chambers of cactus remains from dryland ecosystems of Mexico (Estrada-Torres & al., 2009), but lower than that for the whole study area of the Colorado Plateau (Novozhilov & al., 2003). One of the important microenvironmental factors affecting the abundance and diversity of myxomycetes is the pH of the substrate, as has been indicated in other studies (Wrigley de Basanta, 2004; Wrigley de Basanta & al., 2008). The moist chamber cultures from this survey had a broad substrate pH range from 4.4-9.2, but the majority of myxomycete collections were harvested from substrates with a pH falling between 6 and 7.9 (Fig. 60), and the mean pH of all cultures was circumneutral at pH 7.03. All but one of the cultures with a basic pH of 8.0 or above, were made with different cactus remains, and although some of the cultures made with epidermis of cacti had a pH nearer to 7, the mean of the cactus cultures was almost 8 (pH 7.95). The basic pH may favour the development of the lime-producing Physarales predominant in these results. The moist chamber cultures made with leaf bases of Puya sp., the next most productive of all the substrates after the cacti, had a mean pH of 6.95, again close to neutral as with other productive substrates (Wrigley de Basanta & al., 2008). Badhamia melanospora and Echinostelium colliculosum were the most tolerant species from the moist chamber cultu- Fig. 60. Proportion of moist chamber collections at different substrate pH values. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 92 92 C. Lado & al. res, and were recovered from substrates with pH across the range. All the species of Arcyria were found on substrates with a circumneutral pH, as were Licea eremophila and Perichaena calongei, but Physarum spectabile was found only on basic substrates. During this survey 73 taxa, 72 species and one variety (Craterium leucocephalum var. scyphoides) were obtained from various substrates. The number of species expected, if the sampling effort were exhaustive, according to the estimators ACE and CHAO1, are calculated at 104 and 94 respectively (Fig. 61). This means that the sampling effort of this survey was 7078% complete. If the results of the field collections and collections recovered from moist chamber culture are assessed separately, the former recovered 6972% of the expected species by each estimator and the moist chamber cultures 81-84% respectively. This suggests that the survey recovered a large proportion of the assemblage of myxomycetes to be expected in the Monte Desert. The results are surprisingly high when considering the fact that field sampling may not coincide with the best phenological moment for some species. In addition the substrates for moist chamber cultures may have spores, microcysts or sclerotia that have been exposed for too long to the harsh conditions of the desert, and are no longer viable. To assess community similarities between this and studies of other arid areas in the literature, the Sørensen coefficient of community (CC) index was used (Table 4). The myxomycete assemblage from the TehuacánCuicatlán desert of Mexico was the most similar to the Monte Desert of Argentina (CC = 0.46). The relationships between the myxomycete assemblages of these areas could be due to similarities in their substrates. The floristic affinities between arid zones of North America and the Monte Desert were researched by Solbrig (1972) and Roig & al. (2009), who found that the genera Larrea, Cercidium and Prosopis, and members of the families Cactaceae, Agavaceae and Bromeliaceae, are the dominant plants shared in common between these areas. The microhabitats of the related plants in these taxa are similar, and so the community of myxomycetes developing in them overlaps, as pointed out by Wrigley de Basanta & al. (2010a) for the myxobiota of the genera Hechtia and Puya. The apparently least similar assemblage (CC= 0.22) was from the Atacama Desert in Chile, on the other side of the Andes mountain chain. This is not surprising given the extreme arid environment, which limits the myxomycete species richness (Lado & al., 2008), and therefore produced a much smaller sample. However, despite the low number of common species (14) this number supposed almost 60% of the species found in that study of one of the driest places on earth. Fig. 61. Curves of abundante (ACE and CHAO1 estimators) compared to these species accumulation curves (Sobs) of this Surrey. White lines indicate the polinomial best fit curve. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 93 Biodiversity of Myxomycetes 93 Table 4. Community similarity between myxobiota of arid areas using the coefficient of community index (CC). (Top right CC, number of species in common bottom left). Monte Tehuacán-Cuicatlán Atacama Colorado Plateau – 0.46 0.29 0.35 41 – 0.28 0.45 Atacama desert, Chile (Lado & al., 2007) 14 18 – 0.22 Colorado Plateau, USA (Novozhilov & al., 2003) 29 44 13 – Monte desert, Argentina Tehuacán-Cuicatlán Valley, Mexico (Estrada-Torres & al., 2009) More recently, succulenticolous species Didymium infundibuliforme (Wrigley de Basanta & al., 2009) and Licea eremophila (Wrigley de Basanta & al., 2010a) were found associated with species of succulent plants on both sides of the Andes. The Colorado Plateau species composition was between the other two. The uniformity of method has to be considered when comparing these studies. In all of them, the results are a combination of field results and those from moist chamber cultures. However the total number of collections in each study was different, as was the emphasis placed on fieldwork or cultures. In addition, in the Colorado Plateau study, the vegetation was very different from the other three areas since the research there centered on sagebrush and woodland communities, concentrating on woody substrates absent or very rare in the Monte Desert, and included herbivore dung, a substrate less common in the Monte. The present study adds data to confirm some of the factors that appear to be critical for the development of myxomycetes. It is evident that the macroenvironmental factors such as temperature, rainfall, elevation and indirectly latitude, while influencing the growth of the substrate plants, do not show a direct influence on the species composition or abundance of the myxomycetes found in this area. On the other hand, the microenvironmental factors do seem to influence which myxomycetes develop and how abundantly they appear. The microhabitat in and on specific plants is a complex of many variable factors. The chemical composition of different plant species is different which influences the pH of the tissue, its capacity for water retention, and release of nutrients, which in turn determine the microbial flora living in or on and decomposing the plant, as suggested by Mosquera & al., (2003). The microbial biota, the food organisms for myxomycetes, probably alter the microhabitat in the course of decay and a succession of flora and other organisms, such as fly larvae, occurs (Fogleman & Foster, 1989; Foster & Fogleman, 1993). This complex of interacting abiotic and biotic factors within the microhabitat could explain the differences in myxobiota on different substrates and the apparent substrate specificity seen for example in Licea eremophila, that flourished in the Monte Desert on one type of rosette-leaved plant, Puya sp., and was absent from other similar plants, e.g. Hechtia sp., in Mexico (Wrigley de Basanta & al., 2010a). Other substrate, or substrate-group, specificity has been seen with Didymium infundibuliforme on Puya sp. (Wrigley de Basanta & al., 2009), Didymium tehuacanense on Agave sp., and Didymium subreticulosporum, D. wildpretii and Badhamia melanospora on cacti (Estrada-Torres & al., 2009). On the succulent plant species and cacti of the Monte Desert, several of the succulenticolous species of myxomycetes, Didymium wildpretti, D. vaccinum, Licea succulenticola, Physarum spectabile, Trichia agaves, found in other desert areas have re-appeared. There are many species that do not seem to have such a narrow specificity for substrate, and are found on a large variety of plants, as can be seen with Arcyria cinerea, Didymium squamulosum, Perichaena vermicularis or Physarum pusillum, obviously responding to a general number of conditions common to those microhabitats. It is interesting to note, as mentioned above, that where there were different plant species within similar habitats, studied in a similar manner, genus Didymium had only 17% of the species in common. The myxomycetes found in the Monte Desert represent 8% of the number of species known worldwide, confirming an unexpected species richness of the area. Their colonization and ability to live in such arid environments demonstrate the importance of the resistant stages in their life cycle, where three different life stages, microcysts, sclerotia and spores, increase their ability to survive the extreme adverse environmental conditions of this dryland ecosystem. In the sampling for this survey, both in the field and in culturing by moist chamber, as mentioned above, sclerotia Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 94 94 C. Lado & al. were very common, attesting to their importance as resistant stages. Some of these sclerotia collected from the field were placed on agar and produced viable plasmodia. If the fact that myxomycetes require water, at least for their feeding stages as amoebae or plasmodia, a desert could be considered a totally hostile and impossible place for their development, but these results, showing an unexpectedly high biodiversity of myxomycetes, refute this assumption. Acknowledgements This research was supported by the Ministry of Science and Innovation, Spain (projects CGL2005-00320/BOS and CGL200800720/BOS). We are grateful to Laura Lorenzo, Comahue University, Argentina, for logistical help and the personnel of the Parques Nacionales de Argentina, and the Parque Natural Provincial Ischigualasto, for help and permission to collect. We also thank Yolanda Ruiz for her technical assistance with SEM and Carlos de Mier for his help with the light photomicrographs. References Abraham, E., Valle , H.F. del, Roig, F., Torres, L., Ares, J.O., Coronato, F. & Godagnone, R. 2009. Overview of the geography of the Monte Desert biome (Argentina). J. Arid Environm. 73(2): 144-153. Adl, S.M., Simpson, A.G.B., Farmer, M.A., Andersen, R.A., Anderson, O.R., Barta, J.R., Bowser, S.S., Brugerolle, G., Fensome, R.A., Fredericq, S., James, T.Y., Karpov, S., Kugrens, P., Krug, J., Lane, C.E., Lewis, L.A., Lodge, J., Lynn, D.H., Mann, D.G., McCourt, R.M., Mendoza, L., Moestrup, Ø., MozleyStandridge, S.E., Nerad, T.A., Shearer, C.A., Smirnov, A.V., Spiegel, F.W. & Taylor, M.F.J.R. 2005. The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. J. Eukaryot. Mycrobiol. 52(5): 399-451. Anonymous. 1976. ISCC-NBS color-name charts illustrated with centroid colors. Washington, DC: Inter-Society Color Council, National Bureau of Standards. Baldauf, S.L. 2008. An overview of the phylogeny and diversity of eukaryotes. J. Syst. Evol. 46(3): 263-273. Blackwell, M. & Gilbertson, R. L. 1980. Sonoran desert Myxomycetes. Mycotaxon 11(1): 139-149. Chung, C.-H. & Tzean, S.-S. 1998. Observations on Physarum hongkongense (Physarales, Myxomycetes) from Taiwan. Fungal Sci. 13(3,4): 109-112. Colwell, R.K. & Coddington, J.A. 1994. Estimating terrestrial biodiversity through extrapolation. Philos. Trans. Roy. Soc. Lond. B 345: 101-118. Colwell, R.K., Mao, C.X. & Chang, J. 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 85(10): 2717-2727. Crespo, E.M. & Lugo, M. 2003. Catalogue of Myxomycetes from Argentina. Mycotaxon 87: 91-102. Digilio, A.P.L. 1946. Contribución al catálogo de los "Myxomycetes" argentinos. I. Lilloa 12: 177-203. Digilio, A.P.L. 1950. Myxomycetes de Tucumán. Lilloa 23(4): 365-414. Deschamps, J.R. 1972. El género Stemonitis (Myxomycetidae) en Argentina. Bol. Soc. Argent. Bot. 14(3): 139-153. Deschamps, J.R. 1976. Los Myxomycetes de la Argentina. Catálogo crítico, distribución y clave de las especies. (Conclusión). Physis (Buenos Aires) 35(91): 319-339. Estrada-Torres, A., Wrigley de Basanta, D., Conde, E. & Lado, C. 2009. Myxomycetes associated with dryland ecosystems of the Tehuacán-Cuicatlán Valley Biosphere Reserve, Mexico. Fungal Diversity 36: 17-56. Evenson, A.E. 1961. A preliminary report of the Myxomycetes of Southern Arizona. Mycologia 53: 137-144. Fogleman, J.C. & Foster, J.L.M. 1989. Microbial colonisation of injuried cactus tissue (Stenocereus gummosus) and its relationship to the ecology of cactophilic Drosophila mojavensis. Appl. Environm. Microbiol. 55: 100-105. Foster, J.L.M. & Fogleman, J.C. 1993. Identification and ecology of bacterial communities associated with necroses of three cactus species. Appl. Environm. Microbiol. 59: 1-6. Fries, R.E. 1903. Myxomyceten von Argentinien und Bolivia. Ark. Bot. 1: 57-70. Härkönen, M. 1979. Additions and corrections to the Finnish flora of Myxomycetes. Karstenia 19: 1-7. Ing, B. 1994. The phytosociology of Myxomycetes. New Phytol. 126: 175-201. Kosheleva, A.P., Novozhilov, Y.K. & Schnittler, M. 2008. Myxomycete diversity of the state reserve "Stolby" (south-eastern Siberia, Russia). Fungal Diversity 31: 45-62. Lado, C. 2005-2010. An on line nomenclatural information system of Eumycetozoa. http://www.nomen.eumycetozoa.com (14VI-2010). Lado, C. & Wrigley de Basanta, D. 2008. A review of Neotropical Myxomycetes (1828-2008). Anales Jard. Bot. Madrid 65(2): 211254. Lado, C., Estrada-Torres, A. & Stephenson, S.L. 2007a. Myxomycetes collected in the first phase of a north-south transect of Chile. Fungal Diversity 25: 81-101. Lado, C., Mosquera, J. & Beltrán-Tejera, E. 1999. Cribraria zonatispora, development of a new myxomycete with unique spores. Mycologia 91(1): 157-165. Lado, C., Mosquera, J., Estrada-Torres, A. & Wrigley de Basanta, D. 2007b. Description and culture of a new succulenticolous Didymium (Myxomycetes). Mycologia 99(4): 602-611. Lado, C., Wrigley de Basanta, D., Estrada-Torres, A., García Carvajal, E., Aguilar, M. & Hernández, J.C. 2009. Description of a new species of Perichaena (Myxomycetes) from arid areas of Argentina. Anales Jard. Bot. Madrid 66S1: 63-70. Lizárraga, M., Moreno, G. & Illana, C. 2006. Macbrideola herrerae sp. nov., a new myxomycete from Mexico. Bol. Soc. Micol. Madrid 30: 265-269. Martin, G.W. 1948. Two new species of Physarum. J. Wash. Acad. Sci. 38: 238-240. Martin, G.W. & Alexopoulos, C.J. 1969. The Myxomycetes. Univ. Iowa Press. Iowa. Meylan, C. 1927. Recherches sur les Myxomycètes du Jura en 1925-26. Bull. Soc. Vaud. Sci. Nat. 56: 319-328. Mitchell, D.W., McHugh, R. 2000. Recent discoveries of corticolous Licea species from the British Isles with descriptions of two new species. Karstenia 40: 103-109. Moreno, G., Lizárraga, M. & Illana, C. 2006. Macbrideola lamprodermoides sp. nov., a new myxomycete from Mexico. Bol. Soc. Micol. Madrid 30: 255-263. Mosquera, J., Lado, C., Estrada-Torres, A., Beltrán Tejera, E & Wrigley de Basanta, D. 2003. Description and Culture of a new Myxomycete, Licea succulenticola. Anales Jard. Bot. Madrid 60(1): 3-10. Nannenga-Bremekamp, N.E. 1965. Notes on Myxomycetes IX. The genus Licea in the Netherlands. Acta Bot. Neerl. 15: 131147. Nannenga-Bremekamp, N.E. 1985. Notes on Myxomycetes XXII. Three new species, two new families and four new combinations. Proc. Kon. Ned. Akad. Wetensch., Ser. C 88(1): 121-128. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 95 Biodiversity of Myxomycetes Nannenga-Bremekamp, N.E. & Yamamoto, Y. 1983. Additions to the Myxomycetes of Japan. I. Proc. Kon. Ned. Akad. Wetensch., Ser. C 86(2): 207-241. Nannenga-Bremekamp, N.E. & Yamamoto, Y. 1990. Additions to the Myxomycetes of Japan. Proc. Kon. Ned. Akad. Wetensch. IV 93(3): 265-280. Nannenga-Bremekamp, N.E., Yamamoto, Y. & Sharma, R. 1984. Stemonaria, a new genus in the Stemonitaceae and two new species of Stemonitis (Myxomycetes). Proc. Kon. Ned. Akad. Wetensch., Ser. C 87(4): 449-469. Ndiritu, G.G., Winsett ,K.E., Spiegel, F.W. & Stephenson, S.L. 2009. A checklist of African myxomycetes. Mycotaxon 107: 353-356. Novozhilov, Y.K., Mitchell, D.W. & Schnittler, M. 2003. Myxomycete biodiversity of the Colorado Plateau. Mycol. Progress 2(4): 243-258. Pando, F. & Lado, C. 1988. Two new species of corticolous Myxomycetes from Spain. Mycotaxon 31: 299-303. Patouillard, N. & Lagerheim, G. de. 1895. Champignons de l'Equateur (Pugillus V). Bull. Soc. Mycol. France 11: 205-234. Rammeloo, J. 1981a. Five new Myxomycete species (Trichiales) from Rwanda. Bull. Jard. Bot. Natl. Belgique 51(1/2): 229-230. Rammeloo, J. 1981b. Trichiales (Myxomycetes). In: Flore illustrée des champignons d' Afrique Centrale. Ministère de l'Agriculture. Jardin Botanique National de Belgique. Meise. Roig, F.A., Roig-Juñent, S. & Corbalán, V. 2009. Biogeography of the Monte Desert. J. Arid Environm. 73(2): 164-172. Rojas, C. & Stephenson, S.L. 2008. Myxomycete ecology along an elevation gradient on Cocos Island, Costa Rica. Fungal Diversity 29: 117-127. Rossman, A.Y., Tulloss, R.E., O’Dell, T.E. & Thorn, R.G. 1998. Protocols for an all taxa biodiversity inventory of Fungi in a Costa Rican conservation area. Parkway Oublishers, Inc. Boone, North Carolina. Solbrig, O.T. 1972. The floristic disjunctions between the “Monte” in Argentina and the “Sonoran Desert” in Mexico and the United States. Ann. Missouri. Bot. Gard. 59: 218-223. 95 Spegazzini, C. 1899. Fungi argentini novi vel critici. Anales Mus. Nac. Hist. Nat. Buenos Aires 6: 81-367. Stephenson, S.L. 1989. Distribution and eology of myxomycetes in temperate forests. II. Patterns of occurrence on bark surface of living trees, leaf litter, and dung. Mycologia 81: 608-621. Stephenson, S.L. & Nannenga-Bremekamp, N.E. 1990. Five new species of Myxomycetes from North America. Proc. Kon. Ned. Akad. Wetensch., Ser. C 93(2): 187-195. Stephenson, S.L., Kalyanasundaram, I. & Lakhanpal, T.N. 1993. A comparative biogeographical study of myxomycetes in the mid-Appalachian of eastern North America and two regions of India. J. Biogeogr. 20: 645-657. Whitney, K.D. 1980. The Myxomycete genus Echinostelium. Mycologia 72(5): 950-987. Wrigley de Basanta, D. 2004. The effect of simulated acid rain on corticolous myxomycetes. Syst. Geogr. Pl. 74: 175-181. Wrigley de Basanta, D. & Lado, C. 2005. A taxonomic evaluation of the stipitate Licea species. Fungal Diversity 20: 261-314. Wrigley de Basanta, D., Stephenson, S.L., Lado, C., Estrada-Torres, A. & Nieves-Rivera, A.M. 2008. Lianas as a microhabitat for myxomycetes in tropical forests. Fungal Diversity 28: 109125. Wrigley de Basanta, D., Lado, C., Estrada-Torres, A. & Stephenson, S.L. 2009. Description and life cycle of a new Didymium (Myxomycetes) from arid areas of Argentina and Chile. Mycologia 101(5): 707-716. Wrigley de Basanta, D., Lado, C. & Estrada-Torres, A. 2010a. Licea eremophila a new Myxomycete from arid areas of South America. Mycologia 103: in press. doi:10.3852/09-309. Wrigley de Basanta, D., Lado, C., Estrada-Torres, A. & Stephenson, S.L. 2010b. Biodiversity of myxomycetes in subantarctic forests of Patagonia and Tierra del Fuego, Argentina. Nova Hedwigia 90(1-2): 45-79. Anales del Jardín Botánico de Madrid 68(1): 61-95, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2266 Associate Editor: J. Guarro Received: 27-VII-2010 Accepted: 16-II-2011 2266_Myxomycetes:Anales 68(1).qxd 13/06/2011 12:12 Página 96 2276_Artemisa:Anales 68(1).qxd 13/06/2011 12:22 Página 97 Anales del Jardín Botánico de Madrid Vol. 68(1): 97-105 enero-julio 2011 ISSN: 0211-1322 doi: 10.3989/ajbm.2276 Self-incompatibility, floral parameters, and pollen characterization in the narrow endemic and threatened species Artemisia granatensis (Asteraceae) by Julio Peñas1, Juan Lorite1, Francisca Alba-Sánchez1 & María Angélica Taisma2 2 1 Plant Conservation Unit, Department of Botany, University of Granada, E-18071 Granada, Spain. [email protected] Instituto de Biología Experimental, Centro de Botánica Tropical, Universidad Central de Venezuela, Caracas, Venezuela Abstract Resumen Peñas, J., Lorite, J., Alba-Sánchez, F. & Taisma, M.A. 2011. Selfincompatibility, floral parameters, and pollen characterization in the narrow endemic and threatened species Artemisia granatensis (Asteraceae). Anales Jard. Bot. Madrid 68(1): 97-105. Peñas, J., Lorite, J., Alba-Sánchez, F. & Taisma, M.A. 2011. Autoincompatibilidad, parámetros florales y caracterización de polen en la especie endémica y amenazada Artemisia granatensis (Asteraceae). Anales Jard. Bot. Madrid 68(1): 97-105 (en inglés). Artemisia granatensis Boiss. is a paradigmatic species for plant conservation in Spain and Europe. It is a critically endangered (CR) endemic species growing above 2500 m in the Sierra Nevada (southern Spain). Natural populations have been considerably devastated in the past due to intensive human exploitation for folk medicine. The sparse available data concerning the reproductive biology of this species under natural conditions indicate a low reproductive success. To provide additional information on the reproductive biology of A. granatensis, and consequently information useful for the management and conservation of this species, we studied the breeding system through pollen-tube growth. In addition, some floral and pollen traits were recorded. No differences were found between populations in terms of the morphological traits of flowers and inflorescences. A. granatensis is an anemophilous species, and the data indicate that pollen transfer may be limited between isolated populations, and so contributing to an extremely low fruit-set. Results show A. granatensis is selfincompatible, probably with a sporophytic self-incompatibility system, and with no evidence of partial self-incompatibility. Reproductive traits, related to pollen morphology and settling speed may explain the low rate of recruitment in the small populations separated by geographical barriers. Artemisia granatensis Boiss. es una especie paradigmática en la conservación de flora a nivel español y europeo. Es una especie catalogada como En Peligro Crítico (CR) endémica de Sierra Nevada (sur de España), donde habita por encima de los 2500 m. Las poblaciones naturales han sido casi exterminadas en el pasado debido a una recolección masiva de la especie, utilizada en medicina popular. Los escasos datos disponibles acerca de su biología reproductiva en condiciones naturales indican que existe un bajo éxito reproductivo. Con el objetivo de proporcionar información adicional acerca de la biología reproductiva de A. granatensis, útil para la conservación y el manejo de la especie, evaluamos el sistema de compatibilidad a través del crecimiento del tubo polínico. Además se registraron datos sobre algunos rasgos florales y polínicos de la especie. No se encontraron diferencias entre poblaciones en términos de rasgos morfológicos de flores e inflorescencias. A. granatensis es una especie anemófila para la cual los datos obtenidos sobre capacidad de dispersión sugieren que la transferencia de polen podría ser difícil entre poblaciones aisladas o muy distanciadas, pudiendo ser un factor más a tener en cuenta entre las causas que provocan un limitado éxito reproductivo y una paupérrima producción de semillas. Los resultados muestran que A. granatensis tiene autoincompatibilidad esporofítica sin evidencias de autoincompatibilidad parcial. Los rasgos reproductivos relacionados con la morfología y la velocidad de sedimentación del polen pueden explicar la baja tasa de reclutamiento de poblaciones pequeñas, a veces separadas por barreras geográficas. Keywords: reproductive biology, pollen-tube growth, endemic and threatened species, conservation. Palabras clave: biología reproductiva, crecimiento del tubo polínico, especie endémica y amenazada, conservación. 2276_Artemisa:Anales 68(1).qxd 13/06/2011 12:22 Página 98 98 J. Peñas & al. Introduction The southern area of the Iberian Peninsula has long been recognized as a centre of plant diversity and endemicity (Molero, 1994; Domínguez & al., 1996; Peñas & al., 2005), and a “phytogeographical hotspot”; that is, significant reservoirs of unique genetic diversity favourable to the evolutionary processes of Mediterranean plant species (Médail & Diadema, 2009). The largest number of endemic plant species, and indeed one of the largest in Europe, is found in Sierra Nevada (Blanca & al., 1998). This mountain harbours the narrow endemic Artemisia granatensis Boiss. (Asteraceae), a paradigmatic species for the plant conservation in Spain and Europe (Council Directive 92/43/EEC; Fay, 1992), since it was included in the first Spanish catalogue of threatened species (BOE, 1990). Today, this species is considered critically endangered (CR) in the latest national (Moreno, 2008) and regional (Cabezudo & al., 2005) list of threatened species, with the main threats being overgrazing and collection for folk medicine. Estimates of population sizes suggest that about 3000 individuals of the species survive (Blanca & al., 1998), distributed in 12 populations (Blanca, 2002). Habitat fragmentation leading to small isolated populations may be the most apparent cause for reproductive failure and species loss (Koul & Bhatnagar, 2007). Up to the present, the efforts to recover natural populations of A. granatensis have not been based on a knowledge of the reproductive biology of natural populations. Furthermore, the difficult access to the small and distant populations of A. granatensis has constrained reproductive studies in situ. However, field studies on reproductive traits, compatibility systems, and pollination mechanisms are necessary to define conservation strategies for the species. Reproductive-biology studies, thus, should be an integral feature of any conservation project (Weller, 1994; Weekly & Race, 2001; Koul & Bhatnagar, 2007). Self-incompatibility, a genetic barrier to prevent inbreeding that is broadly distributed among angiosperms, could be a main constraint against reproductive success in A. granatensis because in selfincompatible species there may be a loss of genetic diversity among individuals in fragmented and scattered populations that dooms such isolated populations to extinction (Weller, 1994). Since A. granatensis, belonging to Asteraceae, a family with selfincompatible established in around 40 genera (Charlesworth, 1985), and with some reports on partial self-incompatibility (Ortiz & al., 2006), could have a self-incompatibility system which has not previously been tested. Some studies have related self-incompatibility systems to floral and inflorescence size (Gibbs & al., 1975; Ortiz & al., 2006). For example, in the genus Hypochaeris L. partial self-compatible heads are larger than self-incompatible heads (Ortiz & al., 2006). Thus, the evaluation of a self-incompatibility system must include the flower and inflorescence morphology in order to gain a full understanding of the reproductive potential. Artemisia is known to be an anemophilous genus (O’Brien, 1980; Watson & al., 2002), and ecological characteristics of in A. granatensis populations could hamper the transfer of pollen between distant populations due to geographical barriers. Currently we have no data about the potential A. granatensis pollen movement in the atmosphere. To help fill this gap, theoretical data on the settling speed and residence time in are presented. This information was obtained based on some morphological pollen features. Data about some physical properties of the A. granatensis pollen are important building blocks in a model of its pollen movement and, as such, later will be helpful in establishing the main factors influencing the dispersion degree of pollen between A. granatensis populations. Consequently, the settling speed of this pollen is fundamental for determining the distance that this particle can be transported in the atmosphere as well as its probability of being deposited on the plants or on the ground (Aylor & al., 2005). The aim of the study is provide information on the reproductive biology of A. granatensis, in relation to pollen features and breeding system, useful for the management and conservation of this species. For this propose, the present study i) characterizes the basic pollen morphological and functional parameters and compares floral morphology within and between populations, and ii) evaluates the operation presence of a self-incompatibility system by means of hand pollinations and pollen-tube growth. Material and methods Study area and selected sites Sierra Nevada, a mountain range of some 2,100 km2 located in SE Spain (37ºN, 3ºW), has a complex orography, bedrock, and soil composition (e.g. areas with dolomitic soils), and reaches a height of 3482 m. This massif marks the southernmost limit of the influence of the Quaternary glaciations in Europe, when this mountain range was covered with glaciers in areas above around 2,500 m while large areas remained free of permanent ice (Gómez & al., 2001). All these features have contributed to make the Sierra Nevada a Anales del Jardín Botánico de Madrid 68(1): 97-105, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2276 2276_Artemisa:Anales 68(1).qxd 13/06/2011 12:22 Página 99 Artemisia granatensis refuge for many plant species during glacial ages (Blanca & al., 1998), thus harbouring isolated populations that have evolved under particular conditions (e.g. soil type or isolated summit areas), and which have encouraged speciation and a high level of endemicity (Blondel & Aronson, 1999; Peñas & al., 2005; Thompson, 2005). In fact, the area above 2,000 m contains about 100 endemic or rare taxa. Many of these species are threatened by different factors (Blanca & al., 1998; Bañares & al., 2003), with 38 taxa included on the regional protection list (Blanca & al., 1999). Today most of this entire area lies within National and/or Natural parks. Climatic conditions are typically alpine (with a Mediterranean character), with mean temperatures below 0 °C during winter months and snow cover that can persist up to 8 months in the highest places (occasionally up to 10 months in small, protected areas). See Gómez (2002) for a detailed description of climatic conditions in Sierra Nevada range. Two populations of A. granatensis were selected in the summit area of the Sierra Nevada. The first (AG1 hereafter), at 2790 m on the northern slope, included 45 mature individuals, and the second (AG2 hereafter), at 3045 m on the southern slope, had 55 mature individuals. In both cases there are no other patches or isolated individuals in the surroundings (at least 300 m around). The populations were c. 6 km apart. Because of the difficult access to the populations, the low number of individuals per population, and the use of a semi-extractive sampling design, we limited the sampling effort to the minimum necessary to allow the statistical analysis of the data. Studied species Artemisia granatensis Boiss. belongs to the large family of Asteraceae. Artemisia is the largest genus in the tribe Anthemideae and one of the largest in the family, with over 500 species (Martin & al., 2001). Taxonomically, A. granatensis is closely related to other alpine species such as A. splendens Willd. and A. umbelliformis Lam. (Watson & al., 2002), and it occasionally forms a hybrid, A. × fragosoana Font Quer (Blanca, 2002) with the latter. A. granatensis is a longlived perennial herb, caespitose and white-sericeous, with stems 5-12 cm long, erect, simple or scarcely branched. Basal leaves are numerous, petiolate, divided with segments of flabellate outline. Flowers are arranged in terminal discoid capitula of 5-8 mm in diameter, with 1-5 capitula per stem. The capitula have external flowers female and internal ones hermaphrodite, being a gynomonoecious species. Flowers are actinomorphic, tubulose, dark purple in 99 colour, with a papillose stigma. Flowering ranges from July to August and fruit ripening (small achene) occur in September. Although information concerning the breeding system of the species is scant, A. granatensis is known to be an anemophyilous species (e.g. Blanca & al., 1999). A. granatensis appears in perennial high-mountain pastures on mica-schists, from 2500 m to the highest peaks (above 3400 m). Traditionally, the main threat has been the harvest of complete individuals for medicinal purposes; also ungulates (wild and domestic ones) browse a large percentage of the reproductive stems, despite the production of sesquiterpenes that make the foliage bitter (Watson & al., 2002). The result of these pressures is a major decrease in seed set (decreasing 20-90 % of the total seed set, depending on the population; author’s unpublished data). Inflorescence and flower morphological data A total of 18 mature inflorescences (heads/capitula) were randomly collected from each population (AG1 and AG2). The heads were dissected under a bifocal magnifying glass and the number of female and hermaphrodite flowers per capitulum was counted. A subsample of 25 mature hermaphrodite flowers were randomly taken from these 18 inflorescences, and stamen, ovary and style length were measured using a digital calliper (± 0.001). Data were analysed by means of one-way ANOVA. Pollen morphological analysis and settling speed Samples of 30 anthers (15 per population) from 30 individuals were randomly selected and collected before anthesis and acetolysed using the method of Erdtman (1960) as modified by Hideux (1972), and then mounted in glycer-gelatin for light microscopy. The pollen terminology used is based on Punt & al. (2007). For scanning electron microscopy (SEM), non-acetolysed grains were dehydrated in an alcohol series, pipetted onto a SEM stub in a few drops of 100% alcohol, and allowed to dry. Samples were coated with gold/palladium and examined using a SEMmicroscopy. In parallel, the principal features of individual pollen grains (emphasizing diameters i.e. polar axis and equatorial diameter) were measured using a light microscope at a magnification of 400× in order to determinate their characteristic dimensions and shapes. Then, the diameters measures (i.e. the major and minor diameters P and E) were used to calculate the volume-equivalent sphere of the pollen grain given by 3PE2. Taking into account the latter measure both the “theoretical settling speed” (Fuchs, 1964; Anales del Jardín Botánico de Madrid 68(1): 97-105, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2276 2276_Artemisa:Anales 68(1).qxd 13/06/2011 12:22 Página 100 100 J. Peñas & al. Leith, 1987), as well as, the “theoretical residence time” of A. granatensis pollen (see Chatigny & al. (1979) for further information about residence times estimation) assuming dry deposition were calculate. The settling speed (Vt) of a pollen grain falling in still air at a constant temperature and pressure is described by Fuchs (1964). Taking into account the volume-equivalent diameter (d) of the pollen grains, the Vt of Artemisia pollen was calculated based on Stokes’ Law for a sphere (in the absence of electrostatic forces). This equation calculates the settling speed as a function of particle size, particle density, acceleration due to gravity, and the density and viscosity of the air. Stokes’ Law Equations: solving for settling speed or terminal velocity gd2(ρp - ρm) 18µ Inputs: acceleration of gravity (g); particle diameter (d); density of particle (ρp); density of medium (ρm); viscosity of medium (µ). Vt = Pollen-tube growth and compatibility system Pollinations were achieved by rubbing a dissection needle against the anthers of pollen-bearing flowers and then against the stigmas of outer female flowers of the same head (self-crosses, SC) or the female flowers in the head of a different individual in the same population (intra-population outcrosses, IPC) or against those of a different head in the other population (AG1 and AG2) i.e. inter-population outcrosses (InPC). All hand-pollinated heads were bagged with cellophane bags. Some naturally pollinated heads (no hand-pollinated, and no bagged) were collected to measure natural tube growth (no hand-pollinated NP). Prior to hand pollination stylar arms were observed with a magnifying glass to ensure that the arms were fully expanded (mature) and had no pollen on them. We collected five capitula per population and placed individually in cellophane bags in order to use as donors in the inter-population outcrosses (lnPC), the time passed between collection and hand-pollination was two to three hours, roughly. Afterwards all hand-pollinated (SC, IPC, InPC) flowers were collected (24h-48 h after) and fixed in 70% ethanol. After rinsing in distilled water, isolated gynoecia were softened and cleared in 8 mol/l NaOH for 48-72 h. Softened gynoecia were placed in distilled water for at least 1 hour before staining with a 0.1% solution of aniline blue in 0.1 mol/L K3PO4 for 12 h (as described in Martin, 1959). Each gynoecium was examined under UV light (range 360-390 nm for selective excitation of DAPI fluorescence) to observe tube growth. Pollen-tube growth was observed of selfs and crosses at the stigma, and along the style. Pollinations were classified as SI or SC based on whether pollen tubes reached the base of the style or not. Pollen-tube growth was compared using the non-parametric X2 test. Results Floral morphology (Table 1) Artemisia granatensis capitula had around 80 flowers each. Heads from plants in AG1 and AG2 populations showed no differences in hermaphrodite and female flower number. There were no significant differences in total flowers/head, hermaphrodite flowers/heads, female flowers/head and natural fruitset/head between populations, although the natural fruit:flower ratio was higher in AG2. We found no differences between morphological traits of reproductive structures between AG1 and AG2 (Table 2) with the exception of mature stamen length, which was higher for AG2 flowers (Table 2). Table 1. Mean (SD) of inflorescence traits gathered from Artemisia granatensis populations (AG1 and AG2) (n = 18 inflorescences). AG1 AG2 F-ratio P Total flowers/ head 74.0 (19.1) 83.1 (30.3) F (1, 35) = 0.877 0.356 n.s. Hermaphrodite flowers/ head 70.7 (18.1) 74.2 (30.5) F (1, 35) = 0.701 0.408 n.s. Female flowers/ head 5.4 (3.7) 6.8 (3.7) F (1, 35) = 0.573 0.454 n.s. Fruits/ head 0.4 (2.2) 2.1 (1.0) F (1, 35) = 4.108 0.051 n.s. 0 2.29% Fruit:flower ratio Mean (SD) values for floral traits and fruit set of n.s.: not significant. Anales del Jardín Botánico de Madrid 68(1): 97-105, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2276 2276_Artemisa:Anales 68(1).qxd 13/06/2011 12:22 Página 101 Artemisia granatensis 101 Table 2. Mean (SD) values of female and male floral traits of Artemisia granatensis flowers at AG1 and AG2 populations (n = 25). AG1 AG2 Anova Results F P Ovary length (mm) 0.88 (0.12) 0.87 (0.09) F (1,405) = 0.330 0.569 n.s. Style length (mm) 1.50 (0.32) 1.59 (0.26) F (1,405) = 1.091 0.302 n.s. Style arms length (mm) 0.74 (0.12) 0.77 (0.12) F (1,405) = 0.277 0.601 n.s. Stamen length (mm) 2.12 (0.16) 2.35 (0.23) F (1,405) = 5.447 0.035 * Mean (SD) values for floral traits of * = significant, n.s.: no significant. Pollen morphology and settling speed A. granatensis pollen has the typical anemophilous syndrome, based on morphological features and settling speed. It is isopolar, with radial symmetry. In polar view it is circular-lobate (Fig. 1a), and in equatorial view it is circular-elliptical (Fig. 1b). It is spheroidal or prolate-spheroidal, with a P:E ratio of 1.0, small to medium in size, the length of the polar axis (P) being 18-20 (19.2 ± 0.8) µm and the equatorial diameter (E) being 17-22 (19.3 ± 1.3) µm. The pollen grain is trizonocolporate, fossaperturate. The ectoaperture is a colpus 12-16 (13.7 ± 1.6) µm long; the endoaperture is a circular or lalongate porous 1.5-3 (1.9 ± 0.5) µm long. The surface relief of the pollen grain is microechinate, ornamentation of spines shorter than 1 µm, (c. 0.7 µm), showing delicate verrucate surface sculpturing, broader than high and less than 1 µm in diameter (around 0.24 µm). The resulting volume-equivalent diameter of A. granatensis pollen is c. 19.3 µm. Known this measure and under the assumption of a dry deposition as well as a density of 1 g/cm3, the theoretical settling speed or terminal velocity (Vt) was calculated at around 1.18 cm/s (or 42.48 m/h); taking into account the above parameter, the resulting theoretical residence time in the atmosphere was 0.6 days (c. 14 h). AG2 plants (Fig. 2), signifying a rejection of self pollination and sporophytic self-incompatibility. The X2 test showed that SC produced a significantly lower a b Pollen-tube growth and compatibility system Self-incompatibility was determined based on pollen-tube growth inhibition. The inhibition of incompatible pollen occurred at the stigma surface, where grains either failed to germinate, or the emerging pollen tube was usually inhibited before penetrating the stigma surface. Compatible pollen grains produced pollen tubes growing through the style. Viability tests showing full fluorochromatic reaction demonstrated that A. granatensis pollen collected from populations AG1 and AG2 was viable. Self crosses (SC) produced failed to germinate, or did not penetrate in the stigma both for AG1 and Fig. 1. View of Artemisia granatensis pollen grain (SEM microscopy). a, whole grain: polar view showing three apertures; b, whole grain: meridional view showing two apertures and details of spines. Anales del Jardín Botánico de Madrid 68(1): 97-105, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2276 2276_Artemisa:Anales 68(1).qxd 13/06/2011 12:22 Página 102 102 J. Peñas & al. Discussion No-growth pollen-tube Fig. 2. Inhibition of pollen tube at the stigmas of Artemisia granatensis for a SC cross in AG2. number of gynoecia with pollen tubes than did IPC in both AG1 and AG2 (Table 3). The number of gynoecia with pollen tubes was higher for IPC than for SC in both AG1 and AG2. (Table 3; Fig.3). The X2 test for the comparison of NP and IPC in AG1 and AG2 showed no significant differences with respect to the number of gynoecia with pollen tubes (Table 3). InPC showed a high number of gynoecia with pollen tubes (Table 3; Fig. 4) and also a higher number of gynoecia with pollen tubes than NP in both AG1 and AG2 (Table 3). Artemisia granatensis, as with many Asteraceae, has a gynomonoecious sexual system, in which female and bisexual flowers occur in the same inflorescence (capitulum). The isolated A. granatensis populations studied showed no significant differences between morphological traits. Flower number per head and the ratio of hermaphrodite to female flowers showed no differences between AG1 and AG2. Additionally, stamen, ovary, and style sizes were comparable. Previous reports in some Asteraceae showed that flowersize differences could be related to the loss of self-incompatibility in small populations (Ortiz & al., 2006), and this trend has also been found for another selfincompatible species in fragmented habitats (Taisma & Varela, 2005). Our results indicate that AG1 and AG2 plants have no morphological differences and, therefore, these traits could not be related to differential self-incompatibility expression. Pollen-tube growth after controlled hand pollination in natural populations of A. granatensis showed that the species has a sporophytic self-incompatibility system with no evidence of increased self-incompatibility or partial-self incompatibility as has been found in small, isolated and peripheral populations (Fausto & al., 2001; Vallejo-Marin & Uyenoyama, 2004; Taisma & Varela, 2005). These results agree with findings for other members of Asteraceae and suggest that, although population size is small and isolation is high, there is no breakdown of the self-incompatibility system. These data support the idea that the main constraint on reproductive success in A. granatensis under natural conditions (Fig. 5) is the reduced number of compatible mates due to failure in wind pollination. Table 3. Results of pollination treatments in Artemisia granatensis individuals in populations AG1 and AG2. Cross Type AG1 AG2 n Number of gynoecia with growing tubes Number of gynoecia without growing tubes X2 critical value (1gl; p < 0.01 = 6.63) SC 72 0 72 SC vs. IPC = 77.88 * IPC 6 6 0 InPC 66 56 10 SC vs. InPC = 11.25 * NP 29 8 21 InPC vs. NP = 2.03 n.s. SC 122 0 122 SC vs. IPC = 20.69 * IPC 104 26 78 InPC 48 16 32 SC vs. InPC = 33.49 * NP 48 7 41 InPC vs. NP = 4.62 n.s. SC = self crosses, IPC = intra-population outcrosses, InPC = inter-population outcrosses, NP = no hand-pollinated, natural tube growth; n.s.: not significant. Anales del Jardín Botánico de Madrid 68(1): 97-105, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2276 2276_Artemisa:Anales 68(1).qxd 13/06/2011 12:22 Página 103 Artemisia granatensis 103 Stigma Style Pollen tubes Pollen tubes Stylar arm Fig. 3. Growing pollen tubes at the style after a IPC cross in AG1. Fragmentation of wind-pollinated populations seriously reduces pollen availability, limiting reproduction (Knapp & al., 2001). Davis & al. (2004) found 9fold more pollen on stigmas of high-density Spartina alterniflora plants than on those occurring at low density; they also found that the consequences of loss of appreciable numbers of seed caused by pollen limitation persists for decades. The low natural fruit set of A. granatensis (Hernández-Bermejo & al., 2003) and the low number of gynoecia with viable pollen tubes after NP (natural tube growth) agree with the expected reduced seed-set in small populations of self-incompatible, wind-pollinated species (Widen, 1993, Lienert & Fisher, 2003; Davis & al., 2004). A critical event for A. granatensis reproduction could be pollen deposition, because this species is self-incompatible in very small isolated populations, and thus compatible pollen flow could be a critically limiting condition for fruit set. In this sense, further studies addressing pollen limitation in relation with population size and isolation are needed. Fig. 4. Growing pollen tubes at the stigma, stylar arms and style after a InPC cross in AG2. Transport and dispersal of pollen grains by the moving atmosphere as well as their residence time as airborne particles are strongly linked to physical atmospheric characteristics at their time of flight (Comtois & al., 2000). The settling speed and residence time estimated for A. granatensis pollen can vary greatly in nature (Sierra Nevada) owing to turbulence and atmospheric humidity, which can alter the density of biological particles (Aylor, 2002); including topographic barriers against pollen displacement. However, a cornerstone parameter in any future model of pollen transport in the atmosphere is calculating the gravitational settling speed in still air, Vt, of individual pollen grains, because Vt largely determines both the distance of travel and the efficiency of deposition on target organs (Aylor & al., 2005). According to Kohler & al. (2007) this theoretical information is an essential parameter for reliably modelling the atmospheric dispersal of pollen in situ. Despite this, very little is Anales del Jardín Botánico de Madrid 68(1): 97-105, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2276 2276_Artemisa:Anales 68(1).qxd 13/06/2011 12:22 Página 104 104 J. Peñas & al. Stigma Pollen tubes Style required for acquiring a thorough knowledge the reproductive biology and pollination ecology in the case of A. granatensis. In any case, it seems obvious that A. granatensis reproductive efficiency has been critically affected by the devastation of natural population due to human use. Current population size and plant densities may be a serious limitation to guarantee enough pollen from compatible mates. Additionally, present populations are isolated by major geographic barriers that could also seriously limit compatible pollen flow between populations. Programmes for the recovery of A. granatensis may need to include reintroduction of compatible mates in order to enhance pollen flow and fruit-set efficiency by means of connecting isolated patches. Acknowledgements Fig. 5. Growing pollen tubes at the stigma and style level after NP in AG2. known about A. granatensis pollen mobility in the atmosphere. As far as we know, this study is the first to determine physical characteristics of A. granatensis pollen, which can contribute to defining the main parameters involved in reproductive biology in this species. The model equations for settling speed presented here offers a means for evaluating dispersal potential for a range of environmental conditions. Our results suggest the potential distance that A. granatensis pollen can be transported during the estimated residence time (14 h) was 600 m. According to Mandrioli (1998) small or medium particles, such as A. granatensis pollen, have a relatively long or medium residence time in the atmosphere, supporting the hypothesis of intra- and inter-populational pollination success. Nevertheless, natural pollen-tube growth suggests a pollen-deposition limitation, probably related to the viability and longevity of pollen grain as well as pollen dispersion capacity, both intimately linked to the likelihood of reproductive success (Mandrioli, 1998). These facts suggest that the reproductive success depends on the time spent by the viable pollen grain to reach the nearest population and the inter-population range. Obviously, further details about pollen flow within and between populations is We wish to thank E. Rico (University of Salamanca) for valuable comments on the manuscript, and we thank B. Forot, B. Benito and C. Ruiz Rejón (University of Granada) for their contribution in the field and laboratory work. The authors are indebted to David Nesbitt for linguistic advice. This work has been partly financed by the Spanish Education and Science Ministry (project reference REN2003-09427-C02), and partly by the Consejería de Innovación, Ciencia y Tecnología de la Junta de Andalucía (project reference P05-RNM1067). Science Faculty of Universidad Central de Venezuela supported Dra. Taisma’s research at the University of Granada. References Aylor, D.E. 2002. Settling speed of corn (Zea mays) pollen. Aerosol Science 33: 1601-1607. Aylor, D.E., Baltazar, B.M., Schoper & J.B. 2005. Some physical properties of teosinte (Zea mays subsp. parviglumis) pollen. Journal of Experimental Botany 56 (419): 2401-2407. Bañares, A., Blanca, G., Güemes, J., Moreno, J.C. & Ortiz, S. (eds.). 2003. Atlas y libro rojo de la flora vascular amenazada de España. Madrid. Blanca, G. & Valle, F. 1991. Las plantas endémicas de Andalucía Oriental, IV: Artemisia granatensis. Monografias de Flora y Vegetación Béticas 6: 23-27. Blanca, G. (ed.). 2002. Flora amenazada y endémica de Sierra Nevada. Granada. Blanca, G., Cabezudo, B., Hernández-Bermejo, E., Herrera, C. M., Molero-Mesa, J., Muñoz, J. & Valdés, B. 1999. Libro Rojo de la Flora Silvestre Amenazada de Andalucía. Tomo I: Especies en Peligro de Extinción. Sevilla. Blanca, G., Cueto, M., Martínez-Lirola, M.J. & Molero, J. 1998. Threatened vascular flora of Sierra Nevada (Southern Spain). Biological Conservation 85: 269-285. Blondel, J. & Aronson, J. 1999. Biology and wildlife of the Mediterranean region. Oxford. Anales del Jardín Botánico de Madrid 68(1): 97-105, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2276 2276_Artemisa:Anales 68(1).qxd 13/06/2011 12:22 Página 105 Artemisia granatensis BOE (Boletín Oficial del Estado de España). 1990. Real Decreto 439/1990, de 30 de Marzo, por el cual se regula el Catálogo Nacional de Especies Amenazadas. Madrid. Cabezudo, B., Talavera, S., Blanca, G., Salazar, C., Cueto, M., Valdés, B., Hernández-Bermejo, E. & Herrera, C. M. 2005. Lista roja de la flora vascular de Andalucía. Sevilla. Charlesworth, D. 1985. Distribution of dioecy and self-incompatibility in angiosperms. In: Greenwoog, P.J., Harvey, P.H. & Slatkin, M. (eds.), Evolution: essays in honor of John Maynard Smith:237-268. Cambridge University Press. Cambridge. Comtois, P., Fernández-González, D., Valencia-Barrera, R.M., Sánnchez, J.L., Fraile, R. & Rodier, S. 2000. Pollen content study of the lower atmosphere in León (Spain) by use of a tethered balloon. Aerobiologia 16: 187-191. Davis, H.G., Taylor, C. M., Lambrinos, J.G. & Strong, D.R. 2004. Pollen limitation causes an Allee effect in a wind-pollinated invasive grass (Spartina alterniflora). Proceedings of the National Academy of Sciences of The United States of America 101(38): 13804-13807. Dominguez Lozano, F., Galicia Herbada, D., Moreno Rivero, L., Moreno, J.C. & Sainz, H. 1996. Threatened plants in Peninsular and Balearic Spain: A report based on the EU Habitats Directive. Biological Conservation 76: 123-133. Erdtman, G. 1960. The acetolysis method - a revised description. Svensk Botanisk Tidskrift 54: 561-564. Fausto, J.A., Eckhart, V.M. & Geber, M.A. 2001. Reproductive assurance and the evolutionary ecology of self-pollination in Clarkia xantiana (Onagraceae). American Journal of Botany 88(10): 1794-1800. Fay, M.F. 1992. Conservation of rare and endangered plants using Pollen in vitro. In Vitro Cellular & Developmental Biology – Plant tubes 28(1): 1-4. Fuchs, NA. 1964. The mechanics of aerosols. Macmillan Publishing Company. New York. Gibbs, P.E., Milne, C., & Vargas-Carrillo, M. 1975. Correlation between the breeding system and recombination index in five species of Senecio. New Phytologist 75: 619-626. Gómez, A. 2002. Geomorphological map of Sierra Nevada; glacial a periglacial geomorphology. Sevilla. Gómez, A., Palacios, A., Ramos, M., Tanarro, L.M., Schulte, L. & Salvador, F. 2001. Location of Permafrost in Marginal Regions: Corral del Veleta, Sierra Nevada, Spain. Permafrost Periglacial Processes 12: 93-110. Hernández-Bermejo, E., Contreras, P., Clemente, M. & Prados, J. 2003. Artemisia granatensis Boiss. In: Bañares, A., Blanca, G., Güemes, J., Moreno, J.C. & Ortiz, S. (eds.), Atlas y libro rojo de la flora vascular amenazada de España. Pp: 124-125. Madrid. Hideux, M. 1972. Techniques d’étude du pollen au MEB: effects comparés des differents traitments physicochimiques. Micron 3: 1-31. IUCN. 2001. IUCN Red List Categories and Criteria: Version 3.1. IUCN species survival Commission. Gland and Cambridge. Knap, E.E., Goedde, M.A. & Rice, K.J. 2001. Pollen-limited reproduction in blue oak:implications for wind pollination in fragmented populations. Oecologia 128: 48-55. Kohler, F., Schultz, E. & Helm, H. 2007. Study of morphology and settling velocity of airborne pollen captured in a Paul trap. Abstract European Aerosol Conference 2007, Salzburg, Koul, M. & Bhatnagar, A.K. 2007. Plant Reproductive biology studies crucial for conservation. Current Science 92(9): 1207. Leith D. 1987. Drag on non-spherical objects. Aerosol Science and Technology 6: 153-161. 105 Lienert, J. & Fisher, M. 2003. Habitat fragmentation affects the common wetland specialist Primula farinosa in north-east Switzerland. Journal of Ecology 91(4): 587-599. Mandrioli, P. 1998. Basic aerobiology. Aerobiologia 14: 89-94. Martin, F.W. 1959. Staining and observing pollen tubes by means of fluorescence. Stain Technology 34: 125-128. Martin, J., Torrell, M. & Valles, J. 2001. Palynological features as a systematic marker in Artemisia s.l. and related genera (Asteraceae, Anthemideae): implications for subtribe Artemisiinae delimitation. Plant Biology 3(4): 372-378. Médail, F & Diadema, K. 2009. Glacial refugia influence plant diversity patterns in the Mediterranean basin. Journal of Biogeography 36: 1333-1345. Molero, J. 1994. Baetic and Sub-Baetic Mountains, Spain. In: Davis, S.D., Heywood, V.H. & Hamilton, A.C. (eds.), Centres of plant diversity. Europe, Africa, South West Asia and the Middle East: 55-58. IUCN Publication Unit. Cambridge. Moreno, J.C. (coord.). 2008. Lista Roja 2008 de la flora vascular española. Dirección General de Medio Natural y Política Forestal (Ministerio de Medio Ambiente, y Medio Rural y Marino, y Sociedad Española de Biología de la Conservación de Plantas). Madrid. O’Brien, M.H. 1980. The Pollination Biology of a Pavement Plain: Pollinator Visitation Patterns. Oecologia 47: 213-218. Ortiz, M.A., Talavera, S., García-Castaño, J.L., Tremetsberger, K., Stuessy, T., Balao, F. & Casimiro-Soringuer, R. 2006. Self-incompatibility and floral parameters in Hypochaeris sect. Hypochaeris (Asteraceae). American Journal of Botany 93: 234-244. Peñas, J., Pérez-García, F. & Mota, J.F. 2005. Patterns of endemic plants and biogeography of the Baetic high mountains (south Spain). Acta Botanica Gallica 152(3): 247-360. Punt, W., Hoen, P.P., Blackmore, S., Nilsson, S. & Le Thomas, A. 2007.Glossary of pollen and spore terminology. Review of Palaeobotany and Palynology 143(1-2): 1-81. Taisma, M.A. & Varela, C.W. 2005. Sistema de compatibilidad en la especie distilica Cordia curassavica (Jacq.) R&S (Boraginaceae). Interciencia 30(7): 431-435. Thompson, J.D. 2005. Plant evolution in the Mediterranean. Oxford University Press. Oxford. Vallejo-Marin, M. & Uyenoyama, M.K. 2004. On the evolutionary costs of self-incompatibility : incomplete reproductive compensation due to pollen limitation. Evolution 58(9): 1924-1935. Watson, L.E., Bates, P.L., Evans, T.M., Unwin, M.M. & Estes, J.R. 2002. Molecular phylogeny of Subtribe Artemisiinae (Asteraceae), including Artemisia and its allied and segregate genera. BCM Evolutionary Biology 2(17): 1-12. Weekly, C.W. & Race, T. 2001. The breeding system of Ziziphus celata Judd and D.W. Hall (Rhamnaceae), a rare endemic plant of the Lake Wales Ridge, Florida, USA: Implications for recovery. Biological Conservation 100(2): 207-213. Weller, S.G. 1994. The relationships of rarity to plant reproductive biology. In: Bowles, M.L. & Whelan, C.J. (eds.), Restoration of endangered species: 90-117. Cambridge University Press. Cambridge. Widen, B. 1993. Demographic and genetic effects on reproduction as related to population size in a rare perennial herb, Senecio integrifolius (Asteraceae). Biological Journal of the Linnean Society 50(3): 179-195. Anales del Jardín Botánico de Madrid 68(1): 97-105, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2276 Associate Editor: C. Herrera Received: 10-XI-2010 Accepted: 28-II-2011 2276_Artemisa:Anales 68(1).qxd 13/06/2011 12:22 Página 106 2245_Baccharis:Anales 68(1).qxd 13/06/2011 12:14 Página 107 Anales del Jardín Botánico de Madrid Vol. 68(1): 107-116 enero-julio 2011 ISSN: 0211-1322 doi: 10.3989/ajbm.2245 Microsculpture of cypselae surface of Baccharis sect. Caulopterae (Asteraceae) from Brazil by Angelo Alberto Schneider & Ilsi Iob Boldrini Programa de Pós-Graduação em Botânica, Instituto de Biociências, Universidade Federal do Rio Grande do Sul, Campus do Vale, prédio 43433, 91501-970, Porto Alegre, Rio Grande do Sul, Brazil Corresponding author: [email protected] Abstract Resumen Schneider, A.A. & Boldrini, I.I. 2011. Microsculpture of cypselae surface of Baccharis sect. Caulopterae (Asteraceae) from Brazil. Anales Jard. Bot. Madrid 68(1): 107-116. Schneider, A.A. & Boldrini, I.I. 2011. Microescultura de la superficie de las cipselas de Baccharis sect. Caulopterae (Asteraceae) de Brasil. Anales Jard. Bot. Madrid 68(1): 107-116 (en inglés). The aim of this study was to characterize the microsculpture of the cypselae surface of the Brazilian species of Baccharis L. sect. Caulopterae DC. (Asteraceae), and to compare this data it with the taxonomy of the group. Scanning electron microscopy was used to examine the cypsela surface of 25 taxa of Baccharis sect. Caulopterae from Brazil. According to the micromorphology of the cypsela surface, the species can be classified into five distinct groups. The cypselae of the species of the Baccharis trimera species complex (B. crispa, B. cylindrica, B. jocheniana, B. myriocephala, and B. trimera) share the same micromorphological features. Para examinar la superficie de cipselas de 25 táxones de Baccharis L. sect. Caulopterae DC. de Brasil se ha utilizado la microscopía electrónica de barrido. El objetivo del estudio fue caracterizar la microescultura de la superficie de cipselas de la sección y colaborar con la delimitación taxonómica a nivel específico. Las especies fueron clasificadas en cinco grupos distintos según la micromorfología y asignados a la terminología existente. El complejo Baccharis trimera (B. crispa, B. cylindrica, B. jocheniana, B. myriocephala y B. trimera) mostró afinidades micromorfológicas de las cipselas. Keywords: achene, carpology, carqueja, Compositae, micromorphology, SEM, taxonomy. Palabras clave: aquenio, carpología, carqueja, Compositae, MEB, micromorfología, taxonomía. Introduction fraspecific taxa included in the section is variable in the literature, especially due to different taxonomic concepts (Heiden & al., 2009). The section is restricted to South America, occurring extensively in the Andes, from Colombia to the Central Argentina, and in Brazil, where the highest number of species in this section is concentrated in the south and southwest (Barroso, 1976; Müller, 2006). Velez (1981) studied the American genera of Astereae and provided a detailed overview of cypsela morphology and anatomy in the genus Baccharis. Other studies that have contributed to our knowledge of cypsela morphology in this genus are Ariza (1973), Hellwig (1990), Mukherjee & Sarkar (2001) and Müller (2006). The latter author reviewed cypsela characteristics for the genus, and present data with re- The fruits of the Asteraceae, denominated cypselae, are dry, indehiscent, unilocular, with a single seed that is usually not adnate to the pericarp (linked only by the funicle) and originating from an inferior ovary (Marzinek & al., 2008). The pappus, a modified calyx, is inserted in the apical region of the cypsela, whilst basally, an abscission region is located in relation to the inflorescence axis (clinanthium) denominated the carpopodium (Roth, 1977). Cypsela microsculpture analysis has been considered more and more a taxonomic tool, being also important for higher and medium level classification within the family (Bremer, 1994; Anderberg, 1991). Baccharis sect. Caulopterae is represented by around 35 species, but the number of species and in- 2245_Baccharis:Anales 68(1).qxd 13/06/2011 12:14 Página 108 108 A.A. Scheneider & I.I. Boldrini gard to form, shape, colour, indumentum, number of vascular bundles encircling the fruits (ribs), details of the pericarp, and some general considerations concerning microsculpture of the cypsela cuticle. Here we characterize the microsculpture of the cypselae surface of the Brazilian species of genus Baccharis sect. Caulopterae, and we were particularly interested to see if cypsela characters could help resolve the taxonomy of the B. trimera complex, comprising B. crispa, B. cylindrica, B. jocheniana, B. myriocephala, and B. trimera (Barroso 1976)". Material and methods Scanning Electron Microscopy Mature cypselae of 25 Brazilian species of Baccharis sect. Caulopterae were collected from herbarium specimens (HBR, ICN, PACA and UEC - see Table 1). One representative dry cypsela was then selected per species and mounted on a metallic stub using a carbon adhesive tape and sputter-coated with 20 nm gold using BAL-TEC SCD-050. Electromicrographs of the cypselae were obtained under 10 kV in magnification using a Scanning Electron Microscope (SEM) JEOLJSM 6060 at the Centro de Microscopia Eletrônica (CME) of the Universidade Federal do Rio Grande do Sul. Analyses and terminology The characters we recorded were based on analyses of morphology of the secondary and tertiary sculpture of the surface morphology: smooth or folded, presence or absence of papillae, degree of surface rugosity and presence of cavities. The primary sculpture was not examined since in most species it was lacking. The terminology used for cypselae shape (solid structure) follows Radford (1986), and microsculpture characterization was based on Barthlott (1981; 1990), Mukherjee & Sarkar (2001) and Müller (2006). Baccharis burchellii Baker and B. regnellii Sch. Bip. ex Baker, although belonging to Baccharis sect. Caulopterae and occurring in Brazil were not included in this study because we could not obtain material with mature cypselae. The nomenclature of species follows recent works, and the following names were considered: B. pentaptera (Less.) DC. (=B. stenocephala Baker, according to Schneider & al., 2009), B. sagittalis (Less.) DC. (=B. heeringiana Malag., =B. macroptera D.J.N. Hind), B. subtropicalis Heiden (=B. sagittalis var. montevidensis Baker), and B. junciformis (Less.) DC. (=B. usterii Heering, all according to Heiden & al., 2009). For species belonging to the Baccharis trimera species complex (B. crispa Spreng., B. cylindrica (Less.) DC., B. myriocephala DC. and B. trimera (Less.) DC.) we followed the circumscription proposed by Barroso (1976). Results The cypselae of Baccharis sect. Caulopterae can be divided into three groups with regard to the number of ribs: 18 species with 5-7 ribs; six with ~14 ribs (B. crispa, B. cylindrica, B. jocheniana, B. myriocephala, B. aff. opuntioides, B. trimera); one species with ~20 ribs (B. riograndensis). Cypsela shape varies from narrowly oblong, narrowly oblong to cylindric, narrowly oblong to obovoid, oblanceoloid, oblong and obovoid form (Table 1). The cypselae surface presents a variable microsculpture, mostly with a folded cuticle, but also rugose with the presence of cavities in two species: B. articulata and B. glaziovii. Most species are papillose with digitiform (B. trimera) or globose papillae (B. articulata and B. sagittalis), with length ranging from 220 µm by 2-5.5 µm in diameter, with variable distribution on the cypselae surface but they are longer on the vascular bundles (ribs). B. heeringiana, B. pseudovillosa, B. ramboi, B. riograndensis, B. stenocephala, B. usterii and B. vincifolia did not present papillae. A ringshaped carpopodium was observed in all species, and its diameter ranges from 50-120 µm. The SEM study of the cypselae provided important characters which allowed us to distinguish five groups among the sampled species. GROUP I. Papillose rugose cypselae In this group, the cypselae present an evenly distributed rugose surface with digitiform papillae. The papillae are longer on the ribs. Carpopodium present, ring-shaped, 90-120 µm diameter. Ribs 5-7. Two species: B. microcephala (Fig. 1A-D) and B. penningtonii (Fig. 1E-H). GROUP II. Epapillose rugose cypselae Cypselae present a rugose surface, but papillae are absent, or just relictual and slightly salient on the ribs. Cypsela surface slightly folded and with delicate and irregular rugosities. Carpopodium present, ringshaped, 80-110 µm diameter. Ribs 5-7. Five species: B. palustris (Fig. 2A-D), B. paranensis (Fig. 2E-H), B. pseudovillosa (Fig. 2I-L), B. ramboi (Fig. 2M-P) and B. vincifolia (Fig. 2Q-T). Anales del Jardín Botánico de Madrid 68(1): 107-116, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2245 2245_Baccharis:Anales 68(1).qxd 13/06/2011 12:14 Página 109 Microsculpture of cypselae of Baccharis 109 Table 1. Relation of studied species and some characters analyzed with respective vouchers: length (L), diameter (D); approximate (~). Taxon Shape Size ~(length × L/D Ribs (~) Source width, mm) B. apicifoliosa A.A.Schneid. & Boldrini oblanceloid 1.29 × 0.41 3/1 7 R. Wasum 802 (PACA) obovoid 0.74 × 0.38 2/1 5 L.A. Mentz s.n. (ICN 59169) B. crispa Spreng. narrowly oblong 1.27 × 0.38 3/1 14 I. Fernandes 641 (ICN) B. cylindrica (Less.) DC. narrowly oblong 1.38 × 0.32 4/1 14 R. Schmidt s.n (ICN 153106) B. flexuosiramosa A.A.Schneid. & Boldrini narrowly oblong 1.37 × 0.46 3/1 7 C.F. Jurinitz s.n. (ICN 153107) B. glaziovii Baker narrowly oblong 1.02 × 0.37 3/1 5 J. Mattos 15953 ( UEC) B. jocheniana G. Heiden & L. Macias narrowly oblong 1.24 × 0.37 3/1 14 B. junciformis (Less.) DC. narrowly oblong 1.12 × 0.32 4/1 5 K.D. Barreto & G.D. Fernandes 752 (ESA) B. microcephala (Less.) DC. narrowly oblong 1.27 × 0.40 3/1 7 J. Dutra 1488 (ICN) B. milleflora (Less.) DC. narrowly oblong 1.28 × 0.42 3/1 7 B. Rambo 49318 (PACA) B. myriocephala DC. narrowly oblong 1.16 × 0.29 4/1 14 A.A. Schneider 1161 (ICN) B. aff. opuntioides Mart. ex Baker narrowly oblong 1.40 × 0.38 4/1 14 A.A. Schneider 1326 (ICN) B. organnensis Baker narrowly oblong 1.26 × 0.34 4/1 7 A. Sehnem 5119 (PACA) B. palustris Heering narrowly oblong 1.14 × 0.43 3/1 7 B. Rambo 52024 (HBR) B. paranensis Dusén narrowly oblong 1.46 × 0.34 4/1 7 J. Iganci 507 (ICN) B. penningtonii Heering narrowly oblong 1.00 × 0.31 3/1 7 J.C. Sacco 808 (PACA) B. pentaptera (Less.) DC. oblong 1.73 × 0.70 2/1 7 A.A. Schneider 1261 (ICN) B. phyteumoides (Less.) DC. narrowly oblong 1.01 × 0.38 3/1 7 A.A. Schneider 1586 (ICN) B. pseudovillosa Malag. & J.E. Vidal narrowly oblong 1.73 × 0.34 5/1 7 O. Camargo 2843 (PACA) B. ramboi G. Heiden & L. Macias narrowly oblong 1.19 × 0.28 4/1 7 A.A. Schneider 1282 (ICN) B. riograndensis Malag. & J.E. Vidal narrowly oblong-cylindric 2.55 × 0.37 7/1 20 B. sagittalis (Less.) DC. narrowly oblong 1.00 × 0.29 3/1 7 A.A. Schneider 1296 (ICN) obovoid 0.76 × 0.42 2/1 5 M. Sobral & al. 5031 (ICN) B. trimera (Less.) DC. narrowly oblong 1.19 × 0.30 4/1 14 B. vincifolia Baker narrowly oblong-obovoid 1.37 × 0.42 3/1 5 B. articulata (Lam.) Pers. B. subtropicalis G. Heiden GROUP III. Papillose folded cypselae without cavities In this group, the cypselae present a folded surface with evenly distributed papillae. Longer papillae on the ribs, the number, concentration, and shape of papillae vary in each species. Carpopodium present, ring-shaped, 40-110 µm diameter. This group pre- A.A. Schneider 1267 (ICN) L.T. Pereira 16 (ICN) L.T. Pereira 87 (ICN) B. Rambo 60054 (PACA) sents 12 species and can be divided in two subgroups by the number of ribs: a. with 5-7 ribs consists of B. apicifoliosa (Fig. 3A-D), B. flexuosiramosa (Fig. 3EH), B. milleflora (Fig. 3I-L), B. organensis (Fig. 3M-P), B. phyteumoides (Fig. 3Q-T) and B. sagittalis (Fig. 4AD); b. with ~14 ribs is composed of B. crispa (Fig. 4EH), B. cylindrica (Fig. 4I-L), B. jocheniana (Fig. 4M-P), Anales del Jardín Botánico de Madrid 68(1): 107-116, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2245 2245_Baccharis:Anales 68(1).qxd 13/06/2011 12:14 Página 110 110 A.A. Scheneider & I.I. Boldrini Fig. 1. Scanning electron micrographs of cypselae surface of Baccharis sect. Caulopterae - Group I: B. microcephala: A, cypsela; B, C, papillae detail; D, carpopodium. B. penningtonii: E, cypsela; F, G, papillae detail; H, carpopodium enlarged. B. myriocephala (Fig. 4Q-T), B. aff. opuntioides (Fig. 5A-D), and B. trimera (Fig. 5E-H). GROUP IV. Papillose folded cypselae with cavites The cypselae present a folded surface with evenly distributed papillae. The papillae are globose (B. articulata) or cylindrical and longer on the ribs than in the intercostal region (B. glaziovii). Slight cavities occur on the folded surface. Carpopodium present, ring-shaped, 20-60 µm diameter. Ribs ~ 5. Two species: B. articulata (Fig. 6A-D) and B. glaziovii (Fig. 6E-H). GROUP V. Epapillose folded cypselae The cypselae present a folded surface, with longitudinal folds. Papillae are absent in the intercostal areas, or just relictual, and slightly prominent on the ribs. Carpopodium present, ring-shaped, 50-130 µm diameter. Ribs 5-7 or ~ 20 (B. riograndensis). Four species: B. junciformis (Fig. 7A-D), B. pentaptera (Fig. 7E-H), B. riograndensis (Fig. 7I-L) and B. sagittalis (Fig. 7M-P). Discussion Patterns observed in cypselae morphology in Baccharis sect. Caulopterae were similar to those previously reported for species of other sections of Baccha- ris (Velez, 1981; Hellwig, 1990; Mukherjee & Sarkar, 2001; Müller, 2006), which also show a ring-shaped carpopodium and papillose surface. For the tribe Astereae, and for other Baccharis species, the presence of glandular and non-glandular trichomes was also observed (Mukherjee & Sarkar, 2001; Müller, 2006). Velez (1981) reported that the cypselae of genus Baccharis are not uniform, and he separated them in two groups. Velez (1981) also reported that cypselae may vary from papillose to epapillose, with epidermal cells with slightly lignified walls or not, and a folded or flat cuticle, and these features were also observed in the present study. Additionally, we found that B. articulata and B. glaziovii present cavites, a condition structures not previously reported, although it is possible that this feature is an artefact. The Baccharis sect. Caulopterae species present a carpopodium as characterized by Haque & Godward (1984), who emphasized that most species of the Asteraceae family have this abscission structure. Species taxonomically close presented similarities in cypsela surface morphology, reflecting on the groups formed. However, since species of the “Baccharis trimera complex” (B. crispa, B. cylindrica, B. jocheniana, B. myriocephala, and B. trimera), all showed a similar cypsela micromorphology (Group III.b), cypsela characters were unhelpful to distinguish the species of this group. Anales del Jardín Botánico de Madrid 68(1): 107-116, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2245 2245_Baccharis:Anales 68(1).qxd 13/06/2011 12:14 Página 111 Microsculpture of cypselae of Baccharis 111 Fig. 2. Scanning electron micrographs of cypselae surface of Baccharis sect. Caulopterae - Group II: B. palustris: A, cypsela; B, C, epapillose surface; D, carpopodium. B. paranensis: E, cypsela; F, G, epapillose surface; H, carpopodium. B. pseudovillosa: I, cypsela; J, K, epapillose surface; L, carpopodium. B. ramboi; M, cypsela; N, O, epapillose surface; P, carpopodium. B. vincifolia; Q, cypsela; R, S, epapillose surface; T, carpopodium. Anales del Jardín Botánico de Madrid 68(1): 107-116, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2245 2245_Baccharis:Anales 68(1).qxd 13/06/2011 12:15 Página 112 112 A.A. Scheneider & I.I. Boldrini Fig. 3. Scanning electron micrographs of cypselae surface of Baccharis sect. Caulopterae - Group III.a: B. apicifoliosa: A, cypsela oblong-cylindrical; B, C, papillae detail; D, carpopodium [C]. B. flexuosiramosa: E, cypsela; F, G, papillae; H, carpopodium. B. milleflora: I, cypsela; J, K, papillae detail; L, carpopodium. B. organensis: M, cypsela; N, O, papillae; P, carpopodium. B. phyteumoides: Q, cypsela; R, S, papillae; T, carpopodium. Anales del Jardín Botánico de Madrid 68(1): 107-116, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2245 2245_Baccharis:Anales 68(1).qxd 13/06/2011 12:15 Página 113 Microsculpture of cypselae of Baccharis 113 Fig. 4. Scanning electron micrographs of cypselae surface of Baccharis sect. Caulopterae - Group III.a: B. subtropicalis: A, cypsela; B, C, papillae; D, carpopodium. Group III.b: B. crispa: E, cypsela; F, G, papillae; H, carpopodium. B. cylindrica: I, cypsela; J, K, papillae; L, carpopodium. B. jocheniana: M, cypsela; N, O, papillae; P, carpopodium. B. myriocephala: Q, cypsela; R, S, papillae; T, carpopodium. Anales del Jardín Botánico de Madrid 68(1): 107-116, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2245 2245_Baccharis:Anales 68(1).qxd 13/06/2011 12:15 Página 114 114 A.A. Scheneider & I.I. Boldrini Fig. 5. Scanning electron micrographs of cypselae surface of Baccharis sect. Caulopterae - Group III.b: B. aff. opuntioides: A, cypsela; B, C, papillae; D, carpopodium. B. trimera: E, cypsela; F, G, papillae; H, carpopodium. Fig. 6. Scanning electron micrographs of cypselae surface of Baccharis sect. Caulopterae - Group IV: B. articulata: A, cypsela oblong-cylindrical; B, C, papillae; D, no carpopodium. B. glaziovii: E, cypsela; F, G, papillae; H, carpopodium. [Cav, cavites, Pap, papilla]. Anales del Jardín Botánico de Madrid 68(1): 107-116, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2245 2245_Baccharis:Anales 68(1).qxd 13/06/2011 12:15 Página 115 Microsculpture of cypselae of Baccharis 115 Fig. 7. Scanning electron micrographs of cypselae surface of Baccharis sect. Caulopterae - Group V: B. junciformis: A, cypsela; B, C, epapillose surface; D, carpopodium. B. pentaptera: E, cypsela; F, G, epapillose surface; H, carpopodium. B. riograndensis: I, cypsela; J, K, epapillose surface; L, carpopodium. B. sagittalis: M, cypsela; N, O, epapillose surface; P, carpopodium. Acknowledgements We thank the curators of the herbaria ESA, ICN, PACA and UEC that provided material for this study, to Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the financial support, to Centro de Microscopia Eletrônica da Universidade Federal do Rio Grande do Sul (CEM) and technics Carlos Barboza dos Santos and Karina Marckmann for facilities in using of SEM. Particular thanks to Pedro Maria de Abreu Ferreira for suggestios on language, Cláudio Augusto Mondin, Mara Rejane Ritter, Rinaldo Pires dos Santos, Nádia Roque, Jochen Müller and Gustavo Heiden for the critical revision and suggestions for this manuscript. References Anderberg, A.A. 1991. Taxonomy and phylogeny of the tribe Gnaphalieae (Asteraceae). Opera Botanica 104: 1-195. Anales del Jardín Botánico de Madrid 68(1): 107-116, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2245 2245_Baccharis:Anales 68(1).qxd 13/06/2011 12:15 Página 116 116 A.A. Scheneider & I.I. Boldrini Ariza, L. 1973. Las especies de Baccharis de Argentina Central. Boletín de la Academia Nacional de Ciencias 50: 175-305. Barroso, G.M. 1976. Compositae – Subtribo Baccharidinae Hoffmann. Estudo das espécies ocorrentes no Brasil. Rodriguésia 28(40): 3-273. Barthlott, W. 1981. Epidermal and seed surface characteres of plants: systematic applicability and some evolutionary aspects. Nordic Journal of Botany 1(3): 345-355. Barthlott, W. 1990. Scanning electron microscopy of the epidermal surface in plants. In: Claugher, D. (ed.), Scanning Electron Microscopy in Taxonomy and Functional Morphology. Oxford, Clarendon Press, 69-94. Bremer, K. 1994. Asteraceae: Cladistics and classification. Portland. Timber Press. Haque, M.Z & Godward, M.B.E. 1984. New records of the carpopodium in Compositae and its taxonomic use. Botanical Journal of the Linnean Society 89: 321-340. Heiden, G., Iganci, J.R.V. & Macias, L. 2009. Baccharis sect. Caulopterae (Asteraceae, Astereae) no Rio Grande do Sul, Brasil. Rodriguésia 60(4): 943-983. Hellwig, F.H. 1990. Die Gattung Baccharis L.(Compositae-Asteraceae) in Chile. Mitteilungen der Botanischen Staatssammlung München 29: 1-456. Marzinek, J., De-Paula, O.C. & Oliveira, D.M.T. 2008. Cypsela or achene? Refining terminology by considering anatomical and historical factors. Revista Brasileira de Botânica 31(3): 549-553. Mukherjee, S.K. & Sarkar, A. 2001. Morphology and structure of cypselae in thirteen species of the tribe Astereae (Asteraceae). Phytomorphology 51(1): 17-26. Müller, J. 2006. Systematics of Baccharis (Compositae-Astereae) in Bolivia, including an overview of the genus. Systematic Botany Monographs 76: 1-339. Radford, A.E. 1986. Fundamentals of plants systematics. New York. Harper and Row. Roth, I. 1977. Fruits of angiosperms: encyclopaedia of plant anatomy. Berlin. Gebrüder Borntraeger. Schneider, A.A., Heiden, G. & Boldrini, I. 2009. Notas nomenclaturais em Baccharis L. sect. Caulopterae DC. (Asteraceae). Revista Brasileira de Biociências 7(2): 225-228. Velez, M.C. 1981. Karpologische untersuchungen an amerikanischen Astereae (Compositae) Mitteilungen der Botanischen Staatssammlung München 17: 1-170. Associate Editor: J. Müller Received: 9-II-2010 Accepted: 1-II-2011 Anales del Jardín Botánico de Madrid 68(1): 107-116, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2245 2280_Gelidiales:Anales 68(1).qxd 13/06/2011 12:21 Página 117 Anales del Jardín Botánico de Madrid Vol. 68(1): 117-124 enero-julio 2011 ISSN: 0211-1322 doi: 10.3989/ajbm.2280 Photosynthetic response and zonation of three species of Gelidiales from Tenerife, Canary Islands by S. Domínguez-Álvarez1, J.M. Rico2 & M.C. Gil-Rodríguez1 1 2 Departamento de Biología Vegetal (Botánica), Universidad de La Laguna, E-38071 La Laguna, Spain. Departamento de Biología de Organismos y Sistemas, Universidad de Oviedo, E-33071 Oviedo, Spain. [email protected] Abstract Resumen Domínguez-Álvarez, S., Rico, J.M. & Gil-Rodríguez, M.C. 2011. Photosynthetic response and zonation of three species of Gelidiales from Tenerife, Canary Islands. Anales Jard. Bot. Madrid 68(1): 117-124. Domínguez-Álvarez, S., Rico, J.M. & Gil-Rodríguez, M.C. 2011. Respuestas fotosintéticas y zonación de tres especies de Gelidiales de Tenerife, Islas Canarias. Anales Jard. Bot. Madrid 68(1): 117-124 (en inglés). Three species of Gelidiales (Gelidium arbuscula, Gelidium canariense and Pterocladiella capillacea) (Rhodophyta) were selected due to their abundance in the marine lower intertidal of the north coast of the island of Tenerife (Canary Islands), to assess, using PAM fluorescence, the importance of irradiance and exposure to air on vertical distribution. We compared tolerance to emersion by air-drying fronds under simulated emersion, and results suggest that recovery of photosynthesis after emersion plays a major role in the vertical distribution of these three species. Morphological traits such as clumped fronds explain the higher tolerances, and reduced water loss of the species upper on the shore. Local differences between sites may be related to slight differences in the light regime related to topography. Se han seleccionado tres especies de Gelidiales (Gelidium arbuscula, Gelidium canariense y Pterocladiella capillacea) que son abundantes en los niveles inferiores del intermareal de la costa N de Tenerife para establecer, utilizando fluorescencia tipo PAM, la importancia de la cantidad de luz y la exposición al aire en su zonación vertical. Se ha comparado la tolerancia a la emersión en frondes expuestas al aire, y los resultados sugieren que la capacidad de recuperación de la fotosíntesis tras la emersión tiene un papel fundamental en la explicación de la posición vertical de estas tres especies. Además, características morfológicas como el apelotonamiento de las frondes pueden coadyuvar a la mayor tolerancia, al reducir la pérdida de agua en emersión en las especies de niveles más altos. Las variaciones entre sitios se pueden deber a desigualdades locales en la cantidad de luz provocadas por diferencias topográficas. Keywords: Canary Islands, ecophysiology, Gelidiales, Gelidium, photosynthesis, Pterocladiella. Palabras clave: ecofisiología, fotosíntesis, Gelidiales, Gelidium, Islas Canarias, Pterocladiella. Introduction Brown, 1982; Rico & Fredriksen, 1996; Wiencke & al., 2000; Bischof & al., 2006). Intertidal seaweeds are exposed to rhythmic emersion and submersion periods. The dehydration of algal thalli during the emersion imposes very stressful conditions and it has been supposed that the recovery of photosynthesis after emersion could be a factor involved in the zonation pattern of seaweeds. For example, the recovery of photosynthesis of brown algae, after desiccation during the low tide, was correlated with their heights in the zonation of European shores (Dring & Brown, 1982). A similar correlation Zonation of seaweeds is the most conspicuous characteristic of rocky shores worldwide (Lewis, 1964; Stephenson & Stephenson, 1972; Pérès, 1982). Algal zonation seems to be due to several causes acting simultaneously, but the relative contribution of each factor is under debate. Thus, inter-specific competition has usually been the consensus for the main cause of zonation (Chapman, 1973, 1974). However, experimental studies propose that physiological limits of seaweeds are also involved in zonation (Dring & 2280_Gelidiales:Anales 68(1).qxd 13/06/2011 12:21 Página 118 118 S. Domínguez-Álvarez & al. was argued for the zonation pattern of different species in Gelidiaceae in N Iberian Peninsula (Rico & Fredriksen, 1996). However, more experimental studies are needed/required to improve our knowledge the relationship between photosynthetic performance of intertidal algae and their heights in the zonation pattern. It is also known that dynamic photo-inhibition of photosynthetic quantum yield (Hall & Rao, 1999) occurs in seaweeds exposed to high levels of irradiance at noon, but such photo-inhibition is more important the lower in the zonation that the species occurs. Thus, although a very low level of photo-inhibition was detected in the red alga Rissoella verruculosa growing in the upper intertidal (Flores-Moya & al., 1998) photoinhibition is more important in seaweeds from the lower part of the intertidal (Huppertz & al., 1990; Hanelt & al., 1993); moreover, the reduction of photosynthesis was very high in a subtidal species transplanted close to sea surface (Gevaert & al., 2002). The tides that affect the coasts of the Canaries are of semidiurnal character, with highest high waters that do not surpass 3 m above Lowest Astronomical Tide (LAT), whereas the lowest low tides reach, occasionally, values of 0 m. The steep northern coast of the island of Tenerife is very exposed, and this constitutes one of the most decisive factors determining the species composition of the lower intertidal vegetation that is formed, characteristically, by species that tolerate big swell. In these lower intertidal and shal- low subtidal environments, three species of Gelidiales form successive belts. The upper one is an intense red colour, dominated by Pterocladiella capillacea (S.G. Gmelin) Santelices & Hommersand; then P. capillacea is progressively substituted by Gelidium arbuscula Bory de Saint-Vincent ex Børgesen (around 0.7-0.8 m above LAT). Below it there is a wide belt, reaching the upper subtidal, darker in colour, dominated by Gelidium canariense (Grunow) Seoane-Camba ex Haroun, Gil-Rodríguez, Díaz de Castro et Prud' homme van Reine. The three species are present just in the lower intertidal where they are under the influence of spray even during low tides (Mercado & al., 2001). Thus, the zonation pattern of P. capillacea, G. arbuscula and G. canariense in northern Tenerife shores is an appropriate model to test the hypothesis that their heights on the shore may be correlated with their photosynthetic performance. The objective of this study was to characterize the photosynthetic response of the three species of Gelidiales at two localities on Tenerife and to relate it to the stress of emersion and changes in irradiance levels associated with their different vertical distribution. In particular, the two hypothesis to be tested were: (i) that the time of recovery of photosynthetic activity after desiccation in the selected species (estimated via fluorescence parameters) is higher the lower is the location on the zonation pattern; and (ii) the photoinhibition of the photosynthetic quantum yield around noon is higher the lower is the location in the zonation. Table 1. ANOVA results for Ik and ETRrmax. *the factor ‘Species’ was tested against ‘Species x Run’. ‘Species x Site’ was removed from the analysis (non-significant, P = 0.27). Ik Source of variation df ETRrmax MS F P MS F P Species* 2 281.20 1.00 0.421 164184 4.50 0.064 Site 1 2990.00 5.02 0.111 176026 12.60 0.038 Run 3 840.23 1.41 0.392 45483 3.26 0.179 Species × Site 2 197.43 0.78 0.499 18461 1.64 0.270 Species × Run 6 280.36 1.11 0.451 36515 3.25 0.089 Site × Run 3 596.03 1.26 0.293 13972 1.11 0.350 Species×Site×Run 6 252.66 0.53 0.782 11242 0.89 0.505 Error 96 474.03 Total 119 12624 Anales del Jardín Botánico de Madrid 68(1): 117-124, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2280 2280_Gelidiales:Anales 68(1).qxd 13/06/2011 12:21 Página 119 Photosynthetic response of Gelidiales 119 Fig. 1. Daily cycle of photosynthetic quantum yield (ΦPSII; dimensionless) in Gelidium canariense. Values are represented as mean ± SD (n = 5) from samples of Puerto de la Cruz (䡬) and Garachico (䢇). Fig. 2. Daily cycle of photosynthetic quantum yield (ΦPSII; dimensionless) in Gelidium arbuscula. Values are represented as mean ± SD (n = 5) from samples of Puerto de la Cruz (䡬) and Garachico (䢇). Material and methods Experiment A: daily variation in photosynthetic quantum yield Sampling sites, algal collection and pre-treatment of samples The Northern coast of Tenerife is very exposed, with frequent periods of intense swell 2 to 3 m height, and occasional calm periods throughout the year, mainly in spring and summer. During autumn the storms with waves up to 5 m height are frequent. Water temperature ranges 17 ºC in February to 23 ºC in August. Two localities separated 30 km were selected: Puerto de la Cruz (UTM 3675447/3588716) and Garachico (UTM 3700288/3590165). Both sites haves similar patterns of marine vegetation, characterized by a belt of blue-green algae in the upper intertidal, a lower caespitose belt dominated by Gelidium pusillum (Stackhouse) Le Jolis, Caulacanthus ustulatus (Merthens ex Turner) Kützing and Ulva rigida C. Agardh, another below mainly of species of the Corallinaceae and Rhodomelaceae and, in the lowest levels, the three species of Gelidiales down to the subtidal (with Pterocladiella capillacea in the upper part, G. arbuscula in the middle and G. canariense in the lowest part) (Gil-Rodríguez & Wildpret, 1980; Pinedo & Afonso-Carrillo, 1994). Thalli free from epiphytes of the three Gelidiales cited above were collected in July 1999. They were transferred to the laboratory in seawater and darkness and kept in 50 L aquaria at constant temperature (ca. 20 °C). Natural solar irradiance was reduced by using plastic mesh that acted as neutral density filters reducing incident PAR to 60-80% of full sunlight. A system of continuous seawater circulation was designed to keep 10 L glass aquaria with 10-20 g fresh weight under a constant temperature of 20 ºC during incubations. Measurements of photosynthetic activity were carried out by using a DIVING-PAM (Heinz Walz GmbH, Germany) during a whole daily cycle, from sunset to sunrise. Every hour from sunrise, five Fig. 3. Daily cycle of photosynthetic quantum yield (ΦPSII; dimensionless) in Pterocladiella capillacea. Values are represented as mean ± SD (n = 5) from samples of Puerto de la Cruz (䡬) and Garachico (䢇). Anales del Jardín Botánico de Madrid 68(1): 117-124, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2280 2280_Gelidiales:Anales 68(1).qxd 13/06/2011 12:21 Página 120 120 S. Domínguez-Álvarez & al. Fig. 4. Photosynthetic quantum yield (ΦPSII; dimensionless) of Gelidium canariense from Puerto de la Cruz (left) and Garachico (right) during emersion (values to the left of the vertical broken line) and after resubmergence (values to the right of the broken line). Each line corresponds to a different run. Fig. 5. Photosynthetic quantum yield (ΦPSII; dimensionless) of Gelidium arbuscula from Puerto de la Cruz (left) and Garachico (right) during emersion (values to the left of the vertical broken line) and after resubmergence (values to the right of the broken line). Each line corresponds to a different run. fronds from the aquaria for each locality and species were selected and photosynthesis was measured. During the central hours of the day (–2 to +2 hours around noon) measurements were taking at 30 min intervals. According to Genty & al. (1989), photosynthetic activity was estimated from the effective quantum yield of chlorophyll a from photosystem II (ΦPSII defined as ⌬F/Fm’). The use of the ΦPSII values avoid problems derived from the potential differences be- tween absorptances in different species (Franklin & Badger, 2001) although these differences can be minor in species from the same genus/family and with similar morphology. Additionally, five fronds per species and locality were exposed to nine levels of increasing PAR (0, 12, 55, 122, 213, 317, 443, 653, 889 µmol m-2 s-1) during 45 s each, after which a saturating pulse was applied and ΦPSII values measured. The relative electron transport Anales del Jardín Botánico de Madrid 68(1): 117-124, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2280 2280_Gelidiales:Anales 68(1).qxd 13/06/2011 12:21 Página 121 Photosynthetic response of Gelidiales 121 Fig. 6. Photosynthetic quantum yield (ΦPSII; dimensionless) of Pterocladiella capillacea from Puerto de la Cruz (left) and Garachico (right) during emersion (values to the left of the vertical broken line) and after resubmergence (values to the right of the broken line). Each line corresponds to a different run. rate (ETRr) was computed as PAR × ΦPSII, when ΦPSII value was >0.1 (Beer & Axelsson, 2004). The plot of ETRr versus PAR was used to calculate the irradiancesaturated ETR over the saturation point (ETRrmax) and the value of PAR above which ETRrmax was detected (Ik); for this purpose, a non-linear fit to a hyperbolic curve was applied. The experiment was repeated (using different fronds) every 2 h from sunrise to sunset. Experiment B: response of photosynthetic performance to desiccation stress and rehydration Five fronds from each locality and species were exposed to air by placing them in plastic trays after vigorous shaking to simulate desiccation stress during low tide exposure to air. Fronds were exposed during the hours of highest PAR, since low tides occurred around noon during the experiments. Desiccation stress was estimated from ΦPSII measurements at 30 min intervals. Fronds were also weighed to the nearest mg at the start and at the end of the experiment to calculate the proportion of water loss. After an exposure of 2.5 h (equivalent to the longest emersion period during spring tides), fronds were rehydrated by submersion in seawater and fluorescence measurements were conducted for 2.5 h at 30 min intervals. Statistical analysis of data Data were analyzed using factorial ANOVAs (see results for details on each analysis). All data were analyzed using Statistica for Windows. Results The daily cycle of ΦPSII in the three species showed a similar pattern: the highest ΦPSII values were recorded at the sunrise and decreased abruptly to minimum values (about 50% lower than those on earlier hours) around solar noon; then, ΦPSII increased gradually at the sunset (Figs. 1-3). A marked reduction in ΦPSII was found in the three species when thalli were exposed to air for 2.5 h simulating emersion conditions (Figs 4-6); however, the highest reduction of ΦPSII was recorded in Gelidium canariense (Fig. 4) while Pterocladiella capillacea was the species with the lowest reduction of ΦPSII (Fig. 6). The reduction of ΦPSII in G. arbuscula after desiccation was intermediate to that of the two other species (Fig. 5). However, the relative water loss was similar in the three species from both localities (overall mean % water loss = 50.3 ± 7.4, n = 30). The values of ΦPSII after 1.5 h rehydration in G. arbuscula was around 3050% than to those measured prior to exposure to air (Fig. 5) while recovery in P. capillacea and G. canariense was lower (Figs. 4, 6). Photoinhibition was not detected in the ETRr-PAR curves for the three species, and both ETRrmax and Ik values were similar for the three species at both localities (Figs. 7, 8, Table 1). Saturation occurred above 150 µmol m-2 s-1 for the three species, reaching around 500 µmol m-2 s-1 for some samples of G. arbuscula (Fig. 7). Anales del Jardín Botánico de Madrid 68(1): 117-124, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2280 2280_Gelidiales:Anales 68(1).qxd 13/06/2011 12:21 Página 122 122 S. Domínguez-Álvarez & al. Run Fig. 7. Values of saturating PAR (Ik) for the three species of Gelidiales from Puerto de la Cruz (䢇) and Garachico (䡬). ‘Run’ refers to the time period (UTC+2) when the measurements for the curves where taken (1: from 10 to 11 a.m.; 2: from 12 to 13; 3: from 14 to 15; 4: from 16 to 17). Run Fig. 8. Values of irradiance-saturated, relative electron transport rate (as ETRrmax) for the three species of Gelidiales from Puerto de la Cruz (䢇) and Garachico (䡬).‘Run’ refers to the time period (UTC+2) when the measurements for the curves where taken (1: from 10 to 11 a.m.; 2: from 12 to 13; 3: from 14 to 15; 4: from 16 to 17). Discussion The presence of several species of Gelidiales in contiguous altitudinal belts is a common feature of many temperate coasts (Santelices, 1991) and it has been suggested that this pattern indicates close similarities among species with respect to environmental tolerance (Santelices, 1991; Montalva & Santelices, 1981). In the Gelidium-dominated lower shore in N Spain, irradiance did not appear as a critical factor for photosynthesis (Rico, 1991), but different Gelidium species showed different saturation values (Ik) which could be related to their altitudinal position on the shore (Rico & Fredriksen, 1996). This is not the case with the three Gelidiales from Teneriffe, which exhibited similar patterns of both photoinhibition of ΦPSII along the daily cycle and ETRr-PAR curves, and it has been shown that ETR values can be used to estimate photosynthesis in Gelidium species since they compare well with photosynthetic estimations using, for instance, O2 electrodes (Beer & Axelsson, 2004; Franklin & Badger, 2001; Silva & al., 1998). Anales del Jardín Botánico de Madrid 68(1): 117-124, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2280 2280_Gelidiales:Anales 68(1).qxd 13/06/2011 12:21 Página 123 Photosynthetic response of Gelidiales Since the photosynthetic response under standard conditions did not differ between species, the position on the shore could be explained by negative effects of desiccation on net photosynthesis as it has been demonstrated in fucoids (Williams & Dethier, 2005). Desiccation occurs during low tide almost every day, its length determined by the duration of the tide. Gelidium sesquipedale from Asturias, a lower intertidal species, showed a higher reduction in net photosynthesis after exposure to air (Rico & Fredriksen, 1996), and an exposure longer than 2 h caused a negative net production, thus photosynthesis was overcome by respiration. The same occurred for G. canariense, the species lowest on the shore in Teneriffe, which was more affected by desiccation and also recovered to a lesser degree after rehydration. Recovery of photosynthesis after emersion is one of the major factors explaining the upper limits of seaweeds in the intertidal (Dring & Brown, 1982). It has been suggested that both tolerance to sustain photosynthesis under desiccation and reduction in the rate of water loss can help intertidal species to tolerate longer emersion periods (Oates & Murray, 1983). This could be the case with the two species located upper in the shore in our study: G. arbuscula is morphologically similar to G. pulchellum, which showed a reduced rate of water loss after emersion periods of more than 3 h (Rico & Fredriksen, 1996), while P. capillacea showed a rapid recovery after rehydration. All three species were able to recover after exposure to periods shorter than 2.5 h which suggests that small differences in the recovery rate could explain upper tolerance limits on the shore. Duration of natural exposure to air in intertidal seaweeds determines the degree of drying, but during these periods it is the rate of drying which explains the stress (Hunt & Denny, 2008). Thus, tolerance to desiccation (as opposed to the desiccation stress) and the light environment (Mercado & al., 2001) are the main factors to explain the zonation pattern of these three species in the coast of Teneriffe. Apart from the obvious effect of reduced irradiance levels during high tides affecting photosynthetic production, exposure to full solar radiation during low tides can also affect photosynthetic performance, mainly at noon (Gómez & Figueroa, 1998). Accordingly, P. capillacea showed the most intense noon-depleted ETRr rate, while showing intermediate values of the photosynthetic parameters. Gelidium arbuscula did not show noon depletion in ΦPSII and showed the highest values both in ETRrmax and in Ik, and G. canariense showed a moderate reduction in ΦPSII at noon but not so high values in the photosynthetic parameters. It could be concluded that the upper species, P. capillacea, is adapted to exposure to air and 123 high irradiance levels during emersion by reducing ΦPSII and showing fast recovery after rehydration, while G. arbuscula is able to reduce water loss due to its compact morphology, which may also produce frond shading and so attenuate the effect of high irradiance, and ‘clump’ morphology represents a significant advantage to reduce water loss in central portions of the thallus (Hunt & Denny, 2008). Differences in zonation between sites are intriguing, although between site variation has been reported before and explained by a multiarray of factors other than exposure (Williams & Dethier, 2005). Further studies will be needed to fully assess these additional factors which may explain small scale differences in the zonation pattern, although small topographic differences may produce differences also in the local light regime. Acknowledgements Personnel from the Centro Oceanográfico de Canarias - Instituto Español de Oceanografía de Canarias (IEO), gave support to experiments. The contribution of J.M. Cejas is especially acknowledge. References Beer, S. & L. Axelsson. 2004. Limitations in the use of PAM fluorometry for measuring photosynthetic rates of macroalgae at high irradiances. European Journal of Phycology 39: 1-7. Bischof, K., Gómez, I., Molis, M., Hanelt, D., Karsten, U., Lüder, U., Roleda, M.Y., Zacher, K. & Wiencke, C. 2006. Ultraviolet radiation shapes seaweed communities. Reviews in Environmental Science and Biotechnology 5: 141-166. Chapman, A.R.O. 1973. A critique of prevailing altittudes towards the control of seaweed zonation on the seashore. Botanica Marina 16: 80-82. Chapman, A.R.O. 1974. The ecology of macroscopic marine algae. Annual Review of Ecology and Systematics 5: 65-80. Dring, M.J. & Brown, F.A. 1982. Photosynthesis of intertidal brown algae during and after periods of emersion: a renewed search for physiological causes of zonation. Marine Ecology Progress Series 8: 301-308. Flores-Moya, A., Gómez, I., Viñegla, B., Altamirano, M., PérezRodríguez, E., Maestre, C., Caballero, R. & Figueroa, F.L. 1998. Effects of solar radiation on the endemic Mediterranean red alga Rissoella verruculosa: photosynthetic performance, pigment content and the activities of enzymes related to nutrient uptake. New Phytologist 139: 673-683. Franklin, L.A. & Badger, M.R. 2001. A comparison of photosynthetic electron transport rates in macroalgae measured by pulse amplitude modulated chlorophyll fluorometry and mass spectrometry. Journal of Phycology 37: 756-767. Genty, B., Briantais, J.M. & Baker, N.R. 1989. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll. Biochimica et Biophysica Acta 990: 87-92. Gevaert, F., Creach, A., Davoult, D., Holl, A.-C., Seuront, L. & Lemoine, Y. 2002. Photo-inhibition and seasonal photosynthetic performance of the seaweed Laminaria saccharina during a simulated tidal cycle: chlorophyll fluorescence measurements and pigment analysis. Plant, Cell and Environment 25: 859-872. Anales del Jardín Botánico de Madrid 68(1): 117-124, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2280 2280_Gelidiales:Anales 68(1).qxd 13/06/2011 12:21 Página 124 124 S. Domínguez-Álvarez & al. Gil-Rodríguez, M.C. & Wildpret, W. 1980. Contribución al estudio de la vegetación ficológica marina del litoral canario. Aula de Cultura Excmo. Cabildo Insular de Tenerife. Gómez, I. & Figueroa, F.L. 1998. Effects of UV stress on chlorophyll fluorescence kinetics of intertidal macroalgae from southern Spain: a case study in Gelidium species. Journal of Applied Phycology 10: 285-294. Hall, D.O. & Rao, K.K. 1999. Photosynthesis. Cambridge University Press, Cambridge. Hanelt, D., Huppertz, K. & Nultsch, W. 1993. Daily courses of photosynthesis and photo-inhibition in marine macroalgae investigated in the laboratory and field. Marine Ecology Progress Series 97: 31-37. Hunt, L.J.H. & Denny, M.W. 2008. Desiccation protection and disruption: a trade-off for an intertidal marine alga. Journal of Phycology 44: 1164-1170. Huppertz, K., Hanelt, D. & Nultsch, W. 1990. Photo-inhibition of photosynthesis in the marine brown alga Fucus gardneri as studied in field experiments. Marine Ecology Progress Series 66: 175182. Lewis, J.R. 1964. The Ecology of Rocky Shores. English University Press, London. Mercado, J.M., Niell, F.X. &. Gil-Rodríguez, M.C. 2001. Photosynthesis might be limited by light, not inorganic carbon availability, in three intertidal Gelidiales species. New Phytologist 149: 431-439. Montalva, S. & Santelices, B. 1981. Interspecific interference among species of Gelidium from Central Chile. Journal of Experimental Marine Biology and Ecology 53: 77-88. Oates, B.R. & Murray, S.N. 1983: Photosynthesis, dark respiration and dessication resistance of the intertidal seaweeds Hesperophycus harveyanus and Pelvetia fastigiata f. gracilis. Journal of Phycology 19: 371-380. Pérès, J.M. 1982. Zonations and organismic assemblages. In: Kinne, O. (ed.), Marine Ecology, Vol. V, Ocean management, Part 1. Wiley, Chichester, pp. 9-576. Pinedo, S. & J. Afonso-Carrillo. 1994. Distribución y zonación de las algas marinas bentónicas en Puerto de la Cruz, Tenerife. Vieraea 23: 109-123. Rico, J.M. 1991. Field studies and growth experiments on Gelidium latifolium from Asturias (N Spain). Hydrobiologia 221: 67-75. Rico, J.M. & Fredriksen, S. 1996. Effects of environmental factors on net photosynthesis and growth of intertidal species of the genus Gelidium (Gelidiaceae, Rhodophyta) in northern Spain. Scientia Marina 60: 265-273. Santelices, B. 1991. Production ecology of Gelidium. Hydrobiologia 221/222: 31-44. Silva, J., Santos, R., Serodio, J. & Melo, R.A. 1998. Light response curves for Gelidium sesquipedale from different depths, determined by two methods: O2 evolution and chlorophyll fluorescence. Journal of Applied Phycology 10: 295-301. Stephenson, T.A. & Stephenson. A. 1972. Life Between TideMarks on Rocky Shores. W.H. Freeman, USA. Wiencke, C., Gómez, I., Pakker, H., Flores-Moya, A., Altamirano, M. Hanelt, D., Bischof, K. & Figueroa, F.L. 2000. Impact of UV-radiation on viability, photosynthetic characteristics and DNA of brown algal zoospores: implications for depth zonation. Marine Ecology Progress Series 197: 217-229. Williams, S.L. & Dethier, M.N.. 2005. High and dry: variation in net photosynthesis of the intertidal seaweed Fucus gardneri. Ecology 86: 2373-2379. Associate Editor: A. Flores Received: 27-XII-2010 Accepted: 10-II-2011 Anales del Jardín Botánico de Madrid 68(1): 117-124, enero-julio 2011. ISSN: 0211-1322. doi: 10.3989/ajbm. 2280 00 PUBLICIDAD FLORA ANALES:00 PUBLICIDAD FLORA ANALES.qxd 13/06/2011 12:25 Página 125 Flora iberica Plantas vasculares de la Península Ibérica e Islas Baleares “La qualité, à tous égards, de ce qui est en train de se faire à Madrid et ailleurs en Espagne aurait été inimaginable, il y a peu d‘années, en dehors du monde anglophone. Et bien, aujourd’hui ce son nos confrères espagnols qui ont pris la tête en la matière –à nous autres, anglophones ou non, de suivre le mouvement”. W. Greuter in Optima Newsletter 25-29: 22-23. “Im Still und Layout könnte die Flora Iberica als Beispiel für manche anderen Florenwerke dienen”. K.H. Rechinger in Willdenowia 20: 318. Flora iberica “It is practically impossible to foul this Flora...”. S.L. Jury in Bot. J. Linn. Soc. 107: 112-114. “All in all, a flora of a very high standard and a worthy successor to Willkomm & Lange ‘Prodromus...’. For botanists working in European Flora... this multi-volume work is indis pensable...”. K. Larsen in Nordic J. Bot. 11: 92. sculares Plantas va ula Ibérica ns de la Pení Baleares as e Isl Flora iberica Flora iberica Vol. III RACEAE )-CAPPA AE (partim NACE UMBAGI PL es vascular Plantas nsula Ibérica ní de la Pelas Baleares e Is Plantas de la Penínvasculares su e Islas Ba la Ibérica leares . CO, C.S.I.C BOTÁNI Flora iberica RDÍN REAL JA ares Plantas vasculIbérica de la Península res e Islas Balea LYCOPO E Vol. I ACEA PAVER AE-PA DIACE a Florica iber REAL Vol. V EAE-SAX IFRAGAC EBENAC EAE I.C. O, C.S. NIC ÍN BOTÁ JARD REAL JA RDÍN BO lares ascu Ibérica tas v Plan enínsulaleares P a de lae Islas B CSIC BOTÁNICO, REAL JARDÍN ra Florica ibe TÁNICO, CSIC Vol. VII (I) SAE (partim) LEGUMINO Flo iberi ra ca Flora iberica P de lalantas va scu Pe e Islanínsula lares s Ba Ibéric leare a s Plantas vascula de la Península res Ibér e Islas Baleares ica res cula rica vas Ibé ntasnínsulaeares a l P Pe Bal a s de l e Isla IAC XIII LAR Vol. ROPHU -SC PLA NTA GIN EAE AE ACE SIC O, C ÁNIC BOT DÍN JAR L A RE PLA TAN A CEA VI Vol. EAE SAC RO SIC O, C NIC TÁ BO N Í D JAR AL RE E-PL Vol. II UMB AGIN ACE AE (p Vol. VIII HALORAGACEAE -EUPHORBIACE AE REA L JA artim RDÍN BOT ÁNIC O, C REAL JARDÍN BOTÁNICO, CSIC .S.I.C Vols. I - VII (I), VII (II), VIII, X, XII - XV, XVII, XVIII y XXI . ) Total ❏ Mastercard ❏ Eurocard ❏ 4B 21.88 23.59 41.00 36.00 24.00 27.05 27.05 31.00 27.00 30.00 36.00 30.00 21.02 30.00 23.00 30.00 30.00 Price (€) Total No. of copies Euros Distribution and sale: Departamento de Publicaciones del CSIC C/ Vitruvio, 8. E-28006 Madrid Tel.: +34 915 612 833 / +34 915 159 742/744/717 - Fax: +34 915 629 634 E-mail: [email protected] - www.publicaciones.csic.es C/ Vitruvio, 8. 28006 Madrid Tel.: 915 612 833 / 915 159 742/744/717 - Fax: 915 629 634 E-mail: [email protected] - www.publicaciones.csic.es Date ..................... Signature ............................................................ ❏ Payment is made by international bank check (enclosed) of ....................................... € made payable to DEPARTAMENTO DE PUBLICACIONES DEL CSIC. ❏ Payment is made by international money order (copy enclosed) of ......................... € to the following account: D EPARTAMENTO DE P UBLICACIONES DEL CSIC, Account no. 1943/2, Banco de Santander, Paseo de la Castellana, 86. E-28046 Madrid. ❏ Please, send me a proforma invoice. VAT included for EU countries I II III IV V VI VII (I) VII (II) VIII X XII XIII XIV XV XVII XVIII XXI Volume Distribución y venta: Departamento de Publicaciones del CSIC Fecha ................... Firma ................................................................... ❏ Credit Card ❏ Contra reembolso. ❏ Envíenme, por favor, factura pro-forma. IVA incluido Euros ❏ Please, send me the following volumes: ❏ Les ruego me envíen los siguientes volúmenes: N.º de ejemplares Telephone ............................ Fax ............................. E-mail ............................... Teléfono ......................... Fax ............................. E-mail ..................................... 21,88 23,59 41,00 36,00 24,00 27,05 27,05 31,00 27,00 30,00 36,00 30,00 21,02 30,00 23,00 30,00 30,00 Code ....................... City ......................................... Country ............................. Código postal ......................... Localidad ............................................................ Precio (€) Address ................................................................................................................. Dirección ................................................................................................................ I II III IV V VI VII (I) VII (II) VIII X XII XIII XIV XV XVII XVIII XXI Name and surname ............................................................................................... Nombre y apellidos ............................................................................................... Volumen Order form for Flora iberica (FOREIGN COUNTRIES) Boletín de pedido de Flora iberica (ESPAÑA) 01 BOLETIN DE SUSCRIPCION FLORA:01 BOLETIN DE SUSCRIPCION FLORA.qxd 13/06/2011 12:47 Página 126 01 flora guinea:flora guinea.qxd 13/06/2011 12:32 Página 127 Flora de Guinea Ecuatorial Este es el segundo volumen de una serie de 11 previstos, que rellenarán un importante hueco en los estudios de biodiversidad vegetal del golfo de Guinea, una de las zonas de más alta diversidad de África. El objetivo es facilitar la identificación de las plantas vasculares autóctonas o naturalizadas de Guinea Ecuatorial. En las claves se incluyen las especies cuya presencia consta también en los países vecinos: Camerún (Cam), Gabón (Gab) o Santo Tomé y Príncipe (ST), pues podrían aparecer fácilmente en la zona de estudio, pero no se numeran ni se incluye más informacion sobre ellas. Sobre cada especie se ofrecen las informaciones siguientes: nombre aceptado, autor y referencia bibliográfica del lugar de publicación del nombre; sinónimos; tipos; descripciones; iconografía; hábitat y altitud; distribución; material estudiado y mapa. This is the second volume in a series of a planned 11, fills an important gap in the studies of plant biodiversity of the Gulf of Guinea, one of the areas of greatest diversity in Africa. The aim of this work is to facilitate the identification of autochthonous or naturalised vascular plants from Equatorial Guinea. The keys do include species whose presence is recorded in neighbouring countries: Cameroon (Cam), Gabon (Gab) or São Tomé and Príncipe (ST); these may indeed appear in the zone under study, although they shall neither be numbered nor shall any other information be included about them. We offer the following information on each species: accepted name, author and bibliographic reference of where the name was published; synonyms; types; descriptions; iconography; habitat and altitude; distribution; material studied and map. Total Euros C/ Vitruvio, 8. E-28006 Madrid Tel.: +34 915 612 833 / +34 915 159 742/744/717 - Fax: +34 915 629 634 E-mail: [email protected] - www. publicaciones.csic.es Signature: C/ Vitruvio, 8. E-28006 Madrid Tel.: +34 915 612 833 / +34 915 159 742/744/717 - Fax: +34 915 629 634 E-mail: [email protected] - www. publicaciones.csic.es Reimbursement (only for single issues) Distribution and sale: Departamento de Publicaciones del CSIC Firma: Total No. of copies Pro Forma Invoice Bank Transfer of Account Number: 0049 5117 2621 1010 5188 SWIFT/BIC Code: BSCHESMM IBAN number: ES83 0049 5117 2621 1010 5188 Nominal Cheque to Departamento de Publicaciones del CSIC Credit Card: Visa / Master Card / Eurocard / 4B Number: _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ Expiry date: _ _ / _ _ Manner of Payment: 36.00 49.00 Distribución y venta: Departamento de Publicaciones del CSIC Reembolso (sólo para números sueltos) Factura Transferencia bancaria a la cuenta número: 0049 5117 2621 1010 5188 SWIFT/BIC Code: BSCHESMM IBAN number: ES83 0049 5117 2621 1010 5188 Cheque nominal al Departamento de Publicaciones del CSIC Tarjeta de crédito: Visa / Master Card / Eurocard / 4B Número: _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ Fecha de caducidad: _ _ / _ _ Forma de Pago: 49,00 V I V Price (€) Please, send me the following volumes: Les ruego me envíen los siguientes volúmenes: Volume E-mail ________________________________ Date of subscription ____/____/_______ E-mail _______________________________ Fecha de la solicitud ____/____/_______ Euros Country/State ____________________ Telephone _____________ Fax _____________ País/Estado ___________________ Teléfono _______________ Fax _______________ N.º de ejemplares City ______________________________ Province _______________________________ Localidad __________________________ Provincia _____________________________ 36,00 Direction __________________________________________ Post Code ____________ Dirección _________________________________________________ CP ____________ Precio (€) Name of Organization ______________________________________________________ Razón social ________________________________________ NIF/CIF ______________ I Name and surname ________________________________________________________ Nombre y apellidos ________________________________________________________ Volumen Order form for Flora de Guinea Ecuatorial Boletín de pedido de Flora de Guinea Ecuatorial 02 BOLETIN SUSCRIPCION FLORA guinea:01 BOLETIN DE SUSCRIPCION FLORA.qxd 13/06/2011 12:48 Página 128 10 instrucciones a los autores:01 primeras.qxd 13/06/2011 12:33 Página 129 CONDICIONES DE PUBLICACIÓN / CONDITIONS FOR PUBLICATION Anales del Jardín Botánico de Madrid acepta artículos originales de interés general que no coincidan sustancialmente con alguno que haya sido publicado anteriormente. Los originales deben seguir las normas especificadas en la Guía para los autores (http://www.rjb.csic.es/publicaciones/anales/). Los autores, ante cualquier duda, pueden contactar con el Director ([email protected]) antes de enviar sus originales, lo que puede facilitar el proceso de edición. Todos los originales serán revisados por dos evaluadores externos a la entidad editora, cuyas sugerencias se enviarán a los autores para que realicen las modificaciones pertinentes, de acuerdo con los criterios de calidad científica. Será el Consejo de Redacción el que emita la decisión final a la vista de los informes de los evaluadores. El método de evaluación empleado es “doble ciego”, manteniendo el anonimato tanto del autor como de los evaluadores. Anales del Jardín Botánico de Madrid accepts original articles of general interest that have not appeared, or will not appear, elsewhere in substantially the same or abbreviated form. Authors are asked to carefully follow the Instructions to the authors (http://www. rjb.csic.es/publicaciones/anales/) below concerning preparation of the manuscripts. If in doubt, authors are urged to contact the Editor-inChief ([email protected]) prior to submitting the manuscripts, in order to facilitate the editing process. All the originals will be checked by two evaluators outside the publishing entity, whose suggestions will be sent to the authors so that they may mane the pertinent modifications, in accordance with the criteria of scientific quality. The Editorial Board will make the final decision, with consideration of the reviews of the external evaluators. This “double-blind” method of evaluation supports the anonymity of the author and of the evaluators. 10 instrucciones a los autores:01 primeras.qxd 13/06/2011 12:33 Página 130 11 Bolsuscrip. Anales 2011:10. Bolsuscrip. Anales 2009.qxd 13/06/2011 12:34 Página 131 SUSCRIPCIÓN Y PEDIDOS DATOS DEL PETICIONARIO Nombre y apellidos ________________________________________________________________________________________ Razón social ________________________________________________ NIF/CIF ______________________________________ Dirección ___________________________________________________________ CP _________________________________ Localidad ______________________________________________ Provincia _________________________________________ País/Estado _______________________________________________ Teléfono _______________________________________ Fax ________________________ Correo electrónico _____________________________ Fecha de solicitud _____ / _____ / _____ ■ Suscripción: Precios suscripción año 2011 Año completo: España: 30,37 € Extranjero: 48,37 € ■ Números sueltos CANTIDAD AÑO VOLUMEN NÚMERO Precios números sueltos año 2011 España: 19,17 € (más gastos de envío) Extranjero: 26,37 € (más gastos de envío) A estos precios se les añadirá el 4% (18% en soporte electrónico) de IVA. Solamente para España y países de la UE Forma de Pago: Factura pro forma ■ Transferencia bancaria a la Cuenta Número: C/c 0049 5117 2621 1010 5188 SWIFT/BIC CODE: BSCHESMM - IBAN NUMBER: ES83 0049 5117 2621 1010 5188 ■ Cheque Nominal al Departamento de Publicaciones del CSIC ■ Tarjeta de Crédito: Visa / Master Card / Eurocard / 4B Número: _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ Fecha de caducidad: _ _ / _ _ ■ Reembolso (solamente para números sueltos) Distribución y venta: Departamento de Publicaciones del CSIC C/ Vitruvio, 8 E-28006 Madrid Teléfonos: +34 915 612 833 / +34 915 681 619/620/640 Fax: +34 915 629 634 Correo electrónico: [email protected] www. publicaciones.csic.es Firma ___________________________________________________________________________________________________ 11 Bolsuscrip. Anales 2011:10. Bolsuscrip. Anales 2009.qxd 13/06/2011 12:34 Página 132 SUBSCRIPTIONS AND ORDERS CUSTOMER DETAILS Name, Surname ___________________________________________________________________________________________ Company/institucion name __________________________________________________________________________________ Tax registration code (Spain only)/VAT number (EU only) _________________________________________________________ Street/Address ____________________________________________________________________________________________ Town/City ________________________________________________ Province _______________________________________ Country/State _______________________________________________ Telephone ____________________________________ Fax _______________________________ E-mail: ____________________________ Date of subscription _____ / _____ / _____ ■ Subscription: Price 2011 Susbscription: two issues per year: Spain: 30.37 € International: 48.37 € ■ Separate issues QUANTITY YEAR VOLUME NUMBER Price 2011 single issue Spain: 19.17 € (More postage and handling) International: 26.37 € (More postage and handling) To these prices will be added 4% (18% in electronic support) of VAT. Only for Spain and EU countries Payment ■ Bank wire to account No.: (in Spain) c/c: 0049 5117 26 2110105188 ■ Cheque written to the order of the "Departamento de Publicaciones del CSIC" ■ Credit card, VISA / Master Card / Eurocard / 4B Number: _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ Expiration date: _ _ / _ _ ■ (From abroad) SWIFT/BIC CODE: BSCHESMM. / IBAN NUMBER: ES83 0049 5117 2621 1010 5188 Distribution and sale: Departamento de Publicaciones del CSIC C/ Vitruvio, 8 E-28006 Madrid Telephones: +34 915 612 833 / +34 915 681 619/620/640 Fax: +34 915 629 634 E-mail: [email protected] www. publicaciones.csic.es Signature ________________________________________________________________________________________________ portada y contra 68(1) NOVEDADES2:portada y contra 66(1).qxd 16/06/2011 11:22 Página 1 ISSN: 0211-1322 Zamora, N.A. El caso de Lonchocarpus costaricensis (Leguminosae, Papilionoideae), una especie endémica de Costa Rica: un complejo taxonómico-nomenclatural, y una nueva especie / The case of Lonchocarpus costaricensis (Leguminosae, Papilionoideae), an endemic species of Costa Rica: a taxonomic-nomenclatural complex, and a new species ................................................................ Cano-Maqueda, J. & Talavera, S. A taxonomic revision of the Campanula lusitanica complex (Cam pa nulaceae) in the Western Mediterranean region / Una revisión taxonómica del com plejo Campanula lusitanica (Campanulaceae) en la región occidental mediterránea ........................................................................................................................................... Venhuis, C. & Oostermeijer, J.G.B. Distinguishing colour variants of Serapias perez-chiscanoi (Orchidaceae) from related taxa on the Iberian Peninsula / Distinción de variantes en color de Serapias perezchiscanoi (Orchidaceae) en relación con táxones de la Península Ibérica .......................................... Lado, C., Wrigley de Basanta, D. & Estrada-Torres, A. Biodiversity of Myxomycetes from the Monte Desert of Argentina / Biodiversidad de Myxomycetes en el Desierto de Monte (Argentina) ............... Peñas, J., Lorite, J., Alba-Sánchez, F. & Taisma, M.A. Self-incompatibility, floral parameters, and pollen characterization in the narrow endemic and threatened species Artemisia granatensis (Asteraceae) / Autoincompatibilidad, parámetros florales y caracterización de polen en la especie endémica y amenazada Artemisia granatensis (Asteraceae) ...................................................................................... Schneider, A.A. & Boldrini, I.I. Microsculpture of cypselae surface of Baccharis sect. Caulopterae (Asteraceae) from Brazil / Microescultura de la superficie de las cipselas de Baccharis sect. Caulopterae (Asteraceae) de Brasil ...................................................................................................................... Domínguez-Álvarez, S., Rico, J.M. & Gil-Rodríguez, M.C. Photosynthetic response and zonation of three species of Gelidiales from Tenerife, Canary Islands / Respuestas fotosintéticas y zonación de tres especies de Gelidiales de Tenerife, Islas Canarias ...................................................................... http://rjb.revistas.csic.es N.º 1 enero-junio 2011 Madrid (España) ISSN: 0211-1322 Madrid SUMARIO / CONTENTS Volumen 68 7/14 15/47 2011 Madrid (España) N.º 1 enero-junio 2011 49/59 61/95 97/105 107/116 117/124 Volumen 68 N.º 1 Anales del Jardín Botánico de Madrid Volumen 68 REAL JARDÍN BOTÁNICO www.publicaciones.csic.es CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS