Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
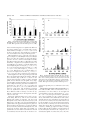
ECOLOGY AND POPULATION BIOLOGY Effect of Allegheny Mound Ant (Hymenoptera: Formicidae) Presence on Homopteran and Predator Populations in Michigan Jack Pine Forests D. B. BISHOP1 AND C. M. BRISTOW Department of Entomology, Michigan State University, East Lansing, MI 48824 Ð1115 Ann. Entomol. Soc. Am. 94(1): 33Ð40 (2001) KEY WORDS Formica exsectoides, Cinara aphids, ants, scale insects, mutualism, homopteran community TENDING HONEYDEW-PRODUCING HOMOPTERA by ants can provide many beneÞts to the tended homopterans, including sanitary removal of excess honeydew (Strickland 1947, Majer 1982), increased growth rate (Banks and Nixon 1958), decreased development time (El-Ziady 1960, Bristow 1984), or even a transfer of parental care to the tending ants (Bristow 1983). The primary beneÞt, though, appears to be protection from predators and parasitoids (Way 1963, Buckley 1987, Bristow 1991). In agricultural and forest settings, the exclusion of tending ants can result in an increase of natural enemies and a resulting decrease in homopterans (DeBach et al. 1951, Bartlett 1961 Bradley 1973, Tilles and Wood 1982, Stechmann et al. 1996). The effectiveness of ant protection, however, appears to depend on the aggressiveness of the ants involved; more aggressive ants are likely to provide better protection for the tended homopterans (Bristow 1984, Buckley and Gullan 1991). Although many of these studies address the effect mutualist ants can have on a single homopteran species, fewer studies indicate how ant mutualists can inßuence the relative abundance of different members of the homopteran community (Addicott 1986, 1 Department of Biology, Concordia College, Moorhead, MN 56562. Bronstein 1994). In one example, the removal of two mutualist ants, Dolichoderus taschenbergi (Mayer) and Formica obscuripes Forel, resulted in the extinction of four myrmecophilous aphid species on jack pines near the destroyed ant nests (Bradley and Hinks 1968). The interplay between tended and untended species may be important, because many homopterans are not tended by ants. A survey of 479 aphid species from the Rocky Mountain region indicated that 75% do not associate with ants (Bristow 1991). Ants will often prey upon nonmyrmecophilous homopterans (Skinner and Whittaker 1981, Mahdi and Whittaker 1993). In some areas of the jack pine, Pinus banksiana (L.), forests of north-central Michigan, Allegheny mound ants, Formica exsectoides Forel, are the most abundant ant encountered, forming large, locally abundant, but patchy populations (Bristow et al. 1992). Formica exsectoides have been described as “the most common mound building ant in North America” (Andrews 1926), and they readily associate with several species of honeydew producing Homoptera (Cory and Haviland 1938, Haviland 1947, Dimmick 1951). Where established, they can become extremely numerous (Andrews 1925, Cory and Haviland 1938, Bristow et al. 1992) and dominate the ant fauna (Hölldobler and 0013-8746/01/0033Ð0040$02.00/0 䉷 2001 Entomological Society of America Downloaded from http://aesa.oxfordjournals.org/ by guest on October 21, 2016 ABSTRACT The impact of different densities of Allegheny mound ant, Formica exsectoides Forel, populations on myrmecophilous (tended) and nonmyrmecophilous (untended) aphid and scale species and their potential predators was examined in jack pine forests of north-central Michigan. A summer-long survey indicated that areas with large populations of F. exsectoides had larger populations of the aphid Cinara banksiana Pepper & Tissot and the soft scale Toumeyella parvicornis (Cockerell), two obligate myrmecophiles, than areas with low or no F. exsectoides populations. Cinara ontarioensis Bradley was also obligately tended by ants but did not show a signiÞcant response to F. exsectoides densities, having a patchy distribution. In contrast, in areas without F. exsectoides, these homopterans were replaced by the woolly aphid Schizolachnus piniradiatae (Davidson), a species that never forms ant associations. Mound ant exclusion experiments and observations indicated that F. exsectoides preyed upon S. piniradiatae, potentially reducing their numbers in areas with this ant. Aphid and scale predators showed a mixed response to F. exsectoides presence: densities of lacewing larvae and salticid spiders were inversely related to F. exsectoides populations, whereas mirid populations did not show a response to different F. exsectoides densities. This study suggests that the presence of large populations of aggressive, honeydew seeking F. exsectoides can shift the homopteran community from one composed predominantly of nonmyrmecophilous species to one composed of myrmecophilous species. 34 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA Materials and Methods Survey. Fifteen plots (30 by 30 m) were selected in the Huron-Manistee National forests of north-central Michigan. The 15 plots were grouped into three treatments based on Allegheny mound ant abundance, high-ant, low-ant, and no-ant treatments. Each ant abundance plot was replicated Þve times. All Þve replicate sites were within a 6 by 15-km area of southeast Crawford and southwest Oscoda counties but were separated by a minimum of 2 km. All replicate sites for plots were in jack pine stands and chosen to be as similar as possible in stand age, plant composition and soil moisture. In selecting sites, we Þrst scouted jack pine forest areas with Allegheny mound ant populations. High-ant plots were then placed in areas where mounds were at a high density (mean mound density per plot ⫾ SE: 15.4 ⫾ 3.8), whereas low-ant plots were placed near the edges of mound distribution, areas where mound density was decreasing (mean mound density per plot ⫾ SE: 3.8 ⫾ 1.5). If possible, we also marked off a no-ant plot, not closer than 100 m to the nearest ant mound. In most cases this represented a cross section through and beyond the local ant population; however, two of the no-ant plots were between 2 and 6 km from their paired ant-plots because adjacent stands without ants had been cut within the last 10 Ð15 yr. All plot sites had 40- to 72-yr-old jack pine as the dominant tree, with blueberry (Vaccinium angustifolium Aiton), bracken fern (Pteridium sp.), and sweet-fern [Compotonia peregrina (L.) Coulter] forming the majority of understory plants. All jack pine saplings between 1.5 and 3 m tall were identiÞed in each ant-density plot. Taller trees would have required the use of tall ladders and much more time to thoroughly examine higher branches than required for the shorter saplings. Fifteen of these trees were then randomly selected in each plot. However, one site had only seven trees within this size range in the high-ant plot. For this site, all three ant-density plots had seven trees marked for the study. Thus, a total of 67 trees was marked for each of the antdensities. The mean distance (⫾SE) from each marked tree to the nearest mound in the high-ant and low-ant density plots was 4.38 ⫾ 0.29 m and 12.48 ⫾ 0.73 m, respectively. To reduce the time needed to examine 201 trees, a half side of each tree was randomly chosen. We conducted a biweekly census of the same side beginning the week of 8 June through 5 August and an end of season census in early September. For each census period, an absolute sample (number of animals per unit habitat, i.e., one-half side of tree) was taken of aphids, scales, ants, predators, and aphid mummies or parasitized scales (Southwood 1979). The time spent examining each tree depended on its size and number of branches and the number of animals on the tree. Representative specimens were collected for identiÞcation. Voucher specimens were deposited in the Michigan State University Entomology Museum. Homopteran and predator species having large enough numbers for statistical analysis were analyzed as a split-plot repeated measures analysis of variance (ANOVA) with ant densities (Þxed effect) the among-site factor and census period (Þxed effect) the within-site factor (SAS Institute 1995, Zar 1996). Site (random effect) was nested within the ant densities treatment and used to test the ant density effect. Because our main question concerned how these arthropod populations varied between different antdensities, we took the mean number of each arthropod species found on all the surveyed trees for each antdensity plot at each site and used this mean in the analysis. Data were tested for the main effect of antdensity on homopteran and predator numbers and the ant-density by census period interaction. Because some homopteran and predator species did not appear until the third sampling date (8 July), earlier dates could not be used in the repeated measures because Downloaded from http://aesa.oxfordjournals.org/ by guest on October 21, 2016 Wilson 1990). This ant exhibits no colony boundaries (Hölldobler and Wilson 1990), so trees can be patrolled by a large number of ants from different mounds. F. exsectoides workers are very aggressive, attacking and overpowering almost every arthropod they encounter, including honeydew-seeking yellow jackets (Vespa sp., unpublished data). We hypothesized that the presence of a large population of aggressive, mutualist ants in an area could cause the Homoptera community to be made up predominately of tended homopterans, whereas areas without these ants would be dominated by nontended species by providing beneÞts to myrmecophilous homopterans and suppressing nonmyrmecophilous homopterans by preying upon them. Additionally, the aggressive nature of F. exsectoides could also act to lower the numbers of potential homopteran predators in areas with these ants. Preliminary observations documented differences in the homopteran community on jack pine saplings between areas with and without F. exsectoides. Two myrmecophilous homopterans, the aphid Cinara banksiana and the pine tortoise scale, Toumeyella parvicornis (Cockerell), appeared to be very common in areas with F. exsectoides but virtually absent from other areas. Conversely, the nonmyrmecophilous woolly aphid Schizolachnus piniradiatae (Davidson) was more common in areas without F. exsectoides. To determine whether a repeatable pattern existed between the presence of F. exsectoides, aphid and scale species, and potential predators of homopterans, we conducted surveys on jack pine saplings at sites with high mound ant densities (hereafter referred to as high-ant density), areas with low mound ant densities (low-ant density), and areas that contained no F. exsectoides (no-ants). This survey was conducted during the summer of 1993. In addition to the survey, we performed experiments in 1994 to determine whether F. exsectoides attacks the S. piniradiatae. Because other studies have addressed the positive role tending ants have for Homoptera (Way 1963, Boucher et al. 1987, Bishop 1998), no parallel experiments were conducted to determine the nature of the interaction between F. exsectoides and tended homopterans. Vol. 94, no. 1 January 2001 BISHOP AND BRISTOW: HOMOPTERAN AND PREDATOR POPULATIONS WITH ANTS Results Survey. In high-ant plots, the overall mean Allegheny mound ant activity (ants encountered on sample side of tree) during the Þve 2-wk census periods was 6.2 ⫾ 1.2 (⫾SE), and the mean activity in low-ant plots was 2.8 ⫾ 1.1 (⫾SE) (Fig. 1). However, by late July the mean number of ants between the high and Fig. 1. Mean ⫾ SE number of F. exsectoides found per half side of each sampled tree in high-ant areas and low-ant areas. n ⫽ 67 trees. low-ant plots was similar (Fig. 1). Other ant species were an order of magnitude less abundant than F. exsectoides (other ant species overall mean ⫾ SE: 0.2 ⫾ 0.04, 0.7 ⫾ 0.14, and 0.5 ⫾ 0.15 for high, low, and no-F. exsectoides areas, respectively). Other ant taxa included Crematogaster spp. Lund, F. fusca L., F. obscuripes Forel, Camponotus spp. Mayr, and at least three unidentiÞed species. None of these species was common. Aphids. We found Þve aphid species feeding on jack pine during this survey. Three aphid species with large enough numbers for statistical analysis were C. banksiana (Pepper & Tissot), C. ontarioensis Bradley, and S. piniradiatae. C. pergandei (Wilson), a large solitary aphid, occurred in very low numbers, about one aphid for every 10 trees in the high and no-ant areas, and one aphid for every Þve trees in the low-ant areas. The other Cinara aphid was not identiÞed to species. The most common aphid present in high ant areas was C. banksiana (Fig. 2A). It Þrst was found on second and third year growth very early in the season but moved to new shoots when they began to elongate. By late-season most aphids moved back to older growth. This aphid was nearly always observed with ant attendants; the only exceptions were alates that had newly settled in no-ant areas. Established colonies of this species were found in all Þve of the high-ant plots, four of the low-ant plots, and two of the no-ant plots. Cinara banksiana numbers were signiÞcantly larger in high-ant plots than no-ant plots (F ⫽ 16.37; df ⫽ 1, 8; P ⫽ 0.004) and the interaction between census date and ant-density was also signiÞcant (F ⫽ 5.69; df ⫽ 4, 32; P ⫽ 0.001). C. banksiana populations in high-ant areas grew much faster than those in the low-ant areas, but by 5 August they declined to levels similar to the low-ant areas. C. banksiana numbers did not signiÞcantly differ for any of the other ant-density comparisons. The second most common aphid in high-ant areas was C. ontarioensis (Fig. 2B). We only observed this aphid in the presence of tending ants. Population Downloaded from http://aesa.oxfordjournals.org/ by guest on October 21, 2016 their variance on those dates was zero. Mean arthropod numbers were log-transformed to normalize error distributions. Untransformed data are presented in Þgures. Spearman rank correlations were performed on the different homopteran species and predators (Sokal and Rohlf 1981) to test whether different homopteran species may be associated with each other or with predators. The correlation was tested on each sampling date when appropriate. Trees that had both C. banksiana and C. ontarioensis were not included in the analysis to eliminate confounding effects. Schizolachnus piniradiatae Experiments. To determine the types of interactions between the untended S. piniradiatae and F. exsectoides, we used the high-ant density areas at three of the survey sites to conduct ant-exclusion experiments and timed observations during 1994. We randomly selected 10 trees that had colonies of C. banksiana with tending ants on at least two branches at each site. A total of 60 branches was used (10 trees ⫻ two treatments ⫻ three sites). For exclusion branches, one branch was randomly selected and all ants were removed. A 4-cm band of Tanglefoot (Grand Rapids, MI) was applied around the base of this branch to exclude ants, and any branches touching this branch were clipped back. The second branch was left as an untreated control. Each branch had three S. piniradiatae placed on the tip of the branch, away from the C. banksiana colony. For branches with ants, data were gathered on the interactions between S. piniradiatae and ants for 15 min. After 72 h, the numbers of S. piniradiatae remaining on branches were recorded. Cinara banksiana aphids were also placed on other branches with C. banksiana colonies and ants as a comparison to the S. piniradiatae. Newly placed C. banksiana aphids were observed for 15 min, but because they mixed in with the other C. banksiana aphids we could not assess their survival 72 h later. A survival analysis was conducted comparing the survival times (time till removal by ants) of S. piniradiatae to C. banksiana for the 15-min observations. A split-plot ANOVA, with ant treatment (ants present or excluded) and site as Þxed effects, was used to test for a difference in the percent change ([S. piniradiatae number at 72 h ⫺ S. piniradiatae number at start]/S. piniradiatae number at start) in S. piniradiatae numbers after 72 h between the two ant treatments. Trees (random effect) were nested within site and used to test the ant treatment effect. All analyses for both the survey and S. piniradiatae experiments were conducted using the JMP statistical package (SAS Institute 1995). 35 36 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA Vol. 94, no. 1 Fig. 2. Population trends in high-, low-, and no- mound ant density plots across six census dates for (A) C. banksiana, (B) C. ontarioensis, and (C) S. piniradiatae. Sixty-seven half sides of trees were sampled in Þve plots for each ant density. No data were collected for low-ant density plots on 6 September. Bars represent mean aphid numbers per tree (⫾SE). numbers increased later in the season than C. banksiana, but C. ontarioensis never reached the numbers C. banksiana attained. Also it had a more patchy distribution than C. banksiana, occurring in fewer sites (four high-ant plots and one plot each of both the lowand no-ant plots) and on fewer trees within a site. Most individuals of this species occurred in the highand low-ant plots at one site. At another high-ant site it was found on only one tree and had disappeared by 22 July. C. ontarioensis numbers did not signiÞcantly differ between any of the ant-density plots. The last aphid common enough for statistical tests was the nontended, pine woolly aphid, S. piniradiatae. Woolly aphid populations increased late in the season and were found primarily in no-ant plots (Fig. 2C). This aphid was found in all Þve of the no- and low-ant plots, and in four of the high-ant plots, albeit at lower numbers. Population size was signiÞcantly larger in no-ant plots compared with the high-ant density plots (F ⫽ 9.13; df ⫽ 1, 8; P ⫽ 0.02) and the interaction between ant density and time was also signiÞcant (F ⫽ 6.04; df ⫽ 2, 16; P ⫽ 0.01). Woolly aphid numbers also differed signiÞcantly between the high-ant and lowant density plots (F ⫽ 8.22; df ⫽ 1, 8; P ⫽ 0.02). No difference in S. piniradiatae numbers was detected between the low-ant density and no-ant plots. Scales. Toumeyella parvicornis (Cockerell), the pine tortoise scale, numbers were signiÞcantly greater in high-ant density plots than in no-ant density plots (Fig. 3, P ⫽ 7.37; df ⫽ 1, 8; P ⫽ 0.03), and this became larger over the season (F ⫽ 5.21; df ⫽ 4, 32; P ⫽ 0.002). Pine tortoise scales were found in all Þve high-ant density plots and only two plots of both the low- and no-ant density plots. Crawlers were released in late June and early July, causing scale numbers to increase greatly in high-ant density plots and to a lesser extent in the low-ant density and no-ant plots (Fig. 3). However, by 6 September second-instar scales had declined by ⬇78% in the high ant plots, leaving scale abundance close to that observed at the start of the season. No other signiÞcant differences in scale numbers were detected between other ant-density comparisons. The association between F. exsectoides and tended homopterans did not necessarily remain constant throughout the summer. Ant numbers on trees with T. parvicornis tended to ßuctuate sharply during and after crawler release (24 June and 22 July, respectively; Fig. 4). F. exsectoides numbers also declined on trees with C. banksiana in late July, but ant numbers for both groups had rebounded by early August (Fig. 4). No signiÞcant correlation was detected between any of the homopteran populations. Predators. Three principal aphid predators were encountered and analyzed in this survey: lacewings, salticid spiders, and mirids (Fig. 5). Lacewings were signiÞcantly more common in the no-ant plots than in the high-ant density plots (F ⫽ 5.91; df ⫽ 1, 8; P ⫽ 0.04) and the interaction with time was also signiÞcant (F ⫽ 2.89; df ⫽ 4, 32; P ⫽ 0.04). No other comparisons Downloaded from http://aesa.oxfordjournals.org/ by guest on October 21, 2016 Fig. 3. Population trends in high-, low-, and no- mound ant density plots across six census dates for T. parvicornis. Sixty-seven half sides of trees were sampled in Þve plots for each ant density. No data were collected for low-ant density plots on 6 September. Bars represent mean scale numbers per tree (⫾SE). January 2001 BISHOP AND BRISTOW: HOMOPTERAN AND PREDATOR POPULATIONS WITH ANTS 37 Fig. 4. Ant numbers found on half-sides of 67 trees that had either T. parvicornis colonies or C. banksiana colonies but not both. Bars represent mean ant number per tree (⫾SE). Fig. 5. Population trends in (A) high-, (B) low-, and (C) no-mound ant density plots across six census dates for the three most common predators observed. Sixty-seven half sides of trees were sampled in Þve plots for each ant density. No data were collected for low-ant density plots on 6 September. Bars represent mean predator numbers per tree. parasitoid ovipositing in a second-instar scale in early August. Schizolachnus piniradiataes Experiments. Observations indicated that, when encountered by ants, S. piniradiataes were more likely to be attacked and carried off than newly placed C. banksiana (2 ⫽ 5.36, df ⫽ 1, P ⫽ 0.02). Ten percent of the S. piniradiatae were removed within 2.5 min of being placed on shoots, and within 15 min, 24% had been removed by ants. The majority of S. piniradiatae encountered by an ant (21 of 26 encounters) were attacked and removed by the ant. Only one was attacked and released after the attack. The remaining four fell off the branch during the attack. Most S. piniradiatae avoided encounter by climbing up adjacent needles, a feeding behavior that also allowed them to remain undetected by ants during the observation. By comparison, of 63 C. banksiana aphids encountered by ants, only nine Downloaded from http://aesa.oxfordjournals.org/ by guest on October 21, 2016 between ant-density plots were signiÞcantly different. Lacewings numbers were correlated with S. piniradiatae numbers on 22 July (rs ⫽ 0.39, n ⫽ 67, P ⫽ 0.001) in the no-ant plots. No other signiÞcant correlations were found between lacewings and homopterans. Mirid populations were composed of at least two species, Pilophorus urhlei Knight and P. furvus Knight. These were the only predators we detected that appeared to be more common in the high-ant density plots than in other plots (Fig. 5), although none of the ant-density comparisons were signiÞcant at the 0.05 level. Mirids were positively correlated with C. banksiana in high-ant plots on 5 August (rs ⫽ 0.310, n ⫽ 67, P ⫽ 0.009) and in low-ant plots on 22 July (rs ⫽ 0.422, n ⫽ 67, P ⬍ 0.001) and 5 August (rs ⫽ 0.265, n ⫽ 67, P ⫽ 0.030). Mirids were also correlated with C. ontarioensis on 22 July in low-ant sites (rs ⫽ 0.393, n ⫽ 67, P ⫽ 0.001). No other correlations were signiÞcant. Salticid spiders were the only predator present at all three ant-densities throughout the summer, although at low densities (Fig. 5). On two occasions we witnessed salticid spiders feeding in small, untended aphid colonies. In both cases, the aphid colony disappeared within 3 d. Salticid numbers were signiÞcantly lower in high-ant density plots than in both the lowant density (F ⫽ 7.12; df ⫽ 1, 8; P ⫽ 0.03) and no-ant plots (F ⫽ 11.27; df ⫽ 1, 8; P ⫽ 0.01). The interaction between census dates and ant-density treatments were not signiÞcant in either case, indicating that these differences did not change over time. No signiÞcant correlations between homopterans and salticids were detected. Parasitoids. Aphid mummies were rarely encountered during this survey. The no-ant area had the highest total number of mummies (34) discovered through the season, followed by the low-ant (23) and high-ant area (18). One-year-old scales that had been parasitized became apparent during the two July sampling periods by the appearance of exit holes. Out of 118 1-yr-old scales observed in the high ant sites, 23% developed exit holes during this period. Only one parasitized scale was found in each of the low-ant and no-ant density sites. We observed one incidence of a 38 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA were removed by ants within 15 min. Although low, the number of C. banksiana carried off by ants may be inßated because the aphidÕs behavior was altered by this experiment. Aphids were attacked when they did not stop moving when antennated by an ant. However, even nonmoving S. piniradiatae were always attacked immediately when contacted by an ant. After 72 h, S. piniradiatae numbers decreased signiÞcantly more when ants were present (F ⫽ 36.60; df ⫽ 1, 27; P ⬍⬍ 0.001); S. piniradiatae on branches with ants declined by 81% ⫾ 0.07 (mean ⫾ SE) compared with a decline of only 4% ⫾ 0.11 (mean ⫾ SE) for S. piniradiatae on branches without ants. Discussion distribution was also detected by Bradley and Hinks (1968) for this aphid. Whether this pattern of distribution is due to the aphidÕs biology, the tending ants, or another factor needs more study. S. piniradiatae showed a pattern opposite C. banksiana and T. parvicornis. It appeared to be sensitive to high populations of F. exsectoides but could tolerate the lower ant-densities found on the edges of ant areas. F. exsectoides were constantly patrolling shoots and needles of trees. The large number of ants would ensure that many alate S. piniradiatae that settled in ant areas would be found and removed by these foraging ants. Despite the higher numbers of predators in the no-ant areas, S. piniradiatae were relatively numerous in those areas. This aphid secreted a waxy covering, which may have helped protect it or camoußage it from enemies, although some lacewings are capable of feeding on such aphids (Eisner et al. 1978). It also responded differently to close proximity of foreign objects. Whereas, Cinara aphids would twist, kick, or run from an unfamiliar object, S. piniradiatae did not move unless physically touched. This behavior may have allowed them to remain cryptic to visual hunters like salticids. Further studies are needed to determine what traits allow this aphid to succeed in no-ant areas. The association between ants and myrmecophilous homopterans, especially with T. parvicornis, did not remain constant throughout the season. The decrease observed in late July of ant numbers on trees with only scales may be a response of the ants in part to the biology of pine tortoise scales. Rabkin and Lejeune (1954) reported that female pine tortoise scales do not begin producing honeydew until after mating, 2Ð3 wk after the crawler stage. This decline in honeydew production for these few weeks could result in the abandonment of scales by ants, potentially exposing scales to their natural enemies. We did record a drop in the number of ants tending scales on 22 July, when early-instar scale populations were at their highest. It is unclear whether this drop was due to scale biology or some other factor(s) because the number of ants on trees with only C. banksiana colonies also declined at this time. However, 2 wk later (8 August) ant numbers on trees with scales had increased sharply, when honeydew production would have increased. Like the homopterans, predators appeared to differ in their sensitivity to ant-densities and in their response to ants. Lacewings appeared to be the most sensitive to disruption by F. exsectoides. Slow moving lacewing larvae on shoots are easily caught by F. exsectoides workers (Bishop 1998). Salticid numbers were also lower in the high-ant areas than in the no-ant areas, although their low densities overall probably had little biological impact on aphid populations. Other recent studies have also indicated fewer spiders in areas with foraging ants (Halaj et al. 1997, Heikkinen 1999). Salticids were probably rarely directly preyed upon by ants. On a number of occasions, we observed salticids escape detection by backing away and moving to the opposite side of the shoot or jumping to adjacent shoots when Downloaded from http://aesa.oxfordjournals.org/ by guest on October 21, 2016 The results of this study indicate that the homopteran community on jack pine saplings differed between areas with F. exsectoides and areas without this ant. Ant-tended homopteran species were most abundant in high-F. exsectoides density areas, but the nontended S. piniradiatae was dominant in areas without F. exsectoides. This distribution would be expected if myrmecophilous Homoptera were dependent on these ants for certain beneÞts. Other species of ants (e.g., Crematogaster spp. and F. obscuripes) in plots either with low or zero densities of F. exsectoides were observed tending the few aphids and scales in these areas. Similar observations, where different ant species tend aphids when another ant species is not present, have been reported (Addicott 1979). However, except for one population of scales tended by F. obscuripes, these homopteran populations disappeared by the end of the season. Due to its high density and aggressiveness, F. exsectoides appeared to act as a dominant species for homopterans in these areas with low densities of other ant species. An alternative hypothesis is that F. exsectoides were tracking the honeydew producing Homoptera. We reject this hypothesis because F. exsectoides colonies are stationary in large mounds that remain year after year (Andrews 1926, Bristow et al. 1992) even when aphid populations were reduced by late frosts (unpublished data). Other studies (Bradley and Hinks 1968, Bishop 1998) have shown that the exclusion of F. exsectoides from tended aphid colonies resulted in the disappearance of the aphid colonies. This survey suggests that there may be a critical density of F. exsectoides needed before ant-tended homopterans succeed. C. banksiana reached the largest population sizes in the high-ant density sites, and their population grew faster than those in the low-ant sites and T. parvicornis appeared to do well only in areas with large populations of F. exsectoides. A larger population of ants would Þnd newly settled or dispersed homopterans more quickly, insuring their survival (Addicott 1978). Cinara ontarioensis showed a different pattern with their largest populations found in the low-ant areas. However, this is hard to interpret because they had a more patchy distribution than C. banksiana, with most of their numbers due to just one site. A similar, patchy Vol. 94, no. 1 January 2001 BISHOP AND BRISTOW: HOMOPTERAN AND PREDATOR POPULATIONS WITH ANTS Acknowledgments We thank R. Mech (Michigan Department of Natural Resources) for information on Þeld sites and lodging, and Miles Howard for his assistance in the Þeld. We also thank the Systematic Entomology Laboratory-TSU for the identiÞcation of the Cinara aphids and two anonymous reviewers for their helpful comments on earlier drafts. This research was funded in part by a grant from the USDA (No. 91-3373026190) and from the Department of Entomology at Michigan State University Hutson Fund. References Cited Addicott, J. F. 1978. Niche relationships among species of aphids feeding on Þreweed. Can. J. Zool 56: 1837Ð1841. Addicott, J. F. 1979. A multispecies aphid-ant association: density dependence and species-speciÞc effects. Can. J. Zool. 57: 558Ð569. Addicott, J. F. 1986. On the population consequences of mutualism, pp. 425Ð436. In J. Diamond and T. J. Case [eds.], Community ecology. Harper and Row, New York. Andrews, E. A. 1925. McCookÕs ant mounds in Pennsylvania, revisited (Hymen: Formicidae). Entomol. News 36: 173Ð179. Andrews, E. A. 1926. Sequential distribution of Formica exsectoides Forel. Psyche (Lexington) 33: 127Ð150. Banks, C. J., and H. L. Nixon. 1958. Effects of the ant, Lasius niger L., on the feeding and excretion of the bean aphid, Aphis fabae Scop. J. Exp. Biol. 35(4): 703Ð711. Bartlett, B. R. 1961. The inßuence of ants upon parasites, predators and scale insects. Ann. Entomol. Soc. Am. 54: 543Ð551. Bishop, D. B. 1998. Differential effects of the Allegheny mound ant, Formica exsectoides Forel, on aphid, scale and predator populations and their interactions in jack pine forests. Ph.D. dissertation, Michigan State University, East Lansing, MI. Boucher, D. H., S. James, and K. S. Keeler. 1987. The ecology of mutualism. Annu. Rev. Ecol. Syst. 13: 315Ð347. Bradley, G. A. 1973. Effect of Formica obscuripes (Hymenoptera: Formicidae) on the predator-prey relationship between Hyperaspis congressis (Coleoptera: Coccinellidae) and Toumeyella numismaticum (Homoptera: Coccidae). Can. Entomol. 105: 1113Ð1118. Bradley, G. A., and J. D. Hinks. 1968. Ants, aphids, and jack pine in Manitoba. Can. Entomol. 100: 40Ð50. Bristow, C. M. 1983. Treehoppers transfer parental care to ants: a new beneÞt of mutualism. Science 220: 532Ð533. Bristow, C. M. 1984. Differential beneÞts from ant attendance to two species of Homoptera on New York ironweed. J. Anim. Ecol. 53: 715Ð726. Bristow, C. M. 1991. Why are so few aphids ant-tended?, pp. 105Ð171. In C. R. Huxley and D. F. Cutler [eds.], Antplant interactions. Oxford University Press, New York. Bristow, C. M., D. Cappert, N. J. Campbell, and A. Heise. 1992. Nest structure and colony cycle of the Allegheny mound ant, Formica exsectoides Forel (Hymenoptera: Formicidae). Insectes Soc. 39: 385Ð402. Bronstein, J. L. 1994. Our current understanding of mutualism. Q. Rev. Biol. 69: 31Ð51. Buckley, R. 1987. Ant-plant-homopteran interactions. Adv. Ecol. Res. 16: 53Ð85. Buckley, R., and P. Gullan. 1991. More aggressive ant species (Hymenoptera: Formicidae) provide better protection for soft scales and mealybugs (Homoptera: Coccidae, Pseudococcidae). Biotropica 23(3): 282Ð286. Cory, E. N., and E. E. Haviland. 1938. Population studies of Formica exsectoides Forel. Ann. Entomol. Soc. Am. 31: 50Ð57. DeBach, P., C. A. Fleschner, and E. J. Dietrick. 1951. A biological check method for evaluating the effectiveness of entomophagous insects. J. Econ. Entomol. 44: 763Ð766. Downloaded from http://aesa.oxfordjournals.org/ by guest on October 21, 2016 ants were approaching. Despite sharing similar prey, Halaj et al. (1997) did not Þnd sufÞcient evidence to support exploitative competition between hunting spiders and Camponotus spp. Their observations suggest that interference competition between hunting spiders and ants may cause spiders to disperse from ant areas. A similar situation may exist between salticids and F. exsectoides. Various mirid species that feed on aphids, including P. furvus and P. uhrlei, have been reported to associate with ant-tended aphid colonies (Bradley and Hinks 1968, Wheeler 1991), a result we also found. This association may be due to a larger population of aphids present in areas with F. exsectoides, whereas the nonant areas had small, sporadic aphid populations that did not increase until late in the season. Mirids were observed close to aphid colonies, rushing in when ants were not present, and piercing an aphid and feeding until an ant appeared. Mirids would run up needles or shoots to escape any ants that came too close (⬇3 cm). These behaviors may allow the mirids to specialize in preying on tended aphid colonies. Any predator that can overcome or avoid the ant defenses would have access to the rich food resource provided by the tended aphids. Aphid parasitoids appeared not to play a signiÞcant role in this system. However, their low numbers may be due in part to the wandering of parasitized Cinara aphids. We typically found mummies on shoots away from the main colony. Scale parasitoids may be of more importance in this system because almost 25% of 1-yr-old scales we observed were parasitized. However, if scales are parasitized in late summer, the subsequent consumption of these parasitized scales by predators could decrease their importance relative to the predators. (Rosenheim 1998). In conclusion, F. exsectoides may play a keystone role as a mutualist predator for homopterans and their natural enemies in jack pine forests. Those homopterans that have the strongest mutualistic relation with F. exsectoides appeared to beneÞt from its large numbers and aggressiveness. As a predator, F. exsectoides preyed on both nonmyrmecophilous homopterans and homopteran predators such as lacewings. But mirids appear to beneÞt from its tending of aphids. We speculate that the presence of the F. exsectoides shifts the composition of the Homoptera community from one dominated by nonmyrmecophilous homopterans and generalist predators to one dominated by myrmecophilous homopterans and predators that can circumvent ant defenses. These results, along with others (e.g., Bradley and Hinks 1968; Skinner and Whittaker 1981; Warrington and Whittaker 1985a, 1985b), imply that large populations of aggressive mound or wood ants play important roles in shaping both herbivore and predator communities in forest systems. 39 40 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA ant (Formica rufa) and some tree-canopy herbivores. J. Anim. Ecol. 50: 313Ð326. Sokal, R. R., and F. J. Rohlf. 1981. Biometery. Freeman, New York. Southwood, T.R.E. 1979. Ecological methods with particular reference to the study of insect populations, 2nd ed. Methuen, London. Stechmann, D. H., W. Volkl, and P. Stary. 1996. Ant-attendance as a critical factor in the biological control of the banana aphid Pentalonia nigronervosa Coq. (Hom. Aphididae) in Oceania. J. Appl. Entomol. 120: 119 Ð123. Strickland, A. H. 1947. Coccids attacking cacao (Theobroma cacoa, L.), in West Africa, with descriptions of Þve new species. Bull. Entomol. Res. 38: 497Ð523. Tilles, D. A., and D. L. Wood. 1982. The inßuence of carpenter ant (Camponotus modoc) (Hymenoptera: Formicidae) attendance on the development and survival of aphids (Cinara spp.) (Homoptera: Aphididae) in a giant sequoia forest. Can. Entomol. 114: 1133Ð1142. Warrington, S., and J. B. Whittaker. 1985a. An experimental Þeld study of different levels of insect herbivory induced by Formica rufa predation on sycamore (Acer pseudoplatanus). I. Lepidoptera. J. Appl. Ecol. 22: 775Ð785. Warrington, S., and J. B. Whittaker. 1985b. An experimental Þeld study of different levels of insect herbivory induced by Formica rufa predation on sycamore (Acer pseudoplatanus). II. Aphidoidea. J. Appl. Ecol. 22: 787Ð796. Way, M. J. 1963. Mutualism between ants and honeydewproducing Homoptera. Annu. Rev. Entomol. 8: 307Ð344. Wheeler, Jr., A. G. 1991. Plant bugs of Quercus ilicifolia: myriads of mirids (Heteroptera) in pitch pine-scrub-oak barrens. J. N.Y. Entomol. Soc. 99: 405Ð 440. Zar, J. H. 1996. Biostatistical Analysis. Prentice-Hall, Upper Saddle River, NJ. Received for publication 19 November 1999; accepted 11 September 2000. Downloaded from http://aesa.oxfordjournals.org/ by guest on October 21, 2016 Dimmick, J. F. 1951. An ecological study of Formica exsectoides. Ill. Acad. Sci. Trans. 44: 197Ð204. Eisner, T., K. Hicks, M. Eisner, and D. S. Robson. 1978. “Wolf-in-sheepÕs-clothing” strategy of a predaceous insect larvae. Science 199: 790 Ð794. El-Ziady, S. 1960. Further effects of Lasius niger L. on Aphis fabae Scopoli. Proc. R. Entomol. Soc. Lond. 35(1Ð3): 30 Ð38. Halaj, J., D. W. Ross, and A. R. Moldenke. 1997. Negative effects of ants foraging on spiders in Douglas-Þr canopies. Oecolgia 109: 313Ð322. Haviland, E. E. 1947. Biology and control of the Allegheny mound ant. J. Econ. Entomol. 40: 413Ð 419. Heikkinen, M. W. 1999. Negative effects of the western thatching ant (Formica obscuripes) on spiders (Araneae) inhabitating big sagebrush (Artemisia tridentata). Great Basin Nat. 59: 380 Ð383. Hölldobler, B., and E. O. Wilson. 1990. The Ants. The Belknap Press of Harvard University Press, Cambridge, MA. Mahdi, T., and J. B. Whittaker. 1993. Do birch trees (Betula pendula) grow better if foraged by wood ants? J. Anim. Ecol 62: 101Ð116. Majer, J. D. 1982. Ant-plant interactions in the Darling Botanical District of Western Australia, pp. 42Ð 63. In R. C. Buckley [ed.], Ant-plant interactions in Australia. Junk, The Hague. Rabkin, F. B., and R. R. Lejeune. 1954. Some aspects of the biology and dispersal of the pine tortoise scale, Toumeyella numismaticum (Pettit and McDaniel) (Homoptera: Coccidae). Can. Entomol. 86: 570 Ð575. Rosenheim, J. A. 1998. Higher-order predators and the regulation of insect herbivore populations. Annu. Rev. Entomol. 43: 421Ð 447. SAS Institute. 1995. JMP statistics and graphics guide, version 3.1. SAS Institute, Cary, NC. Skinner, G. J., and J. B. Whittaker. 1981. An experimental investigation of inter-relationships between the wood- Vol. 94, no. 1